Introduction
Pancreatic cancer is the fifth common malignant
tumor in the gastrointestinal tract, with the incidence rate rising
four times in the last twenty years in China (1).
Furthermore, most patients with pancreatic cancer
are diagnosed with ductal adenocarcinoma, which account for 90% of
pancreatic cancer (1). Due to its
high degree of malignancy, the prognosis of patients with
pancreatic cancer is very poor. The global average annual incidence
is 230,000, with an overall 5-year survival rate of <5%
(1,2). At present, radical resection remains
the best therapeutic option for patients with early stage
pancreatic cancer; however, due to the special anatomy of the
pancreas, early symptoms of pancreatic cancer remain discrete in
most patients and the early diagnosis is therefore difficult
(2). The majority of patients are
diagnosed at an advanced stage and cannot receive the optimal
treatment strategy. The early diagnosis of pancreatic cancer is
therefore essential (2–4).
UBE2T has been recently discovered as an oncogenic
protein that belongs to the family of ubiquitin-conjugating enzymes
(E2) (5). Ubiquitin can be
transferred to E2 enzymes via the ubiquitin-activating enzyme (E1),
and transferred to substrate proteins by ubiquitin ligase (E3)
(5). The protein encoded by the
UBE2T gene consists of 197 amino acids and catalyzes the covalent
attachment of ubiquitin to protein substrates (6). Previous studies reported that detecting
UBE2T expression could directly detect Fanconi anemia (FA) and
interfere with the DNA damage-repair response (5,6). Two
transcript variants encoding different isoforms have been
identified for this gene (6). There
are two isoforms including NP-054895.1 and NP-001297255.1, which
consist of 197 amino acids and 167 amino acids, respectively. Some
of the UBE2T gene-associated pathways are the FA and the ubiquitin
proteasome pathway (6). Data from
Gene Ontology reported that UBE2T gene is associated with
‘chromatin binding’ and ‘ubiquitin-protein transferase activity’
(6) biological processes.
Furthermore, the UBE2T gene participates in the protein
monoubiquitination catalysis and is involved in the mitomycin-C
induced DNA repair (5). In addition,
UBE2T protein acts as a specific E2 enzyme for the FA core complex
by binding to the E3 ligase named FA Complementation Group L
(FANCL) and catalyzing the monoubiquitination of FA Complementation
Group D2, which are crucial steps in DNA damage repair (6). The UBE2T gene is also involved in FANCL
and FA Complementation Group I monoubiquitination (6). It has been reported that UBE2T might
contribute to the ubiquitination and degradation of breast cancer
type 1 susceptibility protein homolog, which serves an important
role in regulating breast cancer cell proliferation and metastasis
(7). Furthermore, the reactome
pathways in which the UBE2T gene is involved include ‘DNA repair’,
‘Fanconi anemia pathway’, ‘Metabolism of proteins’,
‘Post-translational protein modification’, ‘Protein ubiquitination’
and ‘synthesis of active ubiquitin-roles of E1 and E2 enzymes’
(8). Whether UBE2T serves a role in
pancreatic cancer remains unclear.
The present study investigated the role of UBE2T in
pancreatic cancer initiation and progression. The results
demonstrated that UBE2T is significantly upregulated in human
pancreatic cancer tissues compared with normal tissues.
Furthermore, the results demonstrated that high expression of UBE2T
may promote proliferation, invasion and metastasis of pancreatic
cancer cells. In addition, UBE2T may promote epithelial-mesenchymal
transition (EMT). These data suggested that UBE2T may be considered
as an oncogenic protein and a potential therapeutic target in
pancreatic cancer.
Materials and methods
Tissues collection
A total of 121 patients, 76 males and 45 females,
with pancreatic cancer were included in this study, aging from
36–78 years old (average age, 44.7 years). Pancreatic cancer
resection tissues and normal adjacent tissues were collected from
the same patients who underwent surgery at the Department of Organ
Transplantation and Second Hepatobiliary Surgery of Shandong
Provincial Hospital Affiliated to Shandong University between July
2009 and July 2017. The fresh tissues were stored at liquid
nitrogen immediately. The present study was approved by the Ethics
Committee of Shandong Provincial Hospital and all patients provided
signed informed consent.
Immunohistochemistry (IHC)
IHC staining of UBE2T was performed on pancreatic
cancer and normal tissues. The tissue sections were fixed using 10%
formalin and dehydrated with ethanol of gradient concentration (75,
85 and 95% ethanol for 90 min each at room temperature; waterfree
alcohol I and waterfree alcohol II for 30 min each; immersion wax
temperature 58–65°C). Then the tissues sections were embedded with
paraffin at 60°C. The tissues sections were then cut into 5 µm
thickness slides and rehydrated using xylene and ethanol of
gradient concentration. Besides, antigen retrieval was performed
using citrate buffer for 20 min at 90°C. Endogenous peroxidase
activity was inhibited with 3% hydrogen peroxide for 30 min at room
temperature. Tissues were incubated with a primary antibody against
UBE2T (1:200 dilution; cat. no. ab179802; Abcam) for 15 h at 4°C.
Tissue slides were then incubated with secondary antibody (goat
anti-rabbit IgG H&L horseradish peroxidase-conjugated
preadsorbed; 1:500; cat. no. ab7090; Abcam) at room temperature for
1 h. Signal was visualized using 3′-diaminobenzidine that reacts
with HRP-labeled secondary antibody. The chromogenic region was
observed and images were captured using a fluorescence microscope
(Leica Instrument Co., Ltd.; magnification, ×200).
Cell lines and cell culture
The human pancreatic cancer cell lines MIApaca-2 and
Bxpc-3 were purchased from The Cell Bank of Type Culture Collection
of the Chinese Academy of Sciences. The cell line Panc-1 was
donated by Joe Xinzhou, Shanghai. All cells were cultured with
RPMI-1640 medium (MIApaca-2 and Bxpc-3 cells) or Dulbecco's
Modified Eagle Medium (DMEM) (All purchased from Hyclone; GE
Healthcare Life Sciences) for panc-1 cells, supplemented with 10%
fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.) and
1× penicillin (100 IU/ml)/streptomycin (100 µg/ml) and placed at
37°C in a humidified incubator containing 5% CO2.
Cells transfection
All the cells (5×104/ml) were cultured in
6-cm culture dishes without penicillin/streptomycin. Plasmid (2 µg;
purchased from GeneCopoeia, Inc.) transfection was performed by the
lentivirus transfection system, using REV, GAG and VSV package
vectors and Lipofectamine™ 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's instructions,
once cell density reached ~80% for ~48 h. Cells were divided into
four groups, according to the plasmid transfected: Psuper,
psuper-UBE2TD, HA-psuper and HA-UBE2T-psuper. HA tag is a tag
protein that can be fused to target gene and detected using HA
tags. HA-UBE2T is the recombinant plasmid, which contains the HA
tag and UBE2T ORF clone, in order to better observe the
experimental results. The transfection efficiency is presented in
Fig. S1. The following experiments
were performed 48 h after transfection.
Cells were also transfected with shRNAs, the
sequences were as follows: shUBE2T-A,
5′-GATCCCCAGACCAAATGGATGACCTGTTCAAGAGACAGGTCATCCATTTGGTCTTTTTTGGAAA-3′;
shUBE2T-B,
5′-GATCCCCTGCCAGACAGTGGACAGAGTTCAAGAGACTCTGTCCACTGTCTGGCATTTTTGGAAA-3′
(GENEWIZ, Inc.). The following experiments were performed 48 h
after transfection.
Western blotting
Once cell density reached ~80%, cells were lysed
using RIPA lysis buffer (Beyotime Institute of Biotechnology)
supplemented with protease inhibitor cocktail on ice. Cell lysates
were centrifuged at 12,000 × g for 30 min at 4°C following shaking
and sonication every 1 min for three times on ice. Protein
concentration detection was carried out using the BCA assay
(Beijing Solarbio Science & Technology Co., Ltd.) Supernatant
was collected and mixed with 5X loading buffer (contain SDS and
Bromphenol blue) and diluted to a final concentration of 1X sample
buffer (from 5X buffer dilution). The samples were boiled at 95°C
for ~5 min, in order to denature the lysate protein. Proteins (40
µg) were separated by 10% SDS-PAGE gels and transferred onto
polyvinylidene fluoride membranes. Membranes were blocked in 5%
bovine serum albumin (BSA; 5 g/l, purchased from Sigma-Aldrich;
Merck KGaA) for 1 h at room temperature, and incubated with primary
antibody diluted in 5% BSA and against β-actin (1:1,000; cat. no.
ab69512; Abcam), UBE2T (1:1,000; cat. no. ab179802; Abcam),
vimentin (1:1,000; cat. no. 39325; Cell Signaling Technology,
Inc.), E-cadherin (1:1,000; cat. no. 14472; Cell Signaling
Technology, Inc.), N-cadherin (1:1,000; cat. no. 13116; Cell
Signaling Technology, Inc.) and α-catenin (1:50,000; cat. no.
ab64479; Abcam) overnight at 4°C. Membranes were washed three times
with TBST (0.2% Tween) and incubated with secondary antibodies:
(goat anti-rabbit IgG H&L DyLight488 (cat. no. ab96899; Abcam)
and goat anti-mouse IgG H&L DyLight594 (cat. no. ab96881;
Abcam) (all 1:10,000) for 1 h at room temperature. Membranes were
washed three times with TBST and bands were detected using Odessay
Imaging system (supplied by LI-COR Biosciences).
MTT assay
In order to detect the effects of UBE2T on cell
proliferation, transfected pancreatic cancer cells were seeded into
96-well plate at a density of 3,000 cells per well and placed at
37°C in a humidified incubator containing 5% CO2 for 48
h. Six biological replicates were analyzed per group and a blank
well was also set up with only medium. Cells were cultured for five
days until a confluence of 80–90% was reached. Cells were incubated
with MTT reagent (10 µl/well) and placed for 4 h at 37°C in the
dark. Supernatant was removed and purple formazan was dissolved
with 150 µl DMSO. The plate was oscillated for 10 min and the
optical density was measured at 570 nm with a microplate
reader.
Wound-healing assay
All transfected pancreatic cancer cell lines were
cultured in 6-cm culture dishes at a density of
5×104/ml. Cells were scratched using a 200-µl tip,
floating cells were washed with Hank's Balanced Salt Solution
(Thermo Fisher Scientific, Inc.) and new medium (DMEM contain 10%
bovine serum) was added. Cells were incubated for 48 h at 37°C. The
wound width was observed and recorded every 24 h under a microscope
(magnification, ×100). The results were observed using the Zeiss
inverted fluorescence microscope (Zeiss AG).
Cell invasion and migration assay
Transwell (pore size, 8 µm) chambers (BD
Biosciences) were coated with Matrigel and incubated for 30 min at
37°C and were inserted into a 24-well plate. Transfected pancreatic
cancer cells were cultured with RPMI1640 medium or DMEM
supplemented with 1% FBS. Cell suspension (~8×104
cells/200 µl) was added in the upper chamber and 500 µl
corresponding medium supplemented with 10% FBS was added into the
bottom chamber. Cells were cultured in the incubator for 48 h at
37°C The cells on the upper side of the membrane were removed and
the cells that invaded through the membrane were fixed with
methanol for 30 min at room temperature and stained with 1% crystal
violet for 30 min at room temperature Five random fields from each
membrane were counted under a Zeiss inverted fluorescence
microscope at ×200 magnification.
The protocol for migration assay was identical to
the invasion assay; however, Transwell chambers were not coated
with Matrigel.
Statistical analysis
All data are expressed as the means ± standard
deviation. Differences between the two groups were analyzed using a
Student t-test. P<0.05 was considered to indicate a
statistically significant difference.
Results
UBE2T is upregulated in pancreatic
cancer cells and tissues
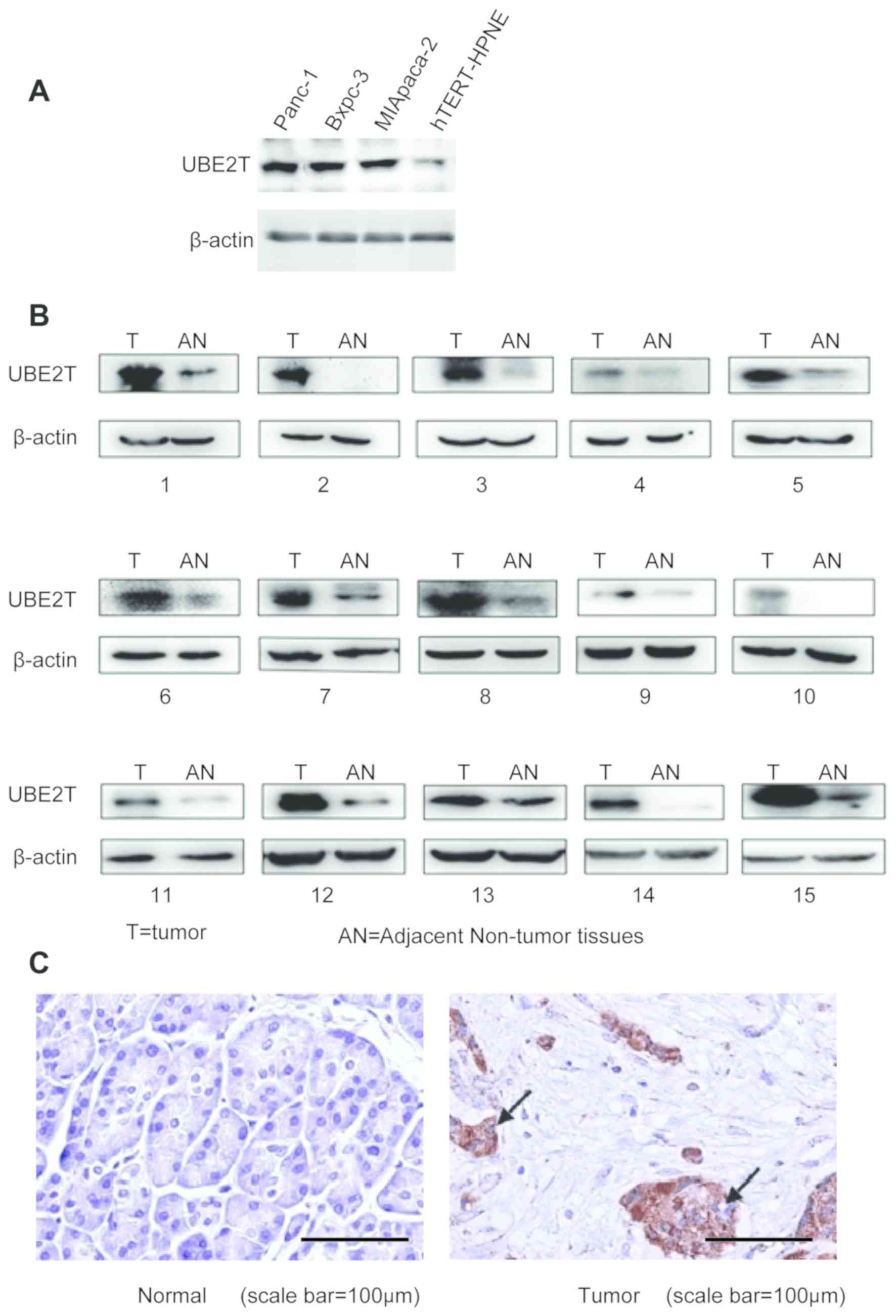
The expression of UBE2T was detected in the three
pancreatic cancer cell lines (MIApaca-2, Bxpc-3 and Panc-1) and the
normal pancreatic cell line (hTERT-HPNE cells). The results from
western blotting demonstrated that UBE2T was upregulated in the
three pancreatic cancer cell lines compared with the normal
pancreatic cell line (Fig. 1A).
Furthermore, the expression of UBE2T was higher in 15 pancreatic
cancer tissues compared with the adjacent normal tissues (Fig. 1B). In addition, results from IHC
revealed that UBE2T expression was increased in pancreatic cancer
tissues compared with adjacent normal tissues (Fig. 1C).
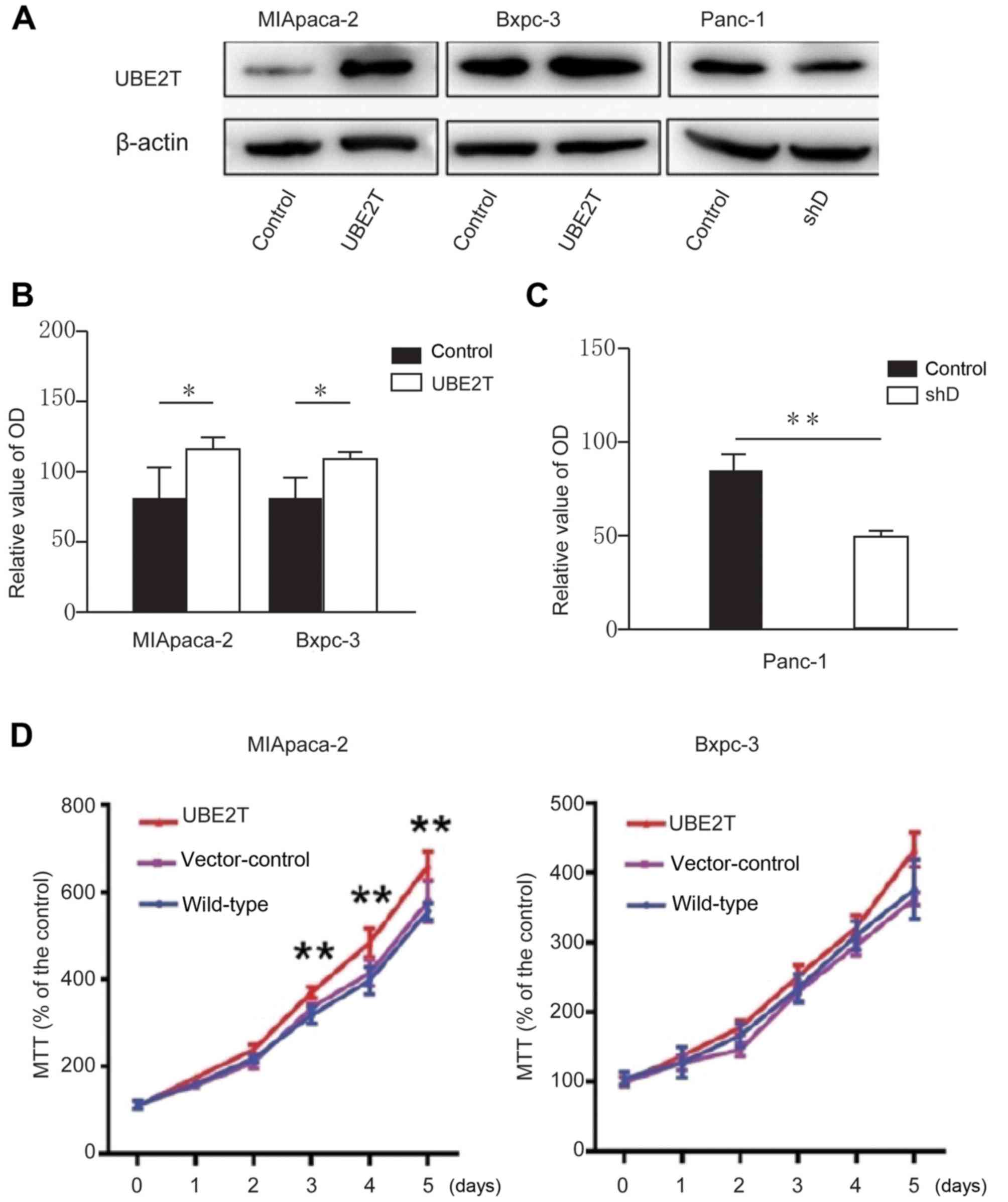
UBE2T promotes pancreatic cancer cell
proliferation
To further explore the role of UBE2T in pancreatic
cancer cells, cells were transfected with psuper, psuper-UBE2T, HA
and HA-UBE2T. The results from western blotting demonstrated that
UBE2T was overexpressed in pancreatic cell lines compared with
control group (Fig. 2A).
Pancreatic cancer cell lines Panc-1, MIApaca-2 and
Bxpc-3 were cultured in a 6-cm dish with penicillin-free medium,
until the confluence was ~80%. The target plasmid and opti-MEM
medium, as well as Lipofectamine™ 2000 and opti-MEM medium, was
added in a ratio of 1:36.5 respectively into two tubes and mixed at
room temperature for 5 min. Subsequently, the two tubes were mixed
and placed on the shelf for 25 min. The mixture in the tube was
added to the petri dish, which was replaced with serum-containing
medium after 6 h. A 10-ml disposable syringe was used to aspirate
the old culture medium, which was filtered with a disposable
filter, and dispensed into a 15-ml disposable centrifuge tube.
The MTT assay was then used to detect the effect of
UBE2T on pancreatic cancer cell proliferation. The results
demonstrated that the proliferation of MIApaca-2 and Bxpc-3 cell
lines overexpressing UBE2T was increased compared with normal cells
(Fig. 2B and D). Conversely, the
proliferation of Panc-1 cells transfected with shRNA-UBE2T was
decreased compared with control cells.
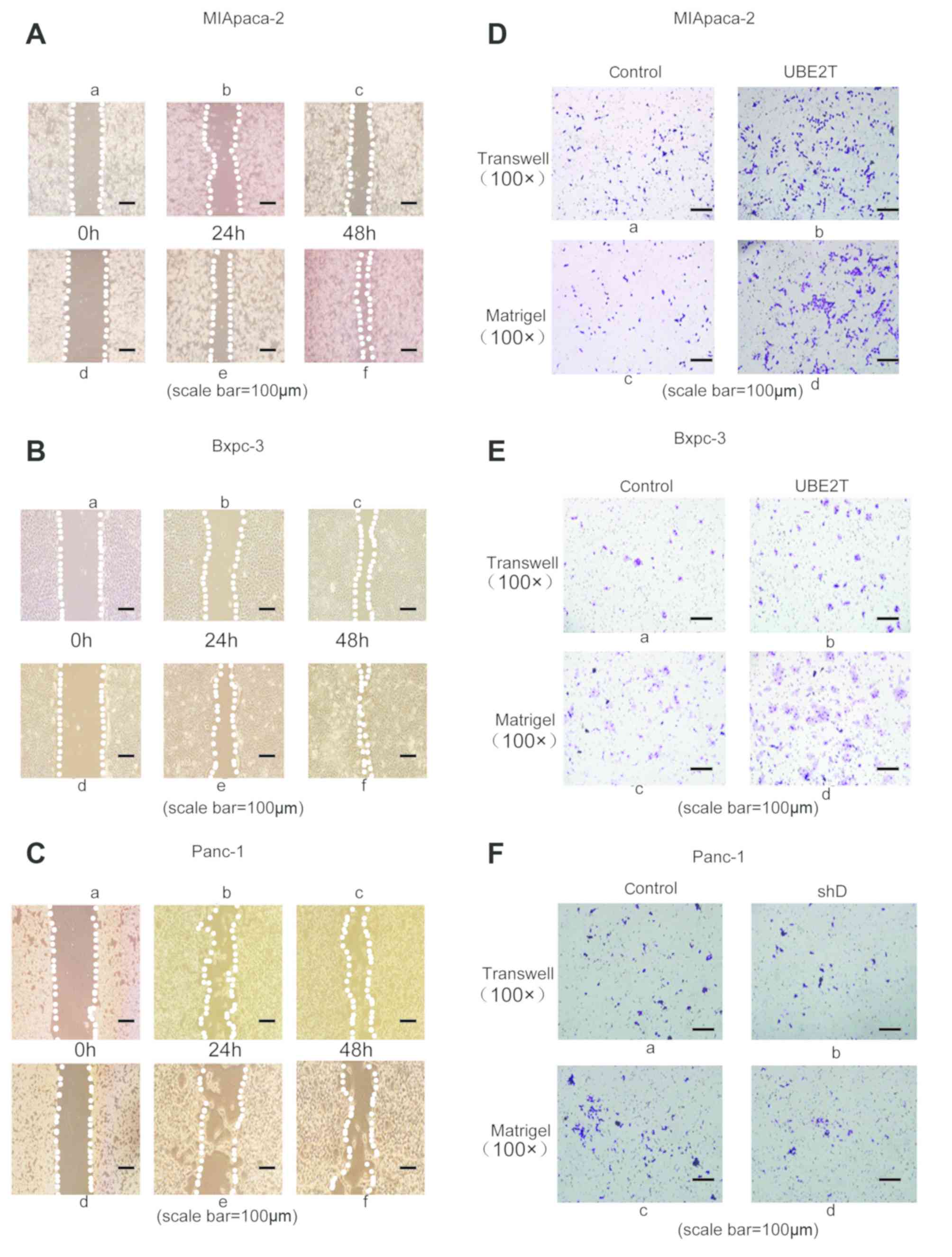
UBE2T promotes pancreatic cancer cell
invasion and migration
The results demonstrated that UBE2T overexpression
could promote pancreatic cell proliferation. Subsequently, wound
healing and Transwell assays were performed to measure the effect
of UBE2T on pancreatic cancer cell invasion and migration.
The results from the wound healing assay
demonstrated that UBE2T overexpression improved the migratory
ability of MIApaca2 (Fig. 3A-d-f)
and Bxpc3 (Fig. 3 B-d-f) cells
compared with corresponding control cells (non-transfected cells)
(Fig. 3A-a-c and B-a-c). However,
the migratory ability of Panc-1 cells transfected with shRNA-UBE2T
(Fig. 3C-d-f) was inhibited compared
with control group (Fig. 3Ca-c). In
order to further confirm these results, Transwell assays were
performed. The results demonstrated that UBE2T overexpression
(Fig. 3D-d; Fig. 3E-d) promoted pancreatic cancer cell
invasion compared with control groups (Fig. 3D-c; Fig.
3E-c). The results demonstrated that UBE2T overexpression
(Fig. 3D-b; Fig. 3E-b) promoted pancreatic cancer cell
migration compared with control groups (Fig. 3D-a; Fig.
3E-a). However, the invasive ability of Panc-1 transfected with
shRNA-UBE2T (Fig. 3F-d) was
inhibited compared with the control group (Fig. 3F-c). Besides, the migration ability
of Panc-1 transfected with shRNA-UBE2T (Fig. 3F-b) was inhibited compared with the
control group (Fig. 3F-a). These
findings indicated that UBE2T may promote pancreatic cancer cell
invasion and migration.
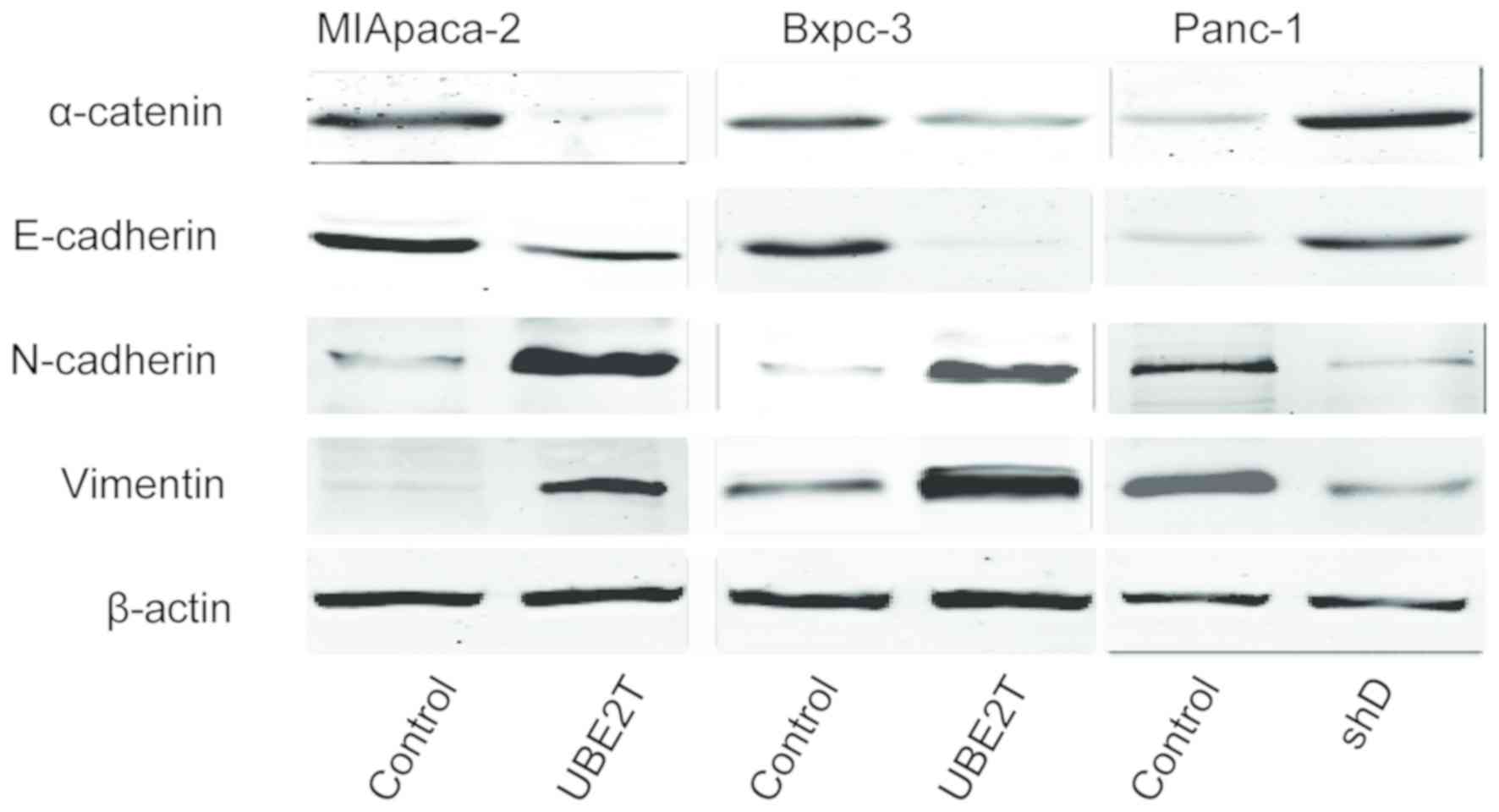
UBE2T promotes EMT in pancreatic
cancer cells
As EMT serves a crucial role in tumor development
(9), the present study investigated
the effect of UBE2T on the expression of epithelial and mesenchymal
marker proteins via western blotting. The results demonstrated that
the expression of the epithelial markers E-cadherin and α-catenin
was decreased whereas the expression of the mesenchymal markers
vimentin and N-cadherin was increased in pancreatic cells
overexpressing UBE2T compared with the control group. Furthermore,
in Panc-1 cells transfected with shRNA-UBE2T, the expression of
mesenchymal markers was decreased whereas the expression of
epithelial markers was increased (Fig.
4).
Discussion
Pancreatic cancer represents a type of solid tumor
that is highly malignant and invasive (7). Furthermore, little is know about the
pathogenesis of pancreatic cancer. It is therefore crucial to
determine some genes that would be associated with the invasive and
metastatic abilities of pancreatic cancer cells and to further
investigate the mechanisms involved in the occurrence and
development of this cancer.
Previous studies reported that UBE2T is highly
expressed in bladder cancer, prostate cancer, gastric cancer and
nasopharyngeal carcinoma (7–9), and that UBE2T is elevated in the
majority of tumors of the digestive system. The present study
demonstrated that UBE2T was upregulated in pancreatic cancer
tissues and that UBE2T overexpression promoted cancer cell
proliferation and invasion in vitro. The underlying
mechanism of UBE2T on the promotion of malignant biological
behavior in pancreatic cancer requires further investigation.
EMT and mesenchymal-epithelial transformation are
considered as key steps in cancer cell metastasis initiation or
endpoint (10). Previous studies on
prostate cancer reported that UBE2T overexpression induces the
upregulation of mesenchymal markers, including vimentin,
fibronectin and α-smooth muscle actin, and the downregulation of
epithelial markers, including E-cadherin (9). It has been reported that UBE2T can
inhibit the degradation of vimentin caused by F-box and WD repeat
domain containing 7 (FBXW7), which is a tumor suppressor (7). The molecular mechanisms underlying the
inhibition of vimentin degradation by UBE2T may be due to the
influence from the interconnection between FBXW7 and vimentin; and
that the 95 amino acid site in UBE2T protein is the binding site
between UBE2T and vimentin (8).
However, to the best of our knowledge, the role of UBE2T in EMT
regulation in pancreatic cancer has not yet been reported. The
results from the present study reported that UBE2T overexpression
could promote EMT in pancreatic cancer cells, which was similar to
findings from previous studies on prostate cancer (8,10).
In conclusion, results from this study suggested
that UBE2T may be considered as a potential therapeutic target in
pancreatic cancer progression. In addition, UBE2T was demonstrated
to promote pancreatic cancer cell invasion, which may be mediated
through EMT regulation and other mechanisms. Although UBE2T may be
considered as a potential diagnostic biomarker and therapeutic
target in pancreatic cancer, further investigation is required to
determine the underlying mechanisms involved in the regulation of
tumor biological behavior by UBE2T.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank Mrs Ling Li (Jinan
Infectious Disease Hospital, Jinan, Shandong) for the help with the
IHC staining assay.
Funding
The present study was supported by the Natural
Foundation of Shandong Province (grant no. ZR2018BH003).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YZ and CL conceived the project and designed the
experiments. PG and MM performed experiments and analyzed data. YC
contributed to the conception and design of the work, as well as to
the analysis and interpretation of data. YZ and CL wrote the
manuscript. All authors accepted the revised version of the
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Shandong Provincial Hospital and all patients provided
signed informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Jemal A, Siegel R, Ward E, Hao Y, Xu J and
Thun MJ: Cancer statistics, 2009. CA Cancer J Clin. 59:225–249.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hidalgo M: Pancreatic cancer. N Engl J
Med. 362:1605–1617. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Witkowski ER, Smith JK and Tseng JF:
Outcomes following resection of pancreatic cancer. J Surg Oncol.
107:97–103. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hassan MM, Bondy ML, Wolff RA, Abbruzzese
JL, Vauthey JN, Pisters PW, Evans DB, Khan R, Chou TH, Lenzi R, et
al: Risk factors for pancreatic cancer: Case-control study. Am J
Gastroenterol. 102:2696–2707. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gong YQ, Peng D, Ning XH, Yang XY, Li XS,
Zhou LQ and Guo YL: UBE2T silencing suppresses proliferation and
induces cell cycle arrest and apoptosis in bladder cancer cells.
Oncol Lett. 12:4485–4492. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wen M, Kwon Y, Wang Y, Mao JH and Wei G:
Elevated expression of UBE2T exhibits oncogenic properties in human
prostate cancer. Oncotarget. 6:25226–25239. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yu H, Xiang P, Pan Q, Huang Y, Xie N and
Zhu W: Ubiquitin-Conjugating enzyme e2t is an independent
prognostic factor and promotes gastric cancer progression. Tumour
Biol. 37:11723–11732. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hu W, Xiao L, Cao C, Hua S and Wu D: UBE2T
promotes nasopharyngeal carcinoma cell proliferation, invasion, and
metastasis by activating the AKT/GSK3β/β-catenin pathway.
Oncotarget. 7:15161–15172. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Rickman KA, Lach FP, Abhyankar A, Donovan
FX, Sanborn EM, Kennedy JA, Sougnez C, Gabriel SB, Elemento O,
Chandrasekharappa SC, et al: Deficiency of UBE2T, the E2 ubiquitin
ligase necessary for FANCD2 and FANCI ubiquitination, causes FA-T
subtype of Fanconi anemia. Cell Rep. 12:35–41. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Brand TM, Iida M, Luthar N, Starr MM,
Huppert EJ and Wheeler DL: Nuclear EGFR as a molecular target in
cancer. Radiother Oncol. 108:370–377. 2013. View Article : Google Scholar : PubMed/NCBI
|