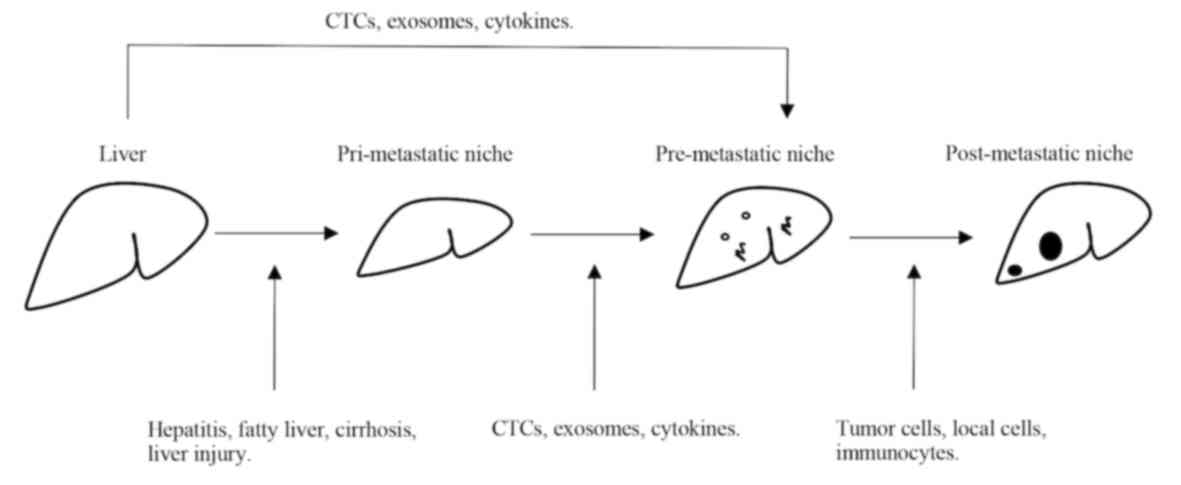
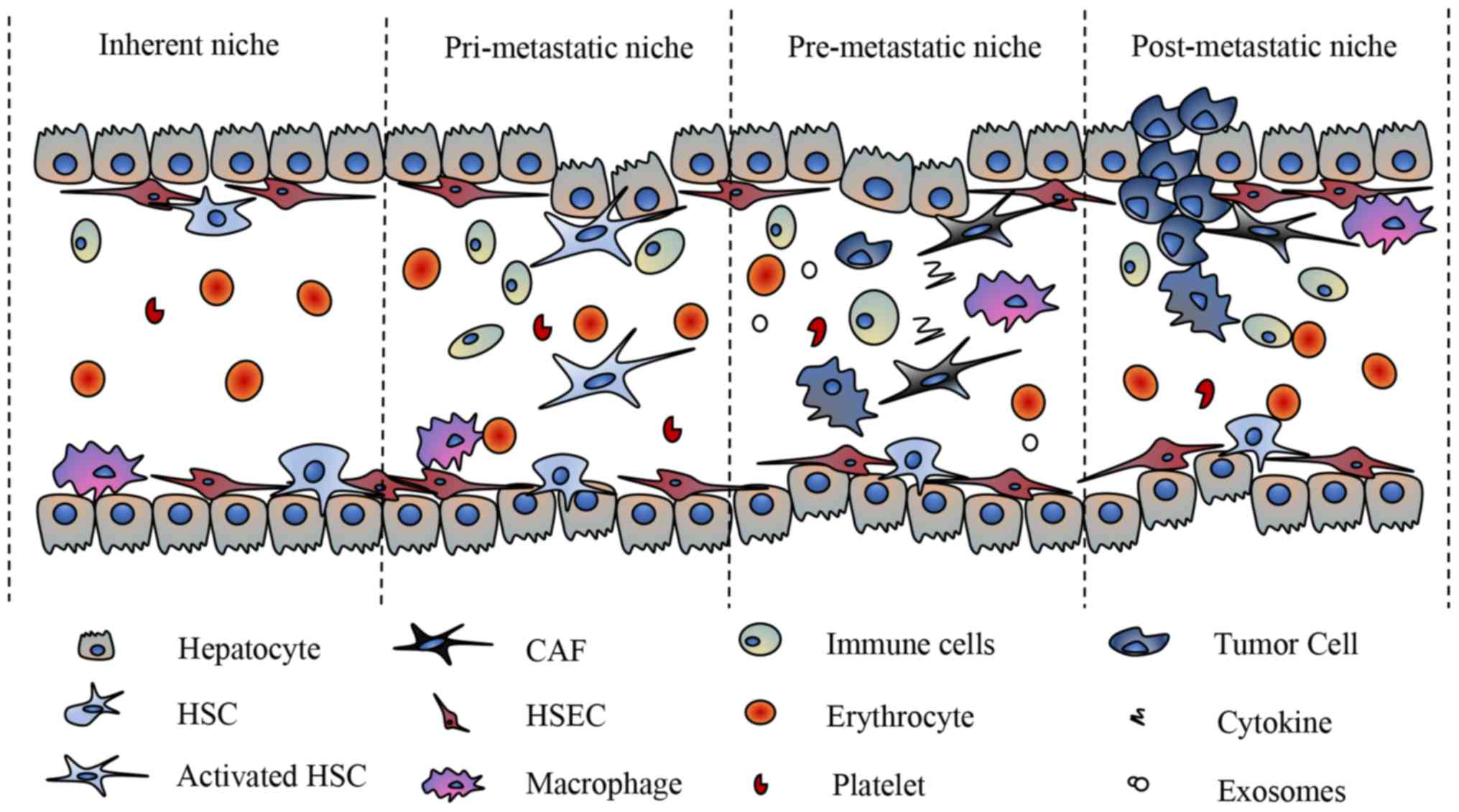
Due to their specific anatomical structures and
biological properties, the majority of the different types of
digestive cancer first metastasise to the liver (1,2). The
timing of, and the molecular determinants underlying the process of
metastasis are largely unknown, and improving the ability to
determine these factors has clinical significance (3). A number of studies (4–6) have
reported that the metastatic microenvironment is altered before
malignant cells metastasise to the liver or lung, forming the
pre-metastatic niche. Furthermore, multiple clinical studies
(7–10) have revealed that patients with
cirrhosis or hepatitis exhibit a lower incidence of hepatic
metastasis despite presenting with the same types of primary
cancer. This indicates that the liver microenvironment may be
modulated by primary liver disease influencing hepatic metastasis,
and this altered microenvironment is defined as the pri-metastatic
niche. After tumor cells migrate to the liver, parenchymal, immune
and mesenchymal cells interact with the tumor cells to modify the
cell state and local microenvironment (including cancer-associated
fibroblasts and macrophages) to perform specific cancer-associated
functions that promote tumor cells colonization, proliferation and
evade immune defence, this is termed the post-metastatic niche
(11–13). These three different niches are the
major components of the IEO model (the pri-, pre- and
post-metastatic niche), as described in Table I, explaining the hepatic metastatic
niche throughout the complete process of tumor progression.
Hepatic metastasis is the primary form of metastasis
in colorectal cancer (CRC) and is the leading cause of
CRC-associated mortality worldwide (3,9).
Colorectal liver metastasis (CRLM) occurs in >25% of patients
diagnosed with primary CRC and 50% of patients during the whole
course of the disease (2–5). Thus, it is important to understand the
underlying mechanisms behind hepatic metastasis, and to develop
measures to prevent or delay this process. According to the seed
and soil theory, the process of metastasis can be stratified into
two major phases (4). The first
phase involves the migration of tumor cells from the primary tumor
site to a targeted metastatic tissue, and the second phase involves
cell proliferation at the site of metastasis (4). This theory emphasises the importance of
the ‘soil’ or microenvironment during the process of metastasis
(12). The liver microenvironment is
comprised of numerous components, including hepatocytes, Kupffer
cells (KCs), hepatic sinusoidal endothelial cells (HSECs), hepatic
stellate cells (HSCs), pit cells and the extracellular matrix (ECM)
(11). The tumor-associated hepatic
niche is regulated via two mechanisms; the tumor-associated
microenvironment formed by the primary tumor lesions and the
intrinsic microenvironment formed by liver disease (7,14,15).
Specific stages include the recruitment of fibroblasts, migration
of immune cells, matrix remodelling and the development of vascular
networks (11). The metastatic cells
pass through the portal vein system into the hepatic sinusoid, and
the anchoring of the circulating tumor cells (CTCs) to the hepatic
sinus, and its attached cells (HSECs, HSCs and KCs), affects
invasion and proliferation (1,4).
Additionally, blood flow in the hepatic sinusoids is regulated by
the vasoconstrictive properties of HSCs that control the oxygen and
nutrient supply (5,16). Fig. 1
provides an overview of the processes underlying pathological
metastasis that form the basis of the IEO model, as described in
detail in the following sections.
The target organ, with its inherent microenvironment
influenced by chronic or acute local disease states, may further
affect the occurrence of cancer metastasis from primary tumors
(5,7–9). The
local microenvironment at the metastatic site forms the
pri-metastatic niche. Various diseases affecting the liver (such as
fatty liver, cirrhosis, liver steatosis, hepatitis B and C
infection or acute liver injury) may affect the liver
microenvironment and the incidence of hepatic metastasis (14,15,17).
Accordingly, the pri-metastatic niche serves an important role in
the process of hepatic metastasis.
A study of 5,092 autopsies of colorectal, breast or
lung tumors indicated that the incidence of liver metastasis is
28.6% compared with non-cirrhosis cases, which is higher than the
4.5% observed in patients with cirrhosis (8). Liver metastasis from CRC is also
infrequent in patients with cirrhosis (10%) compared with
non-cirrhosis cases (25%), indicating that liver cirrhosis
substantially reduces the rate of CRLM, perhaps due to the
differential properties of the liver microenvironment (10). Moreover, a meta-analysis of data
concerning 10,349 patients with CRC from 10 studies was performed
to investigate the association between CRLM and local liver
disease. Based on this meta-analysis, chronically diseased livers
(fatty liver, cirrhosis or chronic hepatitis B and C virus
infection) exhibited a pri-metastatic niche and a significantly
lower incidence of CRLM (14). The
mechanism may involve the remodelling of the inherent liver
microenvironment components, including fibrosis and the hepatic
sinus. Activated immune cells residing in the liver
microenvironment of the diseased liver kill metastatic tumor cells
that circulate in the bloodstream. During cirrhosis, KCs release
pro-inflammatory cytokines to remodel the hepatic immune niche,
such as tumor necrosis factor (TNF)-α and interleukin (IL)-1
(14). In an analysis of rats with
cirrhosis, Song et al (15)
revealed that KCs and CRC cells exhibited upregulated Fas/FasL
protein expression, which induced cell apoptosis, and the apoptotic
cancer cells are further targeted by infiltrating lymphocytes.
Thus, while the activation of KCs in cirrhosis participates in
tissue damage and fibrogenesis, it also exerts a protective effect
by inhibiting the hepatic metastasis of colon cancer. Seitz
(16) reported that liver cirrhosis
is associated with high metalloproteinase inhibitor levels and
decreased levels of lectins or lectin-binding sites. Patients with
liver cirrhosis reduced blood flow from the portal vein that may
decreases tumor cell migration to the liver, which may contribute
to rare occurrences of liver metastasis. However, cytokine analysis
of metastasis of pancreatic cancer to the liver has indicated that
in the early phase of metastasis, pancreatic ductal cancer cells
decrease IL-6/signal transducer and activator of transcription 3
signalling via a negative feedback loop to construct the
microenvironment of liver fibrosis and attract bone marrow-derived
cells, promoting pancreatic duct engraftment to the liver (18).
Patients with CRC infected with hepatitis B virus
(HBV) or hepatitis C virus exhibit a lower incidence of metastasis
(8.1%) and longer survival time compared with that in patients
without infection (21.2%), although they may have a higher
probability of developing liver cancer (19). HBV may affect liver-associated
immunity and increase cytotoxic liver activity on metastatic cells
mediated by T cells, KCs or TNF-α synthesized by liver cells
(8,20,21).
Metalloproteinase inhibitors, such as metalloproteinase tissue
inhibitor-1, have also been isolated from myofibroblasts of
diseased livers (cirrhosis and hepatitis) but less so from healthy
livers. Disease livers with more TIMP1 may inhibit MMPs expression
from tumor cells, and this may explain the lower incidence of
metastasis observed in diseased livers (22,23).
Furthermore, HBV activates cytotoxic T-cells and KCs, which have
the potential to kill metastatic tumor cells when they pass the
liver sinuses (24–26). Additionally, HBV may stimulate HBV
specific T cells to increase the secretion of TNF-α (27).
Non-alcoholic fatty liver disease (NAFLD) and
alcoholic liver disease are major health issues associated with
tumor cell metastasis to the liver (28). According to previous studies, high
alcohol intake is associated with CRLM (29,30).
Both KCs and hepatocytes are affected by alcohol, and high
expression of lipopolysaccharides and pro-inflammatory cytokines,
such as TNFα and IL-1β changes the hepatic niche and CRC metastasis
(28). NAFLD has been used to
demonstrate the higher metastatic burden in steatotic livers
compared with that in normal livers. NAFLD is characterized by fat
accumulation, which alters the local hepatic niche by stimulating
triglyceride, recruiting inflammatory cells, increasing TNFα
expression and disrupting the normal structure of ECM. These
alterations of the liver microenvironment, including
tumor-associated inflammatory cells and aberrant ECM structure may
promote colonization of tumor cells and cancer progression with
poor outcome (28–30). However, Karube et al (31) demonstrated that fat metabolism
disorders inhibit tumor cell proliferation and angiogenesis to
prevent tumor growth and reduce the likelihood of metastasis.
Tumor-derived exosomes are composed of proteins,
mRNAs and microRNAs that regulate pre-metastatic niche formation
and affect metastasis (35–37). In the metastatic process, exosomes
also affect epithelial-mesenchymal transition, cancer stemness,
apoptosis and metastatic angiogenesis via CXCR, integrins or the
TGF-β signaling pathway (38–40).
Wang et al (11) reported
that CRC tumor-derived exosomes affect the hepatic niche and
increase CXCR4 expression in stromal cells, thus creating a
CXCR4-enriched microenvironment suitable for metastasis (11). In addition, pancreatic cancer cells
release migration inhibitory factor-associated exosomes that induce
TGF-β secretion, which results in the production of the
glycoprotein fibronectin by HSCs, and the aggregation of bone
marrow-derived cells to promote hepatic metastasis (33). Integrins in exosomes influence
metastasis to specific organs and prepare the microenvironment for
tumor cell arrival, such as integrin β5 in liver metastasis and
integrin α6 in lung metastasis (41). In addition to tumor-associated
exosomes, CTCs are also necessary elements for successful
metastasis (18). CTCs circulate to
the targeted metastatic organ, evade immune defence via TGF-β
associated signaling pathways and platelet protection, arrive at
the supportive niche and serve as latent tumor seeds in the niche,
ultimately proliferating in the host tissue to form the metastatic
lesion and altering the liver microenvironment (42,43).
CTCs regulate the metastatic microenvironment via cytokine
secretion, including TGF-β and IL-1. The perivascular space around
the small blood vessels supports metastasis; it facilitates the
proliferation of CTCs and hinders antitumor therapy (44,45). The
cytokine signals secreted by the primary tumor may affect the
microenvironment of distant organs and form a pre-metastasis niche
prior to the arrival of CTCs (46,47).
Physical contact between stromal cells and tumor cells, such as the
claudin-2-mediated bridge between metastatic cancer cells and
hepatocytes, induces c-Met signalling and hepatic metastasis in
breast cancer (48). Hepatic
sinusoids lined by endothelial cells and basal lamina gaps
(49) may support the extravasation
of CTCs and result in liver and bone metastasis (50,51). The
major factors in anti-metastatic immunity are cytotoxic T and NK
cells (52). Furthermore, the liver
has a specific immune cell composition, characterised by abundant
NK cells, which affects the susceptibility of the target organ to
metastasis (4). Compared with levels
in the normal mucosa, expression of the inflammatory mediator
cyclooxygenase- is upregulated in CRC and to a greater extent in
hepatic metastases (53), indicating
that inflammation affects disease progression and may serve as a
clinical indicator for malignant tumors (54). In HSECs, the vascular cell adhesion
protein-1 blockade decreases microvascular formation of the hepatic
metastatic lesion (53,54). CTCs, circulating free-DNA, miRNAs and
exosomes may potentially be used for the development of critical
assays for the early detection of metastasis in patients with CRC
and as a therapeutic target.
After tumor cells metastasize to the liver, they
interact with the liver niche and adapt to their new
microenvironment, which is called the post-metastatic niche
(Fig. 2). In the new and challenging
metastatic niche, newly established tumor cells must acquire the
ability to survive immune cell attacks (55–58).
The interaction between the hepatic innate immune
response and tumor cells is a double-edged sword for tumor
metastasis (55,56). Although an initially effective
defence can inhibit CTCs via the cytotoxic attack of KCs and NK
cells (57), immune cells also
promote tumor invasiveness and metastasis via various mechanisms,
such as the activation of angiogenesis (55) and a pro-tumorigenic phenotype to
promote tumor cell proliferation (56,58).
Neutrophils or tumor-associated neutrophils exhibit high levels of
plasticity in order to regulate the tumor microenvironment
(59). They are also associated with
the formation of the pancreatic liver metastatic microenvironment
(60). The neutrophil subtype that
infiltrates at the early phase of metastasis recruits macrophages
and fibroblasts, and promotes the formation of metastatic lesions
(61). Neutrophils promote tumor
invasion (59), and tumor-associated
neutrophils express immunosuppressive factors, such as TGF-β and
FGF2 (62). CXCR2 protein expression
in neutrophils during liver metastasis serves an important role in
the early phase of tumor development and accelerates fibroblast
anchoring to tumor cells in the hepatic sinus (60). Tumor-associated neutrophils may
promote fibroblast growth factor 2 (FGF2), which is primarily
expressed in and released from the ECM (12). The normalisation of microvessels in
the tumor microenvironment is associated with FGF2 (63), which promotes vascular formation in
liver metastases and induces an immune response in endothelial
cells to recruit more immune cells (64). Tumor-infiltrating lymphocytes (TILs),
an indicator of the anticancer immune response, influence cancer
progression, metastasis and chemoresistance and are superior in the
TNM classification as a predictor of survival in patients with
digestive and lung cancer (65). A
high number of TILs in the metastatic tumor is associated with
improved clinical outcomes, overall response rates and
chemotherapeutic outcomes (65).
Moreover, patients with CRC exhibit reduced sensitivity to
programmed cell death protein 1 (PD-1) and programmed cell death 1
ligand 1 (PD-L1), with the exception of patients with mismatch
repair (MMR) genes deficiencies, such as MSH2, MSH6, MLH1 or PMS2
gene (66). The unique immune
microenvironment in the liver forms a special immune tolerance
type. The mechanism underlying immune tolerance involves the
induction of surface immune suppression ligands of T cells and
suppression of the immune receptor expression in liver cells and
HSCs (65–67). The post-metastasis microenvironment
may enhance immune cell infiltration and increase immunosuppression
checkpoints so that sensitivity to PD-1/PD-L1 is greater in
MMR-deficient CRC compared with that in MMR-proficient CRC
(67). Additionally, decreased TGF-β
protein expression levels in the post-metastatic niche may increase
the activation of cytotoxic T cell-dependent processes and change
MMR-proficient CRC cells to immune-hot cells (68). During metastasis, α-SMA-positive
stromal cells present on the interstitial surface of the tumor, and
the residual fibroblasts differentiate into myofibroblasts that
express collagen in the periphery (69,70).
According to microenvironmental change, three distinct growth
patterns have been described in CRC adenocarcinoma liver metastases
(69).
In the replacement growth pattern, the liver
structure is preserved. In the pushing growth pattern, the hepatic
lobules extend to one side and the liver stromal cells surround the
metastatic lesion, and in the desmoplastic growth pattern, a
fibrous ring separates the hepatic stromal tissue from the
metastatic lesion (70). The
survival and proliferation of cancer cells at the metastatic site
is important for the establishment of metastatic tumors and the
re-expression of E-cadherin on cancer cells at the metastatic site
promotes proliferation in breast cancer (71). Further, if the hepatic sinus blood
vessel is blocked by a large number of tumor cells and blood flow
is obstructed, the inflammatory response of ischemia-reperfusion is
initiated (72,73). This may result in the release of
nitric oxide (NO) and reactive oxygen species in local HSECs and
KCs (72,73). The release of NO and interferon-γ by
HSECs entering the hepatic sinus results in the upregulation of
FasL, initiating apoptosis in 95% of metastatic tumor cells
(74).
The liver is the most common site of metastasis for
various types gastrointestinal cancers from the portal vein system
(2–4,41–43). A
deeper understanding of the process of metastasis and cancer
recurrence may be beneficial for the identification of novel
treatment strategies. A novel concept for the metastatic
environment referred to as the IEO model (pri-, pre- and
post-metastatic niche) to explain the key steps in metastasis has
been described in the present review. The liver microenvironment is
formed by invading tumor cells and the immune system. Local cells
in the liver and tumor cells develop complex interaction networks
and adaptions, which may either inhibit or promote tumor
metastasis.
The IEO model provides novel insights into the
prevention of tumor metastasis by identifying interventions
targeting the liver microenvironment mediated by liver diseases.
The model may be used to establish a comprehensive disease
management system for the prevention and treatment of liver
metastasis, from a microenvironmental perspective.
Not applicable.
The present study was funded by the National Natural
Science Foundation of China (grant no. 81773360).
Not applicable.
LW contributed to the conception of the study,
writing the manuscript and performing the literature search. YS, MY
and WZ conducted analysis and revised the manuscript. XY performed
analysis and the quality assessment of the study. All authors read
and approved the final manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Denève E, Riethdorf S, Ramos J, Nocca D,
Coffy A, Daurès JP, Maudelonde T, Fabre JM, Pantel K and
Alix-Panabières C: Capture of viable circulating tumor cells in the
liver of colorectal cancer patients. Clin Chem. 59:1384–1392. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Obenauf AC and Massaguè J: Surviving at a
distance: Organ specific metastasis. Trends Cancer. 1:76–91. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hu Z, Ding J, Ma Z, Sun R, Seoane JA,
Scott Shaffer J, Suarez CJ, Berghoff AS, Cremolini C, Falcone A, et
al: Quantitative evidence for early metastatic seeding in
colorectal cancer. Nat Genet. 51:1113–1122. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Massaguè J and Obenauf AC: Metastatic
colonization by circulating tumor cells. Nature. 529:298–306. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Van den Eynden GG, Majeed AW, Illemann M,
Vermeulen PB, Bird NC, Høyer-Hansen G, Eefsen RL, Reynolds AR and
Brodt P: The multifaceted role of the microenvironment in liver
metastasis: Biology and clinical implications. Cancer Res.
73:2031–2043. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Guo Y, Ji X, Liu J, Fan D, Zhou Q, Chen C,
Wang W, Wang G, Wang H, Yuan W, et al: Effects of exosomes on
pre-metastatic niche formation in tumors. Mol Cancer. 18:392019.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li Destri G, Castaing M, Ferlito F,
Minutolo V, Di Cataldo A and Puleo S: Rare hepatic metastases of
colorectal cancer in livers with symptomatic HBV and HCV hepatitis.
Ann Ital Chir. 84:323–327. 2013.PubMed/NCBI
|
|
8
|
Qiu HB, Zhang LY, Zeng ZL, Wang ZQ, Luo
HY, Keshari RP, Zhou ZW and Xu RH: HBV infection decreases risk of
liver metastasis in patients with colorectal cancer: A cohort
study. World J Gastroenterol. 17:804–808. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kondo T, Okabayashi K, Hasegawa H, Tsuruta
M, Shigeta K and Kitagawa Y: The impact of hepatic fibrosis on the
incidence of liver metastasis from colorectal cancer. Br J Cancer.
115:34–39. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chiou WY, Chang CM, Tseng KC, Hung SK, Lin
HY, Chen YC, Su YC, Tseng CW, Tsai SJ, Lee MS and Li CY: Effect of
liver cirrhosis on metastasis in colorectal cancer patients: A
nationwide population-based cohort study. Jpn J Clin Oncol.
45:160–168. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang X, Ding X, Nan L, Wang Y, Wang J, Yan
Z, Zhang W, Sun J, Zhu W, Ni B, et al: Investigation of the roles
of exosomes in colorectal cancer liver metastasis. Oncol Rep.
33:2445–2453. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gordon-Weeks AN, Lim SY, Yuzhalin AE,
Jones K, Markelc B, Kim KJ, Buzzelli JN, Fokas E, Cao Y, Smart S
and Muschel R: Neutrophils promote hepatic metastasis growth
through fibroblast growth factor 2-dependent angiogenesis in mice.
Hepatology. 65:1920–1935. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mendonsa AM, VanSaun MN, Ustione A, Piston
DW, Fingleton BM and Gorden DL: Host and tumor derived MMP13
regulate extravasation and establishment of colorectal metastases
in the liver. Mol Cancer. 14:492015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cai B, Liao K, Song XQ, Wei WY, Zhuang Y
and Zhang S: Patients with chronically diseased livers have lower
incidence of colorectal liver metastases: A meta-analysis. PLoS
One. 9:e1086182014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Song E, Chen J, Ouyang N, Wang M, Exton MS
and Heemann U: Kupffer cells of cirrhotic rat livers sensitize
colon cancer cells to Fas mediated apoptosis. Br J Cancer.
84:1265–1271. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Seitz G: Why are metastases in cirrhotic
livers so rare? Ultraschall Med. 10:123–126. 1989.(In German).
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Seymour K and Charnley RM: Evidence that
metastasis is less common in cirrhotic than normal liver: A
systematic review of post-mortem case-control studies. Br J Surg.
86:1237–1242. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lee JW, Stone ML, Porrett PM, Thomas SK,
Komar CA, Li JH, Delman D, Graham K, Gladney WL, Hua X, et al:
Hepatocytes direct the formation of a pro-metastatic niche in the
liver. Nature. 567:249–252. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Utsunomiya T, Saitsu H, Saku M, Yoshida K,
Matsumata T, Shimada M and Sugimachi K: Rare occurrence of
colorectal cancer metastasis in livers infected with hepatitis B or
C virus. Am J Surg. 177:279–281. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Qian HG, Zhang J, Leng JH, Zhou GQ, Wu JH,
Tian XY, Yang Y and Hao CY: Association of hepatitis B virus
infection and cirrhosis with liver metastasis in colorectal cancer.
Zhonghua Wei Chang Wai Ke Za Zhi. 13:202–204. 2010.(In Chinese).
PubMed/NCBI
|
|
21
|
Song E, Chen J, Ou Q and Su F: Rare
occurrence of metastatic colorectal cancers in livers with
replicative hepatitis B infection. Am J Surg. 181:529–533. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zeng ZS, Sun Y, Shu WP and Guillem JG:
Tissue inhibitor of metalloproteinase-3 is a basement
membrane-associated protein that is significantly decreased in
human colorectal cancer. Dis Colon Rectum. 44:1290–1296. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cheng L, Geng L, Dai B, Zheng T, Fu J,
Qiao L, Cai W, Wang Y and Yang J: Repression of let-7a cluster
prevents adhesion of colorectal cancer cells by enforcing a
mesenchymal phenotype in presence of liver inflammation. Cell Death
Dis. 9:4892018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Uetsuji S, Yamamura M, Yamamichi K, Okuda
Y, Takada H and Hioki K: Absence of colorectal cancer metastasis to
the cirrhotic liver. Am J Surg. 164:176–177. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wu JF and Chang MH: Natural history of
chronic hepatitis B virus infection from infancy to adult life-the
mechanism of inflammation triggering and long-term impacts. J
Biomed Sci. 22:922015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tordjmann T, Soulie A, Guettier C, Schmidt
M, Berthou C, Beaugrand M and Sasportes M: Perforin and granzyme B
lytic protein expression during chronic viral and autoimmune
hepatitis. Liver. 18:391–397. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lara-Pezzi E, Majano PL, Gómez-Gonzalo M,
García-Monzón C, Moreno-Otero R, Levrero M and López-Cabrera M: The
hepatitis B virus X protein up-regulates tumor necrosis factor
alpha gene expression in hepatocytes. Hepatology. 28:1013–1021.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
McVicker B, Tuma DJ, Lazure KE, Thomas P
and Casey CA: Alcohol, carcinoembryonic antigen processing and
colorectal liver metastases. Adv Exp Med Biol. 815:295–311. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Maeda M, Nagawa H, Maeda T, Koike H and
Kasai H: Alcohol consumption enhances liver metastasis in
colorectal carcinoma patients. Cancer. 83:1483–1488. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Dong H, Tang J, Li LH, Ge J, Chen X, Ding
J, Men HT, Luo WX, Du Y, Li C, et al: Serum carbohydrate antigen
19-9 as an indicator of liver metastasis in colorectal carcinoma
cases. Asian Pac J Cancer Prev. 14:909–913. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Karube H, Masuda H, Hayashi S, Ishii Y and
Nemoto N: Fatty liver suppressed the angiogenesis in liver
metastatic lesions. Hepatogastroenterology. 47:1541–1545.
2000.PubMed/NCBI
|
|
32
|
Pancione M, Giordano G, Remo A, Febbraro
A, Sabatino L, Manfrin E, Ceccarelli M and Colantuoni V: Immune
escape mechanisms in colorectal cancer pathogenesis and liver
metastasis. J Immunol Res. 2014:6868792014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Alderton GK: Metastasis. Exosomes drive
premetastatic niche formation. Nat Rev Cancer. 12:4472012.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kosaka N, Iguchi H, Hagiwara K, Yoshioka
Y, Takeshita F and Ochiya T: Neutral sphingomyelinase 2
(nSMase2)-dependent exosomal transfer of angiogenic microRNAs
regulate cancer cell metastasis. J Biol Chem. 288:10849–10859.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Théry C, Ostrowski M and Segura E:
Membrane vesicles as conveyors of immune responses. Nat Rev
Immunol. 9:581–593. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Valadi H, Ekström K, Bossios A, Sjöstrand
M, Lee JJ and Lötvall JO: Exosome-mediated transfer of mRNAs and
microRNAs is a novel mechanism of genetic exchange between cells.
Nat Cell Biol. 9:654–672. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Bang C and Thum T: Exosomes: New players
in cell-cell communication. Int J Biochem Cell Biol. 44:2060–2064.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hood JL, Pan H, Lanza GM and Wickline SA;
Consortium for Translational Research in Advanced Imaging and
Nanomedicine (C-TRAIN), : Paracrine induction of endothelium by
tumor exosomes. Lab Invest. 89:1317–1328. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Balaj L, Lessard R, Dai L, Cho YJ, Pomeroy
SL, Breakefield XO and Skog J: Tumor microvesicles contain
retrotransposon elements and amplified oncogene sequences. Nat
Commun. 2:1802011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Aliotta JM, Pereira M, Johnson KW, de Paz
N, Dooner MS, Puente N, Ayala C, Brilliant K, Berz D, Lee D, et al:
Microvesicle entry into marrow cells mediates tissue-specific
changes in mRNA by direct delivery of mRNA and induction of
transcription. Exp Hematol. 38:233–245. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hoshino A, Costa-Silva B, Shen TL,
Rodrigues G, Hashimoto A, Tesic Mark M, Molina H, Kohsaka S, Di
Giannatale A, Ceder S, et al: Tumor exosome integrins determine
organotropic metastasis. Nature. 527:329–335. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Labelle M, Begum S and Hynes RO: Direct
signaling between platelets and cancer cells induces an
epithelial-mesenchymal-like transition and promotes metastasis.
Cancer Res. 20:579–560. 2011.
|
|
43
|
Houg DS and Bijlsma MF: The hepatic
pre-metastatic niche in pancreatic ductal adenocarcinoma. Mol
Cancer. 17:952018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Hambardzumyan D, Becher OJ, Rosenblum MK,
Pandolfi PP, Manova-Todorova K and Holland EC: PI3K pathway
regulates survival of cancer stem cells residing in the
perivascular niche following radiation in medulloblastoma in vivo.
Genes Dev. 22:436–448. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Cao ZW, Ding BS, Guo PP, Lee SB, Butler
JM, Casey SC, Simons M, Tam W, Felsher DW, Shido K, et al:
Angiocrine factors deployed by tumor vascular niche induce B cell
lymphoma invasiveness and chemoresistance. Cancer Res. 25:350–365.
2014.
|
|
46
|
Kaplan RN, Riba RD, Zacharoulis S, Bramley
AH, Vincent L, Costa C, MacDonald DD, Jin DK, Shido K, Kerns SA, et
al: VEGFR1-positive haematopoietic bone marrow progenitors initiate
the pre-metastatic niche. Nature. 438:820–827. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
McAllister SS and Weinberg RA: The
tumor-induced systemic environment as a critical regulator of
cancer progression and metastasis. Nat Cell Biol. 16:717–727. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Tabariés S, Dupuy F, Dong Z, Monast A,
Annis MG, Spicer J, Ferri LE, Omeroglu A, Basik M, Amir E, et al:
Claudin-2 promotes breast cancer liver metastasis by facilitating
tumor cell interactions with hepatocytes. Mol Cell Biol.
32:2979–2991. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Aird WC: Phenotypic heterogeneity of the
endothelium: II. Representative vascular beds. Circ Res.
100:174–190. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Nguyen DX, Bos PD and Massagué J:
Metastasis: From dissemination to organ-specific colonization. Nat
Rev Cancer. 9:274–284. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Budczies J, von Winterfeld M, Klauschen F,
Bockmayr M, Lennerz JK, Denkert C, Wolf T, Warth A, Dietel M,
Anagnostopoulos I, et al: The landscape of metastatic progression
patterns across major human cancers. Oncotarget. 6:570–583. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Eyles J, Puaux AL, Wang X, Toh B, Prakash
C, Hong M, Tan TG, Zheng L, Ong LC, Jin Y, et al: Tumor cells
disseminate early, but immunosurveillance limits metastatic
outgrowth, in a mouse model of melanoma. J Clin Invest.
120:2030–2039. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Chen WS, Wei SJ, Liu JM, Hsiao M, Kou-Lin
J and Yang WK: Tumor invasiveness and liver metastasis of colon
cancer cells correlated with cyclooxygenase-2 (COX-2) expression
and inhibited by a COX-2-selective inhibitor, etodolac. Int J
Cancer. 91:894–899. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Wang D, Sun H, Wei J, Cen B and DuBois RN:
CXCL1 is critical for premetastatic niche formation and metastasis
in colorectal cancer. Cancer Res. 77:3655–3665. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Murdoch C, Muthana M, Coffelt SB and Lewis
CE: The role of myeloid cells in the promotion of tumor
angiogenesis. Nat Rev Cancer. 8:618–631. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Flavell RA, Sanjabi S, Wrzesinski SH and
Licona-Limón P: The polarization of immune cells in the tumor
environment by TGFbeta. Nat Rev Immunol. 10:554–567. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Luzzi KJ, MacDonald IC, Schmidt EE,
Kerkvliet N, Morris VL, Chambers AF and Groom AC: Multistep nature
of metastatic inefficiency: Dormancy of solitary cells after
successful extravasation and limited survival of early
micrometastases. Am J Pathol. 153:865–873. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Brodt P: Role of the microenvironment in
liver metastasis: From pre- to prometastatic niches. Clin Cancer
Res. 22:5971–5982. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Fridlender ZG, Sun J, Kim S, Kapoor V,
Cheng G, Ling L, Worthen GS and Albelda SM: Polarization of
tumor-associated neutrophil phenotype by TGF-beta: ‘N1’ versus ‘N2’
TAN. Cancer Res. 16:183–194. 2009.
|
|
60
|
Steele CW, Karim SA, Leach JDG, Bailey P,
Upstill-Goddard R, Rishi L, Foth M, Bryson S, McDaid K, Wilson Z,
et al: CXCR2 inhibition profoundly suppresses metastases and
augments immunotherapy in pancreatic ductal adenocarcinoma. Cancer
Res. 29:832–845. 2016.
|
|
61
|
Hirai H, Fujishita T, Kurimoto K, Miyachi
H, Kitano S, Inamoto S, Itatani Y, Saitou M, Maekawa T and Taketo
MM: CCR1-mediated accumulation of myeloid cells in the liver
microenvironment promoting mouse colon cancer metastasis. Clin Exp
Metastasis. 31:977–989. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Elpek KG, Cremasco V, Shen H, Harvey CJ,
Wucherpfennig KW, Goldstein DR, Monach PA and Turley SJ: The tumor
microenvironment shapes lineage, transcriptional, and functional
diversity of infiltrating myeloid cells. Cancer Immunol Res.
2:655–667. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Yuan H, Li ZM, Shao J, Ji WX, Xia W and Lu
S: FGF2/FGFR1 regulates autophagy in FGFR1-amplified non-small cell
lung cancer cells. J Exp Clin Cancer Res. 36:722017. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Presta M, Andrés G, Leali D, Dell'Era P
and Ronca R: Inflammatory cells and chemokines sustain FGF2-induced
angiogenesis. Eur Cytokine Netw. 20:39–50. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Shibutani M, Maeda K, Nagahara H, Fukuoka
T, Matsutani S, Kashiwagi S, Tanaka H, Hirakawa K and Ohira M: A
comparison of the local immune status between the primary and
metastatic tumor in colorectal cancer: A retrospective study. BMC
Cancer. 18:3712018. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Le DT, Durham JN, Smith KN, Wang H,
Bartlett BR, Aulakh LK, Lu S, Kemberling H, Wilt C, Luber BS, et
al: Mismatch repair deficiency predicts response of solid tumors to
PD-1 blockade. Science. 357:409–413. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Zhou G, Noordam L, Sprengers D, Doukas M,
Boor PPC, van Beek AA, Erkens R, Mancham S, Grünhagen D, Menon AG,
et al: Blockade of LAG3 enhances responses of tumor-infiltrating T
cells in mismatch repair-proficient liver metastases of colorectal
cancer. Oncoimmunology. 7:e14483322018. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Tauriello DVF, Palomo-Ponce S, Stork D,
Berenguer-Llergo A, Badia-Ramentol J, Iglesias M, Sevillano M,
Ibiza S, Cañellas A, Hernando-Momblona X, et al: TGFβ drives immune
evasion in genetically reconstituted colon cancer metastasis.
Nature. 554:538–543. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Vermeulen PB, Colpaert C, Salgado R,
Royers R, Hellemans H, Van Den Heuvel E, Goovaerts G, Dirix LY and
Van Marck E: Liver metastases from colorectal adenocarcinomas grow
in three patterns with different angiogenesis and desmoplasia. J
Pathol. 195:336–342. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Bugyik E, Szabó V, Dezső K, Rókusz A,
Szücs A, Nagy P, Tóvári J, László V, Döme B and Paku S: Role of
(myo)fibroblasts in the development of vascular and connective
tissue structure of the C38 colorectal cancer in mice. Cancer
Commun (Lond). 38:462018. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Padmanaban V, Krol I, Suhail Y, Szczerba
BM, Aceto N, Bader JS and Ewald AJ: E-cadherin is required for
metastasis in multiple models of breast cancer. Nature.
573:439–444. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Wang HH, McIntosh AR, Hasinoff BB, Rector
ES, Ahmed N, Nance DM and Orr FW: B16 melanoma cell arrest in the
mouse liver induces nitric oxide release and sinusoidal
cytotoxicity: A natural hepatic defense against metastasis. Cancer
Res. 60:5862–5869. 2000.PubMed/NCBI
|
|
73
|
Yanagida H, Kaibori M, Yoshida H, Habara
K, Yamada M, Kamiyama Y and Okumura T: Hepatic ischemia/reperfusion
upregulates the susceptibility of hepatocytes to confer the
induction of inducible nitric oxide synthase gene expression.
Shock. 26:162–168. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Braet F, Nagatsuma K, Saito M, Soon L,
Wisse E and Matsuura T: The hepatic sinusoidal endothelial lining
and colorectal liver metastases. World J Gastroenterol. 13:821–825.
2007. View Article : Google Scholar : PubMed/NCBI
|