Introduction
Myelodysplastic syndromes (MDS) are defined as
clonal hematopoietic stem cell (HSC) malignancies with
characteristic manifestation of peripheral blood cytopenias due to
ineffective hematopoiesis and dysplasia of the bone marrow (BM).
MDS has a high risk of conversion to acute myeloid leukemia (AML)
(1). Treatments for patients with
MDS include hematopoietic therapy, differentiation therapy,
demethylation therapy and allogeneic BM HSC transplantation
(allo-HSCT) (2). However, the
efficacy of these treatments is unsatisfactory. Data from the
Center for International Blood and Marrow Transplant Research
(CIBMTR) indicated transplant-related mortality (TRM) and relapse
rates of 30 and 30%, respectively, following allogeneic HCT from
human leukocyte antigen (HLA)-matched related donors; the 3-year
probability of survival is 40% (3).
Upon progression to AML, patients exhibit a poor response to
standard therapy and a high likelihood of mortality. Therefore, it
is urgently necessary to identify effective therapeutic targets to
prevent MDS progression.
Sperm-associated antigen 6 (SPAG6), a member of the
cancer-testis antigen family, was first discovered in human testis
tissue, where its functions include regulating sperm flagella
motility and germ cell maturation (4). However, increasing evidence has
demonstrated that SPAG6 may influence the progression of certain
malignant diseases, including gastric, breast and lung cancer
(5). Furthermore, a number of
reports have demonstrated that, compared with normal controls,
SPAG6 is upregulated in BM from patients with MDS, MDS-AML or de
novo AML, and that high expression levels of SPAG6 are
associated with poor survival (6,7). SKM-1
is an MDS-AML cell line that was established from the peripheral
blood of a male patient with MDS transformation to AML. As there
are currently no MDS cell lines, SKM-1 cells are considered to be
the optimal model for investigating the pathogenesis of MDS
(8). Previous studies have
demonstrated that lentivirus-mediated SPAG6 knockdown suppresses
proliferation and promotes apoptosis and differentiation in SKM-1
cells, confirming that SPAG6 influences the progression of MDS
(7,9–11).
However, the effects of SPAG6 on autophagy and its underlying
mechanisms have not yet been identified.
Autophagy is a physiological process in which
unnecessary intracellular materials are degraded by the lysosome
for recycling. Therefore, it is essential for the maintenance of
cell development, metabolism and cellular homeostasis. However,
previous evidence has demonstrated that autophagy exerts both
pro-survival or pro-apoptotic functions in disease, depending on
the tumor condition, disease progression and cellular
microenvironment (12). There is
growing evidence that autophagy serves a critical role in the
pathogenesis of MDS and AML; autophagy regulates the
differentiation and development of HSCs by removing damaged
mitochondria and reactive oxygen species (ROS) (13,14).
Further studies have indicated that a lack of autophagy in HSCs may
facilitate the progression of MDS and AML (15,16). In
addition, a recent study demonstrated that, compared with normal
controls, autophagy was decreased, and damaged mitochondria and ROS
levels were increased in patients with MDS and MDS-AML (17), suggesting that autophagy functions as
a tumor suppressor in the progression of MDS. However, the possible
cause of decreased autophagy in MDS remains unclear. Thus, studies
are required to investigate the mechanisms underlying decreased
autophagy in MDS to provide guidance for the prevention and
treatment of MDS.
AMP-activated protein kinase (AMPK) is a key factor
in regulating cellular energy and metabolism, and serves an
essential role in the pro-differentiation and anti-leukemic
properties of myeloid malignancies, via regulation of multiple
downstream signaling molecules to induce autophagy (18). For example, it directly or indirectly
inhibits cancer-promoting mTOR complex 1 (mTORC1) signaling and
activates pro-apoptotic p53 (19,20).
However, previous studies have demonstrated that the expression
levels of autophagy, differentiation and AMPK mRNA were
significantly decreased in cells collected from patients with
high-risk MDS compared with patients with low-risk MDS or healthy
controls (7,17,18). A
previous study demonstrated that SPAG6 silencing increases the
differentiation of SKM-1 cells (7),
which suggests that SPAG6 silencing may regulate differentiation
and autophagy in SKM-1 cells via activation of the AMPK signaling
pathway. To the best of our knowledge, however, the potential
molecular mechanism has not yet been clarified,
The present study demonstrated that SPAG6 knockdown
may trigger autophagy and apoptosis in SKM-1 cells and that
autophagy inhibitors attenuated SPAG6 knockdown-induced cell
apoptosis. Furthermore, the present study investigated the
underlying mechanism of SPAG6 knockdown-induced autophagy, which
may be mediated by regulation of the AMPK/mTOR/ULK1 signaling
pathway.
Materials and methods
Cell culture and infection
SKM-1, the human MDS-AML cell line, was provided by
Professor Jianfeng Zhou of Tongji Medical College of Huazhong
University of Science and Technology (Wuhan, China) -and cultured
in RPMI-1640 medium (Corning Inc.) containing 10% fetal bovine
serum (Capricorn Scientific GmbH) without antibiotics. Cells were
grown in complete medium with 3-methyladenine (3-MA; 5 mM/l),
chloroquine (CQ; 10 µM/l) or the AMPK inhibitor Compound C (5
µM/l), purchased from Selleck Chemicals and all cells were
incubated at 37°C with 5% CO2. Lentiviral short hairpin
RNAs (shRNAs) were designed and synthesized by Shanghai GeneChem
Co., Ltd. A total of three different shRNAs for SPAG6 were designed
(sequences listed in Table SI) and
the shRNA used as the negative control (NC) was
5′-TTCTCCGAACGTGTCACGT-3′. SKM-1 cells at the logarithmic stage
were seeded in 24-well plates (5×104 cells/well) before
being transfected with SPAG6-shRNA lentivirus or NC-shRNA
lentivirus at a multiplicity of infection of 80 in the presence of
5 µg/ml polybrene, an enhancing reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). Another 500 µl fresh complete RPMI-1640 medium
supplemented with 10% FBS, without antibiotics was added to cells
in each group 20 h later, and the cells were then resuspended in
fresh complete medium after an additional 24 h. Three days later,
fresh complete medium with puromycin at a concentration of 0.5
mg/ml was used to culture the transfected cells.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted separately from cells
treated with NC-shRNA or SPAG6-shRNA using TRIzol reagent (Takara
Biotechnology Co., Ltd.); then, the RNA samples were reverse
transcribed into cDNA using a PrimeScript RT reagent kit (Takara
Biotechnology Co., Ltd.), according to the manufacturer's
protocols. qPCR was subsequently performed using a CFX-Connect
Real-Time PCR system (Bio-Rad Laboratories, Inc.) and TB Green
Master Mix (Takara Biotechnology Co., Ltd.) for a total of 45
cycles, following standard assay procedures. The thermocycling
parameters used were 95°C for 30 sec, followed by 45 cycles at 95°C
for 5 sec and 62°C for 30 sec. The mRNA expression levels of SPAG6
and autophagy-associated genes were calculated using the
2−ΔΔCq method (21) with
GAPDH as an internal control. The primer sequences are listed in
Table I.
 | Table I.Primer sequences used for RT-qPCR
analysis. |
Table I.
Primer sequences used for RT-qPCR
analysis.
| Gene | Forward
(5′-3′) | Reverse
(5′-3′) |
|---|
| GAPDH |
CTTTGGTATCGTGGAAGGACTC |
GTAGAGGCAGGGATGATGTTCT |
| SPAG6 |
AGTGCGACATTCTTCCACAGCTTG |
GCGTATCCAGTGCTCCACAATCG |
| LC3 |
CCGACTTATTCGAGAGCAGCATCC |
GTCCGTTCACCAACAGGAAGAAGG |
| Beclin1 |
ATCTAAGGAGCTGCCGTTATAC |
CTCCTCAGAGTTAAACTGGGTT |
| ATG5 |
GATGGGATTGCAAAATGACAGA |
GAAAGGTCTTTCAGTCGTTGTC |
| ATG7 |
TGTATAACACCAACACACTCGA |
GGCAGGATAGCAAAACCAATAG |
Western blot analysis and
antibodies
Cells from each group were collected and washed with
PBS three times. Then, all cells were lysed in RIPA lysis buffer
(Beyotime Institute of Biotechnology) containing PMSF and
phosphatase inhibitor (Boster Biological Technology). A
bicinchoninic acid kit (Beyotime Institute of Biotechnology) was
used to measure protein concentrations, according to the
manufacturer's protocol. Total cellular proteins (40 µg) in each
lane were separated via SDS-PAGE on 10 or 12% gels and transferred
onto PVDF membranes (Merck KGaA) (12% gel was used for LC3; 10% gel
was used for SPAG6, P62, Beclin1, ATG5, ATG7, cleaved caspase 3,
Bcl-2, Bax, p-AMPK, AMPK, p-mTOR, mTOR, p-ULK1, ULK1 and GAPDH).
Then, the membranes were blocked with 5% skimmed milk at room
temperature for 2 h before incubation with the corresponding
primary antibodies overnight at 4°C. The next day, the membranes
were incubated with goat anti-rabbit secondary antibody (cat. no.
A0216; 1:3,000; Beyotime Institute of Biotechnology) for 1 h at
room temperature. Finally, the bands were visualized using an
enhanced chemiluminescence kit (Advansta, Inc.), and the relative
protein expression levels were analyzed using Fusion FX Spectra
software (version 7; Vilber Lourmat). The primary antibodies were
as follows: SPAG6 (cat. no. HPA038440; 1:1,000; Sigma-Aldrich;
Merck KGaA); GAPDH (cat. no. AG019, 1:1,500 Beyotime Institute of
Biotechnology); Beclin1, P62, cleaved caspase-3, Bcl-2, Bax, mTOR
and phosphorylated (p-)mTOR (cat. nos. ab210498, ab109012, ab2302,
ab32124, ab32503, ab32028 and ab109268, respectively; all 1:1,000;
all Abcam); LC3, ATG5, ATG7, AMPK, p-AMPK, ULK1 and p-ULK1 (cat.
nos. 12741, 12994, 8558, 2532, 2535, 8054 and 5869, respectively;
all 1:1,000; all Cell Signaling Technology, Inc.).
Transmission electron microscopy
Cells treated with NC-shRNA or SPAG6-shRNA alone, as
well as cells co-treated with SPAG6-shRNA and 3-MA or CQ were
collected, centrifuged at 350 × g for 5 min at room temperature and
subsequently fixed with 2.5% glutaraldehyde and 1% osmic acid for 2
h at 4°C. Next, the prepared cells were washed twice with ddH2O and
embedded following dehydration with acetone. Then, the samples were
solidified and cut into 1-µm sections, which were double-stained
with uranium acetate for 10 min and lead citrate for 30 min, at
35°C. Sections were observed under a Hitachi-7500 transmission
electron microscope (Hitachi, Ltd.; magnification, ×20,000).
Flow cytometry
Cells treated with NC-shRNA or SPAG6-shRNA alone and
cells co-treated with SPAG6-shRNA and 3-MA or CQ, as well as cells
co-treated with Compound C and NC-shRNA or SPAG6-shRNA were
collected and washed with PBS twice before being resuspended in 500
µl PBS. Then, the cells were double-stained with annexin V APC and
DAPI (BD Pharmingen; BD Biosciences) and incubated in the dark at
room temperature for 10 min. The fluorescence intensities of the
cells were examined immediately by flow cytometry (CytoFLEX,
Beckman Coulter, Inc.).
Statistical analysis
SPSS (version 24.0; IBM Corp.) and GraphPad Prism
software (version 8.0; GraphPad Software, Inc.) were used to
analyze all data. Data are presented as the mean ± standard
deviation. An unpaired Student's t-test was used for two-group
analysis and a One-way analysis of variance was used for
comparisons among multiple groups, followed by the Newman-Keuls
post hoc test. All experiments were performed in triplicate.
P<0.05 was considered to indicate a statistically significant
difference.
Results
SPAG6 silencing induces autophagy in
SKM-1 cells
To the best of our knowledge, there is no previous
evidence concerning the association between SPAG6 and autophagy in
MDS. It is well known that the expression of autophagy associated
genes (ATG) increases while the expression of autophagy receptors
decreases when autophagy is activated (22). LC3 is a homologue of ATG8, including
two forms of LC3-I and LC3-II, the latter being a marker of
autophagy; Beclin1 is a homologue of ATG6; these, as well as ATG5
and ATG7 are autophagy associated genes (22). P62 acts as an autophagy receptor,
fusing autophagosomes (including cargo) with lysosomes, and then
P62 and cargo are degraded by autophagolysosomes, resulting in a
decrease in P62 expression levels (22). Therefore, in order to investigate
whether SPAG6 is associated with autophagy in SKM-1 cells, cells
were transfected with shRNAs. The optimal shRNA for SPAG6 was
screened via RT-qPCR and western blot analysis, and was identified
as SPAG6-shRNA3 (Fig. S1).
Following the suppression of the expression levels of SPAG6, the
expression levels of autophagy-related genes were analyzed:
Compared with its expression levels in SKM-1 cells transfected with
NC-shRNA lentivirus, SPAG6 mRNA and protein expression levels in
cells transfected with SPAG6-shRNA lentivirus were significantly
decreased. Moreover, there was an increase in mRNA and protein
expression levels of LC3-II, Beclin1, ATG5 and ATG7, and a decrease
in p62 (Fig. 1A and B). Transmission
electron microscopy is considered to be the gold standard for
observing autophagosome formation. In the present study, a
significant increase in autophagosome formation was detected in the
SPAG6 knockdown group (Fig. 1C and
D). Taken together, the results of the present study indicate
that SPAG6 knockdown may induce autophagy in SKM-1 cells.
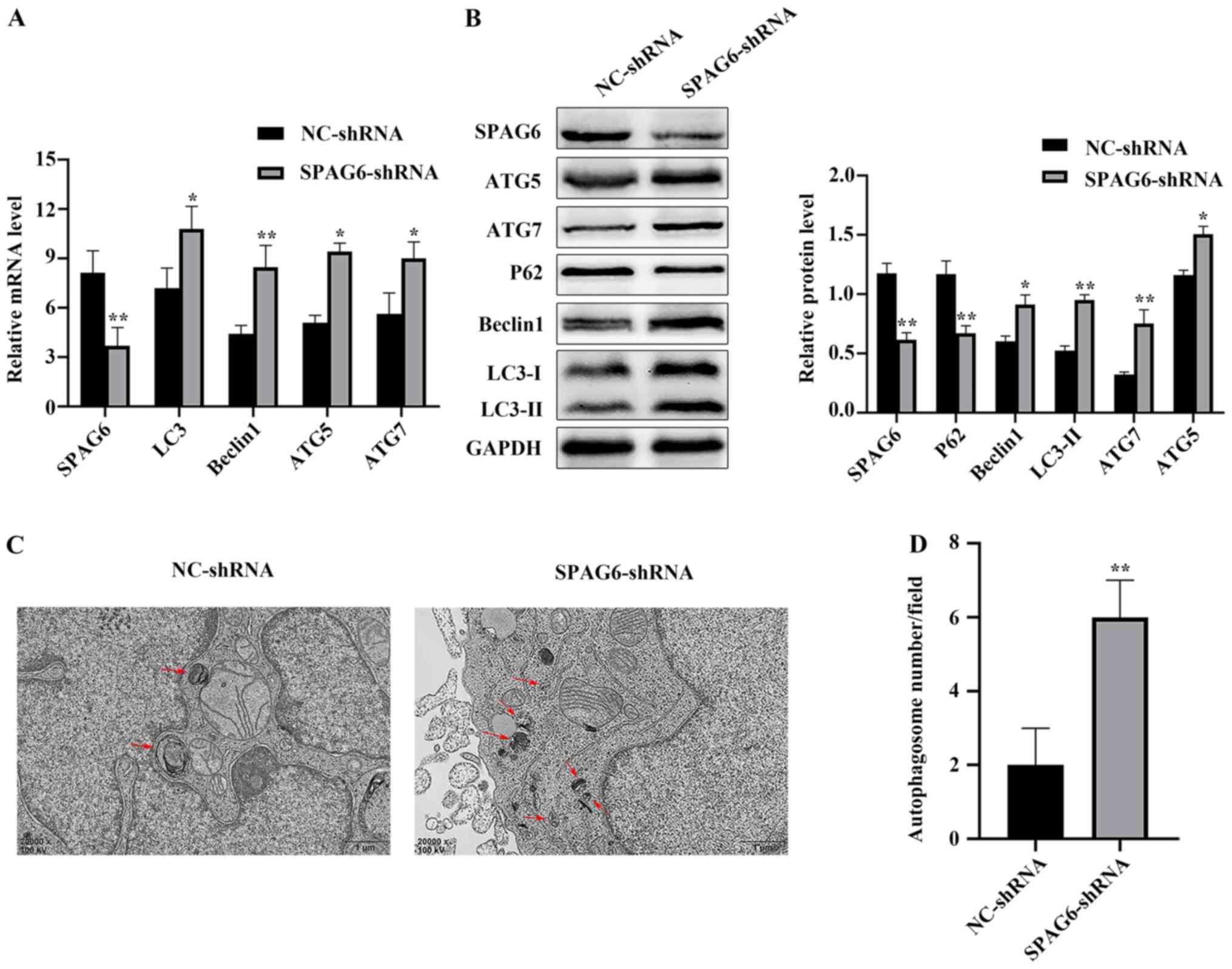 | Figure 1.SPAG6 silencing induces autophagy in
SKM-1 cells. (A) Relative mRNA expression levels of SPAG6, LC3,
Beclin1, ATG5 and ATG7 in each group were detected using RT-qPCR.
(B) Relative protein expression levels of SPAG6, LC3, Beclin1, p62,
ATG5 and ATG7 in each group were detected using western blot
analysis. GAPDH was used as a loading control. (C) Transmission
electron microscopy revealed the intracellular ultrastructures in
each group (magnification, ×20,000). The images display a
representative experiment from three independent experiments. Red
arrows indicate autophagic vacuoles. Scale bar, 1 µm. (D) Number of
autophagosomes per field was quantified. Data are presented as the
mean ± standard deviation of three independent experiments.
*P<0.05; **P<0.01 vs. NC-shRNA. NC, negative control; shRNA,
short hairpin RNA; SPAG6, sperm-associated antigen 6; LC3,
microtubule-associated protein 1 light chain 3; Beclin1,
autophagy-associated protein 6; p62, sequestosome 1; ATG5,
autophagy-related protein 5; ATG7, autophagy-related protein 7. |
SPAG6 knockdown triggers apoptosis in
SKM-1 cells
The present study used western blot analysis and
flow cytometry to detect the apoptosis rate in each group. Compared
with the NC-shRNA group, Bax and cleaved caspase-3 protein levels
were significantly increased in the SPAG6-shRNA group, whereas
Bcl-2 protein levels were decreased (Fig. 2A). These data indicated that SPAG6
knockdown significantly induced cell apoptosis. This was confirmed
using flow cytometry (Fig. 2B and
C). Overall, these results indicated that SPAG6 silencing may
induce apoptosis in SKM-1 cells, which is consistent with previous
evidence (9–11).
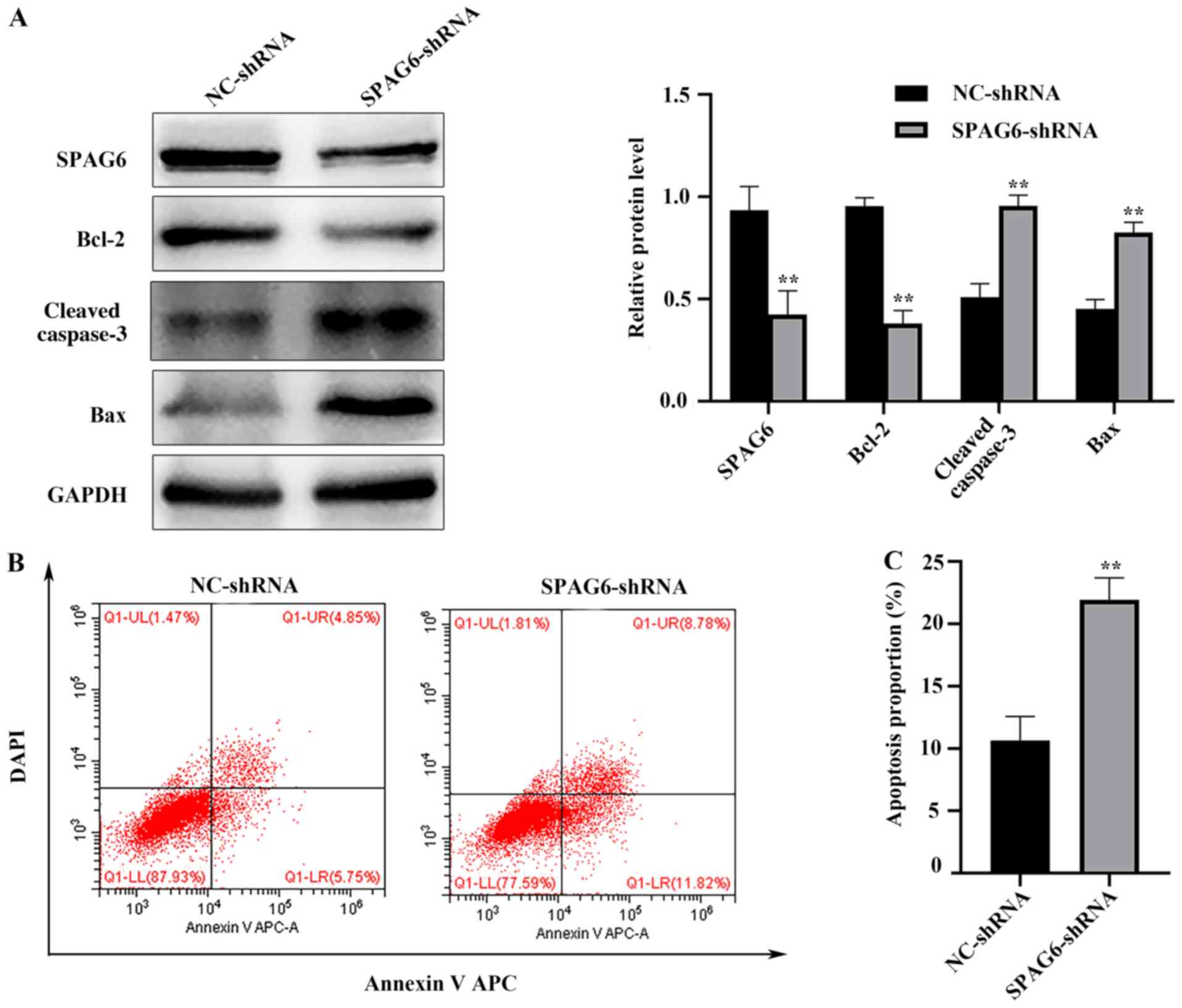 | Figure 2.SPAG6 knockdown triggers apoptosis in
SKM-1 cells. (A) Relative protein expression levels of SPAG6,
Bcl-2, Bax and cleaved caspase-3 in each group were detected using
western blot analysis. (B) After annexin V APC/DAPI
double-staining, the total apoptosis rate of SKM-1 cells treated
with NC-shRNA or SPAG6-shRNA was detected by flow cytometry. The
images display a representative experiment from three independent
experiments. (C) Analysis of the apoptotic rate shown in (B). Data
are presented as the mean ± standard deviation of three independent
experiments. **P<0.01 vs. NC-shRNA. NC, negative control; shRNA,
short hairpin RNA; SPAG6, sperm-associated antigen 6; APC,
allophycocyanin; LL, lower left; UL, upper left; LR, lower right;
UR, upper right. |
Inhibition of autophagy attenuates
SPAG6 knockdown- induced apoptotic cell death in SKM-1 cells
Autophagy activation has previously been reported to
induce apoptosis in certain types of human cancer (23,24).
Thus, the present study used the autophagy inhibitors 3-MA and CQ
to block autophagy in order to investigate the effect of autophagy
on apoptosis induced by SPAG6 silencing. It has previously been
reported that 3-MA and CQ inhibit autophagy at different stages:
3-MA inhibits autophagy at an early stage, resulting in decreased
LC3-II and Beclin1 expression levels, and increased P62 expression
levels (25). However, CQ suppresses
autophagy by inhibiting the degradation of autophagy lysosomes,
which results in additional LC3-II and Beclin1 accumulation and
decreased p62 degradation (26). The
present study demonstrated that, compared with the expression
levels in cells transfected with SPAG6-shRNA alone, LC3-II and
Beclin1 protein expression levels were significantly decreased in
SKM-1 cells cotreated with SPAG6-shRNA/3-MA, while P62 protein
expression levels were increased; these changes were accompanied by
decreased cleaved caspase-3 and Bax protein expression levels and
increased Bcl-2 protein expression levels (Fig. 3A and D). Furthermore, SKM-1 cells
treated with SPAG6-shRNA/CQ yielded the same results, except
increased LC3-II and Beclin1 protein expression levels (Fig. 3A and D). In addition, the number of
autophagosomes was notably decreased in SPAG6-shRNA/3-MA-cotreated
SKM-1 cells but was increased in SKM-1 cells treated with
SPAG6-shRNA/CQ compared with cells transfected with SPAG6-shRNA
alone (Fig. 3B and C). Furthermore,
the present study used flow cytometry to analyze the effects of
3-MA and CQ on apoptosis. Levels of annexin V APC/DAPI positive
cells were decreased in cells co-treated with SPAG6-shRNA and 3-MA
or CQ compared with cells treated with SPAG6-shRNA alone (Fig. 3E and F). Collectively, the current
results indicate that inhibiting autophagy with 3-MA and CQ may
attenuate SPAG6 knockdown-mediated apoptosis in SKM-1 cells.
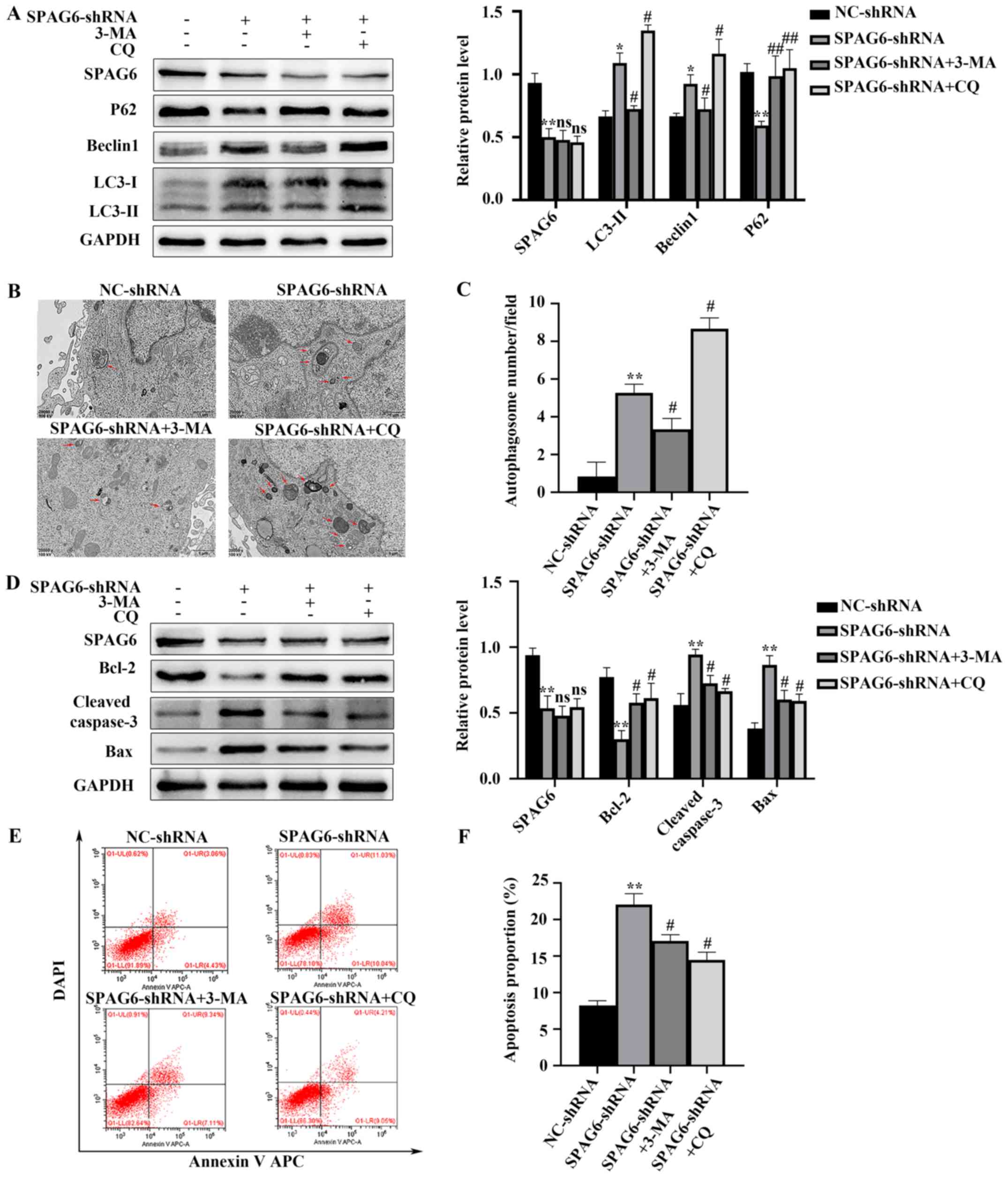 | Figure 3.Inhibiting autophagy with autophagy
inhibitors attenuates SPAG6 knockdown-induced apoptosis. Cells were
treated with SPAG6-shRNA lentivirus or NC-shRNA lentivirus alone,
or co-treated with 3-MA or CQ and SPAG6-shRNA. (A) Relative protein
expression levels of SPAG6, LC3, Beclin1 and P62 in each group were
detected via western blot analysis. (B) Transmission electron
microscopy revealed the intracellular ultrastructures of each group
(magnification, ×20,000). The images display a representative
experiment from three independent experiments. Red arrows indicate
autophagic vacuoles. Scale bar, 1 µm. (C) Number of autophagosomes
per field was quantified. (D) Relative protein expression levels of
SPAG6, Bcl-2, Bax and cleaved caspase-3 in each group were detected
using western blot analysis. (E) Following annexin V APC/DAPI
double-staining, the total apoptosis rate of SKM-1 cells treated
with NC-shRNA or SPAG6-shRNA alone and SKM-1 cells co-treated with
SPAG6-shRNA and 3-MA or CQ was detected using flow cytometry. The
images display a representative experiment from three independent
experiments. (F) Analysis of the apoptotic rate shown in (E) Data
are presented as the mean ± standard deviation of three independent
experiments. *P<0.05; **P<0.01 vs. NC-shRNA,
#P<0.05; ##P<0.01 vs. SPAG6-shRNA. NC,
negative control; shRNA, short hairpin RNA; SPAG6, sperm-associated
antigen 6; LC3, microtubule-associated protein 1 light chain 3;
Beclin1, autophagy-associated protein 6; p62, sequestosome 1; CQ,
chloroquine; 3-MA, 3-methyladenine; APC, allophycocyanin; LL, lower
left; UL, upper left; LR, lower right; UR, upper right; ns, not
significant. |
SPAG6 knockdown induces autophagy via
the AMPK/mTOR/ULK1 pathway
There is evidence to indicate that autophagy may be
induced via the AMPK/mTOR/ULK1 pathway (27,28). In
order to investigate whether this pathway participates in SPAG6
knockdown-mediated autophagy in SKM-1 cells, the present study
detected AMPK, p-AMPK, ULK1, p-ULK1, mTOR and p-mTOR protein levels
using western blotanalysis. As demonstrated in Fig. 4A, although the total AMPK, ULK1 and
mTOR protein levels were not altered, p-AMPK and p-ULK1 protein
levels were notably increased following SPAG6 knockdown, while the
p-mTOR protein level was significantly decreased; these changes
were accompanied by increases in protein levels of LC3-II, Beclin1,
cleaved caspase-3 and Bax, and decreases in protein levels of P62
and Bcl-2 (Fig. 4A and B). However,
compared with the levels in cells transfected with SPAG6-shRNA
alone, p-AMPK and p-ULK1 protein levels were decreased, whereas the
p-mTOR protein level was increased in cells co-treated with SPAG6
and Compound C. Furthermore, the protein level of LC3-II was
significantly decreased, indicating that autophagy was suppressed
via AMPK inhibition (Fig. 4A and B).
Moreover, annexin V APC/DAPI double-staining analysis revealed
fewer apoptotic cells among those co-treated with SPAG6-shRNA and
Compound C compared with those treated with SPAG6-shRNA alone
(Fig. 4C and D). Taken together, the
current findings indicate that SPAG6 silencing induced autophagy
via the AMPK/mTOR/ULK1 signaling pathway.
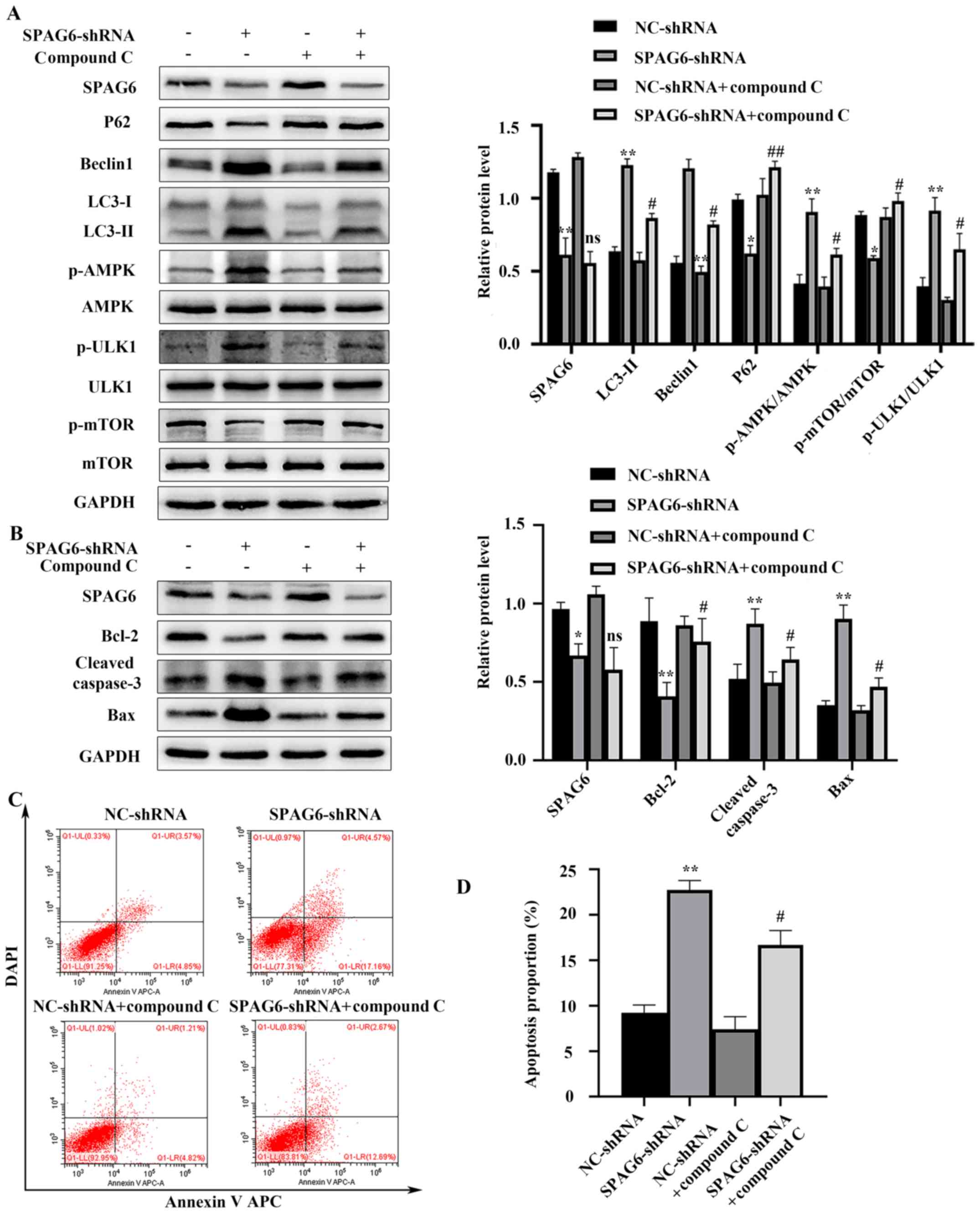 | Figure 4.Inhibition of AMPK by Compound C
attenuates SPAG6 knockdown-mediated autophagy and apoptosis in
SKM-1 cells. Cells were treated with SPAG6-shRNA lentivirus or
NC-shRNA lentivirus alone, or co-treated with Compound C and
NC-shRNA lentivirus or SPAG6-shRNA lentivirus. (A) Relative protein
expression levels of SPAG6, LC3, Beclin1, P62, p-AMPK, p-ULK1 and
p-mTOR in each group were detected using western blot analysis. (B)
Relative protein expression levels of SPAG6, Bcl-2, Bax and cleaved
caspase-3 in each group were detected using western blot analysis.
(C) Following annexin V APC/DAPI double-staining, the total
apoptosis rate of SKM-1 cells treated with NC-shRNA or SPAG6-shRNA
alone and SKM-1 cells cotreated with Compound C and NC-shRNA or
SPAG6-shRNA was detected using flow cytometry. The images display a
representative experiment from three independent experiments. (D)
Analysis of the apoptotic rate shown in (C). Data are presented as
the mean ± standard deviation of three independent experiments.
*P<0.05; **P<0.01 vs. NC-shRNA, #P<0.05;
##P<0.01 vs. SPAG6-shRNA. NC, negative control;
shRNA, short hairpin RNA; SPAG6, sperm-associated antigen 6; LC3,
microtubule-associated protein 1 light chain 3; Beclin1,
autophagy-associated protein 6; p62, sequestosome 1; p,
phosphorylated; AMPK, AMP-activated protein kinase; ULK1,
unc-51-like autophagy activating kinase; APC, allophycocyanin; LL,
lower left; UL, upper left; LR, lower right; UR, upper right; ns,
not significant. |
Discussion
A number of studies have demonstrated that
chromosomal abnormalities and aberrant gene expression levels serve
essential roles in the evolution and progression of MDS (29,30). It
has been reported that SPAG6 is upregulated in patients with MDS
and AML and that it may regulate apoptosis, proliferation and
differentiation in SKM-1 cells (9–11).
However, to the best of our knowledge, there are currently no data
concerning the role of SPAG6 in autophagy in SKM-1 cells. The
present study demonstrated that SPAG6 silencing induced autophagy
and apoptosis in SKM-1 cells and that pharmacologically inhibiting
autophagy decreased SPAG6 knockdown-induced apoptosis. Furthermore,
the present study demonstrated that SPAG6 downregulation mediated
p-AMPK and p-ULK1 activation and that blocking the AMPK/mTOR/ULK1
signaling pathway attenuated SPAG6 knockdown-induced autophagy and
apoptosis. Hence, the present study indicated that SPAG6 silencing
induced autophagy, which further promoted apoptosis in SKM-1 cells,
and that the AMPK/mTOR/ULK1 signaling pathway was involved in
regulating autophagy.
As a major mechanism of cell component degradation
and recycling, autophagy is indispensable in the maintenance of
cell homeostasis and metabolism. However, dysfunction of autophagy
is involved in a number of pathological conditions that may lead to
tumorigenesis. Previous studies have shown that abnormal gene
expression levels are important in regulating autophagy in tumor
cells (15,31). For example, mice lacking the
autophagy-associated gene ATG7 in HSCs develop atypical hyperplasia
in the BM, which is similar to MDS and can progress to AML
(32). In bladder cancer cells, G9a
inhibition induces autophagy, which further results in autophagic
cell death (31). It has been
reported that SPAG6, an oncogene that is aberrantly upregulated in
patients with MDS and AML, may regulate proliferation, apoptosis
and differentiation in SKM-1 cells (7,9–11). The present study used a shRNA
lentiviral vector to knockdown SPAG6 expression levels in order to
investigate the impact of SPAG6 on autophagy in SKM-1 cells. It was
demonstrated that lentivirus-mediated SPAG6 knockdown induced
autophagy in SKM-1 cells, as evidenced by an increase in
autophagosome formation (detected using transmission electron
microscopy) and increases in LC3-II and Beclin1 protein expression
levels, and a decrease in p62 protein expression level (detected
using western blot analysis). Previous studies have indicated that
SPAG6 regulates differentiation, proliferation, apoptosis and drug
sensitivity in neurons and MDS (7,9–11,33,34). To
the best of our knowledge, this is the first study to provide
evidence for the role of SPAG6 in regulating autophagy in SKM-1
cells. The present study investigated the association between SPAG6
and apoptosis in SKM-1 cells, as growing evidence has demonstrated
that abnormal proliferation and apoptosis are related to the
pathogenesis of MDS. Previous studies have demonstrated that
lentiviral transfection-mediated knockdown of SPAG6 inhibits the
proliferation of SKM-1 cells, which may be initiated via the
AKT/FOXO/p27Kip1 pathway (7,9). In addition, SPAG6 downregulation
significantly increases apoptosis in SKM-1 cells, and is
accompanied by a significant increase in tumor suppressor genes
(P53 and PTEN) (9). Further studies
have indicated that SPAG6 knockdown-induced apoptosis in SKM-1
cells may occur via a caspase-independent mechanism, such as
PTEN/PI3K/AKT pathway activation or TNF-related apoptosis-inducing
ligand pathway induction (10,11). In
accordance with these findings, the present study confirmed that
lentivirus-mediated SPAG6 knockdown increased apoptosis in SKM-1
cells.
Autophagic cell death and apoptosis are both types
of programmed cell death. The association between autophagy and
apoptosis is reported to be complicated because molecules that
affect apoptosis also play an important role in autophagy
activation. For example, downregulating Bcl-2 may induce autophagy
by decreasing its binding affinity to the BH3 domain of Beclin.
Subsequently, autophagy activation may increase/decrease apoptosis
or sensitivity to chemotherapeutic drugs (35–37).
Additionally, autophagy can be both cytotoxic and cytoprotective;
previous studies have demonstrated that the autophagy inhibitor
3-MA decreases apoptosis triggered by ATG3 overexpression in SKM-1
cells (36), and in non-small cell
lung cancer cells, scutellaria radix-triggered apoptosis is
attenuated by autophagy inhibitors (23), which indicates that autophagy has a
cytotoxic role in carcinoma treatment. By contrast, certain studies
have demonstrated that the autophagy inhibitor CQ enhances
apoptosis in SKM-1 cells with long-term exposure to azacytidine
(37), and that in ovarian cancer
cells, inhibition of autophagy increases paeonol-induced apoptosis
(38), which indicates that
autophagy may exert a pro-survival role in cancer cells. In order
to investigate the association between autophagy and apoptosis in
SKM-1 cells, the present study applied the autophagy inhibitors
3-MA and CQ and demonstrated that inhibition of autophagy
significantly decreased SPAG6 knockdown-induced apoptosis in SKM-1
cells. This was determined by the decreased number of annexin V
APC/DAPI-positive cells and apoptosis-associated protein expression
levels, which were detected using flow cytometry and western blot
analysis, respectively. Collectively, the results of the present
study indicate that SPAG6 knockdown-mediated autophagy functions as
a cytotoxic mechanism, which may further induce apoptosis in SKM-1
cells. However, the underlying mechanism of SPAG6
knockdown-mediated autophagy in regulating apoptosis in SKM-1 cells
remains unclear. It has been reported that the regulation of
apoptosis by autophagy involves numerous mechanisms, such as the
Bcl-2 homology-3 domain of Beclin1 and Bcl-2, P62 and
PI3K/PTEN/AKT/mTOR (35,39,40). In
the present study, SPAG6 knockdown increased Beclin1 expression
levels in SKM-1 cells. Thus, it was hypothesized that Beclin1
activation participated in regulating apoptosis via SPAG6
knockdown-mediated autophagy in SKM-1 cells. Further studies are
required in order to identify the underlying molecular mechanisms
involved in the interaction between autophagy and apoptosis both
in vivo and in vitro.
A number of signaling pathways have been reported to
regulate autophagy, including the AMPK and the PI3K/AKT/mTOR
pathways (41,42). The role of AMPK in regulating
differentiation and autophagy in HSCs has been a focus of recent
research. Mounting evidence has confirmed that autophagy may be
regulated by AMPK, which is activated by the tumor suppressor liver
kinase B1 and calcium/calmodulin-dependent protein kinase kinase 2;
then, activated AMPK stimulates autophagy by directly
phosphorylating and activating ULK1 at Ser555, or by inhibiting
mTORC1, which further upregulates p-ULK1 and ATG13 (27,43). The
present study demonstrated that, compared with levels in NC-shRNA
lentivirus transfected-SKM-1 cells, LC3-II, Beclin1, p-AMPK and
p-ULK1 protein levels increased in cells that were transfected with
SPAG6-shRNA lentivirus, whereas p62 and p-mTOR protein levels were
significantly decreased. Furthermore, inhibition of the
AMPK/mTOR/ULK1 signaling pathway using Compound C significantly
decreased autophagy, which further decreased the cytotoxicity
mediated by SPAG6 knockdown. Collectively, the results of the
present study demonstrated that SPAG6 knockdown induced autophagy
via the AMPK/mTOR/ULK1 axis. In addition, these findings are
consistent with a recent report demonstrating that iron overload
induces autophagy via mitochondrial dysfunction, ROS accumulation
and activation of the AMPK signaling pathway in mesenchymal stromal
cells from patients with MDS (44),
indicating that the AMPK signaling pathway regulates autophagy in
MDS cells. It has been reported that the Nod-like receptor protein
3 (NLRP3) inflammasome is activated in patients with MDS and plays
a key role in driving the progress of MDS phenotype (45). As a key regulator in NLRP3-dependent
inflammation and immunity, AMPK may negatively regulate the
activation of the NLRP3 inflammasome, indicating an inverse
association between them (46). The
present study demonstrated that knockdown of SPAG6 triggered
autophagy via the activation of AMPK, which further increased
apoptosis in SKM-1 cells. It was hypothesized that SPAG6 mediates
AMPK downregulation, which further activates the NLRP3 inflammasome
during the progression of MDS. This must be verified by future
studies. In addition, a previous study demonstrated that SPAG6
knockdown increased the differentiation level in SKM-1 cells
(7). Therefore, further research is
required in order to determine whether SPAG6 knockdown-mediated
activation of the AMPK signaling pathway participates in this
molecular mechanism.
In conclusion, the present study indicated that
SPAG6 knockdown induced autophagy via the AMPK/mTOR/ULK1 signaling
pathway in SKM-1 cells, and that inhibiting autophagy decreased
SPAG6 knockdown-mediated apoptosis. The results of the present
study contribute to a better understanding of the mechanisms
underlying the role of SPAG6 in the tumor microenvironment and
further demonstrate that SPAG6 may be a potential target to
attenuate the progression of MDS.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank Dr Lixue Chen and Dr
Xiaojuan Deng (The First Affiliated Hospital of Chongqing Medical
University, Chongqing, China) for providing technical support.
Funding
The present study was supported by funds from the
National Natural Sciences Foundation of China (grant no. 81570109)
and the Natural Science Foundation of Chongqing (grant no.
CSTC2013jjB0145).
Availability of data and materials
The datasets used or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
MZ designed and performed the experiments and wrote
the manuscript. JL helped to analyze the data. XL contributed to
the conception of the study. LL contributed to the study design and
provided financial support. All authors read and approved the final
version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Corey SJ, Minden MD, Barber DL, Kantarjian
H, Wang JC and Schimmer AD: Myelodysplastic syndromes: The
complexity of stem-cell diseases. Nat Rev Cancer. 7:118–129. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Greenberg PL, Stone RM, Al-Kali A, Barta
SK, Bejar R, Bennett JM, Carraway H, De Castro CM, Deeg HJ, DeZern
AE, et al: Myelodysplastic syndromes, Version 2.2017, NCCN clinical
practice guidelines in oncology. J Natl Compr Canc Netw. 15:60–87.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Saber W, Cutler CS, Nakamura R, Zhang MJ,
Atallah E, Rizzo JD, Maziarz RT, Cortes J, Kalaycio ME and Horowitz
MM: Impact of donor source on hematopoietic cell transplantation
outcomes for patients with myelodysplastic syndromes (MDS). Blood.
122:1974–1982. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sapiro R, Kostetskii I, Olds-Clarke P,
Gerton GL, Radice GL and Strauss III JF: Male infertility, impaired
sperm motility, and hydrocephalus in mice deficient in
sperm-associated antigen 6. Mol Cell Biol. 22:6298–6305. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Siliņa K, Zayakin P, Kalniņa Z, Ivanova L,
Meistere I, Endzeliņš E, Abols A, Stengrēvics A, Leja M, Ducena K,
et al: Sperm-associated antigens as targets for cancer
immunotherapy: Expression pattern and humoral immune response in
cancer patients. J Immunother. 34:28–44. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Steinbach D, Schramm A, Eggert A, Onda M,
Dawczynski K, Rump A, Pastan I, Wittig S, Pfaffendorf N, Voigt A,
et al: Identification of a set of seven genes for the monitoring of
minimal residual disease in pediatric acute myeloid leukemia. Clin
Cancer Res. 12:2434–2441. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jiang M, Chen Y, Deng L, Luo X, Wang L and
Liu L: Upregulation of SPAG6 in myelodysplastic syndrome: Knockdown
inhibits cell proliferation via AKT/FOXO signaling pathway. DNA
Cell Biol. 38:476–484. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nakagawa T, Matozaki S, Murayama T,
Nishimura R, Tsutsumi M, Kawaguchi R, Yokoyama Y, Hikiji K, Isobe T
and Chihara K: Establishment of a leukaemic cell line from a
patient with acquisition of chromosomal abnormalities during
disease progression in myelodysplastic syndrome. Br J Haematol.
85:469–476. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang B, Wang L, Luo X, Chen L, Yang Z and
Liu L: SPAG6 silencing inhibits the growth of the malignant myeloid
cell lines SKM-1 and K562 via activating p53 and caspase
activation-dependent apoptosis. Int J Oncol. 46:649–656. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li X, Yang B, Wang L, Chen L, Luo X and
Liu L: SPAG6 regulates cell apoptosis through the TRAIL signal
pathway in myelodysplastic syndromes. Oncol Rep. 37:2839–2846.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yin J, Li X, Zhang Z, Luo X, Wang L and
Liu L: SPAG6 silencing induces apoptosis in the myelodysplastic
syndrome cell line SKM-1 via the PTEN/PI3K/AKT signaling pathway
in vitro and in vivo. Int J Oncol. 53:297–306.
2018.PubMed/NCBI
|
|
12
|
White E and DiPaola RS: The double-edged
sword of autophagy modulation in cancer. Clin Cancer Res.
15:5308–5316. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mortensen M, Ferguson DJ, Edelmann M,
Kessler B, Morten KJ, Komatsu M and Simon AK: Loss of autophagy in
erythroid cells leads to defective removal of mitochondria and
severe anemia in vivo. Proc Natl Acad Sci USA. 107:832–837. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Watson AS, Mortensen M and Simon AK:
Autophagy in the pathogenesis of myelodysplastic syndrome and acute
myeloid leukemia. Cell Cycle. 10:1719–1725. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mortensen M, Watson AS and Simon AK: Lack
of autophagy in the hematopoietic system leads to loss of
hematopoietic stem cell function and dysregulated myeloid
proliferation. Autophagy. 7:1069–1070. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ali AM and Raza A: Two different ‘tales’
of ATG7: Clinical relevance to myelodysplastic syndromes. Mol Cell
Oncol. 3:e12126862016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jiang H, Yang L, Guo L, Cui N, Zhang G,
Liu C, Xing L, Shao Z and Wang H: Impaired mitophagy of nucleated
erythroid cells leads to anemia in patients with myelodysplastic
syndromes. Oxid Med Cell Longev. 2018:63280512018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jacquel A, Luciano F, Robert G and
Auberger P: Implication and regulation of AMPK during physiological
and pathological myeloid differentiation. Int J Mol Sci.
19:E29912018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhao M and Klionsky DJ: AMPK-dependent
phosphorylation of ULK1 induces autophagy. Cell Metab. 13:119–120.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Nieminen AI, Eskelinen VM, Haikala HM,
Tervonen TA, Yan Y, Partanen JI and Klefstrom J: Myc-induced
AMPK-phospho p53 pathway activates Bak to sensitize mitochondrial
apoptosis. Proc Natl Acad Sci USA. 110:E1839–E1848. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Klionsky DJ, Abdelmohsen K, Abe A, Abedin
MJ, Abeliovich H, Acevedo Arozena A, Adachi H, Adams CM, Adams PD,
Adeli K, et al: Guidelines for the use and interpretation of assays
for monitoring autophagy (3rd edition). Autophagy. 12:1–222. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yim NH, Hwang YH, Liang C and Ma JY: A
platycoside-rich fraction from the root of Platycodon grandiflorum
enhances cell death in A549 human lung carcinoma cells via mainly
AMPK/mTOR/AKT signal-mediated autophagy induction. J
Ethnopharmacol. 194:1060–1068. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kim HI, Hong SH, Ku JM, Lim YS, Lee SJ,
Song J, Kim TY, Cheon C and Ko SG: Scutellaria radix promotes
apoptosis in non-small cell lung cancer cells via induction of
AMPK-Dependent autophagy. Am J Chin Med. 47:691–705. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wu Y, Wang X, Guo H, Zhang B, Zhang XB,
Shi ZJ and Yu L: Synthesis and screening of 3-MA derivatives for
autophagy inhibitors. Autophagy. 9:595–603. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yin H, Yang X, Gu W, Liu Y, Li X, Huang X,
Zhu X, Tao Y, Gou X and He W: HMGB1-mediated autophagy attenuates
gemcitabine-induced apoptosis in bladder cancer cells involving JNK
and ERK activation. Oncotarget. 8:71642–71656. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kim J, Kundu M, Viollet B and Guan KL:
AMPK and mTOR regulate autophagy through direct phosphorylation of
Ulk1. Nat Cell Biol. 13:132–141. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xing JJ, Hou JG, Ma ZN, Wang Z, Ren S,
Wang YP, Liu WC, Chen C and Li W: Ginsenoside Rb3 provides
protective effects against cisplatin-induced nephrotoxicity via
regulation of AMPK-/mTOR-mediated autophagy and inhibition of
apoptosis in vitro and in vivo. Cell Prolif. 52:e126272019.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sperling AS, Gibson CJ and Ebert BL: The
genetics of myelodysplastic syndrome: From clonal haematopoiesis to
secondary leukaemia. Nat Rev Cancer. 17:5–19. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Schanz J, Tuchler H, Sole F, Mallo M, Luno
E, Cervera J, Granada I, Hildebrandt B, Slovak ML, Ohyashiki K, et
al: New comprehensive cytogenetic scoring system for primary
myelodysplastic syndromes (MDS) and oligoblastic acute myeloid
leukemia after MDS derived from an international database merge. J
Clin Oncol. 30:820–829. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li F, Zeng J, Gao Y, Guan Z, Ma Z, Shi Q,
Du C, Jia J, Xu S, Wang X, et al: G9a inhibition induces autophagic
cell death via AMPK/mTOR pathway in bladder transitional cell
carcinoma. PLoS One. 10:e01383902015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Mortensen M, Soilleux EJ, Djordjevic G,
Tripp R, Lutteropp M, Sadighi-Akha E, Stranks AJ, Glanville J,
Knight S, Jacobsen SE, et al: The autophagy protein Atg7 is
essential for hematopoietic stem cell maintenance. J Exp Med.
208:455–467. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hu X, Yan R, Cheng X, Song L, Zhang W, Li
K and Zhao S: The function of sperm-associated antigen 6 in
neuronal proliferation and differentiation. J Mol Histol.
47:531–540. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li X, Xu L, Sun G, Wu X, Bai X, Li J,
Strauss JF, Zhang Z and Wang H: Spag6 mutant mice have defects in
development and function of spiral ganglion neurons, apoptosis, and
higher sensitivity to paclitaxel. Sci Rep. 7:86382017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Maiuri MC, Criollo A and Kroemer G:
Crosstalk between apoptosis and autophagy within the Beclin 1
interactome. EMBO J. 29:515–516. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhuang L, Ma Y, Wang Q, Zhang J, Zhu C,
Zhang L and Xu X: Atg3 overexpression enhances bortezomib-induced
cell death in SKM-1 cell. PLoS One. 11:e01587612016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Romano A, Giallongo C, La Cava P,
Parrinello NL, Chiechi A, Vetro C, Tibullo D, Di Raimondo F, Liotta
LA, Espina V and Palumbo GA: Proteomic analysis reveals autophagy
as pro-survival pathway elicited by long-term exposure with
5-azacitidine in high-risk myelodysplasia. Front Pharmacol.
8:2042017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Gao L, Wang Z, Lu D, Huang J, Liu J and
Hong L: Paeonol induces cytoprotective autophagy via blocking the
Akt/mTOR pathway in ovarian cancer cells. Cell Death Dis.
10:6092019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Maiuri MC, Zalckvar E, Kimchi A and
Kroemer G: Self-eating and self-killing: Crosstalk between
autophagy and apoptosis. Nat Rev Mol Cell Biol. 8:741–752. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Martelli AM, Evangelisti C, Chappell W,
Abrams SL, Basecke J, Stivala F, Donia M, Fagone P, Nicoletti F,
Libra M, et al: Targeting the translational apparatus to improve
leukemia therapy: Roles of the PI3K/PTEN/Akt/mTOR pathway.
Leukemia. 25:1064–1079. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zeng J, Liu W, Fan YZ, He DL and Li L:
PrLZ increases prostate cancer docetaxel resistance by inhibiting
LKB1/AMPK-mediated autophagy. Theranostics. 8:109–123. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Li TT, Zhu D, Mou T, Guo Z, Pu JL, Chen
QS, Wei XF and Wu ZJ: IL-37 induces autophagy in hepatocellular
carcinoma cells by inhibiting the PI3K/AKT/mTOR pathway. Mol
Immunol. 87:132–140. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Chan EY, Kir S and Tooze SA: siRNA
screening of the kinome identifies ULK1 as a multidomain modulator
of autophagy. J Biol Chem. 282:25464–25474. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zheng Q, Zhao Y, Guo J, Zhao S, Fei C,
Xiao C, Wu D, Wu L, Li X and Chang C: Iron overload promotes
mitochondrial fragmentation in mesenchymal stromal cells from
myelodysplastic syndrome patients through activation of the
AMPK/MFF/Drp1 pathway. Cell Death Dis. 9:5152018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Basiorka AA, McGraw KL, Eksioglu EA, Chen
X, Johnson J, Zhang L, Zhang Q, Irvine BA, Cluzeau T, Sallman DA,
et al: The NLRP3 inflammasome functions as a driver of the
myelodysplastic syndrome phenotype. Blood. 128:2960–2975. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Bullón P, Alcocer-Gómez E, Carrión AM,
Marín-Aguilar F, Garrido-Maraver J, Román-Malo L, Ruiz-Cabello J,
Culic O, Ryffel B, Apetoh L, et al: AMPK phosphorylation modulates
pain by activation of NLRP3 inflammasome. Antioxid Redox Signal.
24:157–170. 2016. View Article : Google Scholar : PubMed/NCBI
|














