Introduction
In 2018, colorectal cancer (CRC) was reported to be
the third most commonly diagnosed malignancy and the second leading
cause of cancer-associated mortalities worldwide (1). Although many advances in the early
detection and comprehensive therapy of CRC have prolonged the
survival time of patients with CRC, patients with advanced CRC in
the United States have been reported to still experience tumor
progression, relapse and metastasis following initial surgery
(2). Several studies have
demonstrated that complex genomic and epigenomic alterations that
represent the heterogeneous molecular characterization of CRC are
the leading causes of tumorigenesis and progression of CRC
(3–5). In addition to immune response and
influences by exogenous factors, the complexity of molecular
genetic mechanisms remains a challenge to current personalized
therapeutic approaches to CRC (6).
Therefore, it is important to identify genes associated with the
pathogenesis and prognosis of CRC to find candidate molecular
biomarkers and therapeutic targets.
LBX2, located on chromosome 2p13.1, is a
transcription factor gene with barely any previous functional
studies or known animal models (7).
LBX2 serves a role in body pattern formation as well as
morphogenesis of the heart, skeletal muscle and nervous systems in
vertebrates, such as zebrafish and mice (7). In zebrafish, LBX2 has been reported to
regulate gastrulation movements and hypaxial myogenesis through the
canonical or noncanonical Wnt/β-catenin signaling pathway (8,9). Owing
to its diversity in functional domains, previous studies have
revealed that LBX2 is not only involved in diverse physiological
processes, including the regulation of myofibril formation and
development of heart (7,10), but also tumorigenesis. For example,
the aberrant expression of LBX2 has been observed in several types
of cancers, such as non-small cell lung cancer, adenoid cystic
carcinoma and T-cell acute lymphoblastic leukemia (11–13). The
methylation and downregulation of LBX2 are also associated with
adenoid cystic carcinoma development and progression (12). Thus, LBX2 deregulation may contribute
to carcinogenesis; however, the expression levels of LBX2 and its
role in CRC remain unclear.
To investigate the significance of LBX2 expression
in CRC, a comprehensive analysis of the association between the
mRNA expression levels of LBX2 and prognosis in CRC was conducted
by mining data from The Cancer Genome Atlas (TCGA) database
(cancer.gov/tcga). Furthermore, to investigate the
potential underlying molecular mechanisms of LBX2 dysregulation and
corresponding biological pathways in CRC, the regulatory miRNAs of
LBX2 were evaluated and Gene Set Enrichment Analysis (GSEA) was
performed to find potential signaling pathways associated with the
upregulation of LBX2 in CRC. The present study is the first to
describe the deregulation of LBX2 in TCGA colon adenocarcinoma and
rectum adenocarcinoma cohorts of 383 patients with CRC and the
association between LBX2 expression levels and survival.
Materials and methods
Mining data from TCGA database
The gene expression data of mRNA sequencing, miRNA
sequencing and the corresponding clinical datasheets of 51 normal
colorectal issues and 383 primary CRC samples were obtained from
the TCGA website. Both the mRNA sequencing data and clinical
information of all 383 patients with CRC and the miRNA sequencing
data of 324 patients were analyzed. Sequenced data were downloaded
using the Illumina HiSeq_RNA-Seq and Illumina HiSeq_miRNA-Seq
platforms (portal.gdc.cancer.gov). The R software package ‘edgeR’
(version 3.6.0; R-project.org) was used to normalize
and process the downloaded data and identify the differentially
expressed genes between the CRC samples and normal controls.
Log2 conversions were performed for all gene expression
data. The threshold was determined according to the following
values: log2FoldChange >1 and adjusted P-value
<0.05. The gene-expression levels of LBX2 in CRC was reviewed
using the Oncomine database (oncomine.org/resource/login.html) and Tumor Immune
Estimation Resource (TIMER) (cistrome.shinyapps.io/timer/). The
LBX2 DNA copy number alterations (CNA) and methylation data were
obtained from cBioPortal (14).
Prediction of the regulatory miRNAs of
LBX2
The regulatory miRNAs of LBX2 were predicted using
four miRNAs target prediction databases: TargetScan version 7.2
(targetscan.org/vert_72), miRDB
(mirdb.org/), miRWalk (129.206.7.150) and microRNA
(microrna.org/microrna/home.do). The
common miRNAs in at least three of the target prediction databases
were defined as the prediction cohort using Venny version 2.1.0
(bioinfogp.cnb.csic.es/tools/venny/index.html).
Compared with normal controls, the miRNAs downregulated in TCGA
colon adenocarcinoma and rectum adenocarcinoma cohorts (also
present in the prediction cohort) were regarded as the candidate
regulatory miRNAs of LBX2 in CRC. In addition, linear regression
analyses were performed to assess the association between LBX2 mRNA
expression levels and miRNAs expression. Kaplan-Meier analyses were
performed to determine the prognostic value of candidate regulatory
miRNAs of LBX2 in CRC.
Gene set enrichment analysis
(GSEA)
GSEA software version 3.0 was used to evaluate the
correlations between LBX2 expressions (high vs. low) and
tumor-associated pathways in the TCGA colon adenocarcinoma and
rectum adenocarcinoma datasets. The reference gene set C2
(c2.cp.kegg.v6.0.symbols.gmt), which summarizes and represents
specific, well-defined, biological signal pathways and processes,
belongs to the Molecular Signatures Database (software.broadinstitute.org/gsea/msigdb/index.jsp)
(15). The normalized enrichment
score (NES) was obtained by analyzing with permutations for 1,000
times. A gene set was deemed to be significantly enriched when the
normal P-value <0.05 and the false discovery rate (FDR)
<0.25.
Statistical analysis
A two-tailed Student's t-test was used to perform
expression data comparisons in different clinicopathological groups
using the GraphPad Prism 7 software (GraphPad Software). One-way
ANOVA and Tukey's post-hoc test were used for the comparison of
datasets containing three or more groups. Receiver operating
characteristic (ROC) curves were established to estimate the
diagnostic value of LBX2 in CRC. Patients were then divided into
low and high LBX2 expression groups based on survival data using
the X-tile software version 3.6.1 (16), which automatically chooses the
optimum cut-off value. The correlations between LBX2 expression
levels and clinicopathological parameters were evaluated using SPSS
software (version 22.0; IBM Corp.) with the application of the
two-tailed χ2 test. Overall survival (OS) and
disease-free survival (DFS) were determined using Kaplan-Meier
analysis and a log-rank test. SPSS was employed to perform
univariate and multivariable analyses to generate a Cox
proportional hazards model for the analysis of relative risk
factors associated with OS or DFS. The Pearson's correlation
analysis was used to determine the linear relationship between the
LBX2 mRNA expression levels and the LBX2 DNA methylation level or
the miRNAs expression. P<0.05 was considered to indicate a
statistically significant difference.
Results
LBX2 is overexpressed in CRC tissues
compared with normal colorectal tissues
The Oncomine database was used to analyze LBX2 mRNA
expression levels in colorectal tumors and corresponding normal
tissues. As shown in Fig. 1A,
compared with normal tissues, LBX2 expression levels were higher in
CRC. In addition, a lower LBX2 expression was observed in cervical
and gastric cancer.
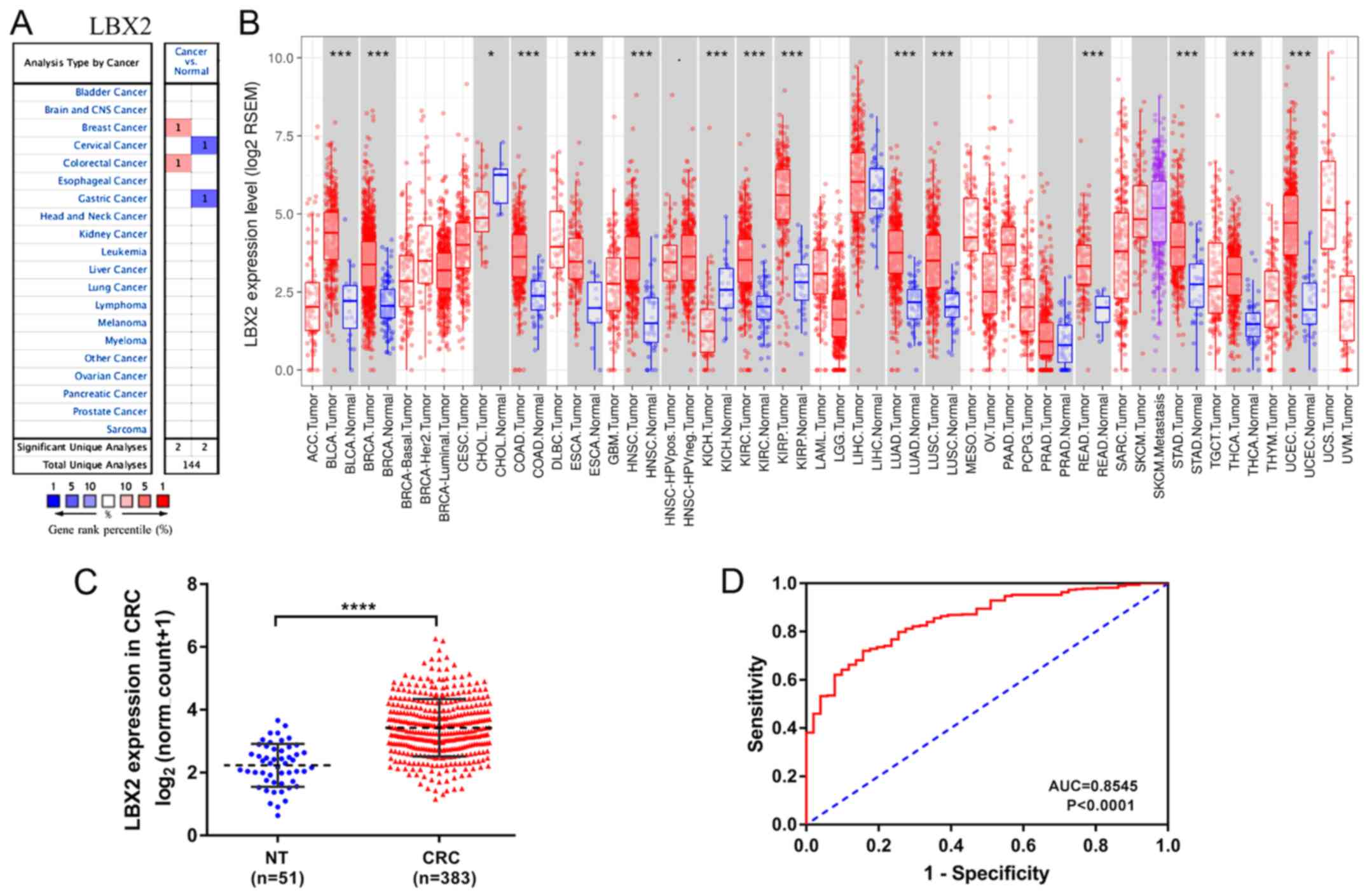 | Figure 1.LBX2 mRNA expression profiles in
different types of cancer. (A) Increased or decreased LBX2 in data
sets of different types of cancer compared with normal tissues in
the Oncomine database. (B) LBX2 expression levels in different
tumor types from the Cancer Genome Atlas database were examined
using the Tumor Immune Estimation Resource. (C) Comparison of LBX2
expression levels in CRC tissues (n=383) and in normal colorectal
tissues (n=51). (D) Receiver Operating Characteristic curves were
used to validate LBX2 overexpression for CRC survival prediction.
*P<0.05, **P<0.01, ***P<0.001, ****P<0.0001. NT, normal
tissue; CRC, colorectal cancer; AUC, area under the curve; ACC,
adrenocortical carcinoma; BLCA, bladder urothelial carcinoma; BRCA,
breast invasive carcinoma; CESC, cervical squamous cell carcinoma
and endocervical adenocarcinoma; CHOL, cholangiocarcinoma; COAD,
colon adenocarcinoma; DLBC, lymphoid neoplasm diffuse large B-cell
lymphoma; ESCA, esophageal carcinoma; GBM, glioblastoma multiforme;
HNSC, head and neck squamous cell carcinoma; KICH, kidney
chromophobe; KIRC, kidney renal clear cell carcinoma; KIRP, kidney
renal papillary cell carcinoma; LAML, acute myeloid leukemia; LGG,
brain lower grade glioma; LIHC, liver hepatocellular carcinoma;
LUAD, lung adenocarcinoma; LUSC, lung squamous cell carcinoma;
MESO, mesothelioma; OV, ovarian serous cystadenocarcinoma; PAAD,
pancreatic adenocarcinoma; PCPG, pheochromocytoma and
paraganglioma; PRAD, prostate adenocarcinoma; READ, rectum
adenocarcinoma; SARC, sarcoma; SKCM, skin cutaneous melanoma; STAD,
stomach adenocarcinoma; TGCT, testicular germ cell tumors; THCA,
thyroid carcinoma; THYM, thymoma; UCEC, uterine corpus endometrial
carcinoma; UCS, uterine carcinosarcoma; UVM, uveal melanoma. |
To assess LBX2 expression in the various types of
cancer further, the LBX2 expression levels in multiple malignant
tumors in TCGA were analyzed using mRNA expression data from TIMER.
The difference in LBX2 expression levels between tumor and adjacent
normal tissues across all TCGA tumors are shown in Fig. 1B. LBX2 expression levels were
significantly higher in bladder urothelial carcinoma, breast
invasive carcinoma, colon adenocarcinoma, esophageal carcinoma,
head and neck cancer, kidney renal clear cell carcinoma, lung
adenocarcinoma, lung squamous cell carcinoma, rectum
adenocarcinoma, stomach adenocarcinoma, thyroid carcinoma and
uterine corpus endometrial carcinoma, compared with normal tissues.
Conversely, LBX2 expression levels were significantly lower in
cholangiocarcinoma and kidney chromophobe compared with adjacent
normal tissues.
Furthermore, the quantitative evaluation of the
expression levels of LBX2 mRNA in CRC samples (n=383) showed that
LBX2 mRNA expression levels were significantly higher compared with
normal colorectal tissues (n=51; P<0.0001; Fig. 1C). The AUC value of LBX2
overexpression for CRC diagnosis was 0.8545 (P<0.0001) (Fig. 1D). These results suggest that LBX2 is
upregulated in multiple types of cancer, particularly in CRC.
Association between LBX2 expression
levels and survival of patients with CRC
Overall, 383 patients with CRC with complete LBX2
mRNA sequencing data and clinical data in TCGA colon adenocarcinoma
and rectum adenocarcinoma datasets were analyzed to explore the
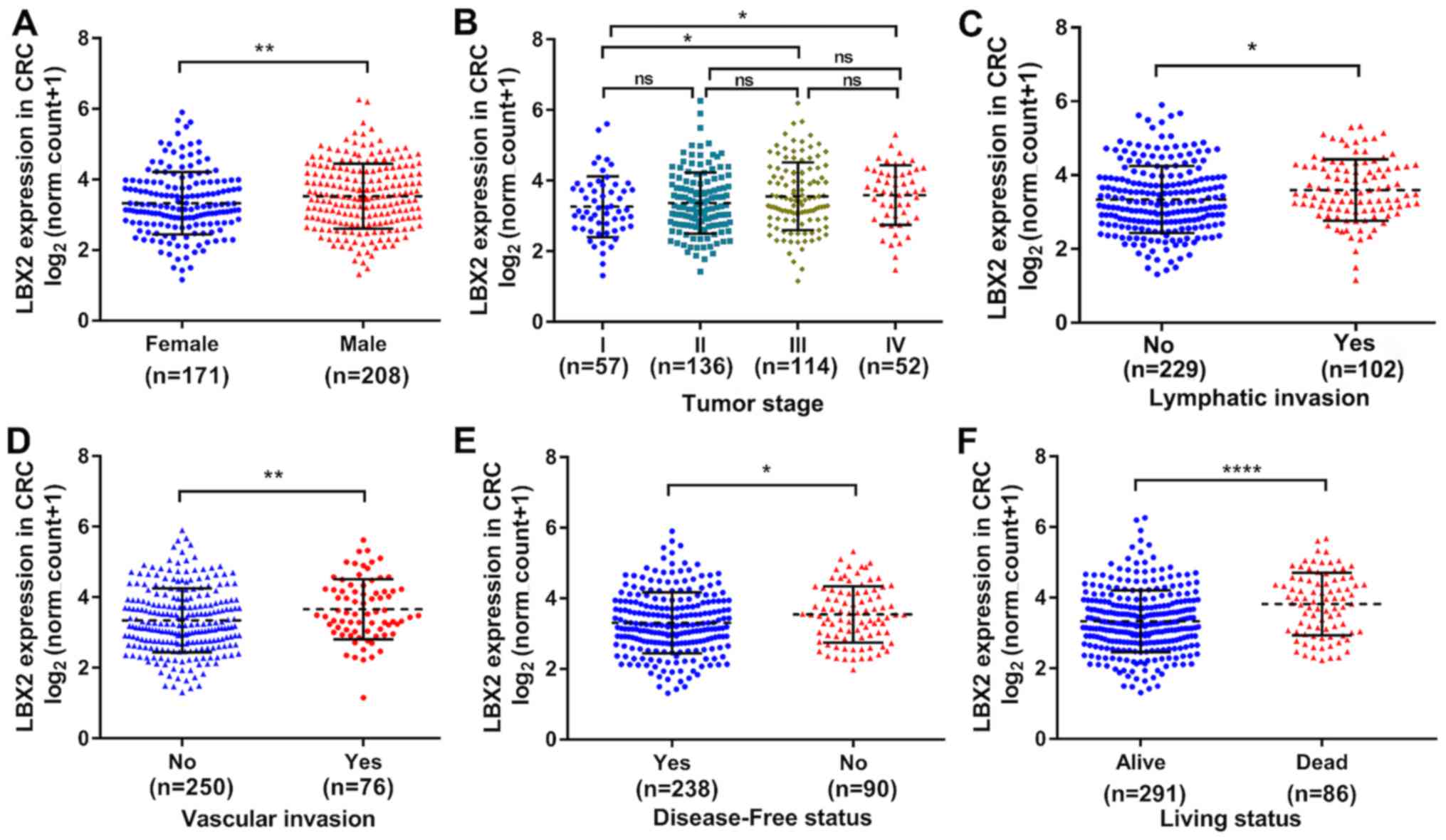
clinical significance of LBX2. As shown in Table I, the expression levels of LBX2 were
not associated with patients' age, perineural infiltration and
residual tumor (all P>0.05), whereas significant correlations
were observed between high LBX2 expression levels and male
patients, advanced tumor stage (17)
(III or IV), vascular invasion, lymphatic invasion, recurrence or
progression and living status (all P<0.05). Consistent with
these findings, LBX2 mRNA expression levels data were analyzed as a
continuous variable, which further demonstrated the different LBX2
expression levels among the different clinicopathological groups
(sex, tumor stage, lymphatic or vascular invasion, disease-free
status and living status) (all P<0.05; Fig. 2).
 | Table I.Association between LBX2 expression
levels and clinical characteristics in patients with colorectal
cancer in The Cancer Genome Atlas. |
Table I.
Association between LBX2 expression
levels and clinical characteristics in patients with colorectal
cancer in The Cancer Genome Atlas.
|
| LBX2
expression |
|
|
|---|
|
|
|
|
|
|---|
| Characteristic | Low, n=191,
(%) | High, n=192,
(%) | χ2
value | P-value |
|---|
| Age, years |
|
| 0.3137 | 0.5754 |
|
<65 | 93 (48.7) | 88 (45.8) |
|
|
|
≥65 | 98 (51.3) | 104 (54.2) |
|
|
| Sex |
|
| 6.841 | 0.0089b |
|
Male | 93 (48.7) | 119 (62.0) |
|
|
|
Female | 98 (51.3) | 73 (38.0) |
|
|
| Tumor stage |
|
| 6.758 | 0.0341a |
|
I–II | 108 (56.6) | 88 (45.9) |
|
|
|
III–IV | 73 (38.2) | 93 (48.4) |
|
|
|
Unknown | 10 (5.2) | 11 (5.7) |
|
|
| Vascular
invasion |
|
| 6.519 | 0.0384a |
| No | 138 (72.3) | 115 (59.9) |
|
|
|
Yes | 31 (16.2) | 45 (23.4) |
|
|
|
Unknown | 22 (11.5) | 32 (16.7) |
|
|
| Lymphatic
invasion |
|
| 8.539 | 0.0140a |
| No | 125 (65.5) | 105 (54.7) |
|
|
|
Yes | 44 (23.0) | 58 (30.2) |
|
|
|
Unknown | 22 (11.5) | 29 (15.1) |
|
|
| Paraneural
infiltration |
|
| 0.7739 | 0.6791 |
| No | 83 (43.5) | 86 (44.8) |
|
|
|
Yes | 32 (16.7) | 26 (13.5) |
|
|
|
Unknown | 76 (39.8) | 80 (41.7) |
|
|
| Residual tumor |
|
| 2.207 | 0.3318 |
| R0 | 121 (63.3) | 126 (65.6) |
|
|
|
R1-R2 | 3 (1.6) | 7 (3.7) |
|
|
|
RX-unknown | 67 (35.1) | 59 (30.7) |
|
|
| Disease-free
status |
|
| 9.209 | 0.0100b |
|
Disease-free | 134 (70.1) | 106 (55.2) |
|
|
|
Recurrence | 37 (19.4) | 54 (28.1) |
|
|
|
Unknown | 20 (10.5) | 32 (16.7) |
|
|
| Living status |
|
| 11.99 | 0.0025b |
|
Alive | 157 (82.2) | 134 (69.8) |
|
|
|
Dead | 30 (15.7) | 57 (29.7) |
|
|
|
Unknown | 4 (2.1) | 1 (0.5) |
|
|
LBX2 overexpression independently
predicts poor OS and DFS in CRC
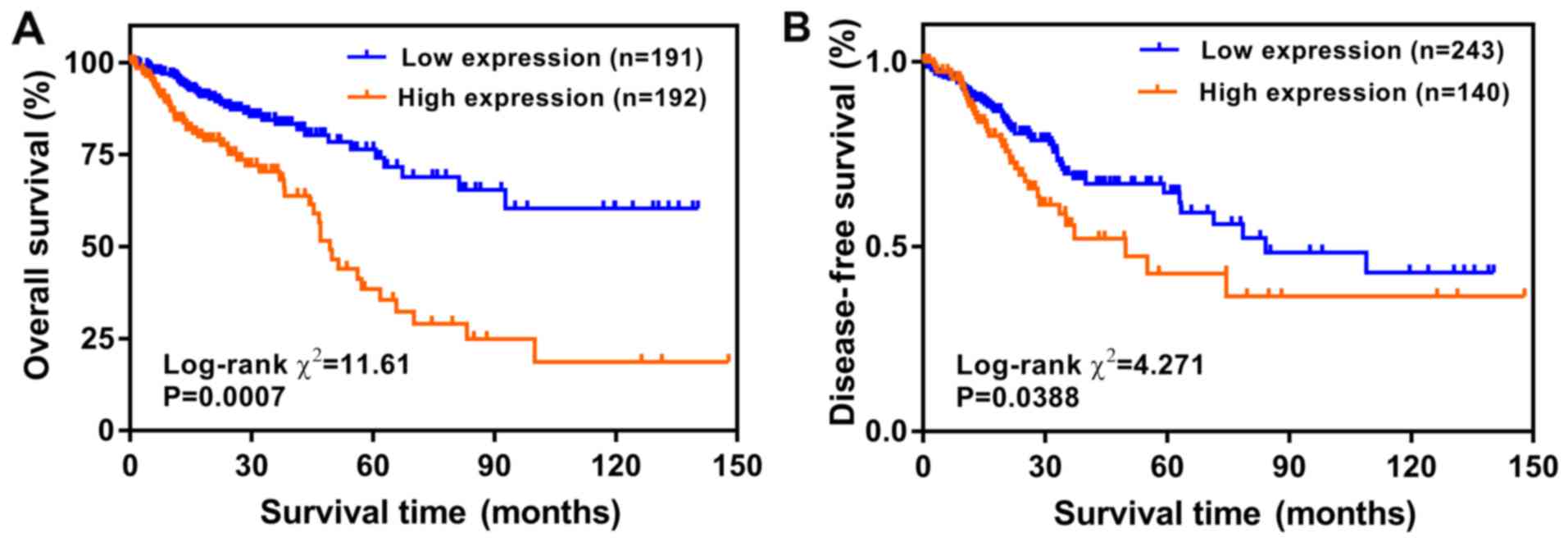
To explore the prognostic value of LBX2 expression
levels, the Kaplan-Meier method was used to analyze the effects of
LBX2 expression on the OS and DFS of patients with CRC. The results
showed that high LBX2 expression levels were significantly
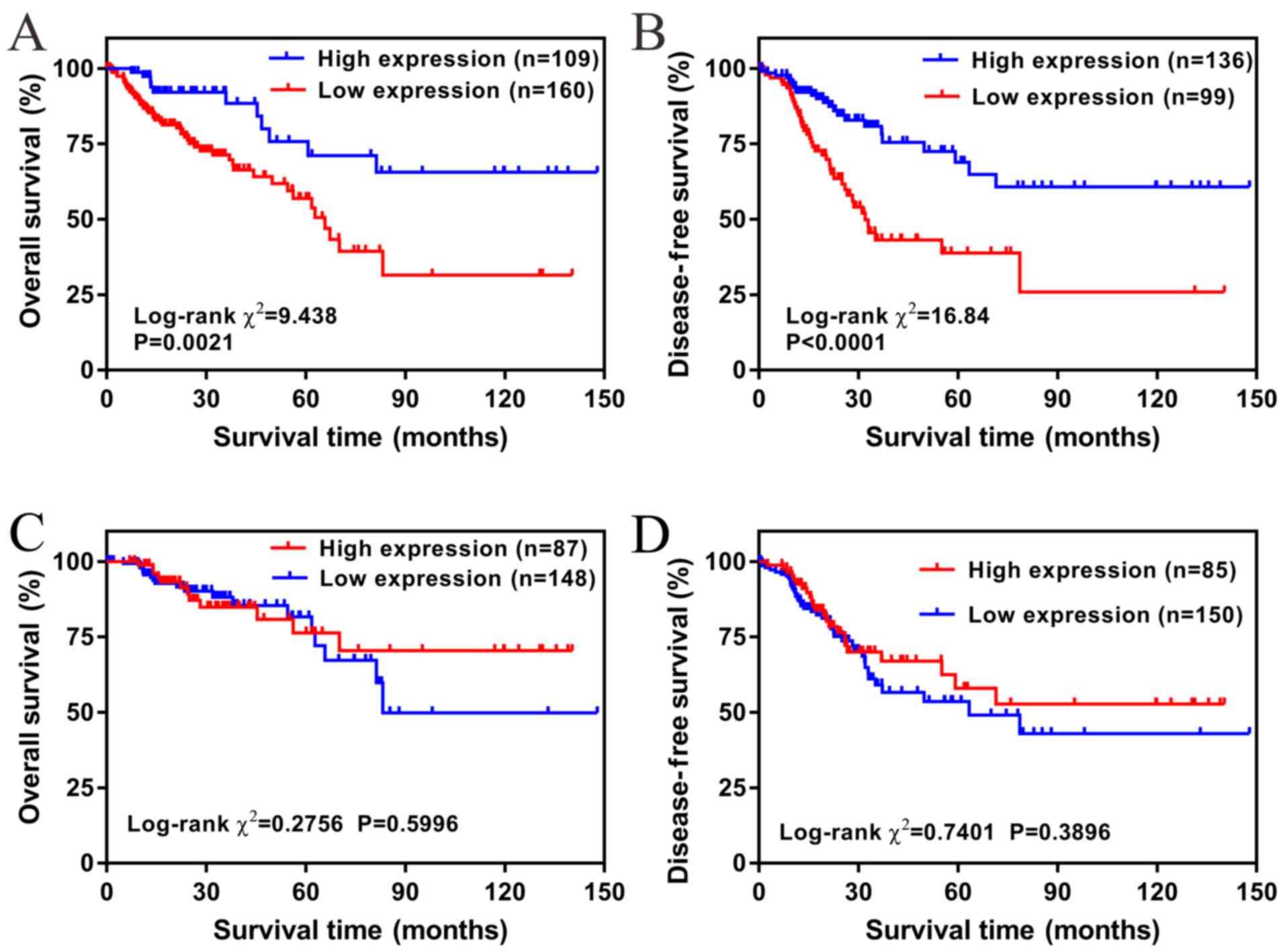
correlated with less favorable OS and DFS (all P<0.05; Fig. 3).
Furthermore, the independent risk factors of OS and
DFS were evaluated using the Cox proportional hazards regression
model. Univariate and multivariate analyses demonstrated that OS
was significantly associated with LBX2 expression levels, age and
tumor stage, and vascular and lymphatic invasion (only univariate
analysis) (all P<0.05; Table
II). Conversely, univariate and multivariate analyses
demonstrated that DFS was significantly associated with LBX2
expression levels and tumor stage, and vascular and lymphatic
invasion and residual tumor (only univariate analysis) (all
P<0.05; Table III).
Multivariate Cox regression analyses further demonstrated that
upregulation of LBX2 was an independent risk factor of a less
favorable OS [high vs. low; hazard ratio (HR), 2.934; confidence
interval (CI), 1.735–4.965; P<0.001] and DFS (high vs. low; HR,
2.135; CI, 1.183–3.853; P=0.012) (Tables II and III).
| Table II.Univariate and multivariate Cox
regression analyses of overall survival for patients with
colorectal cancer. |
Table II.
Univariate and multivariate Cox
regression analyses of overall survival for patients with
colorectal cancer.
|
| Univariate
analysis | Multivariate
analysis |
|---|
|
|
|
|
|---|
| Variables | HR (95% CI) | P-value | HR (95% CI) | P-value |
|---|
| LBX2, high vs.
low | 2.791
(1.816–4.289) |
<0.001b | 2.934
(1.735–4.965) |
<0.001b |
| Age, years, ≥65 vs.
<65 | 1.997
(1.261–3.161) | 0.008a | 2.247
(1.299–3.887) | 0.004a |
| Sex, female vs.
male | 1.219
(0.795–1.870) | 0.363 | − | − |
| Tumor stage, III–IV
vs. I–II | 2.943
(1.829–4.735) |
<0.001b | 3.349
(1.875–5.983) |
<0.001b |
| Vascular invasion,
yes vs. no | 2.446
(1.521–3.934) |
<0.001b | 2.223
(1.137–4.346) | 0.156 |
| Paraneural
infiltration, yes vs. no | 1.552
(0.819–2.938) | 0.178 | − | − |
| Lymphatic invasion,
yes vs. no | 1.703
(1.084–2.675) | 0.008a | 0.653
(0.327–1.304) | 0.325 |
| Residual tumor,
R1-R2 vs. R0 | 1.920
(0.675–5.460) | 0.222 | − | − |
| Table III.Univariate and multivariate Cox
regression analyses of disease-free survival for patients with
colorectal cancer. |
Table III.
Univariate and multivariate Cox
regression analyses of disease-free survival for patients with
colorectal cancer.
|
| Univariate
analysis | Multivariate
analysis |
|---|
|
|
|
|
|---|
| Variables | HR (95% CI) | P-value | HR (95% CI) | P-value |
|---|
| LBX2, high vs.
low | 1.685
(1.112–2.555) | 0.014a | 2.135
(1.183–3.853) | 0.012a |
| Age, years, ≥ 65
vs. < 65 | 0.900
(0.596–1.358) | 0.616 | − | − |
| Sex, female vs.
male | 1.387
(0.906–2.213) | 0.132 | − | − |
| Tumor stage, III–IV
vs. I–II | 2.599
(1.705–3.906) |
<0.001b | 1.952
(1.016–3.750) | 0.045a |
| Vascular invasion,
yes vs. no | 1.762
(1.109–2.801) | 0.017a | 1.187
(0.530–2.658) | 0.677 |
| Paraneural
infiltration, yes vs. no | 1.396
(0.800–2.437) | 0.240 | − | − |
| Lymphatic invasion,
yes vs. no | 2.291
(1.501–3.495) |
<0.001b | 1.630
(0.508–2.664) | 0.721 |
| Residual tumor,
R1-R2 vs. R0 | 2.530
(1.074–5.962) | 0.034a | 2.384
(0.951–5.977) | 0.064 |
DNA hypomethylation and microRNA
(miR)-378a-3p dysregulation contribute to LBX2 upregulation in
CRC
To further explore the potential mechanisms of LBX2
upregulation in CRC, the genetic and epigenetic alterations of LBX2
in TCGA colon adenocarcinoma and rectum adenocarcinoma datasets
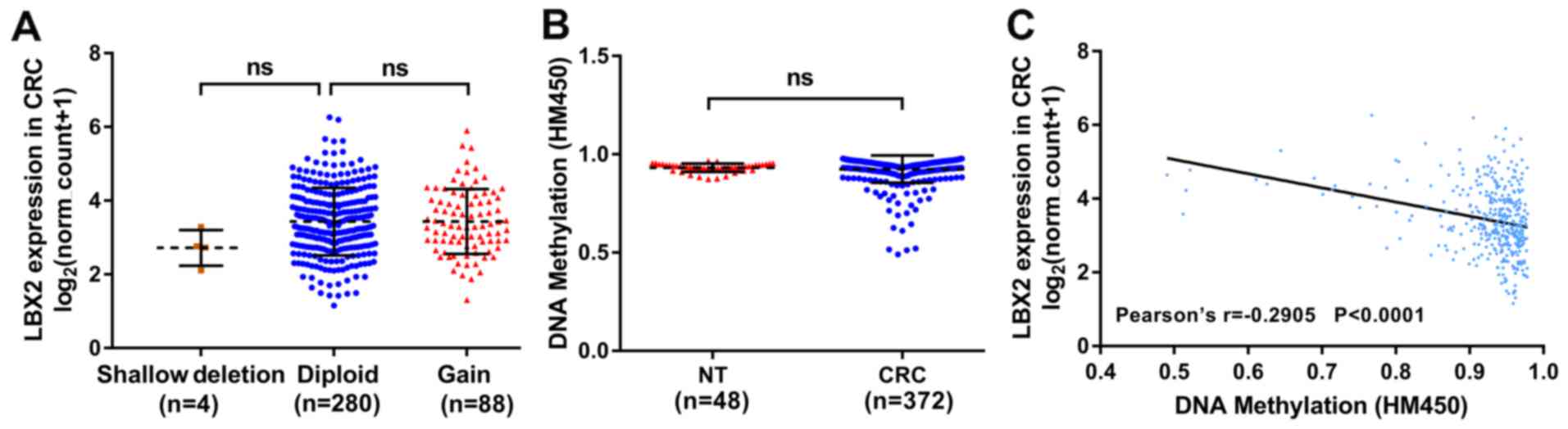
were analyzed. According to the tumor samples with complete mRNA,
CNA and methylation data (n=372), 4 patients with CRC harbored a
LBX2 shallow deletion (single copy deletion) and 88 patients
harbored a LBX2 DNA copy gain (low-level amplification); however
the LBX2 CNA was not significantly correlated with the upregulation
of LBX2 mRNA (P>0.05; Fig. 4A).
The DNA methylation levels of LBX2 between normal colorectal
tissues and CRC samples were compared and no significant difference
was observed (P>0.05; Fig. 4B).
The association between LBX2 mRNA expression levels and LBX2 DNA
methylation was also analyzed. Linear regression analyses indicated
that the DNA methylation levels of LBX2 were correlated moderately
and negatively with LBX2 mRNA expression levels (Pearson's
r=−0.2905; P<0.001; Fig. 4C).
These results suggest that the DNA methylation of LBX2 may not
directly participate in carcinogenesis. However, the upregulation
of LBX2 in CRC may be associated with the hypermethylation of LBX2
and this requires further investigation.
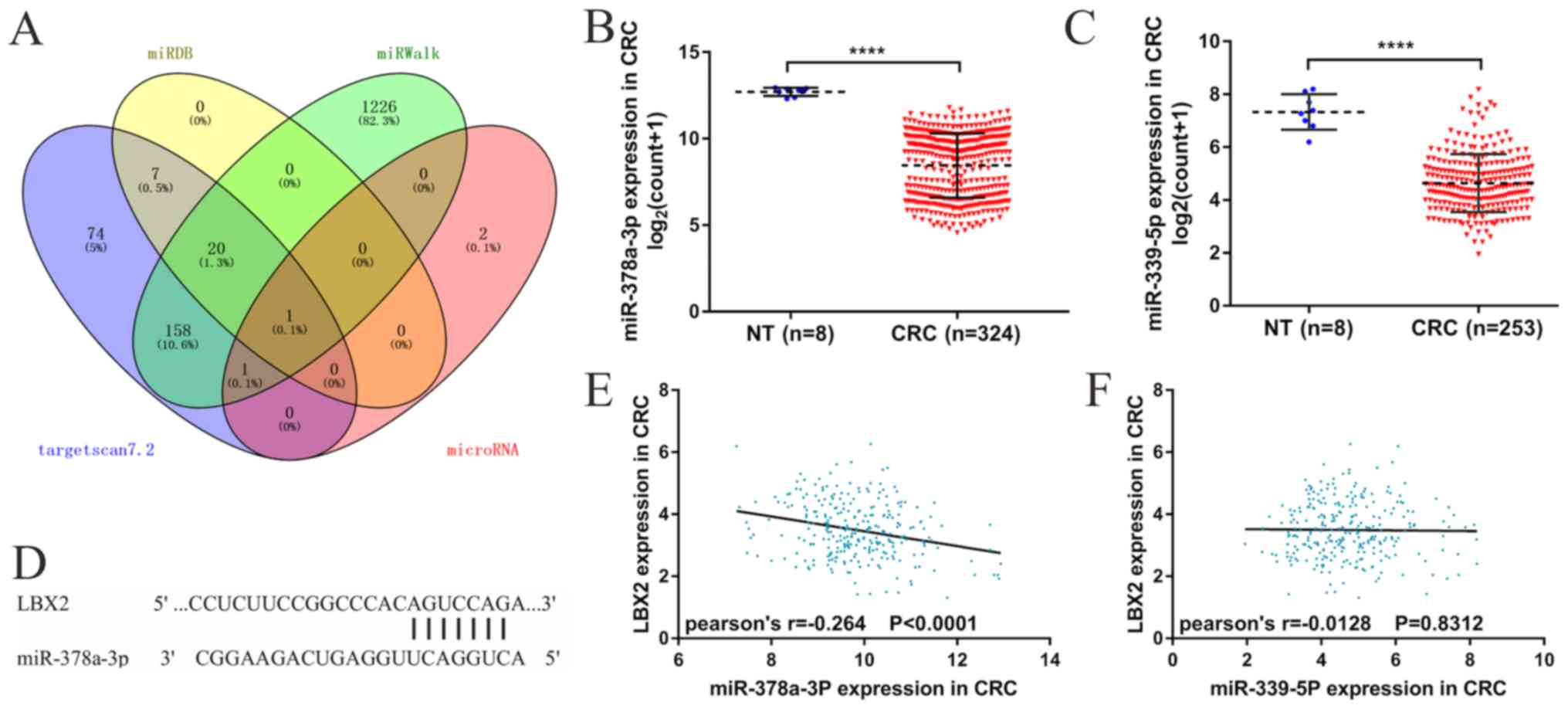
Additionally, the present study sought
to identify the possible regulatory miRNAs of LBX2
Candidate regulatory miRNAs (n=22) of LBX2 were
obtained from four miRNAs target prediction platforms (Fig. 5A). Moreover, data from TCGA database
showed that two miRNAs (miRs), miR-378a-3p and miR-339-5p, of the
22 candidate regulatory miRNAs were significantly downregulated in
CRC tissues compared with normal colorectal tissues and a possible
miR-378a-3p/LBX2 alignment was acquired from miRNAs target
prediction databases (Fig. 5B-D).
Considered as candidates for regulatory miRNAs of LBX2, miR-378a-3p
and miR-339-5p were used for further analyses and validation.
Linear regression analyses revealed that miR-378a-3p expression
levels were correlated moderately and negatively with LBX2 mRNA
expression levels (Pearson's r=−0.264; P<0.001; Fig. 5E), whereas miR-339-5p expression
levels were not significantly correlated with LBX2 mRNA expression
levels (Pearson's r=−0.0128; P=0.8312; Fig. 5E). Kaplan-Meier analyses further
demonstrated that low miR-378a-3p expression levels were linked to
less favorable OS and DFS (all P<0.05; Fig. 6A and B). However, no significance was
observed between miR-339-5p expression levels and OS or DFS (all
P>0.05; Fig. 6C and D). These
results suggest that miR-378a-3p could serve as a potential
regulator of LBX2 in CRC and needs to be researched further.
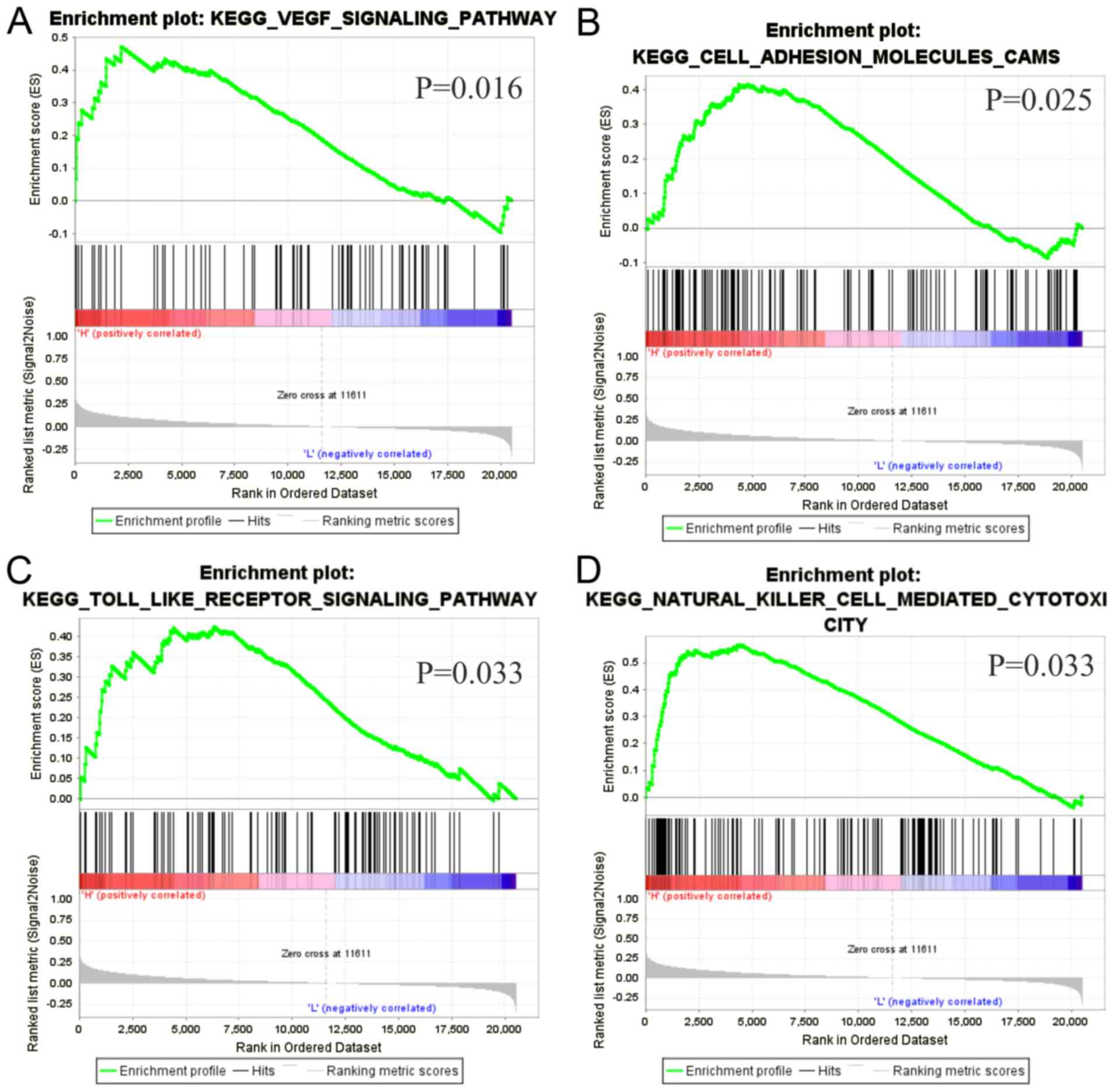
Gene set enrichment analysis
GSEA was performed to investigate the underlying
biological functions of LBX2 upregulation in CRC. Four gene sets,
‘VEGF_SIGNALING_PATHWAY’, ‘CELL_ADHESION_MOLECULES_CAMS’,
‘TOLL_LIKE_ RECEPTOR_SIGNALING_PATHWAY’,
‘NATURAL_LILLER_CELL_MEDIATED_CYTOTOXICITY’, were significantly
enriched (all P<0.05; Fig.
7).
Discussion
In 2018, CRC was reported as the third most commonly
diagnosed type of cancer and the leading cause of cancer-associated
morality worldwide (2). Efforts have
been made in establishing the early detection and comprehensive
therapy of CRC (18). Nevertheless,
individual prognosis prediction remains challenging due to the high
heterogeneity and complex molecular biological characteristics of
CRC (18). Several investigations on
tumor biology from the past few years have provided insights on how
to improve the effectiveness of treatments through patient
stratification, which has improved the prognosis of patients with
CRC (18,19). Furthermore, the availability of
comprehensive genetic and molecular profiling has created promising
treatment opportunities for CRC (19). Hence, it is essential to investigate
the biological characteristics of CRC further, as well as develop
new biomarkers for the stratification of patients with CRC into
different prognostic subgroups to improve treatment decision-making
for CRC.
LBX2 is located on chromosome 2p13.1 and several
previous studies have reported that it is dysregulated in non-small
cell lung cancer, adenoid cystic carcinoma and T-cell acute
lymphoblastic leukemia (11–13). However, the expression levels of LBX2
and its clinical significance in CRC has not been investigated
before, to the best of our knowledge. In the present study, it was
initially demonstrated that LBX2 at the mRNA level was
significantly upregulated in CRC tissues compared with normal
colorectal tissues. Based on ROC curves, the diagnostic value of
LBX2 mRNA expression reached high efficiency, indicating that LBX2
may be a promising biomarker for the diagnosis of CRC.
Investigation into the clinical significance of LBX2
expression levels in CRC tissues revealed that the overexpression
of LBX2 was associated with male patients, advanced tumor stage,
and vascular and lymphatic invasions, suggesting that LBX2 may be
involved in the occurrence and progression of CRC. Kaplan-Meier
analyses showed that LBX2 upregulation was correlated with less
favorable OS and DFS. Multivariate analyses further confirmed that
LBX2 upregulation was an independent risk factor of less favorable
OS in patients with CRC. These results suggest that the expression
levels of LBX2 may be a valuable prognostic indicator and promising
monitoring indicator of postoperative tumor recurrence in patients
with CRC. Approximately 10–35% of patients with CRC suffer a
recurrence within 5 years of diagnosis despite achieving remission
after curative resection and/or chemotherapy (20–22).
Serum carcinoembryonic antigen (CEA) monitoring is
one of the recommended indicators for early detection of disease
recurrence or second primary cancer (23). Although CEA is the most widely used
CRC molecular marker, CEA can also be elevated in other types of
cancer, such as ovarian, pancreatic, gastric, lung and breast
cancer and benign conditions, including inflammatory bowel disease
and pancreatitis (23). Thus,
identifying another clinical biomarker to use independently or
combined with CEA in CRC monitoring for recurrence and metastasis
is important. The analyses of the present study showed that LBX2
was significantly upregulated in CRC tissues compared with normal
colorectal tissues, but the difference in the expression level of
serum LBX2 between patients with CRC and healthy individuals
remains unclear. Therefore, the diagnostic and prognostic values of
independent serum LBX2 or combined with CEA in CRC requires further
study.
The underlying mechanisms of LBX2 upregulation in
CRC were further explored. LBX2 is located on chromosome 2p13.1,
which has been identified differentially expressed in several
malignancies (11–13). From the perspective of genetic and
epigenetic alterations, it was demonstrated that 88 patients with
CRC had low-level LBX2 DNA amplification, but this was not
significantly correlated with LBX2 expression levels. In addition,
DNA methylation is an essential regulator of gene transcription and
hypermethylation represses the transcription of the promoter
regions of tumor suppressor genes leading to gene silencing, while
hypomethylation is considered as a cause of oncogenesis (24). Consistent with this knowledge, the
present study demonstrated that LBX2 DNA hypomethylation was
significantly correlated with the upregulation of LBX2 expression
levels.
Furthermore, miRNAs are critical regulators of gene
expression that can repress gene expression by binding to the 3′
untranslated region (3′UTR) of mRNAs to target them for degradation
and thereby prevent their translation (25). Considering the deregulation of miRNA
functioning involved in several types of human cancer (26), the present study aimed to identify
the candidate regulatory miRNAs of LBX2 and demonstrated that
miR-378-3p and miR-339-5p were significantly downregulated in CRC
samples compared with normal colorectal tissues. Additional linear
regression analyses showed that miR-378-3p expression, but not that
of miR-339-5p, were negatively correlated with LBX2 mRNA expression
levels. Kaplan-Meier analyses also revealed that miR-378-3p
downregulation, but not that of miR-339-5p, was associated with a
less favorable OS and DFS. Several previous studies showed that
miR-378-3p was downregulated in CRC, esophageal squamous cell
carcinoma and prostate cancer (27–29). In
addition, miR-378-3p has been confirmed to be downregulated in CRC
compared with adjacent normal colorectal tissues (28–30),
consistent with the present study, and could serve as an
independent prognostic marker or potential target of novel cancer
therapies. The putative binding of miR-378-3p at the 3′UTR site of
LBX2 further supports the hypothesis that miR-378-3p may be an
upstream regulator of LBX2 in CRC. These results indicate that both
LBX2 DNA hypomethylation and miR-378-3p downregulation contribute
to LBX2 dysregulation in CRC.
The potential biological pathways of LBX2
upregulation in CRC remain to be elucidated further. The findings
of the present study from GSEA suggested that high LBX2 expression
levels in CRC were correlated with ‘VEGF signaling’, ‘Cell adhesion
molecules CAMs’, ‘Toll-like receptor signaling’ and ‘Natural killer
cell-mediated cytotoxicity’ which have been showed to be involved
in CRC development and progression (31–34).
Furthermore, the present study showed that the high expression
levels of LBX2 were significantly associated with advanced tumor
stage of CRC. To the best of our knowledge, no current studies has
investigated the association between LBX2 and CRC. Hence, with
little research on this subject, the underlying regulatory
mechanisms of LBX2 in CRC require further research.
Although the present study investigated several
independent databases, there are some limitations. First, the
clinical significance of LBX2 in CRC was only investigated using
patient samples from TCGA database and lack validation using
further clinical samples. In addition, experiments investigating
the association between LBX2 expression levels and the invasion and
migration of CRC were not performed in the present study. Further
studies are needed to understand the underlying mechanisms of LBX2
function in CRC.
In summary, LBX2 expression levels were
significantly upregulated in CRC tissues compared with normal
colorectal tissues, which might have been caused by LBX2 DNA
hypomethylation and miR-378a-3p downregulation in CRC. Importantly,
the present research indicates that LBX2 overexpression was
associated with tumor progression and was an independent predictor
of less favorable OS and DFS in patients with CRC. The addition of
LBX2 assessment to prognosis may lead to more accurate survival
stratification of patients with CRC and may enable more appropriate
clinical treatment decision-making. Furthermore, LBX2
overexpression was significantly associated with ‘VEGF signaling’,
‘Cell adhesion molecules CAMs’, ‘Toll-like receptor signaling’ and
‘Natural killer cell-mediated cytotoxicity’ pathways in CRC.
Additional studies are needed to clarify the underlying mechanisms
of LBX2 in CRC.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets generated and/or analyzed during the
present study are available in The Cancer Genome Atlas repository
(cancer.gov/tcga).
Authors' contributions
XH and YZ conceived and designed the present study,
performed statistical analysis and drafted the initial manuscript.
YY, CY, HL and HC made substantial contributions to data analysis
and manuscript modifications. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD, Fedewa SA, Ahnen DJ,
Meester RG, Barzi A and Jemal A: Colorectal cancer statistics,
2017. CA Cancer J Clin. 67:177–193. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cancer Genome Atlas Network, .
Comprehensive molecular characterization of human colon and rectal
cancer. Nature. 487:330–337. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mogensen MB, Rossing M, Østrup O, Larsen
PN, Heiberg Engel PJ, Jørgensen LN, Hogdall EV, Eriksen J, Ibsen P,
Jess P, et al: Genomic alterations accompanying tumour evolution in
colorectal cancer: Tracking the differences between primary tumours
and synchronous liver metastases by whole-exome sequencing. BMC
Cancer. 18:7522018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang X, Fang H, Cheng Y, Li L, Sun X, Fu
T, Huang P, Zhang A, Feng Z, Li C, et al: The molecular landscape
of synchronous colorectal cancer reveals genetic heterogeneity.
Carcinogenesis. 39:708–718. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Grasso CS, Giannakis M, Wells DK, Hamada
T, Mu XJ, Quist M, Nowak JA, Nishihara R, Qian ZR, Inamura K, et
al: Genetic mechanisms of immune evasion in colorectal cancer.
Cancer Discov. 8:730–749. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang J, Luo J, Chen Q, Wang X, He J, Zhang
W, Yin Z, Zheng F, Pan H, Li T, et al: Identification of LBX2 as a
novel causal gene of atrial septal defect. Int J Cardiol.
265:188–194. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lou Q, He J, Hu L and Yin Z: Role of lbx2
in the noncanonical Wnt signaling pathway for convergence and
extension movements and hypaxial myogenesis in zebrafish. Biochim
Biophys Acta. 1823:1024–1032. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lu FI, Sun YH, Wei CY, Thisse C and Thisse
B: Tissue-specific derepression of TCF/LEF controls the activity of
the Wnt/β-catenin pathway. Nat Commun. 5:53682014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ochi H and Westerfield M: Lbx2 regulates
formation of myofibrils. BMC Dev Biol. 9:132009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang J, Song J, Gao Z, Huo X, Zhang Y,
Wang W, Qi J and Zheng S: Analysis of gene expression profiles of
non-small cell lung cancer at different stages reveals
significantly altered biological functions and candidate genes.
Oncol Rep. 37:1736–1746. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bell A, Bell D, Weber RS and El-Naggar AK:
CpG island methylation profiling in human salivary gland adenoid
cystic carcinoma. Cancer. 117:2898–2909. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Villarese P, Lours C, Trinquand A, Le Noir
S, Belhocine M, Lhermitte L, Cieslak A, Tesio M, Petit A, LeLorch
M, et al: TCRα rearrangements identify a subgroup of
NKL-deregulated adult T-ALLs associated with favorable outcome.
Leukemia. 32:61–71. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cerami E, Gao J, Dogrusoz U, Gross BE,
Sumer SO, Aksoy BA, Jacobsen A, Byrne CJ, Heuer ML, Larsson E, et
al: The cBio cancer genomics portal: An open platform for exploring
multidimensional cancer genomics data. Cancer Discov. 2:401–404.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Subramanian A, Tamayo P, Mootha VK,
Mukherjee S, Ebert BL, Gillette MA, Paulovich A, Pomeroy SL, Golub
TR, Lander ES, et al: Gene set enrichment analysis: A
knowledge-based approach for interpreting genome-wide expression
profiles. Proc Natl Acad Sci USA. 102:15545–15550. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Camp RL, Dolled-Filhart M and Rimm DL:
X-tile: a new bio-informatics tool for biomarker assessment and
outcome-based cut-point optimization. Clinical cancer research : an
official journal of the American Association for Cancer Research.
10:7252–7259. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Weiser MR: AJCC 8th Edition: Colorectal
Cancer. Ann Surg Oncol. 25:1454–1455. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Punt CJA, Koopman M and Vermeulen L: From
tumour heterogeneity to advances in precision treatment of
colorectal cancer. Nat Rev Clin Oncol. 14:235–246. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sveen A, Kopetz S and Lothe RA:
Biomarker-guided therapy for colorectal cancer: Strength in
complexity. Nat Rev Clin Oncol. 17:11–32. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sargent D, Sobrero A, Grothey A, O'Connell
MJ, Buyse M, Andre T, Zheng Y, Green E, Labianca R, O'Callaghan C,
et al: Evidence for cure by adjuvant therapy in colon cancer:
Observations based on individual patient data from 20,898 patients
on 18 randomized trials. J Clin Oncol. 27:872–877. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Konishi T, Shimada Y, Hsu M, Tufts L,
Jimenez-Rodriguez R, Cercek A, Yaeger R, Saltz L, Smith JJ, Nash
GM, et al: Association of preoperative and postoperative serum
carcinoembryonic antigen and colon cancer outcome. JAMA Oncol.
4:309–315. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yakabe T, Nakafusa Y, Sumi K, Miyoshi A,
Kitajima Y, Sato S, Noshiro H and Miyazaki K: Clinical significance
of CEA and CA19-9 in postoperative follow-up of colorectal cancer.
Ann Surg Oncol. 17:2349–2356. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fletcher RH: Carcinoembryonic antigen. Ann
Intern Med. 104:66–73. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Das PM and Singal R: DNA methylation and
cancer. J Clin Oncol. 22:4632–4642. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lin S and Gregory RI: MicroRNA biogenesis
pathways in cancer. Nat Rev Cancer. 15:321–333. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ding N, Sun X, Wang T, Huang L, Wen J and
Zhou Y: miR 378a 3p exerts tumor suppressive function on the
tumorigenesis of esophageal squamous cell carcinoma by targeting
Rab10. Int J Mol Med. 42:381–391. 2018.PubMed/NCBI
|
|
28
|
Li H, Dai S, Zhen T, Shi H, Zhang F, Yang
Y, Kang L, Liang Y and Han A: Clinical and biological significance
of miR-378a-3p and miR-378a-5p in colorectal cancer. Eur J Cancer.
50:1207–1221. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Valentino A, Calarco A, Di Salle A,
Finicelli M, Crispi S, Calogero RA, Riccardo F, Sciarra A,
Gentilucci A, Galderisi U, et al: Deregulation of MicroRNAs
mediated control of carnitine cycle in prostate cancer: Molecular
basis and pathophysiological consequences. Oncogene. 36:6030–6040.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang G-J, Zhou H, Xiao H-X, Li Y and Zhou
T: MiR-378 is an independent prognostic factor and inhibits cell
growth and invasion in colorectal cancer. BMC Cancer. 14:1092014.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Winder T and Lenz H-J: Vascular
endothelial growth factor and epidermal growth factor signaling
pathways as therapeutic targets for colorectal cancer.
Gastroenterology. 138:2163–2176. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yang LC, Lai CY, Hsieh CC and Lin WC:
Natural killer cell-mediated anticancer effects of an
arabinogalactan derived from rice hull in CT26 colon cancer-bearing
mice. Int J Biol Macromol. 124:368–376. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Paschos KA, Canovas D and Bird NC: The
role of cell adhesion molecules in the progression of colorectal
cancer and the development of liver metastasis. Cell Signal.
21:665–674. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Moradi-Marjaneh R, Hassanian SM, Fiuji H,
Soleimanpour S, Ferns GA, Avan A and Khazaei M: Toll like receptor
signaling pathway as a potential therapeutic target in colorectal
cancer. J Cell Physiol. 233:5613–5622. 2018. View Article : Google Scholar : PubMed/NCBI
|