Introduction
Gliomas are the most aggressive, clinically
intractable and unfortunately the most common type of primary
tumors of the central nervous system. Following decades of
advancements in the diagnosis and combined modality therapy of
gliomas, the median survival following initial diagnosis of their
most aggressive form, glioblastoma multiforme, WHO grade IV,
remains at ~14.6 months, and the 5-year survival rate at 9.8%
(1,2).
Human four-and-a-half LIM domains protein 1 (FHL1)
is a member of the FHL protein family, and is characterized by its
possession of four and a half LIM domains. The LIM domain was first
identified during the isolation and identification of the
lin1 gene in Caenorhabditis elegans, the isl-1
gene in rats, and the mec-1 gene in C. elegans. It
was named after the first letter of each gene mentioned above
(3,4). The LIM domain has protein-protein
binding interfaces that possess cysteine-rich zinc finger motifs.
Therefore, FHL1 serves an important role in cellular events, such
as skeletal muscle growth, by interacting with transcription
factors using its cysteine-rich zinc finger motifs (5).
Recently, FHL1 has been implicated in cancer. FHL1
is downregulated in several types of human cancer; the greatest
levels of downregulation are observed in widely invasive and
metastatic cases and associated with poor clinical prognosis
(6–15). FHL1 overexpression suppresses cell
growth through cyclin D1 and E and p27 in head and neck squamous
cell carcinoma (16). FHL1 may also
be converted from a tumor suppressor to a cell growth accelerator,
when FHL1 is phosphorylated by cytosolic tyrosine kinase Src
(17). However, its exact role and
molecular mechanism in gliomas is poorly understood.
PI3K/AKT signaling controls diverse cellular
functions in glioma, including proliferation, survival and
migration (18,19). When the activation of the PI3K/AKT
pathway is inhibited, the levels of cell proliferation and cycle
arrest are significantly decreased (20). The PI3K inhibitor combined with
adenovirus-mediated PTEN could further suppress malignant glioma
cell growth in vitro and in vivo (21). These studies indicated that PI3K/AKT
signaling may be a key topic in the study of malignant glioma cell
growth.
The present study explored the exact role of FHL1 in
glioma and the molecular interactions between FHL1 and PI3K/AKT
signaling. The results demonstrated that the expression of FHL1 was
able to inhibit glioma cell growth in vitro and in
vivo by modulating PI3K/AKT signaling through its interaction
with AKT. FHL1 was also negatively associated with histological
grades of glioma, suggesting that decreased FHL1 expression was
associated with poor prognosis. In conclusion, the data from the
present study provide evidence for a novel diagnostic and
prognostic marker and a new target for glioma treatment in the
future.
Materials and methods
Cell lines and animals
The human glioblastoma U251 cell line and a
glioblastoma of unknown origin U87 cell line were purchased from
the Cancer Institute of Fudan University. The U87 cell line was
authenticated by STR profiling. All cells were cultured in
Dulbecco's modified Eagle medium (Sigma-Aldrich; Merck KGaA)
supplemented with 10% heat-inactivated fetal bovine serum (Thermo
Fisher Scientific, Inc.) and 1% penicillin and streptomycin at 37°C
in an incubator supplied with 5% CO2.
Male BALB/cA-nu 6-week-old nude mice were purchased
from the Shanghai Laboratory Animal Center, Chinese Academy of
Sciences, and maintained under specific pathogen-free conditions. A
total of 12 mice were randomly divided into two groups. The U87
cells stably expressing FHL1 or vehicle controls were injected
subcutaneously into one side of the flank of each mouse with
5×105 cells in each group. A total of 7 days after the
injection, the tumor size was monitored every 3 or 4 days using
sliding caliper measurements, and tumor volumes were calculated
according to the formula: Volume=0.51 × length × width2.
The mice in the present study were sacrificed after 24 day by
cervical dislocation under intraperitoneal sodium pentobarbital
anesthesia (60 mg/kg) to minimize discomfort. The tumors were
excised for further experiments. No significant differences in the
body weights of the experimental mice compared with those of mice
prior to tumor cells inoculation were observed. The results
demonstrated that the largest tumor volume observed in mice was
3.5×103 mm3.
The animal studies were approved by the Animal
Experiment Administration Committee of the Fourth Military Medical
University (approval no., 20190211), and in accordance with the
recommendations of Guide for the Care and Use of Laboratory Animals
prepared by the National Academy of Sciences and published by the
National Institutes of Health (22).
Patients and tissue specimens
Frozen and paraffin-embedded glioma tissues (n=114)
were obtained from the Department of Neurosurgery of Xijing
Hospital, Fourth Military Medical University. Tumors were graded
according to current World Health Organization guidelines (23). All patients involved in the study
provided written informed consent for the use of their samples, and
the protocols involvi0ng human samples were approved by the Ethics
Committee of Xijing Hospital, Fourth Military Medical University
(approval no., KY20183175-1). The clinical data of the patients are
summarized in Table SI.
Immunohistochemistry
FHL1 expression was detected using
immunohistochemical staining of the clinical samples, according to
standard protocol. The glioma tissues were fixed within 30 min
following resection using 10% formalin at room temperature for 24
h. Block of tissue was cut into 5 µm-thick sections that were
heated at 67°C for 120 min. Sections were then deparaffinized twice
with xylene for 5 min, rehydrated with gradient ethanol (100% for 5
min twice, 95% for 5 min twice, 90% for 5 min, 85% for 5 min and
80% for 5 min) and washed with pure water for 1 h. Sections were
then pretreated with 3% H2O2 for 15 min at
4°C to block endogenous peroxidase activity. Antigens were
retrieved by pressure cooker treatment for 100 sec in 0.01 mmol/l
citrate buffer (pH 6). The primary antibody against FHL1 (cat. no.
10991-1-AP; Wuhan Sanying Biotechnology) was diluted at 1:150 and
then incubated with tissue sections overnight at 4°C. Normal rabbit
IgG isotype was used as the negative control. The next day, the
tissue sections were incubated with horseradish peroxidase
(HRP)-conjugated secondary antibodies (Wuhan Boster Biological
Technology, Ltd. cat. no. BM3894; 1:500) for 2 h at room
temperature. Samples were visualized using a 3′-diaminobenzidine
kit (OriGene Technologies, Inc.), and hematoxylin was used for
counterstaining. Images were captured using a BX51 fluorescence
microscope with a CCD camera (DP70; Olympus Corporation). Five
randomized sights were selected (at least 1,000 tumor cells per
slide) under a high-power microscope. The percentage of positive
cells was calculated as ‘positive cells’ number/‘all cells’
number.
According to the percentage of FHL1 expression in
total cells, tumors were classified into two categories: 0–20% FHL1
positive staining in tumor cells, negative/low FHL1 expression;
>20% FHL1 positive staining in tumor cells, high FHL1
expression.
Western blot analysis
Western blot analysis was performed according to
standard procedure as previously described (24). Cell or tissue lysates were obtained
from clinical specimens and glioma cell lines by using lysis buffer
(Beyotime Institute of Biotechnology) supplemented with complete
protease inhibitor cocktail (Roche Diagnostics). The bicinchoninic
acid protein assay kit (Thermo Fisher Scientific, Inc.) was used to
measure protein concentration. Proteins (25 µg) were separated by
10% SDS-PAGE and transferred onto polyvinylidene difluoride
membranes. The membranes were blocked with 5% non-fat dried milk in
PBST for 2 h and incubated overnight with primary antibodies at an
appropriate dilution at 4°C. Following washing with PBST buffer,
the membranes were then incubated with HRP-conjugated secondary
antibodies for 1 h at room temperature and detected by an enhanced
chemiluminescence detection system (Multi Science (Lianke) Biotech
Co., Ltd). Primary antibodies including anti-FHL1 (1:1,200),
anti-AKT (1:500), anti-phospho-AKT (p-AKT), which detects the
endogenous levels of AKT with phosphorylation at threonine 308
(1:500; Signalway Antibody LLC), anti-β-actin (1:1,000; Merck KGaA)
and anti-FLAG (1:1,000; Merck KGaA). As secondary antibodies,
HRP-conjugated goat anti-rabbit IgG and goat anti-mouse IgG (Wuhan
Boster Biological Technology, Ltd.) antibodies were used.
Cell transfection
Lentiviral vectors expressing FHL1 were constructed
using lentiviral packaging systems as previously described
(24). Briefly, a polymerase chain
reaction (PCR) was performed by using the PCR kit (Takara
Biotechnology Co., Ltd.) to amplify human FHL1 open read frame with
human cDNA library as the template. The PCR thermocycling
conditions were as follows: Initial denaturation at 95°C for 5 min,
35 cycles of denaturation at 95°C for 30 sec, annealing at 58°C for
30 sec, elongation at 72°C for 45 sec and final extension at 72°C
for 5 min. Human FHL1 fragment was inserted into the multiple
cloning sites of the pENTR-3C plasmid (Thermo Fisher Scientific,
Inc.), and then the pLenti6.3/V5-FHL1 plasmid (3 µg) was
constructed by EcoR I/Sal I restriction digestion and ligation
(Takara Biotechnology Co., Ltd.). Next, the pLenti6.3/V5-FHL1
plasmid (3 µg) was co-transfected with the packaging plasmid psPAX2
(2.25 µg) and envelope plasmid pMD2.G (0.75 µg) Promega
Corporation) into 293T cells (ATCC). Following transfection by
using Lipofectamine LTX™ (Invitrogen; Thermo Fisher Scientific,
Inc.) for 48 h, the supernatant was collected for the infection of
the U87 cells, and then the stable FHL1 overexpression cell line
was constructed through 1,000 µg/ml blasticidin selection. The
sequences of the primers were as follows: FHL1, forward
5′-CATGGCGGAGAAGTTTGACTG-3′ and reverse
5′-CCTATTGATGGTATAGGGCAGAAAG-3′; and β-actin, forward
5′-CTGGGACGACATGGAGAAAA-3′ and reverse 5′-GCCCAATACGACCAAATCC-3′.
β-actin was used as the internal control.
Next, a cell line with stable FHL1 knockdown was
established. Small interfering RNA (siRNA) targeting the sequence
of FHL1 and the control were inserted into the pLKO.1 vector
(Addgene, Inc.), and the infection was performed following the
recommended protocols (Shanghai GeneChem) by using Lipofectamine
LTX™ (Invitrogen; Thermo Fisher Scientific, Inc.). Cells were
seeded in 6-well plates at the density of 1×106 cells
per well. Transfection was performed when cells have reached ~80%
confluence the next day by using Lipofectamine LTX™. Briefly, 2 µg
plasmid and 5 µl Lipofectamine LTX™ were mixed gently and incubated
for 30 min at room temperature before transfection. The mixture was
subsequently added to the cells at 37°C. After 6 h, culture medium
was refreshed and cells were further cultured for 48 h before
subsequent experiments. The sequence of siRNA targeting to FHL1 was
AAGGAGGTGCACTATAAGAAC. The sequence of control siRNA was
UUCUCCGAACGUGUCACGUTT. The cell line then underwent 400 µg/ml
puromycin selection. FHL1 overexpression or knockdown was confirmed
by western blot analysis.
Reverse transcription (RT)-PCR
Total RNA was extracted from the 2 glioma U87 and
U251 cell lines using TRIzol® reagent (Thermo Fisher
Scientific, Inc.) quantified using NanoDrop™ (Thermo Fisher
Scientific, Inc.) and transcribed using a kit from Toyobo Life
Science. RT-PCR and semi-quantitative PCR were performed by using a
PrimerScript RT Reagent Kit (Takara Biotechnology Co., Ltd.) as
previously described (24). β-actin
was used as the internal control. Primer sequences were as follows:
β-actin forward, CTGGGACGACATGGAGAAAA; β-actin reverse,
GCCCAATACGACCAAATCC; FHL1 forward, CTGAAGTGCTTTGACAAGTTC; and FHL1
reverse, GTGCCAGTAGCGATTCTTAT.
Cell proliferation and cell cycle
assay
Cell proliferation was analyzed using an MTT
colorimetric assay (Sigma-Aldrich; Merck KGaA). Briefly, cells were
counted and seeded in 4 96-well plates at a density of
2×103 cells/well. Each day, 1 plate was selected and 20
µl MTT (5 mg/ml) was added to each well, and then the cells were
incubated at 37°C for 4 h. Following this, the medium was removed
and 150 µl dimethyl sulfoxide was added to solubilize the formazan
precipitate. Absorbance was measured at 490 nm using a microplate
reader.
The cell cycle analysis was performed as previously
described (24). Briefly cells were
resuspended in PBS and fixed in 70% ethanol for 2 h. They were then
stained with 20 mg/ml propidium iodide in PBS containing 0.1%
Triton X-100 and 0.2 mg/ml RNase A for 30 min on ice. The cells
were the analyzed by a FACSCalibur flow cytometer (BD
Immunocytometry Systems). The data were analyzed using CellQuest™
Pro software (version 5.1; Becton, Dickinson and Company).
Co-immunoprecipitation (IP)
PCR-amplified human FHL1 fragment was inserted into
the multiple cloning sites of pCMV-FLAG vector and then confirmed
by DNA sequencing. Cells were transfected with pCMV-FLAG-FHL1 or
pCMV-FLAG using Lipofectamine® LTX &Plus Reagent
(Thermo Fisher Scientific, Inc.). Co-IP was performed as previously
described (17), with an anti-FLAG
monoclonal antibody (Merck KGaA; cat. no. F1804; 1:1,000.). Western
blot analysis was performed using anti-FHL1 or anti-AKT antibody as
aforementioned.
Statistical analysis
Statistical analysis was performed using Student's
t-test and one-way analysis of variance with Least Significant
Difference post hoc test, as appropriate. The association between
FHL1 expression and clinicopathological variables was examined by
Chi-square tests. The Kaplan-Meier method was used to calculate
overall patient survival, while the difference in survival time
between the two groups was examined using log-rank test. All
P-values are two-tailed. P<0.05 was considered to indicate a
statistically significant difference. Data were analyzed using SPSS
v.13.0 software (SPSS, Inc.).
Results
FHL1 expression in patients with
glioma is negatively associated with prognosis
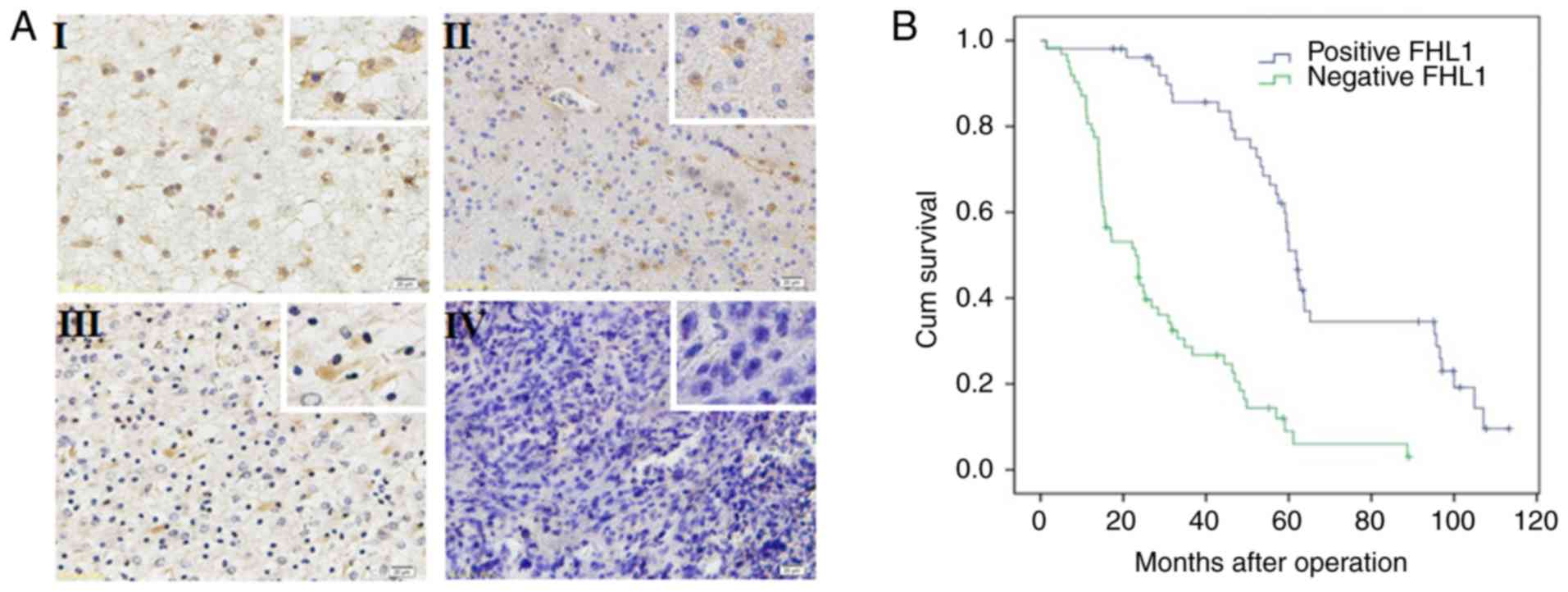
The expression of FHL1 was detected in 114 glioma
specimens by immunohistochemistry. FHL1 was distributed in both the
cytoplasm and the nucleus, but primarily in the cytoplasm. In all
114 glioma specimens, a high FHL1 expression was detected in
low-grade glioma tissues while a low or no expression of FHL1 was
detected in high-grade glioma tissues (Fig. 1A). No association was identified
between FHL1 expression and sex, age, tumor position and extent of
resection. However, FHL1 expression was negatively associated with
histological grades (Table SI). The
association between the survival time in patients with glioma
following surgery and FHL1 expression was further analyzed. The
data indicated that the low FHL1 expression was significantly
associated with poor prognosis. When the expression of FHL1 was
increased, the patients exhibited a longer survival time. These
results suggested that the expression level of FHL1 in glioma was
significantly associated with poor prognosis (Fig. 1B).
FHL1 inhibits glioma cell growth in
vitro
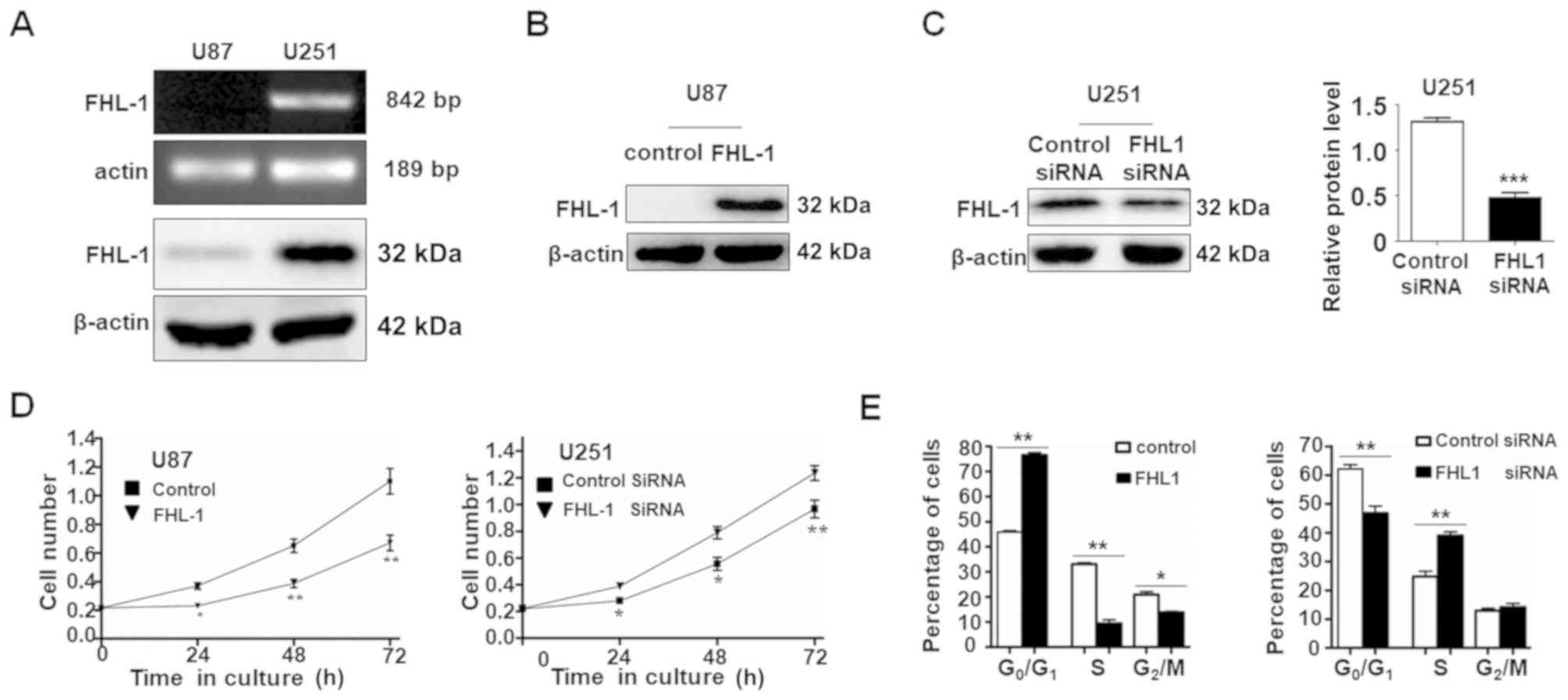
Western blot analysis and RT-PCR were used to
examine the expression level of FHL1 in 2 glioma cell lines (U87
and U251). The mRNA and protein expression levels of FHL1 were not
detected in U87 cells, whereas FHL1 was highly expressed in U251
cells (Fig. 2A). As expected, FHL1
siRNAs specifically suppressed the endogenous expression of FHL1 in
U251 cells and the overexpression of FHL1 also increased the
protein level in U87 cells (Fig.
2B). The effects of FHL1 overexpression or knockdown of
endogenous FHL1 protein on glioma cell growth were also
investigated (Fig. 2C). MTT assay
data indicated that U87 cells transfected with FHL1 expression
vector grew more slowly compared with those transfected with the
empty vector. By contrast, U251 cells transfected with FHL1 siRNAs
grew at an increased rate compared with those transfected with
control siRNA (Fig. 2D). The cell
cycle assays indicated that FHL1 may inhibit cell growth by
maintaining cells in the G0/G1 phase and FHL1 knockdown in U251 may
promote cell proliferation by prompting cells into the S phase
(Fig. 2E). These results suggested
that FHL1 may inhibit glioma cell growth by maintaining cells in
the G0/G1 phase.
FHL1 inhibits glioma growth by
modulating PI3K/AKT signaling by interacting with AKT
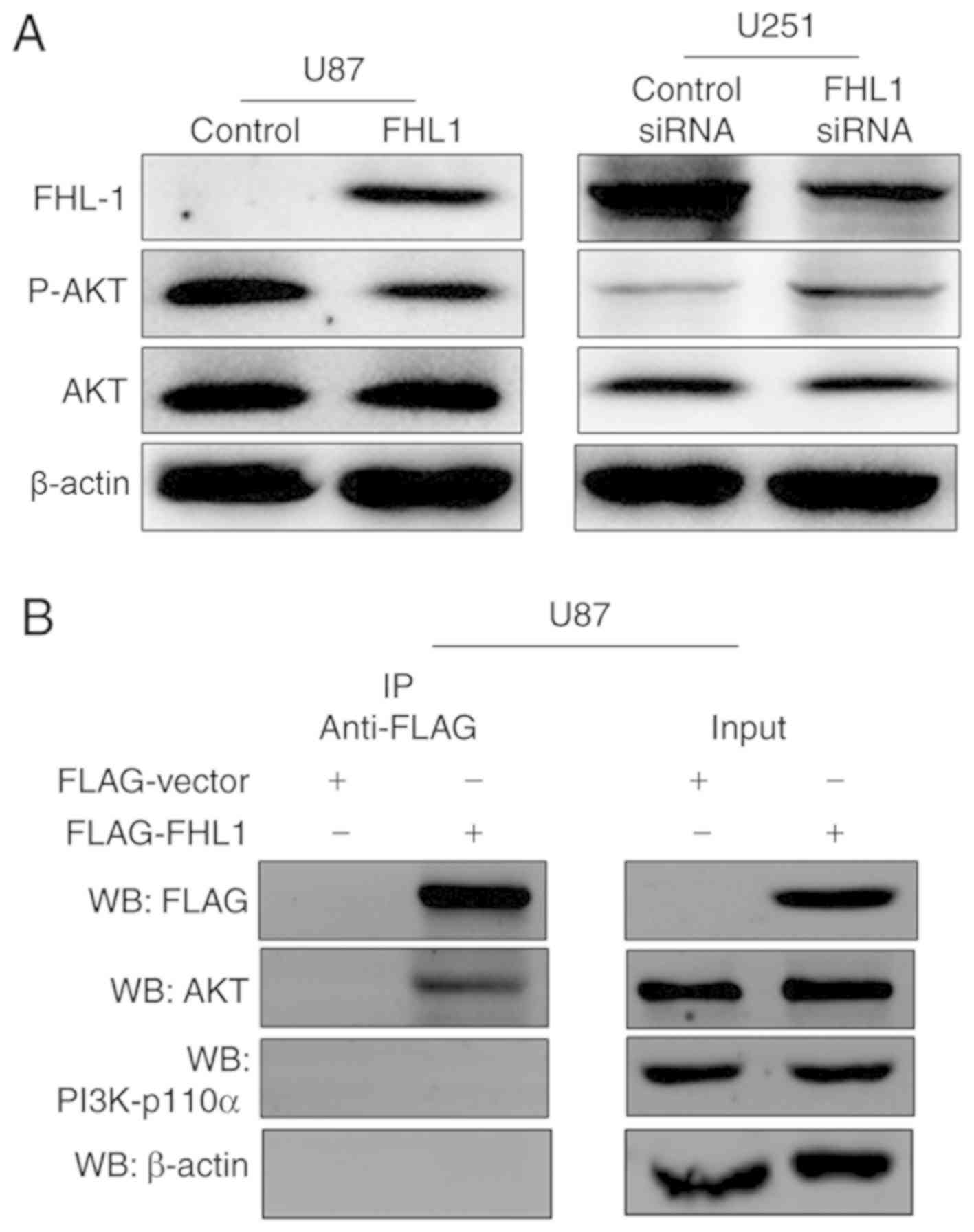
Previous studies have indicated that PI3K/AKT
signaling controls diverse cellular functions in glioma, including
proliferation, suggesting that it may serve a crucial role in
modulating in the studies of glioma cell proliferation (18,19). The
phosphorylation levels of AKT at Thr308 were therefore detected by
western blot analysis, which may significantly increase the
activity of AKT protein. The expression levels of p-AKT were
decreased in the FHL1-overexpressed U87 cell line, as compared with
the control cells (Fig. 3A). By
contrast, the expression levels of p-AKT were increased when FHL1
was knocked down in the U251 cell line, as compared with the
control cells (Fig. 3A). In order to
investigate the association between FHL1 and PI3K/AKT signaling,
Co-IP assays were performed to examine whether FHL1 may modulate
PI3K/AKT signaling by interacting with kinase PI3K-p110α or AKT.
U87 cells were transfected with the FLAG-FHL1 expression or empty
vectors, and Co-IP assays were performed with each antibody. The
results demonstrated that FLAG-FHL1 interacted with AKT,
subsequently preventing the binding of AKT and PI3K and further
phosphorylation, as demonstrated by the lack of interaction between
FLAG-FHL1 and PI3K (p110α). β-actin was considered as a randomized
controlled protein and no detectable interaction was found between
FLAG-FHL1 and β-actin (Fig. 3B).
These data indicated that FHL1 inhibited glioma growth by
modulating PI3K/AKT signaling by interacting with AKT.
FHL1 represses glioma growth in vivo
through PI3K/AKT signaling
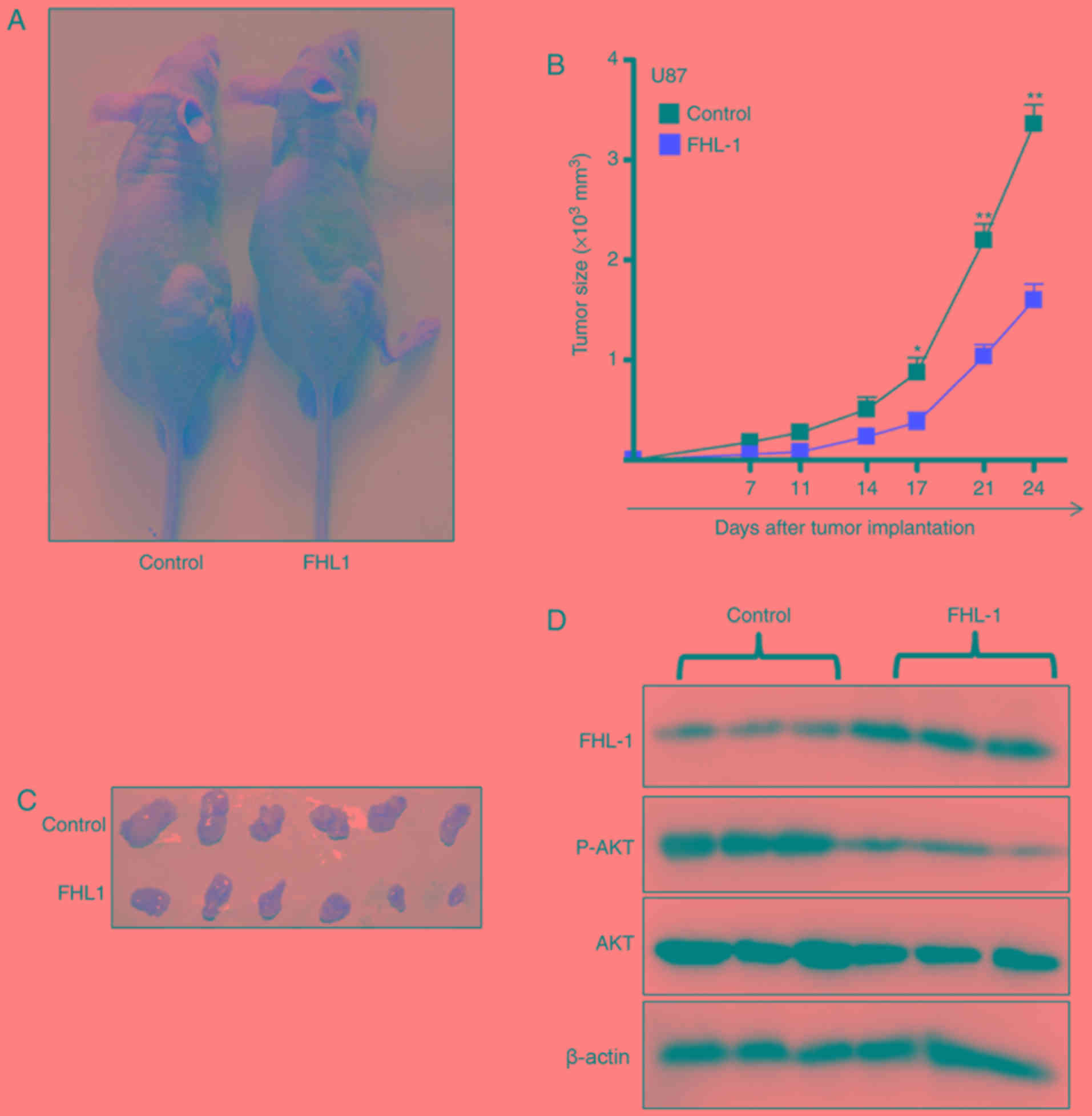
The effects of FHL1 on glioma growth in vivo
were studied by subcutaneously injecting U87-FHL1 overexpression
and U87 control stable cell lines. In the control group, the tumors
gradually grew over time (Fig. 4A),
suggesting that FHL1 overexpression may significantly inhibit tumor
growth. The biggest tumor in the FHL1 overexpression group
exhibited a volume of 499 mm3 (length=15.3 mm;
width=8.00 mm), however the biggest tumor in control group
exhibited a volume of 198 mm3 (length=10.1 mm;
width=6.20 mm). (Fig. 4B and C). In
addition, western blot analysis of the protein extracted from the
subcutaneous transplantation of tumor tissues indicated that FHL1
overexpression was maintained in the transplanted tumors of the
nude mice, and the levels of p-AKT (Thr308) expression decreased in
transplanted tumor tissues formed by the U87-FHL1 overexpression
cells, as compared with the control cells (Fig. 4D). The results indicated that FHL1
was responsible for the inhibition of transplanted tumor growth in
nude mice by modulating PI3K/AKT signaling through the decrease in
phosphorylation at Thr308 of AKT.
Discussion
A growing body of evidence has demonstrated FHL1 to
be significantly downregulated in various types of cancer,
including hepatocellular carcinoma, as well as lung, bladder,
gastric, prostate and breast cancer (6–13). In
gastric and lung cancer, patients with a decreased expression of
FHL1 suffer from significantly shorter survival times compared with
those with increased FHL1 expression levels (7,12).
Furthermore, tumors with a decreased FHL1 expression level are
generally more invasive (11,12,15).
Consistent with the data described for other types of cancer, the
present study demonstrated for the first time, to the best of our
knowledge, that gliomas with a decreased FHL1 expression level are
more malignant compared with those with an increased FHL1
expression level, and that decreased FHL1 expression levels
predicted shorter survival in patients with glioma.
FHL1 is regarded as a tumor suppressor by functions
downstream of Src and Cas to suppress non-anchored tumor growth
(6). In addition, FHL1 has been
demonstrated to inhibit growth of various cancer cells by
physically and functionally interacting with transcription factors.
For example, Ding et al (8)
demonstrated that FHL1 may inhibit hepatocellular carcinoma cell
growth by interacting with Smad proteins, including Smad2-4, which
are mediators of transforming growth factor-β (TGF-β) signaling.
The interaction between FHL1 and Smad complexes in the nucleus has
been demonstrated to regulate TGF-β-responsive target gene
transcription, including the activation of p21 and repression of
the oncogene c-myc (8). Furthermore,
Ding et al (10) also
verified that FHL1 serves an important role in suppressing breast
cancer cell growth by regulating estrogen receptor signaling and
FHL1 interaction with ERs to repress the transcription of
estrogen-responsive genes, trefoil factor 1 (TFF1) and
cathepsin D (10). FHL1 can also
interact with receptor-interacting protein of 140 kDa, a repressor
for ER, to inhibit the transcription of the TFF1 gene in
breast cancer cells (9) and FHL1 may
downregulate estrogen receptor α activity by repressing AKT
phosphorylation in human breast cancer cells. It should be noted
that the human glioblastoma U251 cell line is PTEN-deficient and
exhibits high level of phosphorylated AKT (25). In the present study, we hypothesized
that the interaction between FHL1 with AKT may affect the spatial
structure of AKT and decrease, at least partially, the
phosphorylation efficiency of AKT by PI3K. Therefore, following
disruption of the binding of FHL1 and AKT by FHL1 silencing, normal
PI3K/AKT signaling was restored and activated, as demonstrated by
the increased levels of p-AKT in FHL1-knockdown U251 cells. A
recent study by Niu et al (12) indicated that FHL1 may also inhibit
lung cancer cell growth by markedly inhibiting the expression of
cyclin A, B1 and D and inducing the expression of cyclin-dependent
kinase inhibitors p21and p27, which lead to G1 and the G2/M cell
cycle arrest. Similarly, the present study identified that FHL1 may
inhibit glioma cell proliferation in vitro and impede
tumorigenesis in vivo.
Previous studies have indicated that PI3K/AKT
signaling affects cell growth, proliferation and survival in
numerous types of cancer, including glioma (26). The inactivation of PI3K/AKT signaling
inhibits glioma cell growth (27).
In addition, Cyr61-modulated hepatocyte growth factor-dependent
tumor cell growth is regulated by AKT (28), which results in PI3K/AKT signaling as
the principal target in the studies of glioma cell proliferation.
The present study also demonstrated that FHL1 inhibits
proliferation in vivo and in vitro through its
interaction with AKT, following decreased phosphorylation at Thr308
of AKT. However, only co-immunoprecipitation has been performed to
demonstrate the interaction between FHL1 and AKT. Additional
experiments are required for further functional studies.
The present study indicated that FHL1 may be a tumor
suppressor in glioma. Firstly, FHL1 was downregulated in high-grade
glioma tissues with more malignant biological behaviors, as
compared with low-grade glioma tissues. Secondly, the
downregulation of FHL1 predicted shorter survival in patients with
glioma. Thirdly, the overexpression of FHL1 suppressed cell growth
in human glioma cell lines and endogenous FHL1 knockdown promoted
glioma cell growth. It was also identified that FHL1 may inhibit
glioma cell growth in nude mice. Finally, the mechanism of action
of the inhibitory effect of FHL1 on the proliferation of glioma
cells was demonstrated to function through the regulation of
PI3K/AKT signaling, through direct interaction with AKT. Therefore,
these data suggested that FHL1 may serve an important role in the
development and progression of glioma, and that FHL1 may be a new
diagnostic and prognostic marker and novel target for glioma
treatment in the future.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by The National
Natural Science Foundation of China (grant nos. 81402438, 81502145
and 81071874) and Natural Science Foundation of Shaanxi (grant nos.
2016JQ8017 and 2019SF117).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
SZL performed immunohistochemical staining of the
clinical samples, interpreted the patient data regarding the grade
and prognosis of patients with glioma and performed animal
experiments. YYH performed western blotting and RT-PCR and analysed
the corresponding data. JLZ performed lentiviral packaging, cell
proliferation and cell cycle assays, and co-immunoprecipitation. JZ
performed statistical analysis for clinical data and analyzed the
corresponding data. HYQ was a major contributor in conception and
design of this study. HH analysed and interpreted the whole data of
this study and drafted the manuscript. ZF participated in the whole
process of the present study, including the study design,
experiment guide, data interpretation and manuscript revision. All
of the authors participated sufficiently in the work to take public
responsibility for appropriate portions of the content and agreed
to be accountable for all aspects of the work in ensuring that
questions related to the accuracy or integrity of any part of the
work are appropriately investigated and resolved. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
The animal studies were approved by the Animal
Experiment Administration Committee of the Fourth Military Medical
University (approval no., 20190211), and in accordance with the
recommendations of Guide for the Care and Use of Laboratory Animals
prepared by the National Academy of Sciences and published by the
National Institutes of Health (NIH publication 86–23, revised
1985). All patients involved in the study provided written informed
consent for the use of their samples, and the protocols involving
human samples were approved by the Ethics Committee of Xijing
Hospital, Fourth Military Medical University (approval no.
KY20183175-1.)
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hegi ME, Diserens AC, Gorlia T, Hamou MF,
de Tribolet N, Weller M, Kros JM, Hainfellner JA, Mason W, Mariani
L, et al: MGMT gene silencing and benefit from temozolomide in
glioblastoma. N Engl J Med. 352:997–1003. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Stupp R, Hegi ME, Mason WP, Van den Bent
MJ, Taphoorn MJ, Janzer RC, Ludwin SK, Allgeier A, Fisher B,
Belanger K, et al: Effects of radiotherapy with concomitant and
adjuvant temozolomide versus radiotherapy alone on survival in
glioblastoma in a randomised phase III study: 5-year analysis of
the EORTC-NCIC trial. Lancet Oncol. 10:459–466. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lee SM, Tsui SK, Chan KK, Garcia-Barcelo
M, Waye MM, Fung KP, Liew CC and Lee CY: Chromosomal mapping,
tissue distribution and cDNA sequence of four-and-a-half LIM domain
protein 1 (FHL1). Gene. 216:163–170. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bach I: The LIM domain: Regulation by
association. Mech Dev. 91:5–17. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cowling BS, McGrath MJ, Nguyen MA, Cottle
DL, Kee AJ, Brown S, Schessl J, Zou Y, Joya J, Bönnemann CG, et al:
Identification of FHL1 as a regulator of skeletal muscle mass:
Implications for human myopathy. J Cell Biol. 183:1033–1048. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shen Y, Jia Z, Nagele RG, Ichikawa H and
Goldberg GS: SRC uses Cas to suppress Fhl1 in order to promote
nonanchored growth and migration of tumor cells. Cancer Res.
66:1543–1552. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sakashita K, Mimori K, Tanaka F, Kamohara
Y, Inoue H, Sawada T, Hirakawa K and Mori M: Clinical significance
of loss of Fhl1 expression in human gastric cancer. Ann Surg Oncol.
15:2293–2300. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ding L, Wang Z, Yan J, Yang X, Liu A, Qiu
W, Zhu J, Han J, Zhang H, Lin J, et al: Human four-and-a-half LIM
family members suppress tumor cell growth through a TGF-beta-like
signaling pathway. J Clin Invest. 119:349–361. 2009.PubMed/NCBI
|
|
9
|
Lin J, Ding L, Jin R, Zhang H, Cheng L,
Qin X, Chai J and Ye Q: Four and a half LIM domains 1 (FHL1) and
receptor interacting protein of 140kDa (RIP140) interact and
cooperate in estrogen signaling. Int J Biochem Cell Biol.
41:1613–1618. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ding L, Niu C, Zheng Y, Xiong Z, Liu Y,
Lin J, Sun H, Huang K, Yang W, Li X and Ye Q: FHL1 interacts with
oestrogen receptors and regulates breast cancer cell growth. J Cell
Mol Med. 15:72–85. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Matsumoto M, Kawakami K, Enokida H, Toki
K, Matsuda R, Chiyomaru T, Nishiyama K, Kawahara K, Seki N and
Nakagawa M: CpG hypermethylation of human four-and-a-half LIM
domains 1 contributes to migration and invasion activity of human
bladder cancer. Int J Mol Med. 26:241–247. 2010.PubMed/NCBI
|
|
12
|
Niu C, Liang C, Guo J, Cheng L, Zhang H,
Qin X, Zhang Q, Ding L, Yuan B, Xu X, et al: Downregulation and
growth inhibitory role of FHL1 in lung cancer. Int J Cancer.
130:2549–2556. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang F, Feng F, Yang P, Li Z, You J, Xie
W, Gao X and Yang J: Four-and-a-half-LIM protein 1 down-regulates
estrogen receptor α activity through repression of AKT
phosphorylation in human breast cancer cell. Int J Biochem Cell
Biol. 44:320–326. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xu Y, Liu Z and Guo K: Expression of FHL1
in gastric cancer tissue and its correlation with the invasion and
metastasis of gastric cancer. Mol Cell Biochem. 363:93–99. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li X, Jia Z, Shen Y, Ichikawa H, Jarvik J,
Nagele RG and Goldberg GS: Coordinate suppression of Sdpr and Fhl1
expression in tumors of the breast, kidney, and prostate. Cancer
Sci. 99:1326–1333. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cao W, Liu J, Xia R, Lin L, Wang X, Xiao
M, Zhang C, Li J, Ji T and Chen W: X-linked FHL1 as a novel
therapeutic target for head and neck squamous cell carcinoma.
Oncotarget. 7:14537–14550. 2016.PubMed/NCBI
|
|
17
|
Wang X, Wei X, Yuan Y, Sun Q, Zhan J,
Zhang J, Tang Y, Li F, Ding L, Ye Q and Zhang H: Src-mediated
phosphorylation converts FHL1 from tumor suppressor to tumor
promoter. J Cell Biol. 217:1335–1351. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dey N, Crosswell HE, De P, Parsons R, Peng
Q, Su JD and Durden DL: The protein phosphatase activity of PTEN
regulates SRC family kinases and controls glioma migration. Cancer
Res. 68:1862–1871. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Nakamura JL, Garcia E and Pieper RO: S6K1
plays a key role in glial transformation. Cancer Res. 68:6516–6523.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li Y, Ma X, Wang Y and Li G: miR-489
inhibits proliferation, cell cycle progression and induces
apoptosis of glioma cells via targeting SPIN1-mediated PI3K/AKT
pathway. Biomed Pharmacother. 93:435–443. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nan Y, Guo L, Song Y, Wang L, Yu K, Huang
Q and Zhong Y: Combinatorial therapy with adenoviral-mediated PTEN
and a PI3K inhibitor suppresses malignant glioma cell growth in
vitro and in vivo by regulating the PI3K/AKT signaling pathway. J
Cancer Res Clin Oncol. 143:1477–1487. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bayne K: Revised Guide for the Care and
Use of Laboratory Animals available. American Physiological
Society. Physiologist. 39:199, 208–211. 1996.
|
|
23
|
Louis DN, Ohgaki H, Wiestler OD, Cavenee
WK, Burger PC, Jouvet A, Scheithauer BW and Kleihues P: The 2007
WHO classification of tumours of the central nervous system. Acta
Neuropathol. 114:97–109. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang H, Yan X, Ji LY, Ji XT, Wang P, Guo
SW and Li SZ: miR-139 functions as an antioncomir to repress glioma
progression through targeting IGF-1 R, AMY-1, and PGC-1β. Technol
Cancer Res Treat. 16:497–511. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang YT, Yuan B, Chen HD, Xu L, Tian YN,
Zhang A, He JX and Miao ZH: Acquired resistance of phosphatase and
tensin homolog-deficient cells to poly(ADP-ribose) polymerase
inhibitor and Ara-C mediated by 53BP1 loss and SAMHD1
overexpression. Cancer Sci. 109:821–831. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Vivanco I and Sawyers CL: The
phosphatidylinositol 3-Kinase AKT pathway in human cancer. Nat Rev
Cancer. 2:489–501. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Han L, Yang Y, Yue X, Huang K, Liu X, Pu
P, Jiang H, Yan W, Jiang T and Kang C: Inactivation of PI3K/AKT
signaling inhibits glioma cell growth through modulation of
β-catenin-mediated transcription. Brain Res. 1366:9–17. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Goodwin CR, Lal B, Zhou X, Ho S, Xia S,
Taeger A, Murray J and Laterra J: Cyr61 mediates hepatocyte growth
factor-dependent tumor cell growth, migration, and Akt activation.
Cancer Res. 70:2932–2941. 2010. View Article : Google Scholar : PubMed/NCBI
|