Introduction
Oral cancer is a class of malignant tumors that
usually occurs in the oral cavity, and includes tongue, gingival,
oropharyngeal and lip cancer. Squamous cell carcinoma is the most
common histological type of oral cancer (1–4). Tongue
squamous cell carcinoma (TSCC) accounts for 25–40% of all types of
oral cancer. Due to its high risk of forming secondary or recurrent
tumors in the surrounding areas, the 5-year survival rate of TSCC
was 42% in young patients between 2001 and 2011 in Asia (5–7).
Although advanced diagnostic strategies and therapies, including
surgery, chemotherapy and radiotherapy, are widely used, the
possibility of complete recovery is remote (8,9).
Therefore, a complete recognition of the molecular mechanisms
involved in the multi-step processes of TSCC carcinogenesis and
progression could provide more effective therapeutic targets for
TSCC treatment (10,11).
Long non-coding RNAs (lncRNAs) are defined as a
series of non-coding RNAs >200 nucleotides long (12). Numerous studies have demonstrated
that lncRNAs act as biological controllers to regulate a spectrum
of cellular activities, including cell proliferation, migration,
invasion and apoptosis (13,14). The biological functions and molecular
mechanisms of lncRNAs are complex and varied. lncRNAs can act as
competing endogenous RNAs (ceRNAs) to inhibit the function of
microRNAs (miRNAs/miRs), are involved in chromatin remodeling and
histone protein modification, and bind proteins to regulate their
function (15–17). Additionally, lncRNA dysregulation has
been suggested to be closely associated with several human
diseases, including several types of cancer (18). For example, lncRNA
metastasis-associated lung adenocarcinoma transcript 1 has been
reported to facilitate cancer development by regulating the
alternative splicing process of ‘metastatic signature’ genes, such
as ZEB1 and ZEB2 (19).
Additionally, lncRNA nuclear paraspeckle assembly transcript 1 is
highly expressed in hepatocellular carcinoma tissues and closely
associated with poor prognosis (20). Downregulation of some
cancer-inhibiting lncRNAs and upregulation of some cancer-promoting
lncRNAs have also been reported to be important factors in
accelerating cancer occurrence and development (4). However, identification of these
dysregulated lncRNAs and their role in cancer development is an
ongoing process.
Notably, long intergenic non-coding RNA 00152
(LINC00152) has been reported to be increased in numerous types of
cancer, including glioblastoma, ovarian cancer, urothelial bladder
carcinoma and breast cancer (21–24).
Additionally, a previous study revealed that LINC00152 expression
is significantly upregulated in TSCC tissues, which is
significantly associated with poor prognosis in patients, thereby
suggesting the potential importance of LINC00152 in TSCC
carcinogenesis and progression (25). Dysregulated LINC00152 expression in
TSCC tissues also suggests that it is a potential therapeutic
target. However, the biological functions and the underlying
molecular mechanisms of LINC00152 in TSCC development remain
unclear.
The present study focused on the expression and
roles of LINC00152 in TSCC tissues and cell lines, in order to
elucidate its function in the cancer biology of TSCC. LINC00152
expression was detected and analyzed in TSCC tissues and its roles
in cancer progression were determined in TSCC cell lines.
Additionally, the underlying molecular mechanisms of LINC00152 in
promoting TSCC were analyzed in mechanistic studies. The present
study suggested a potential oncogenic role of LINC00152 in TSCC
development.
Materials and methods
Tissue samples
A total of 15 TSCC tissues and paired adjacent
non-tumor tongue tissues (1 cm away from TSCC tissues) were
obtained from patients who underwent curative surgery at the
Shanghai Changzheng Hospital between July 2014 and October 2016.
The cohort consisted of 10 male and 5 female patients aged between
42–67, with median age 51. All specimens were diagnosed with TSCC
according to histopathological evaluation. None of the patients
underwent chemotherapy or radiotherapy prior to surgery. All
tissues were stored at −80°C. All patients provided written
informed consent for the use of their tissues. The present study
was approved by the Ethics Committee of Xinhua Hospital, Shanghai
Jiao Tong University School of Medicine.
Cell culture
TSCC cell lines, SCC-9 and CAL-27, were purchased
from the Shanghai Institute of Biochemistry and Cell Biology,
Chinese Academy of Sciences. SCC-9 cells were cultured in RPMI-1640
medium (HyClone; GE Healthcare Life Sciences) and 10% FBS (Gibco;
Thermo Fishes Scientific, Inc.), whereas CAL-27 cells were grown in
DMEM (HyClone; GE Healthcare Life Sciences) and 10% FBS. Cells were
maintained in a humidified atmosphere containing 5% CO2
at 37°C.
Subcellular fractionation
The nuclear and cytoplasmic fractions of CAL-27 and
SCC-9 TSCC cells cultured under standard conditions (105
cells at 50% confluence) were separated using the RNA Subcellular
Isolation kit (Active Motif, Inc.), according to the manufacturer's
protocol.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from TSCC tissues and cells
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). For lncRNA and mRNA, 300 ng total RNA was
reverse transcribed to cDNA via random primers using the
PrimeScript RT Reagent kit (Takara Biotechnology Co., Ltd.)
according to the manufacturer's protocols. qPCR analyses were
conducted with specific forward and reverse primers using
SYBR® Premix Ex Taq™ (Takara Biotechnology Co., Ltd.).
The thermocycling conditions were: 95°C for 5 sec; followed by 45
cycles of 55°C for 30 sec and 72°C for 30 sec as described
previously (26–28). Human GAPDH was used as an internal
control. For miRNAs, 300 ng total RNA was reverse transcribed to
cDNA via specific reverse primers using the PrimeScript RT Reagent
kit. qPCR analyses were performed using a unified reverse primer
and specific forward primers using SYBR® Premix Ex Taq™,
according to the manufacturer's protocols. U6 small nuclear (sn)RNA
was used as an internal control. All assays were performed in
triplicate. Relative RNA expression calculated using the
2−ΔΔCq method (28). The
primer sequences were as follows: LINC00152, forward,
5′-GAAAATCACGACTCAGCCCC-3′, reverse 5′-AGACCAGCCCATGACCAAAA-3′;
GAPDH, forward 5′-GCACCGTCAAGGCTGAGAAC-3′, reverse
5′-GGATCTCGCTCCTGGAAGATG-3′; miR-193a-3p, forward
5′-ACACTCCAGCTGGGAACTGGCCTACAAAGT-3′, reverse
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGACTGGGAC-3′; miR-193b-3p,
forward 5′-ACACTCCAGCTGGGAACTGGCCCTCAAAGT-3′, reverse
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGAGCGGGAC-3′; miR-376c-3p,
forward 5′-ACACTCCAGCTGGGAACATAGAGGAAATT-3′, reverse
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGACGTGGAA-3′; unified reverse
primer, 5′-TGGTGTCGTGGAGTCG-3′; and U6 snRNA, forward
5′-CTCGCTTCGGCAGCACA-3′ and reverse 5′-AACGCTTCACGAATTTGCGT-3′.
Plasmid construction and
transfection
The cDNA, reverse transcribed from total RNA,
encoding LINC00152 was PCR-amplified and subcloned into the
pcDNA3.1 vector (Invitrogen; Thermo Fisher Scientific, Inc.). Small
interfering RNAs (siRNAs) specifically targeting LINC00152 and the
negative control (NC) were constructed by Shanghai GenePharma Co.,
Ltd, and the sequences were: si-linc00152-1 sense,
5′-GAAUAACUGGGAGAUGAAATT-3′ and antisense,
5′-UUUCAUCUCCCAGUUAUUCTT-3′; si-linc00152-2 sense,
5′-GGUGGUCUGCCUGUGAUAUTT-3′ and antisense,
5′-AUAUCACAGGCAGACCACCTT-3′; si-linc00152-3 sense,
5′-GUCUUAAUCCCUUGUCCUUTT-3′ and antisense
5′-AAGGACAAGGGAUUAAGACTT-3′. hsa-miR-193b-3p and NC mimics were
also purchased from Shanghai GenePharma Co., Ltd. In addition, a
mutation at the supposed miR-193b-3p response element of LINC00152
was introduced using a QuikChange Site-Directed Mutagenesis kit
(Stratagene; Agilent Technologies, Inc.) according to the
manufacturer's protocol as follows: The target sequence
5′-GGCCAGT-3′ was mutated to 5′-GTCAGTC-3′. A total of 1 µg plasmid
was used for transfections using a jetPEI kit
(Polyplus-transfection SA), according to the manufacturer's
protocol. siRNA or miRNA mimics and NCs were transfected into cells
using the INTERFERin® kit (Polyplus-transfection SA) at
a concentration of 50 nM. Cells were harvested or collected for
other assays 48–72 h post-transfection.
Luciferase assay
A total of 1×104 293T cells per-well were
plated in a 96-well plate. pGL3-promoter-LINC00152-wild type (WT)
or pGL3-promoter-LINC00152-mutant (MUT) (empty vector obtained from
Promega Corporation) were transfected into 293T cells under the
aforementioned culture conditions along with miR-193b-3p mimics or
NC using a jetPEI kit. After 48 h, Renilla luciferase
activity was used as an internal control to normalize relative
firefly luciferase activity. A dual-luciferase reporter gene assay
system (Promega Corporation) was used in this experiment.
Cell proliferation assays
Cell proliferation was detected using a Cell
Counting kit-8 (CCK-8; Dojindo Molecular Technologies, Inc.) assay.
A total of 1×104 SCC-9 and CAL-27 cells transfected with
siRNA (si-)LINC00152/NC or pcDNA3.1-LINC00152/pcDNA3.1 were seeded
into 96-well plates. Cell viability was assessed at 24, 48 and 72 h
post-transfection, according to the manufacturer's protocol.
Absorbance was detected at 450 nm.
Flow cytometric analysis
SCC-9 and CAL-27 cells transfected with
si-LINC00152/si-NC or pcDNA3.1-LINC00152/pcDNA3.1 were collected 48
h post-transfection. A total of 5×104 cells were
analyzed via flow cytometry (FACScan; BD Biosciences) after
staining with the Apoptosis Detection kit (BD Biosciences),
according to the manufacturer's protocol. The percentage of
apoptosis was calculated based on the number of Annexin-V positive
cells. Cell cycle distribution was also analyzed using flow
cytometry after staining with propidium iodide using the Cycletest™
Plus DNA Reagent kit (BD Biosciences), according to the
manufacturer's protocol. The number of cells in
G0/G1, S or G2/M phase were
counted using FlowJo software (version 7.6; FlowJo LLC).
Cell migration and invasion
assays
Cell migration assays were conducted using a 24-well
Transwell chamber (pore size, 8 µm; Corning, Inc.) 24 h post
transfection. Cells (~2×105) were suspended in 200 µl
serum-free medium and seeded into the upper chamber per well. For
the invasion assay, the Transwell chamber was precoated with
Matrigel solution (BD Biosciences), and ~4×105 cells
were seeded into the upper chamber. Next, 500 µl medium containing
10% FBS was added to the lower chamber. After incubation for 48 h
ate 37°C, the cells that remained in the upper chamber were removed
and those below the membrane were fixed using formalin for 5 min
and stained with 0.1% crystal violet for 5 min both at room
temperature. Images of stained cells were captured using a light
microscope at ×40 magnification in five randomly chosen fields, as
described previously (29).
Bioinformatics analysis
Bioinformatics analysis was conducted using starBase
version 3.0 (starbase.sysu.edu.cn). Interactions between LINC00152
and miRNAs were predicted.
Western blot analysis and
antibodies
Total protein was isolated using RIPA lysis buffer
supplemented with protease inhibitors (Beyotime Institute of
Biotechnology). Protein concentration was detected using a
bicinchoninic acid kit (Beyotime Institute of Biotechnology),
according to the manufacturer's protocol. Total protein (20 µg) was
separated by 10% SDS-PAGE and electrophoretically transferred to a
PVDF membrane (EMD Millipore). After blocking in PBS containing
0.1% Tween-20 (Beyotime Institute of Biotechnology) and 5% non-fat
dry milk for 2 h at room temperature, the membranes were incubated
overnight at 4°C with primary antibodies against phosphorylated
(p-)p85 (cat. no. 4228), p85 (cat. no. 4257), p-AKT (cat. no.
4060), AKT (cat. no. 4685), cleaved caspase 3 (cat. no. 9661) and
GAPDH (cat. no. 5174) (all at a 1:1,000 dilution; Cell Signaling
Technology, Inc.). The membranes were then incubated with DyLight™
800-Labeled Antibody to Mouse/Rabbit IgG (H+L) (1:1,000 dilution;
cat. nos. 0412 and 0416, respectively; KPL, Inc.) for 2 h at room
temperature. Finally, immunoblots were visualized using ECL
solution (Pierce; Thermo Fisher Scientific, Inc.), and images were
captured using the FluorChem Imaging system (AlphaView software
version 1.0.2; ProteinSimple; Bio-techne).
Statistical analysis
Statistical analyses were conducted using GraphPad
Prism software (version 6.0; GraphPad Software, Inc.).
Paired-sample or independent-sample t-tests were performed to
examine differences. A one-way ANOVA was used to analyze the
differences among more than two groups. All data are presented as
the mean ± standard deviation of at least three independent
repeats. P<0.05 was considered to indicate a statistically
significant difference.
Results
LINC00152 expression is significantly
upregulated in TSCC tissues
Since the present study primarily focused on the
roles of LINC00152 in TSCC development, LINC00152 expression was
examined in clinical human TSCC tissues. LINC00152 expression in 15
TSCC tissues and paired adjacent non-tumor tissues was detected
using RT-qPCR. As shown in Fig. 1A,
LINC00152 expression levels were significantly higher in TSCC
tissues compared with in the paired adjacent non-tumor tissues
(P<0.01). These findings suggested that LINC00152 expression was
significantly upregulated in TSCC.
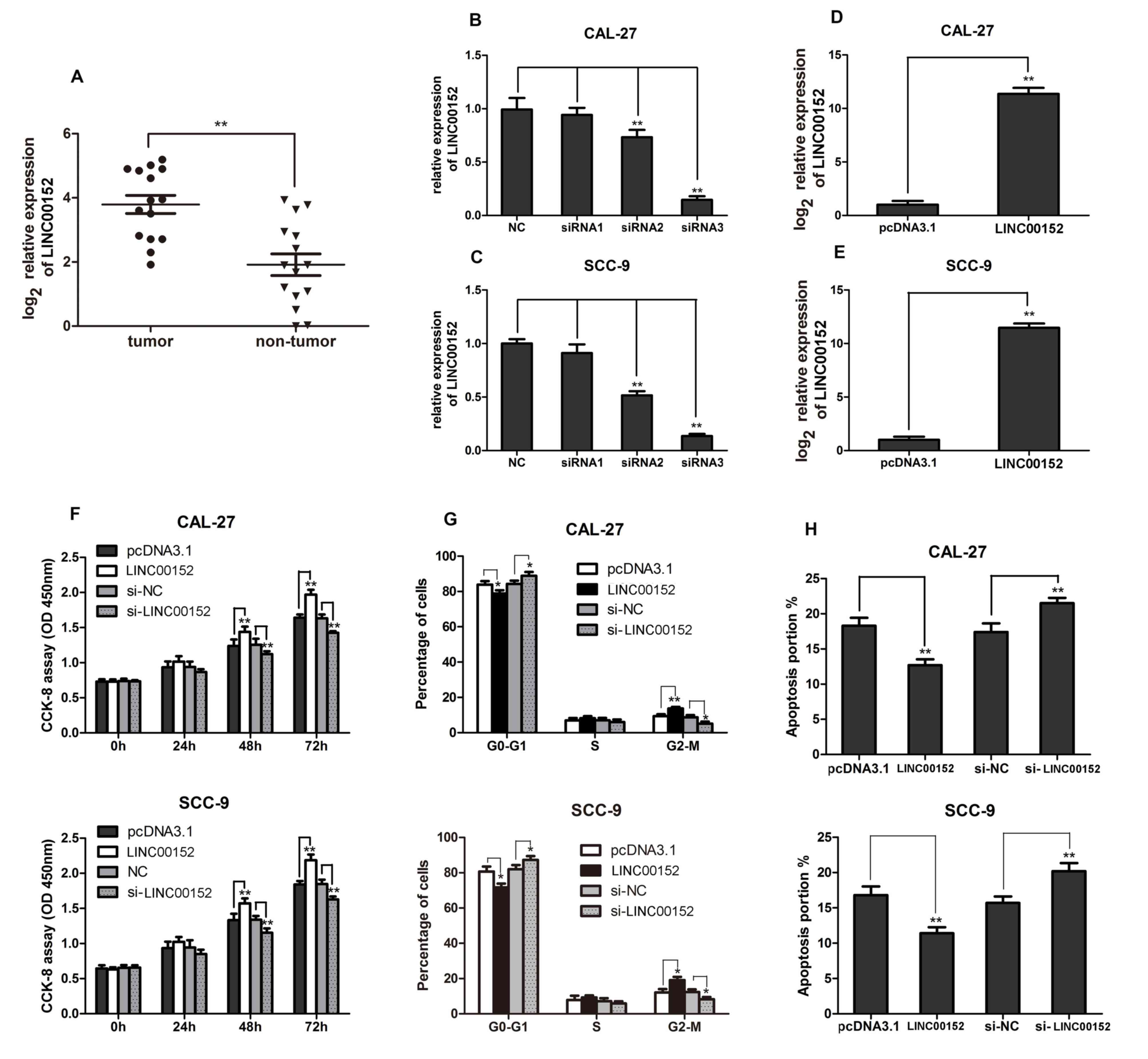 | Figure 1.Increased LINC00152 levels in TSCC
promote cell proliferation, cell cycle progression and apoptosis
inhibition. (A) RT-qPCR analysis was performed to detect LINC00152
expression in 15 TSCC and paired non-tumor tissues. Relative
LINC00152 expression in (B) CAL-27 and (C) SCC-9 cell lines
detected using RT-qPCR 48 h post-transfection with the indicated
siRNAs (n=3). Relative LINC00152 expression in (D) CAL-27 and (E)
SCC-9 cells detected using RT-qPCR 48 h post-transfection with
pcDNA3.1-LINC00152/pcDNA3.1 (n=3). (F) CCK-8 assays were performed
to measure cell growth 0, 24, 48 and 72 h post-transfection with
si-LINC00152/si-NC or pcDNA3.1-LINC00152/pcDNA3.1 in CAL-27 and
SCC-9 cells as indicated (n=6). (G) Flow cytometry was performed to
analyze cell cycle progression in CAL-27 and SCC-9 cells (n=3). (H)
Annexin V-FITC/propidium iodide staining was performed to measure
cell apoptosis in CAL-27 and SCC-9 cells as indicated (n=3). Data
are presented as the mean ± standard deviation. *P<0.05;
**P<0.01. CCK-8, Cell Counting kit-8; LINC00152, long intergenic
non-coding RNA 00152; NC, negative control; OD, optical density;
RT-qPCR, reverse transcription-quantitative PCR; si, small
interfering RNA; TSCC, tongue squamous cell carcinoma. |
LINC00152 promotes TSCC cell
proliferation and cell cycle progression, and inhibits
apoptosis
In order to investigate the biological function of
LINC00152 in TSCC pathogenesis, the TSCC cell lines, CAL-27 and
SCC-9, were transfected with si-LINC00152 or si-NC, and
pcDNA3.1-LINC00152 or empty pcDNA3.1 vector. LINC00152 expression
was then confirmed via RT-qPCR. As shown in Fig. 1B and C, LINC00152 expression was
significantly decreased in siRNA-3-transfected samples compared
with in the NC group. Furthermore, LINC00152 expression in cells
transfected with pcDNA3.1-LINC00152 was significantly increased
compared with the empty pcDNA3.1-transfected cells (Fig. 1D and E). Using a CCK-8 assay to
analyze cell proliferation, it was shown that LINC00152
overexpression significantly promoted the proliferation of the TSCC
cell lines, 48 and 72 h post transfection; however, their
proliferation was significantly inhibited by LINC00152 knockdown
(Fig. 1F). Cell cycle analysis
revealed a reduction in the number of cells in
G0/G1 phase and an increase in cell numbers
in G2/M phase following LINC00152 overexpression,
whereas LINC00152 knockdown increased the number of cells in
G0/G1 phase and decreased the number of cells
in G2/M phase (Fig. 1G).
Furthermore, the percentage of apoptotic cells was decreased by
LINC00152 overexpression and increased by LINC00152 knockdown in
the TSCC cell lines, CAL-27 and SCC-9 (Fig. 1H). Overall, increased LINC00152
expression promoted cell proliferation and cell cycle progression,
and inhibited apoptosis in TSCC cells.
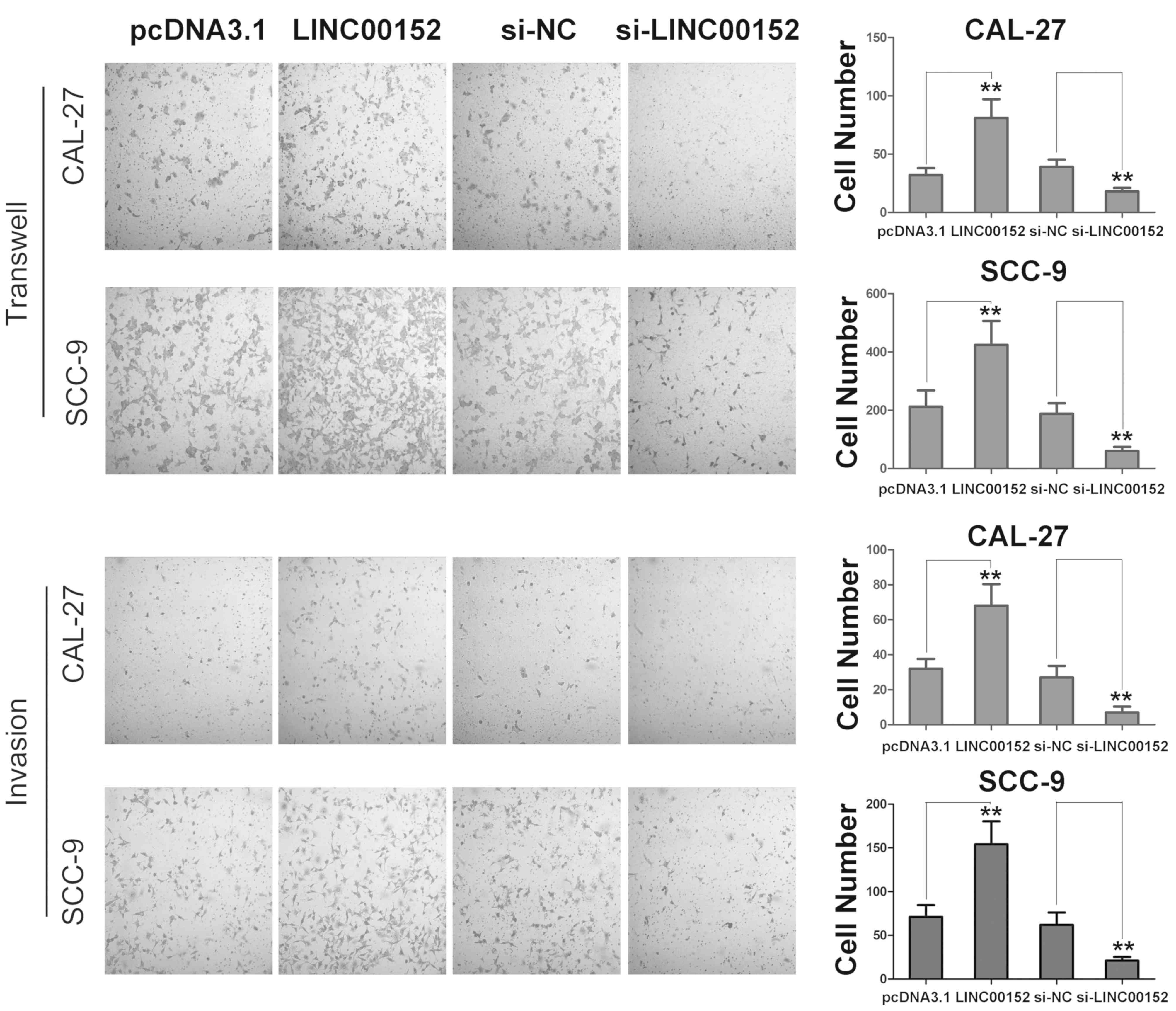
LINC00152 promotes migration and
invasion of TSCC cells
Subsequently, the present study examined whether
LINC00152 regulated migration and invasion of TSCC cells. As shown
in Fig. 2, LINC00152 overexpression
significantly promoted the migration and invasion of the TSCC cell
lines, SCC-9 and CAL-27, compared with control-transfected cells.
Correspondingly, LINC00152 knockdown inhibited the migration and
invasion of TSCC cells. Therefore, these results revealed that
LINC00152 promoted the migration and invasion of TSCC cells and
that increased LINC00152 expression promoted TSCC progression in
vitro.
LINC00152 acts as a miR-193b-3p sponge
to activate the PI3K/AKT signaling pathway in TSCC
The present study further investigated the
mechanisms underlying LINC00152-induced TSCC progression. As shown
in Fig. 3A and B, the subcellular
localization of LINC00152 was investigated. This confirmed that
LINC00152 was mainly found in the cytoplasm of TSCC cells, which
suggested its possible interaction with cytoplasmic miRNAs to exert
its biological functions. Next, the potential interactions between
miRNAs and LINC00152 were analyzed using starBase. Potential
interactions between LINC00152 and miR-193a-3p, miR-193b-3p and
miR-376c-3p were predicted (Fig.
3C). To validate the interaction between LINC00152 and these
miRNAs, relative expression levels of miR-193a-3p, miR-193b-3p and
miR-376c-3p were examined in TSCC CAL-27 and SCC-9 cells. As shown
in Fig. 3D and E, miR-193b-3p
exhibited the highest expression levels in TSCC cells, and the
expression level was similar to that of LINC00152. Notably, miRNA
absorption (sponge activity) is one of the most important
biological functions of lncRNAs and, more importantly, they absorb
miRNAs without inhibiting their expression levels (12). Additionally, miRNAs inhibit the
expression of their target genes when the expression of a
particular miRNA is relatively high (27). Based on this, it may be hypothesized
that LINC00152 exerted its function by sponging miR-193b-3p.
Subsequently, a luciferase reporter plasmid containing the
LINC00152 sequence was constructed and it was revealed that
luciferase activity was significantly inhibited in cells
transfected with miR-193b-3p mimics; whereas, the luciferase
reporter containing the mutated target site was not suppressed by
miR-193b-3p mimics, thus demonstrating the interaction between
LINC00152 and miR-193b-3p (Fig. 3F and
G). Overall, it is possible that LINC00152 acts as a
miR-193b-3p sponge in TSCC cells.
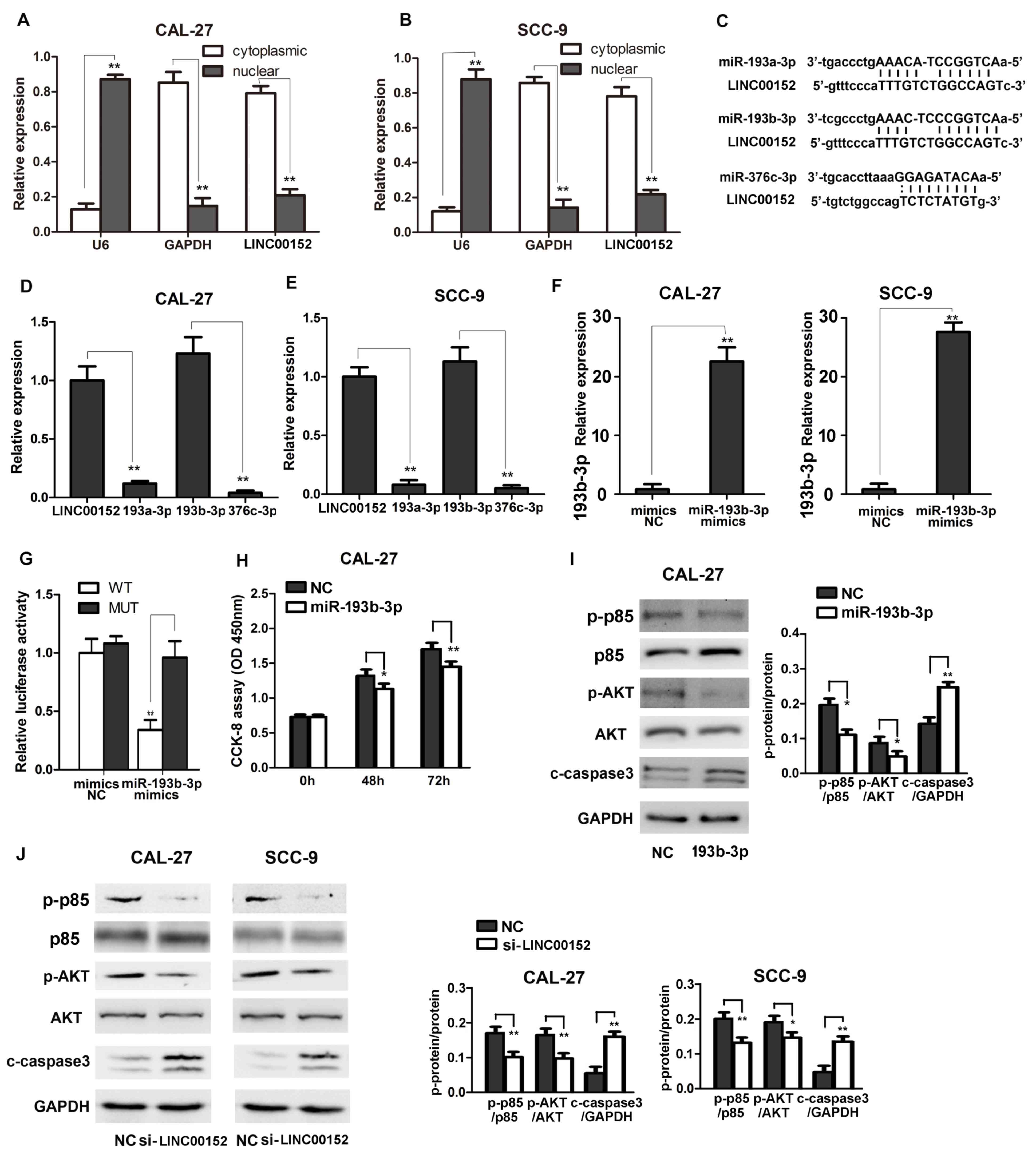 | Figure 3.LINC00152 acts as a miR-193b-3p
sponge to activate the PI3K/AKT signaling pathway in TSCC cells.
Nuclear and cytoplasmic RNAs were separated and relative LINC00152
expression was detected via RT-qPCR in (A) CAL-27 and (B) SCC-9
cells (n=3). (C) Potential LINC00152 miRNAs capable of interaction
were predicted using bioinformatics analysis. Relative miR-193a-3p,
miR-193b-3p, miR-376c-3p and LINC00152 expression in (D) CAL-27 and
(E) SCC-9 cells was detected using RT-qPCR (n=3). (F) Relative
miR-193b-3p expression transfected with miR-193b-3p or NC mimics in
CAL-27 and SCC-9 cells was detected using RT-qPCR. (G) Relative
luciferase activity in 293T cells transfected with LINC00152 WT or
LINC00152 MUT and miR-193b-3p or NC mimics was analyzed as
indicated using a dual luciferase activity assay (n=3). (H) CCK-8
assays were performed to measure cell growth at the indicated time
points after transfection with miR-193b-3p/NC in CAL-27 cells
(n=6). (I and J) p-p85 and AKT and c-caspase 3 were detected using
western blotting as indicated. Data are presented as the mean ±
standard deviation. *P<0.05; **P<0.01. c-, cleaved; CCK-8,
Cell Counting kit 8; LINC00152, long intergenic non-coding RNA
00152; miR, microRNA; MUT, mutant type; NC, negative control; OD,
optical density; p-, phosphorylated; RT-qPCR, reverse
transcription-quantitative PCR; si, small interfering RNA; TSCC,
tongue squamous cell carcinoma; WT, wild-type. |
Since miR-193b-3p has previously been suggested to
inhibit cancer progression by activating the PI3K/AKT signaling
pathway (30–32) and was confirmed to inhibit TSCC
progression. The effect of miR-193b-3p was determined in the TSCC
cell. CAL-27 cells transfected with miR-193b-3p exhibited reduced
proliferation and PI3K/AKT activation (Fig. 3H and I). Additionally, transfection
of miR-139b-3p decreased p85 and AKT phosphorylation (Fig. 3I), LINC00152-regulated activation of
the PI3K signaling pathway in TSCC cells was further examined. As
shown in Fig. 3J, LINC00152
knockdown significantly inhibited the phosphorylation and
activation of p85 and downstream AKT, and activated cleaved caspase
3, thus suggesting that LINC00152 may enhance activation of the
PI3K/AKT signaling pathway to promote TSCC cancer progression.
Overall, these results revealed that LINC00152 acted as a
miR-193b-3p sponge to enhance activation of the PI3K signaling
pathway and inactivation of caspase 3.
Discussion
Numerous studies have reported that lncRNAs serve a
key role in the development of human cancer (13–20).
LINC00152 has been reported to be upregulated in several human
tumor tissues, including gastric cancer, hepatocellular carcinoma,
colon cancer and gallbladder cancer, and to be associated with poor
prognosis (33–36). Functional studies have revealed that
LINC00152 can promote cancer progression in a number of these
cancer types (33–36). The present study revealed that
increased LINC00152 expression in TSCC promoted cell growth, cell
cycle progression, and migration and invasion, while reducing cell
apoptosis. Therefore, LINC00152 may have considerable potential as
a therapeutic target in TSCC progression.
LINC00152 expression was determined to be
upregulated in TSCC tissues in the present study. However, the
mechanisms underlying increased LINC00152 expression remain
unknown. Previous studies have suggested that several gene
regulation mechanisms involving cell signal activation,
transcription factors, epigenetic factors and gene sequences are
dysregulated in cancer cells (19,20).
Future studies will continue to focus on the mechanisms that
increase LINC00152 expression in TSCC cells, particularly the
epigenetic mechanisms, thereby attempting to elucidate TSCC
development in more detail.
Growing evidence has also revealed that lncRNAs can
promote cancer development by acting as ceRNAs in order to inhibit
tumor suppressors (33–36). The present study demonstrated that
LINC00152 was mainly located in the cytoplasm and functioned as a
ceRNA in TSCC cells (Fig.
3A–3G). LINC00152 was further
determined to act as a sponge of miR-193b-3p, which has been
reported to be a tumor suppressor (37,38).
Currently, the possibility that LINC00152 may also absorb other
miRNAs or interact with proteins in TSCC cells to promote cancer
progression cannot be excluded. Future studies will focus on this
issue as well, since LINC00152 has been reported to regulate gene
expression via other mechanisms, including epigenetic modulation
and post-transcriptional regulation (39,40).
Additionally, LINC00152 was demonstrated to promote TSCC migration
and invasion in the present study; however, the roles of
miR-193b-3p in the migration and invasion of TSCC cells remain
elusive. Hence, the mechanisms underlying LINC00152-mediated
migration and invasion need to be further investigated.
Furthermore, a previous study reported that miR-193b could enhance
tumor proliferation and invasion by targeting neurofibromin 1 in
head and neck squamous cell carcinoma (41), which is not in line with the tumor
suppressor roles of miR-193b-3p (39,40).
This suggests that one miRNA may exert different functions in
different types of cancer, and the distinct roles of LINC00152 and
miR-193b-3p in various cancer types may form the basis of
interesting future investigations elucidating their detailed roles
in cancer biology.
In conclusion, the present study demonstrated that
the expression levels of the lncRNA, LINC00152, were significantly
increased in TSCC tissues. Additionally, increased LINC00152
expression promoted cell growth, cell cycle progression, cell
invasion and migration, and inhibited apoptosis, thus enhancing
TSCC progression. Furthermore, mechanistic experiments revealed
that LINC00152 acted as a miR-193b-3p sponge to promote the
phosphorylation and activation of the PI3K signaling pathway and
downstream AKT to contribute to TSCC development.
Acknowledgements
The authors would like to thank Ms. Wei Huang
(Second Military Medical University, Shanghai, China) for her
assistance with the techniques.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81970938) and the
Shanghai Natural Science Foundation (grant no. 15ZR1413000).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HL designed and supervised the study. XL, BR, YC and
XG performed the experiments. HL and XL analyzed the data and wrote
the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Xinhua Hospital, Shanghai Jiao Tong University School
of Medicine (Shanghai, China). All patients provided informed
consent for the use of their tissues.
Patient consent for publication
All patients provided written informed consent for
the use of their tissues and publication of the research data.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
D'souza S and Addepalli V: Preventive
measures in oral cancer: An overview. Biomed Pharmacother.
107:72–80. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yang X, Wu H and Ling T: Suppressive
effect of microRNA-126 on oral squamous cell carcinoma in
vitro. Mol Med Rep. 10:125–130. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
O'Callaghan K, Palagano E, Butini S,
Campiani G, Williams DC, Zisterer DM and O'Sullivan J: Induction of
apoptosis in oral squamous carcinoma cells by
pyrrolo-1,5-benzoxazepines. Mol Med Rep. 12:3748–3754. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tang H, Wu Z, Zhang J and Su B: Salivary
lncRNA as a potential marker for oral squamous cell carcinoma
diagnosis. Mol Med Rep. 7:761–766. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ye X, Wang X, Lu R, Zhang J, Chen X and
Zhou G: CD47 as a potential prognostic marker for oral leukoplakia
and oral squamous cell carcinoma. Oncol Lett. 15:9075–9080.
2018.PubMed/NCBI
|
|
6
|
Jeon JH, Kim MG, Park JY, Lee JH, Kim MJ,
Myoung H and Choi SW: Analysis of the outcome of young age tongue
squamous cell carcinoma. Maxillofac Plast Reconstr Surg. 39:412017.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fu X, Chen S, Chen W, Yang Z, Song M, Li
H, Zhang H, Yao F, Su X, Liu T and Yang AK: Clinical analysis of
second primary gingival squamous cell carcinoma after radiotherapy.
Oral Oncol. 84:20–24. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bachar G, Hod R, Goldstein DP, Irish JC,
Gullane PJ, Brown D, Gilbert RW, Hadar T, Feinmesser R and Shpitzer
T: Outcome of oral tongue squamous cell carcinoma in patients with
and without known risk factors. Oral Oncol. 47:45–50. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Vered M, Dayan D, Dobriyan A, Yahalom R,
Shalmon B, Barshack I, Bedrin L, Talmi YP and Taicher S: Oral
tongue squamous cell carcinoma: Recurrent disease is associated
with histopathologic risk score and young age. J Cancer Res Clin
Oncol. 136:1039–1048. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Farquhar DR, Tanner AM, Masood MM, Patel
SR, Hackman TG, Olshan AF, Mazul AL and Zevallos JP: Oral tongue
carcinoma among young patients: An analysis of risk factors and
survival. Oral Oncol. 84:7–11. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sgaramella N, Gu X, Boldrup L, Coates PJ,
Fahraeus R, Califano L, Tartaro G, Colella G, Spaak LN, Strom A, et
al: Searching for new Targets and Treatments in the battle against
squamous cell carcinoma of the head and neck, with specific focus
on tumours of the tongue. Curr Top Med Chem. 18:214–218. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li X, Wu Z, Fu X and Han W: lncRNAs:
Insights into their function and mechanics in underlying disorders.
Mutat Res Rev Mutat Res. 762:1–21. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Huarte M: The emerging role of lncRNAs in
cancer. Nat Med. 21:1253–1261. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sanchez Calle A, Kawamura Y, Yamamoto Y,
Takeshita F and Ochiya T: Emerging roles of long non-coding RNA in
cancer. Cancer Sci. 109:2093–2100. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yu X and Li Z: Long non-coding RNA HOTAIR:
A novel oncogene (Review). Mol Med Rep. 12:5611–5618. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu Y, Zhang R and Ying K: Long non-coding
RNAs: Novel links in respiratory diseases (Review). Mol Med Rep.
11:4025–4031. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Alvarez-Dominguez JR, Hu W, Gromatzky AA
and Lodish HF: Long noncoding RNAs during normal and malignant
hematopoiesis. Int J Hematol. 99:531–541. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bhan A and Mandal SS: Long noncoding RNAs:
Emerging stars in gene regulation, epigenetics and human disease.
ChemMedChem. 9:1932–1956. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu J, Peng WX, Mo YY and Luo D:
MALAT1-mediated tumorigenesis. Front Biosci (Landmark Ed).
22:66–80. 2017. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yu X, Li Z, Zheng H, Chan MT and Wu WK:
NEAT1: A novel cancer-related long non-coding RNA. Cell Prolif.
502017.doi: 10.1111/cpr.12329.
|
|
21
|
Hu XL, Wang J, He W, Zhao P and Wu WQ:
Down-regulation of lncRNA Linc00152 suppressed cell viability,
invasion, migration, and epithelial to mesenchymal transition, and
reversed chemo-resistance in breast cancer cells. Eur Rev Med
Pharmacol Sci. 22:3074–3084. 2018.PubMed/NCBI
|
|
22
|
Chen X, Li D, Gao Y, Tang W, Iw L, Cao Y
and Hao B: Long intergenic noncoding RNA 00152 promotes glioma cell
proliferation and invasion by interacting with miR-16. Cell Physiol
Biochem. 46:1055–1064. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang J, Yin M, Huang J, Lv Z, Liang S,
Miao X, Huang F and Zhao Y: Long noncoding RNA LINC00152 as a novel
predictor of lymph node metastasis and survival in human cancer: A
systematic review and meta-analysis. Clin Chim Acta. 483:25–32.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Haque SU, Niu L, Kuhnell D, Hendershot J,
Biesiada J, Niu W, Hagan MC, Kelsey KT, Casper KA, Wise-Draper TM,
et al: Differential expression and prognostic value of long
non-coding RNA in HPV-negative head and neck squamous cell
carcinoma. Head Neck. 40:1555–1564. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yu J, Liu Y, Guo C, Zhang S, Gong Z, Tang
Y, Yang L, He Y, Lian Y, Li X, et al: Upregulated long non-coding
RNA LINC00152 expression is associated with progression and poor
prognosis of tongue squamous cell carcinoma. J Cancer. 8:523–530.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hou J, Zhou Y, Zheng Y, Fan J, Zhou W, Ng
IO, Sun H, Qin L, Qiu S, Lee JM, et al: Hepatic RIG-I predicts
survival and interferon-α therapeutic response in hepatocellular
carcinoma. Cancer Cell. 25:49–63. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hou J, Lin L, Zhou W, Wang Z, Ding G, Dong
Q, Qin L, Wu X, Zheng Y, Yang Y, et al: Identification of miRNomes
in human liver and hepatocellular carcinoma reveals miR-199a/b-3p
as therapeutic target for hepatocellular carcinoma. Cancer Cell.
19:232–243. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Han Y, Liu Q, Hou J, Gu Y, Zhang Y, Chen
Z, Fan J, Zhou W, Qiu S, Zhang Y, et al: Tumor-induced generation
of splenic erythroblast-like Ter-cells promotes tumor progression.
Cell. 173:634–648.e12. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang F, Lu J, Peng X, Wang J, Liu X, Chen
X, Jiang Y, Li X and Zhang B: Integrated analysis of microRNA
regulatory network in nasopharyngeal carcinoma with deep
sequencing. J Exp Clin Cancer Res. 35:172016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Pan ST, Qin Y, Zhou ZW, He ZX, Zhang X,
Yang T, Yang YX, Wang D, Qiu JX and Zhou SF: Plumbagin induces G2/M
arrest, apoptosis, and autophagy via p38 MAPK- and
PI3K/Akt/mTOR-mediated pathways in human tongue squamous cell
carcinoma cells. Drug Des Devel Ther. 9:1601–1626. 2015.PubMed/NCBI
|
|
32
|
Bhayadia R, Krowiorz K, Haetscher N,
Jammal R, Emmrich S, Obulkasim A, Fiedler J, Schwarzer A, Rouhi A,
Heuser M, et al: Endogenous tumor suppressor microRNA-193b:
Therapeutic and prognostic value in acute myeloid leukemia. J Clin
Oncol. 36:1007–1016. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Martens-Uzunova ES, Böttcher R, Croce CM,
Jenster G, Visakorpi T and Calin GA: Long noncoding RNA in
prostate, bladder, and kidney cancer. Eur Urol. 65:1140–1151. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ji J, Tang J, Deng L, Xie Y, Jiang R, Li G
and Sun B: LINC00152 promotes proliferation in hepatocellular
carcinoma by targeting EpCAM via the mTOR signaling pathway.
Oncotarget. 6:42813–42824. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yue B, Cai D, Liu C, Fang C and Yan D:
Linc00152 functions as a competing endogenous RNA to confer
oxaliplatin resistance and holds prognostic values in colon cancer.
Mol Ther. 24:2064–2077. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Xia T, Liao Q, Jiang X, Shao Y, Xiao B, Xi
Y and Guo J: Long noncoding RNA associated-competing endogenous
RNAs in gastric cancer. Sci Rep. 4:60882014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang J, Qin J and Su Y: miR-193b-3p
possesses anti-tumor activity in ovarian carcinoma cells by
targeting p21-activated kinase 3. Biomed Pharmacother.
96:1275–1282. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Mets E, Van der Meulen J, Van Peer G,
Boice M, Mestdagh P, Van de Walle I, Lammens T, Goossens S, De
Moerloose B, Benoit Y, et al: MicroRNA-193b-3p acts as a tumor
suppressor by targeting the MYB oncogene in T-cell acute
lymphoblastic leukemia. Leukemia. 29:798–806. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chen QN, Chen X, Chen ZY, Nie FQ, Wei CC,
Ma HW, Wan L, Yan S, Ren SN and Wang ZX: Long intergenic non-coding
RNA 00152 promotes lung adenocarcinoma proliferation via
interacting with EZH2 and repressing IL24 expression. Mol Cancer.
16:172017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wang Y, Liu J, Bai H, Dang Y, Lv P and Wu
S: Long intergenic non-coding RNA 00152 promotes renal cell
carcinoma progression by epigenetically suppressing P16 and
negatively regulates miR-205. Am J Cancer Res. 7:312–322.
2017.PubMed/NCBI
|
|
41
|
Lenarduzzi M, Hui AB, Alajez NM, Shi W,
Williams J, Yue S, O'Sullivan B and Liu FF: MicroRNA-193b enhances
tumor progression via down regulation of neurofibromin 1. PLoS One.
8:e537652013. View Article : Google Scholar : PubMed/NCBI
|