Introduction
The incidence and mortality rates of colorectal
cancer (CRC) vary markedly worldwide, due to heterogeneity in
environmental and genetic factors. A high-fat and low-fiber diet
has been established as a major risk factor for colon cancer
development (1), and cholecystectomy
has been associated with right-sided colon cancer (2). These factors may result in an increased
bile acid (BA) load in the intestine and higher relative
concentrations of fecal secondary Bas (3). Consequently, resistance to BA-induced
DNA oxidative damage, inflammation, nuclear factor-κB activation
and enhanced cell proliferation has been reported to increase colon
tumorigenesis (4).
BAs are endogenous ligands for the nuclear farnesoid
X receptor (FXR; gene name, Nuclear Receptor Subfamily 1 Group H
Member 4), which regulates BA concentrations by modulating BA
influx, efflux and detoxification (5,6). FXR is
downregulated in both mouse and human models of CRC during the
progression from normal intestinal epithelia to dysplastic lesions,
and its expression has been inversely correlated with CRC stage and
clinical outcome (7,8). FXR silencing in chronic colitis mouse
models of intestinal tumorigenesis results in early mortality and
increased tumor progression (9).
The mechanism by which FXR suppresses tumor growth
remains unclear, but it may involve protecting the colonic
epithelium from inflammation and ameliorating BA toxicity by
upregulating intracellular BA binding proteins and efflux
transporters and downregulating influx transporters and de
novo BA synthesis (10–12). However, FXR has additional
antitumorigenic functions independent of BA homeostasis regulation
(13). FXR deficiency increases
colon cancer susceptibility by increasing epithelial permeability
to bacteria, promoting Wnt/β-catenin signaling and increasing
intestinal inflammation (8). The aim
of the present study was to explore the effects and mechanism of
FXR on the Wnt/β-catenin signaling pathway in CRC.
Materials and methods
Cell culture
The human colon cancer cell lines HT-29, Caco-2 and
HCT-116 were obtained from the State Key Laboratory of Molecular
Oncology, National Cancer Center/Cancer Hospital, Chinese Academy
of Medical Sciences (cell line not authenticated). HT-29, Caco-2
and HCT-116 cells were maintained in Dulbecco's modified Eagle's
medium: Nutrient mixture F-12 media (DMEM/F-12; HyClone; GE
Healthcare Life Sciences), minimum essential medium/Earle's
balanced salt solution (MEM/EBSS; HyClone; GE Healthcare Life
Sciences) and Iscove's modified Dulbecco's medium (IMDM; HyClone;
GE Healthcare Life Sciences), respectively. They were supplemented
with 10% fetal bovine serum (FBS; HyClone; GE Healthcare Life
Sciences) and 1% penicillin/streptomycin (HyClone; GE Healthcare
Life Sciences) at 37°C in a humidified incubator containing 5%
CO2. Caco-2 cells were co-treated with 1% non-essential
amino acids (HyClone; GE Healthcare Life Sciences).
Plasmid and luciferase reporter
assays
FXR siRNA (sc-38848) and non-targeting negative
control siRNA (siNC) (sc-37007) were purchased from Santa Cruz
Biotechnology, Inc. and transiently transfected into cells using
Lipofectamine® 2000 according to the manufacturer's
protocol (Invitrogen; Thermo Fisher Scientific, Inc.). The
sequences of these siRNAs was not available from the manufacturer.
A preliminary experiment was conducted to and indicated 40 nM as
the optimal dose, and this was used for the subsequent experiments
(data not shown). At 48 h after cells were transfected with 40 nM
siFXR or siNC, total protein was extracted and FXR was measured via
western blotting. According to the physiological function of FXR in
regulating BA metabolism, FXR activation primes the transcription
of the small heterodimer partner (SHP), ultimately resulting in
decreased expression of CYP7A1, the rate-limiting enzyme of BA
de novo synthesis (14).
Therefore, the target gene of FXR, SHP/NR0B2 was also measured to
verify its successful knockdown.
E. coli DH5α, TOPflash and pRL-TK vectors
were obtained from the State Key Laboratory of Molecular Oncology,
National Cancer Center/Cancer Hospital, Chinese Academy of Medical
Sciences. Luciferase assays were performed using the
Dual-Luciferase Reporter assay system (Invitrogen; Thermo Fisher
Scientific, Inc.). Briefly, 300 ng of TOPflash and 30 ng of pRL-TK
with siFXR or siNC were co-transfected into cells using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions.
Luciferase activity in the harvested cell lysates was measured
using a luminometer, 48 h after transfection. The results were
normalized to Renilla luciferase activity.
Electrophoretic mobility shift assay
(EMSA)
Nuclear extracts from cultured cells were prepared
according to the manufacturer's protocol with Nuclear and
Cytoplasmic Extraction reagents (Pierce; Thermo Fisher Scientific,
Inc.). The biotin-labelled Bio-TCF-4 probe
(Bio-5′-CCCTTTGATCTTACC-3′) and cold-TCF-4 probe
(cold-5′-AGTTGAGGGGACTTTCCCAGGC-3′) (15,16),
were constructed by Sango Biotech Co., Ltd. EMSAs were performed by
incubating 5 µg nuclear extract with 0.5 µl biotin-labelled
Bio-TCF-4 DNA probes in a 15-µl binding reaction mixture containing
1.5 µl 10X binding buffer and 1 µl poly(dI-dC)/poly(dI-dC)
(LightShift™ Poly(dI-dC), cat. no. 20148E, Pierce;
Thermo Fisher Scientific, Inc.). Following incubation at room
temperature for 20 min, the samples were loaded onto a 6.5%
polyacrylamide gel and electrophoresed in Tris/Borate/EDTA (TBE)
buffer at 180 V for 60 min at 4°C. For competition experiments, the
nuclear extracts were preincubated with 2 µl unlabeled
oligonucleotides before the biotin-labeled oligonucleotide DNA
probe was added. The complexes were transferred onto nitrocellulose
in TBE buffer for 40 min at 390 mA at room temperature.
Immunoreactive bands were developed using streptavidin-horseradish
peroxidase (HRP) and visualized via autoradiography. CK represents
the background control group with only the probe and no nuclear
extract; 100× represents the competitive control group with
additional non-labelled probe; + represents the positive control
group [nuclear extracts from cells pre-treated with Wnt signaling
activator BML284 (Abcam)] (17); C
represents the control group pre-treated with siNC. Intensity
analysis was performed to demonstrate the differences between
groups, for which intensity of the positive control group was
recorded as 1 and results of other groups were calculated as the
intensity ratio relative to the positive control group. Analysis
was performed using ImageJ (version 1.52q; National Institutes of
Health).
Western blotting
Cultured cells were lysed in 500 µl RIPA lysis
buffer (Applygen Technologies, Inc.), and total protein
concentration was determined using a BCA protein assay kit
(Applygen Technologies, Inc.). Equal amounts of protein (50
µg/lane) were electrophoresed using 10% SDS-PAGE gel and then
transferred onto nitrocellulose membranes. After blocking with TTBS
containing 5% non-fat milk and 0.1% Tween 20 for 2 h at room
temperature, the membranes were incubated overnight with a primary
antibody at 4°C. After washing with TTBS containing 0.1% Tween-20 3
times for 5 min each, the membranes were incubated with a
HRP-labeled secondary antibody for another 2 h at room temperature.
The membranes were then developed using Pierce ECL Western Blotting
Substrate (Thermo Fisher Scientific, Inc.) and exposed using an
Amersham Imager 600 (GE Healthcare). Primary anti-FXR mouse
monoclonal antibody (1:100,000, cat. no. ab187735, Abcam),
anti-β-catenin rabbit monoclonal antibody (1:100,000, cat. no.
ab32572, Abcam), anti-β-actin mouse monoclonal antibody (1:10,000,
cat. no. sc-8432, Santa Cruz Biotechnology, Inc.), anti-Poly
(ADP-ribose) polymerase (PARP) rabbit polyclonal antibody (1:1,000,
cat. no. #9542,) and secondary anti-mouse (1:10,000, cat. no.
#7076), anti-rabbit (1:10,000, cat. no. #7074) horseradish
peroxidase-conjugated antibodies (all Cell Signaling Technology,
Inc.) were used for protein labeling. β-actin was used here as a
loading control of total protein extract, and PARP was used as a
loading control of nuclear protein extract (18).
RNA extraction and reverse
transcription-quantitative (RT-q)PCR analysis
RNA was isolated from cells using TRIzol®
reagent according to the manufacturer's protocol (Invitrogen;
Thermo Fisher Scientific, Inc.). cDNA was generated from 500 ng
total RNA using SuperScript II Reverse Transcriptase (Thermo Fisher
Scientific, Inc.). RT-qPCR reactions were performed as follows:
25°C for 10 min, 50°C for 15 min and 85°C for 5 min. qPCR analysis
was performed using SYBR Green PCR master mix (Thermo Fisher
Scientific, Inc.) and analyzed using a CFX96 real-time PCR system
(Bio-Rad Laboratories, Inc.). The reaction conditions included an
initial pre-denaturation step at 95°C for 30 sec, followed by 40
cycles of thermal steps consisting of 95°C for 5 sec and 60°C for
30 sec. Values were normalized to GADPH. The fold-change was
calculated by the 2−ΔΔCq method (19). The primer sequences were as follows:
β-catenin (human) forward, 5′-AGAACCCCTTGGATATCGCC-3′ and reverse,
5′-TGGCCACCCATCTCATGTTC-3′; and GAPDH (human) forward,
5′-CTCTGCTCCTCCTGTTCGAC-3′ and reverse,
5′-GCGCCCAATACGACCAAATC-3′.
Cell proliferation assay
The human colon cancer cell lines HT-29, Caco-2 and
HCT-116 (5×104 cells) were seeded in quintuplicate in
96-well plates and incubated at 37°C, 95% humidity, and 5% carbon
dioxide overnight. At the indicated time points, 40 µl MTT (Sangon
Biotech Co., Ltd.) solution (1 mg/ml) was added to each well. The
plates were incubated at 37°C for 1 h. Subsequently, the medium was
removed, and 200 µl DMSO was added to each well to dissolve the
formazan crystals. Next, the plates were measured at 490 nm on a
multi-well plate reader. The effect of the FXR agonist GW4064
(Sigma-Aldrich; Merck KGaA) on cell proliferation was measured.
Based on the present studies on GW4064 (20,21), the
experimental groups in the present study included: i) Different
doses of GW4064 (1, 3 and 5 µM); and ii) different durations of
GW4064 treatment (12, 24, 36 and 48 h). Control groups were treated
with only 1, 3 or 5 µM GW4064 according to the dose, and other
conditions were consistent with the experimental groups.
Mice
Eight-week-old male C57BL/6 mice (21±2 g) were
purchased from Vital River Laboratory Animal Technology Co. Ltd.
All animals were housed under specific pathogen-free conditions in
plastic cages (4–5 mice/cage) with free access to drinking water
and a pellet-based diet, under controlled conditions of humidity
(40–60%), light (12/12 h, light/dark cycle) and temperature
(22±1°C) in the National Cancer Center/Cancer Hospital animal
facilities. Experiments were performed in accordance with local
laws and the Council of the European Communities Directive of 1986
(86/609/EEC) guidelines for the care and use of laboratory animals.
All animal experiments were approved by and conducted in accordance
with the recommendations of the Animal Care Ethics and Use
Committee of Peking Union Medical College (Beijing, China; approval
no. XHDW-2015-0032). All measures were made to minimize animal
suffering and to reduce the number of animals used.
All mice (n=30) were initially housed together (5
animals/cage) for adaption, one week before randomization into the
control group [no ulcerative colitis (UC)-associated carcinogenesis
induction, n=10) and model group (UC-associated carcinogenesis,
n=20). For the UC carcinogenesis model, the mice were injected
intraperitoneally with 12.5 mg/kg body weight azoxymethane (AOM).
After one week, 2.5% dextran sulfate sodium (DSS) (MP Biomedicals,
LLC) was added to the drinking water for 5 days, followed by 10
weeks and 2 days of regular drinking water. This modeling method
was based on a novel inflammation-related mouse colon
carcinogenesis model (22),
indicating that 1-week administration of DSS after initiation with
a low dose of AOM exerted a powerful tumor-promoting activity in
colon carcinogenesis. Body weight, colitis symptoms and other
animal behavior were recorded every week throughout the experiment.
At the 12th week, all mice were sacrificed via injection of 200
mg/kg pentobarbital sodium to induce overdose anesthesia, and colon
tissues were removed.
Histology and
immunohistochemistry
Tissue specimens were fixed in 10% formalin at room
temperature for 12–24 h, dehydrated and paraffin embedded. Graded
ethanol was used in a dehydration series as follows: 70% for 2 h,
80% for 2 h, 90% for 2 h, 95% for 2 h, an two lots of 100% for 2 h.
The sections were deparaffinized in 5% xylene and then dehydrated
in ethanol as following: 100% for 1 min, 95% for 1 min and 75% for
1 min. It was then washed with distilled water and stained with
Harris hematoxylin at room temperature for 3 min and rinsed in
distilled water. The section was differentiated with 1% acid
alcohol, washed under running water, and counterstained with eosin
at room temperature for 1 min. At last, it was washed with water,
dehydrated with ascending grades of ethanol, 95% for 1 min, two
lots of 100% for 1 min, and cleaned with xylene. The mounted slides
were then examined and images were captured with magnifications of
×4, ×10 and ×20 using a light microscope (Olympus).
Standard immunohistochemical procedures were
performed. Briefly, 5-µm-thick sections were treated with 3%
hydrogen peroxide at room temperature for 5 min to quench
endogenous peroxidase activity and subjected to antigen retrieval
by boiling the slides in an antigen unmasking solution (Zsbio
Commerce Store) for 15 min, according to the manufacturer's
instructions. The sections were incubated sequentially for 60 min
at room temperature in 50% non-immune serum (from the host animal
in which the secondary antibody was raised) in PBS (to avoid
non-specific signaling) and overnight at 4°C with the following
primary antibodies: Anti-FXR rabbit polyclonal antibody (1:300,
cat. no. sc-13063, Santa Cruz Biotechnology, Inc.) and
anti-β-catenin rabbit monoclonal antibody (1:2,000, cat. no.
ab32572, Abcam). The sections were washed for 10 min with PBS and
incubated for 30 min at room temperature with the secondary
biotinylated antibody (1:1, cat. no. PV-9000, Zsbio Commerce
Store). After several washes with PBS (3 washes; 5 min each), the
sections were incubated with the avidin-biotin complex (Cell
Signaling Technology, Inc.) for 30 min at room temperature. After
washing with PBS, the peroxidase reaction was developed by
incubation with diaminobenzidine (Sigma-Aldrich; Merck KGaA) 8 min
at room temperature. The staining was observed under a microscope,
when the brown color appeared, all sections were immediately washed
three times for 5 min with PBS to terminate the reaction
simultaneously. Counterstaining was carried out with methylene-blue
(Sigma-Aldrich; Merck KGaA). For negative controls, 1% non-immune
serum in PBS was used instead of the primary antibodies. The
positively stained cells were counted in randomized fields at ×400
magnification using a light microscope (Olympus).
FXR and β-catenin expression was qualitatively
assessed through scored by two independent, experienced
pathologists who were blinded to the clinicopathological
information. Staining intensity was scored as (absent, 0), (weak,
1), (moderate, 2) and (strong, 3). The percentages of
immunoreactive cells were grouped and scored manually as follows:
1, <25%; 2, 25–50%; 3, 50–75%; and 4, >75%. A final score was
obtained by multiplication of the single scores using. There was
~95% concordance between the two observers. Differences were
resolved by consensus evaluation of the slides.
Statistical analysis
The results are expressed as the mean ± SEM, as
indicated in the figure legends. Statistical significance was
determined by two-tailed independent Student's t-test or one-way
ANOVA and Tukey's post hoc test. All statistical calculations were
performed using the SPSS 22.0 software (IBM Corp.). P<0.05 was
considered to indicate a statistically significant difference.
Results
FXR knockdown activates the Wnt
signaling pathway
FXR and β-catenin protein expression levels were
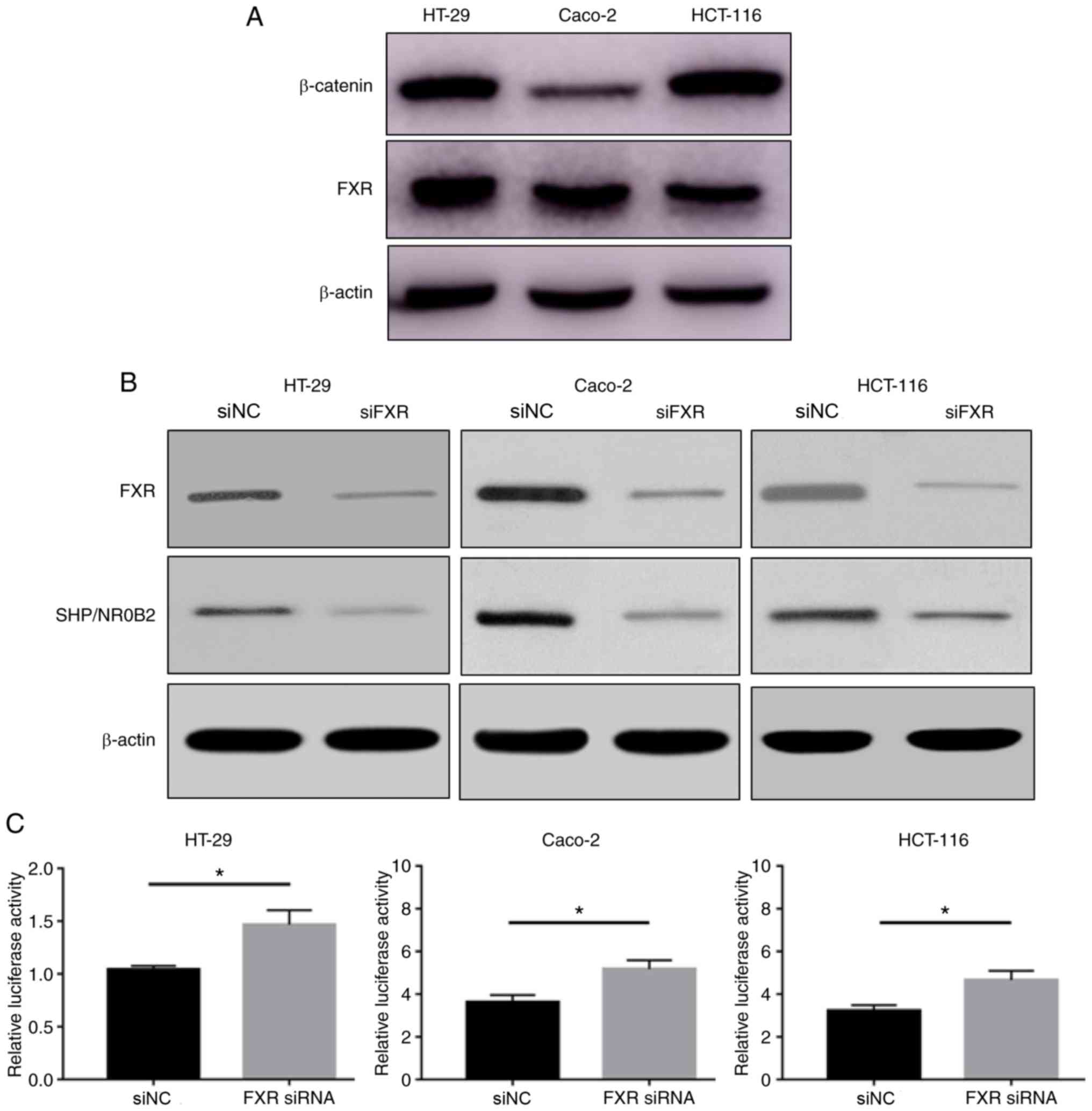
determined in the HT-29, Caco-2 and HCT-116 cell lines. HT-29 and
Caco-2 cells highly express FXR, and HT-29 and HCT-116 cells highly
express β-catenin (Fig. 1A). FXR and
β-catenin expression levels were different between these human
colon cancer cell lines.
siFXR efficiently silenced FXR expression and its
target gene SHP/NR0B2 (Fig. 1B). The
effect of FXR on β-catenin-mediated transcriptional activity was
subsequently tested using a cell-based TOPflash reporter assay. The
TOPflash reporter, pRL-TK and siFXR or siNC were co-transfected
into HT-29, Caco-2 and HCT-116 cells. Silencing FXR significantly
increased the β-catenin-mediated luciferase activity, driven by the
TOPflash reporter, compared with negative control conditions
(Fig. 1C). These results suggested
that FXR knockdown activates Wnt/β-catenin transcriptional
activity.
To explore the mechanism by which FXR interacts with
the Wnt signaling pathway, the β-catenin/TCF4 complex levels were
determined. Because the transcriptional activity of the Wnt
signaling pathway is mediated by the β-catenin/TCF4 complex, EMSA
was used to measure the active β-catenin/TCF4 complex levels. The
silencing of FXR enhanced the formation of the β-catenin/TCF4
complex (P<0.05, Fig. 2A).
However, the β-catenin protein distribution in the cytoplasm and
nuclei was not different following FXR silencing (Fig. 2B). In addition, no difference was
observed in β-catenin mRNA levels (Fig.
2C).
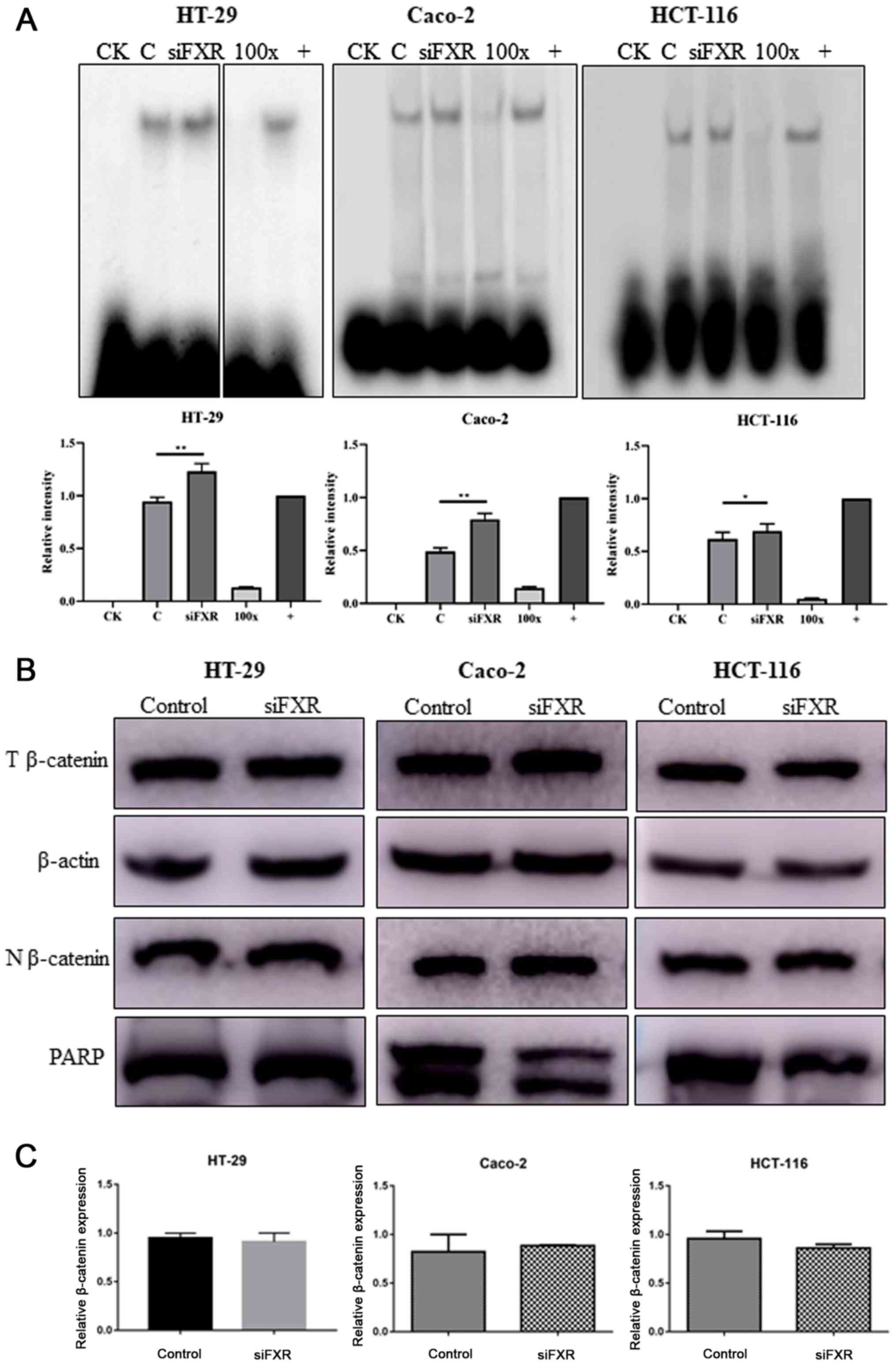 | Figure 2.FXR activates the Wnt signaling
pathway via increasing β-catenin/TCF4 complex levels. (A) Silencing
FXR increases β-catenin/TCF4 complex formation as detected by an
electrophoretic mobility shift assay. Images of HT-29 cells were
from discontinuous parts of the same gel. Intensity analysis using
ImageJ (version 1.52q; National Institute of Health) is also
revealed to demonstrate the difference among groups. (B) T protein
and N protein were extracted separately to determine the expression
and distribution of β-catenin after FXR silencing. (C) β-catenin
mRNA levels were not different after FXR silencing. PARP, Poly
(ADP-ribose) polymerase, which is expressed in only the nucleus,
was used as a loading control. The error bars represent the SEM of
three independent samples. FXR, farnesoid X receptor; CK,
background control; 100×, competitive control; +, positive control;
C, control group pre-treated with negative control siRNA; T, total;
N, nuclear; si, small interfering; NC, negative control.
*P<0.05; **P<0.01. |
The current results indicated that silencing FXR
activates the Wnt signaling pathway mediated by an increase in
β-catenin/TCF4 complex levels, with no changes in the β-catenin
mRNA and protein expression levels.
Effect of an FXR agonist on the Wnt
signaling pathway
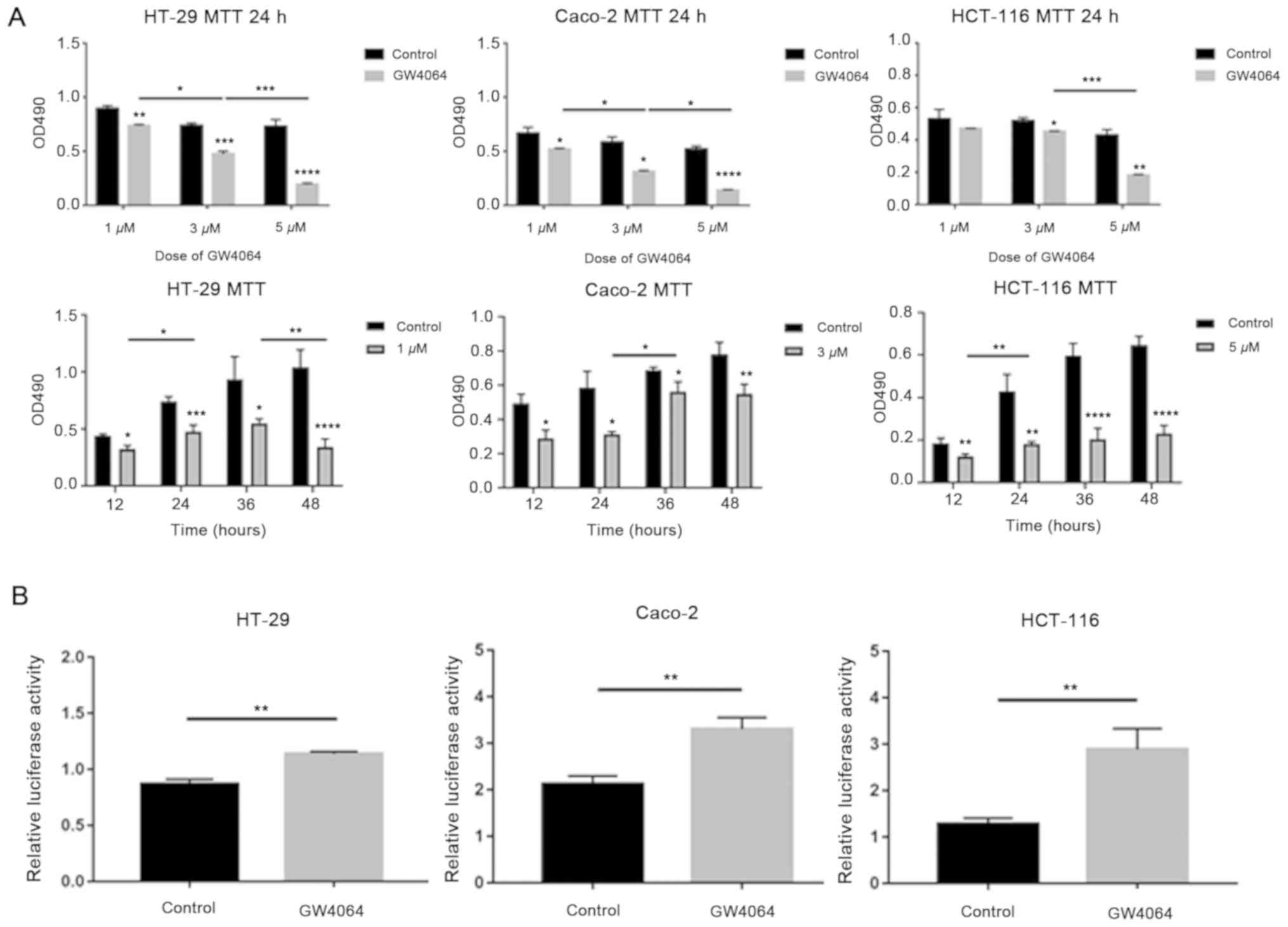
To confirm the appropriate dose and duration time of
the FXR agonist GW4064, cell proliferation was measured using MTT
assays. GW4064 significantly inhibited cell proliferation in a
dose- and time-dependent manner compared with the solvent control,
(P<0.05, Fig. 3A). In HT-29 and
Caco-2 cells with high FXR expression, 3 µM GW4064 was used in the
following experiments, and in HCT-116 cells with low FXR
expression, 5 µM GW4064 was used.
Subsequently, the effect of GW4064 on
β-catenin-mediated transcriptional activity was tested. Cells were
treated with GW4064 following transfection with TOPflash and
pRL-TK. GW4064 promoted β-catenin-mediated luciferase activity
compared with the solvent control (Fig.
3B).
FXR and β-catenin expression in a UC
carcinogenesis mouse model
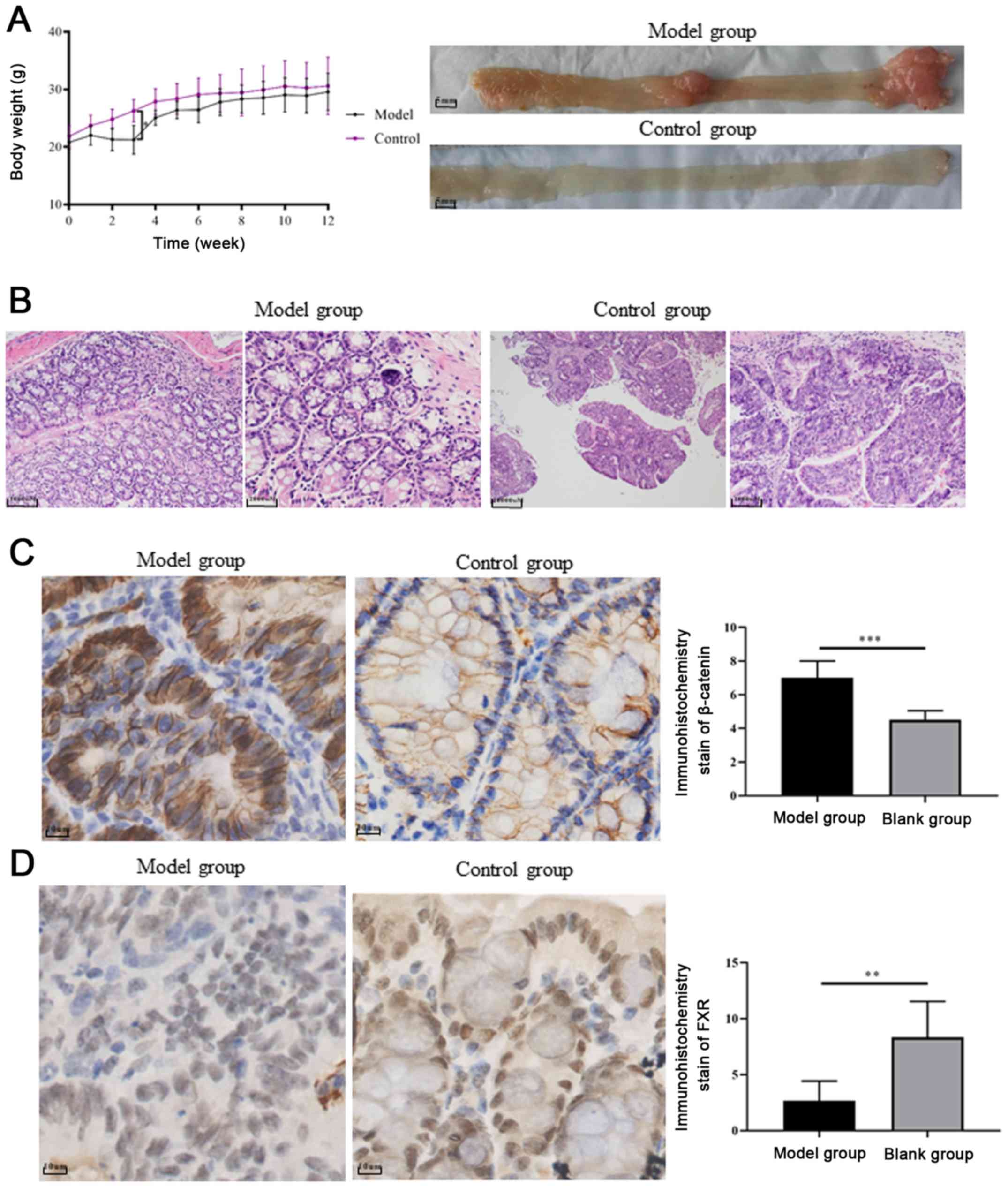
Mice treated with AOM/DSS exhibited significant body
weight loss compared with control mice, following day 10 of
administration (P=0.0001, Fig. 4A).
Colitis symptoms, such as loose and bloody stool, dull body hair,
fatigue and decreased movement were accompanied, which were
alleviated when the mice received ordinary drinking water. In weeks
9–10, two of the mice treated with AOM/DSS exhibited bloody stools
again as well as anal prolapse, anal tumor fusion and ring growth
at the end of the rectum (Fig. 4A).
However, no apparent weight loss was observed among the groups at
the end of the 12th week. Mice treated with AOM/DSS exhibited
mucosal carcinoma or high-grade intraepithelial neoplasia,
manifested as colonic gland structure disorder, large nuclei, deep
staining and nucleoplasmic ratio imbalance (Fig. 4B).
In order to explore whether FXR and β-catenin
activation influences chronic colitis-associated colorectal
carcinoma development, the expression levels of FXR and β-catenin
were evaluated in tissue samples from the UC carcinogenesis mouse
model and control groups. The histopathological grades of FXR
expression based on IHC staining scores were significantly lower in
the model group compared with the control group (n=6; 2.667±0.7149
vs. 8.333±1.308, respectively; P<0.01; Fig. 4D). By contrast, β-catenin
upregulation was detected in the cell nucleus of tumor samples
(Fig. 4C).
Discussion
Wnt/β-catenin signaling is highly conserved from
nematodes to humans, and is essential for various cellular
processes, such as development, growth, survival, regeneration and
self-renewal (23). Mutations in the
Wnt/β-catenin signaling pathway have been detected in >90% of
colon carcinogenesis cases (24). A
close association was recently revealed between FXR and
Wnt/β-catenin signaling (25–27). FXR
knockdown in a chronic colitis mouse model resulted in greater
susceptibility to tumorigenesis and increased tumor progression
(28). Correspondingly, increased
expression of β-catenin and its downstream targets C-Myc and cyclin
D1 was detected in FXR-deficient mice (28). Furthermore, FXR overexpression
resulted in tumor growth suppression in both in FXR deficiency mice
and colon cancer cell in vitro and in vivo (13).
In the present study, the crosstalk and mechanism
between FXR and the Wnt signaling pathway was identified. FXR
expression was significantly decreased and was inversely associated
with β-catenin in both the mouse model and colon cancer cell lines.
Consistent with the observations of the present study, FXR
expression was also downregulated in mouse models and human CRC
samples in previous studies (7,8). In
addition, a similar phenomenon was observed in cholangiocarcinoma,
biliary tract carcinoma and tumor samples from patients with
hepatocellular carcinoma (20),
which may indicate the participation of BAs and FXR in the
mechanism of these types of tumorigenesis. The results suggested
that the interaction between FXR and β-catenin is a critical
component in this process (20).
Since previous studies have demonstrated a correlation between FXR
deficiency and increased expression of β-catenin and its downstream
targets C-Myc and cyclin D1, other stains that are associated with
the development or growth of tumors have not been repeated.
In the present study, FXR knockdown significantly
promoted β-catenin transcriptional activity. FXR appeared to
regulate β-catenin activity via increasing β-catenin/TCF4 complex
levels, without affecting β-catenin localization or total mRNA and
protein levels. Activated β-catenin forms a complex with TCF4 and
is recruited to the corresponding promoter region of Wnt target
genes to elicit transcriptional activity (29). The results also suggest that FXR
activation by GW4064 results in significantly inhibiting cell
proliferation, and that GW4064 promoted β-catenin-mediated
luciferase activity.
The present study demonstrated that the silencing of
FXR by siRNA-FXR, and FXR activation by GW4084 treatment, both
resulted in Wnt/β-catenin signaling activation. This suggests that
the interaction between FXR and the Wnt signaling pathway warrants
further study. Consistent with the observations of the present
study, previous studies have reported that FXR deficiency results
in increased β-catenin transcriptional activity in chronic colitis
mouse models (13,28). However, few studies have focused on
whether the effect of β-catenin occurs via FXR activation or
upregulation. Only one study in the 293 cell line, which originates
from renal epithelial cells, indicated that both GW4064-induced
activation and FXR overexpression decreased β-catenin
transcriptional activity (20).
Since mutations in the Wnt signaling pathway have been detected in
most colorectal cell lines, the effects of abnormal interaction
sites may be relevant to the present study.
GW4064, a potent agonist of FXR, was used in the
present study to explore the interaction between FXR and the Wnt
signaling pathway. GW4064 is a synthetic isoxazole, which was used
to decipher the cellular and physiological functions of FXR.
Similar to previous studies (20,21), the
selected dose of GW4064 was ~–5 µM. In a study by Peng et al
(21), the effect of GW4064 at 0.1,
0.5, 1, 5 and 10 µM on cell proliferation was demonstrated in three
colon cancer cell lines (H508, SNU-C4 and HT-29), and the results
indicated that the effect was significant when the dose of GW4064
reaching ~1 µM. However, in the physiological state, FXR is a
member of the nuclear family and serves as a regulatory center of
BAs. The complexity of the components of the BA pool and the
crosstalk of nuclear family members may all contribute to its
function (6,7,14).
The crosstalk between Wnt/β-catenin ligands and
members of the nuclear receptor (NR) family has been considered a
clinically and developmentally important research area of cancer
biology (30). The NR family
consists of retinoid X receptor (RXR), steroid receptor, peroxisome
proliferator-activated receptor-γ and vitamin D receptor (VDR). A
high proportion of these proteins interact with the Wnt pathway via
different mechanisms; for example, RXR binds β-catenin to degrade
and decrease the levels of the β-catenin/TCF complex (31,32). VDR
activation increases VDR/β-catenin complex formation, in addition
to enhancing E-cadherin expression and sequestering TCF-bound
β-catenin (33). The identification
of functional interactions between Wnt signaling components and NRs
is gaining increasing attention. A growing number of NRs appear to
be activated by β-catenin, resulting in alterations in cell
proliferation and tumorigenesis, whereas Wnt signaling appears to
be compromised by the actions of NRs (30).
Exploration of the interaction of FXR with the
Wnt/β-catenin signaling pathway in the present study demonstrated
that FXR knockdown promotes β-catenin/TCF4 complex formation and,
subsequently, its binding ability to the corresponding promoter.
The data indicate a novel mechanism through which FXR expression is
mediated during tumor progression. Thus, FXR represents a novel
modulator of the Wnt signaling pathway, and a potential molecular
target of the Wnt signaling cascade that may be exploited to
achieve antitumor effects.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (grant nos. 81370500
and 81770559).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JL designed the study. JM and XC performed the cell
experiments, and CW and WL performed the animal experiments. WL
helped interpret the data. JM and XC analyzed the data. JM wrote
the manuscript. CW revised the manuscript for important
intellectual content. All authors read and approved the final
manuscript.
Ethics approval and consent for
participate
All animal experiments were approved by and
conducted in accordance with the recommendations of the Animal Care
Ethics and Use Committee of Peking Union Medical College (approval
no. XHDW-2015-0032).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
FXR
|
farnesoid X receptor
|
|
BA
|
bile acid
|
|
CRC
|
colorectal carcinoma
|
|
UC
|
ulcerative colitis
|
|
EMSA
|
electrophoretic mobility shift
assay
|
|
NR
|
nuclear receptor
|
|
VDR
|
vitamin D receptor
|
References
|
1
|
Reddy BS: Diet and excretion of bile
acids. Cancer Res. 41:3766–3768. 1981.PubMed/NCBI
|
|
2
|
Lagergren J, Ye W and Ekbom A: Intestinal
cancer after cholecystectomy: Is bile involved in carcinogenesis?
Gastroenterology. 121:542–547. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Peterlik M: Role of bile acid secretion in
human colorectal cancer. Wien Med Wochenschr. 158:539–541. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pearson JR, Gill CI and Rowland IR: Diet,
fecal water, and colon cancer-development of a biomarker. Nutr Rev.
67:509–526. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kundu S, Kumar S and Bajaj A: Cross-talk
between bile acids and gastrointestinal tract for progression and
development of cancer and its therapeutic implications. IUBMB Life.
67:514–523. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Degirolamo C, Modica S, Palasciano G and
Moschetta A: Bile acids and colon cancer: Solving the puzzle with
nuclear receptors. Trends Mol Med. 17:564–572. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Modica S, Gofflot F, Murzilli S, D'Orazio
A, Salvatore L, Pellegrini F, Nicolucci A, Tognoni G, Copetti M,
Valanzano R, et al: The intestinal nuclear receptor signature with
epithelial localization patterns and expression modulation in
tumors. Gastroenterology. 138:636–648, 648.e1-12. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bailey AM, Zhan L, Maru D, Shureiqi I,
Pickering CR, Kiriakova G, Izzo J, He N, Wei C, Baladandayuthapani
V, et al: FXR silencing in human colon cancer by DNA methylation
and KRAS signaling. Am J Physiol Gastrointest Liver Physiol.
306:G48–G58. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lax S, Schauer G, Prein K, Kapitan M,
Silbert D, Berghold A, Berger A and Trauner M: Expression of the
nuclear bile acid receptor/farnesoid X receptor is reduced in human
colon carcinoma compared to nonneoplastic mucosa independent from
site and may be associated with adverse prognosis. Int J Cancer.
130:2232–2239. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Frankenberg T, Rao A, Chen F, Haywood J,
Shneider BL and Dawson PA: Regulation of the mouse organic solute
transporter alpha-beta, Ostalpha-Ostbeta, by bile acids. Am J
Physiol Gastrointest Liver Physiol. 290:G912–G922. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
De Gottardi A, Touri F, Maurer CA, Perez
A, Maurhofer O, Ventre G, Bentzen CL, Niesor EJ and Dufour JF: The
bile acid nuclear receptor FXR and the bile acid binding protein
IBABP are differently expressed in colon cancer. Dig Dis Sci.
49:982–989. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gadaleta RM, van Erpecum KJ, Oldenburg B,
Willemsen EC, Renooij W, Murzilli S, Klomp LW, Siersema PD,
Schipper ME, Danese S, et al: Farnesoid X receptor activation
inhibits inflammation and preserves the intestinal barrier in
inflammatory bowel disease. Gut. 60:463–472. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Modica S, Murzilli S, Salvatore L, Schmidt
DR and Moschetta A: Nuclear bile acid receptor FXR protects against
intestinal tumorigenesis. Cancer Res. 68:9589–9594. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gadaleta RM, Garcia-Irigoyen O and
Moschetta A: Bile acids and colon cancer: Is FXR the solution of
the conundrum? Mol Aspects Med. 56:66–74. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Korinek V, Barker N, Morin PJ, van Wichen
D, de Weger R, Kinzler KW, Vogelstein B and Clevers H: Constitutive
transcriptional activation by a beta-catenin-Tcf complex in
APC-/-colon carcinoma. Science. 275:1784–1787. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mizushima T, Nakagawa H, Kamberov YG,
Wilder EL, Klein PS and Rustgi AK: Wnt-1 but not epidermal growth
factor induces beta-catenin/T-cell factor-dependent transcription
in esophageal cancer cells. Cancer Res. 62:277–282. 2002.PubMed/NCBI
|
|
17
|
Ni D, Liu J, Hu Y, Liu Y, Gu Y, Zhou Q and
Xie Y: A1CF-Axin2 signal axis regulates apoptosis and migration in
Wilms tumor-derived cells through Wnt/β-catenin pathway. Vitro Cell
Dev Biol Anim. 55:252–259. 2019. View Article : Google Scholar
|
|
18
|
Xin Y, He L, Luan Z, Lv H, Yang H, Zhou Y,
Zhao X, Zhou W, Yu S, Tan B, et al: E-cadherin mediates the
preventive effect of Vitamin D3 in Colitis-associated
Carcinogenesis. Inflamm Bowel Dis. 23:1535–1543. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu X, Zhang X, Ji L, Gu J, Zhou M and
Chen S: Farnesoid X receptor associates with β-catenin and inhibits
its activity in hepatocellular carcinoma. Oncotarget. 6:4226–4238.
2015.PubMed/NCBI
|
|
21
|
Peng Z, Raufman JP and Xie G: Src-mediated
cross-talk between farnesoid X and epidermal growth factor
receptors inhibits human intestinal cell proliferation and
tumorigenesis. PLoS One. 7:e484612012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tanaka T, Kohno H, Suzuki R, Yamada Y,
Sugie S and Mori H: A novel inflammation-related mouse colon
carcinogenesis model induced by azoxymethane and dextran sodium
sulfate. Cancer Sci. 94:965–973. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cavard C, Colnot S, Audard V, Benhamouche
S, Finzi L, Torre C, Grimber G, Godard C, Terris B and Perret C:
Wnt/beta-catenin pathway in hepatocellular carcinoma pathogenesis
and liver physiology. Future Oncol. 4:647–660. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cancer Genome Atlas Network, .
Comprehensive molecular characterization of human colon and rectal
cancer. Nature. 487:330–337. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fu T, Coulter S, Yoshihara E, Oh TG, Fang
S, Cayabyab F, Zhu Q, Zhang T, Leblanc M, Liu S, et al: FXR
regulates intestinal cancer stem cell proliferation. Cell.
176:1098–1112.e18. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Thompson MD, Moghe A, Cornuet P, Marino R,
Tian J, Wang P, Ma X, Abrams M, Locker J, Monga SP and Nejak-Bowen
K: β-Catenin regulation of farnesoid X receptor signaling and bile
acid metabolism during murine cholestasis. Hepatology. 67:955–971.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang R, Nakao T, Luo J, Xue Y, Cornuet P,
Oertel M, Kosar K, Singh S and Nejak-Bowen K: Activation of
WNT/Beta-catenin signaling and regulation of the Farnesoid X
Receptor/Beta-catenin complex after murine bile duct ligation.
Hepatol Communications. 3:1642–1655. 2019. View Article : Google Scholar
|
|
28
|
Maran RR, Thomas A, Roth M, Sheng Z,
Esterly N, Pinson D, Gao X, Zhang Y, Ganapathy V, Gonzalez FJ and
Guo GL: Farnesoid X receptor deficiency in mice leads to increased
intestinal epithelial cell proliferation and tumor development. J
Pharmacol Exp Ther. 328:469–477. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sawa M, Masuda M and Yamada T: Targeting
the Wnt signaling pathway in colorectal cancer. Expert Opin Ther
Targets. 20:419–429. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mulholland DJ, Dedhar S, Coetzee GA and
Nelson CC: Interaction of nuclear receptors with the
Wnt/β-catenin/Tcf signaling axis: Wnt you like to know? Endocr Rev.
26:898–915. 2006. View Article : Google Scholar
|
|
31
|
Xiao JH, Ghosn C, Hinchman C, Forbes C,
Wang J, Snider N, Cordrey A, Zhao Y and Chandraratna RA:
Adenomatous polyposis coli (APC)-independent regulation of
beta-catenin degradation via a retinoid X receptor-mediated
pathway. J Biol Chem. 278:29954–29962. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Han A, Tong C, Hu D, Bi X and Yang W: A
direct protein-protein interaction is involved in the suppression
of beta-catenin transcription by retinoid X receptor alpha in
colorectal cancer cells. Cancer Biol Ther. 7:454–459. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Palmer HG, González-Sancho JM, Espada J,
Berciano MT, Puig I, Baulida J, Quintanilla M, Cano A, de Herreros
AG, Lafarga M and Muñoz A: Vitamin D(3) promotes the
differentiation of colon carcinoma cells by the induction of
E-cadherin and the inhibition of beta-catenin signaling. J Cell
Biol. 154:369–387. 2001. View Article : Google Scholar : PubMed/NCBI
|