Introduction
Esophageal cancer is one of the most common
malignant tumors and ranks as the sixth leading cause of global
mortalities, and the third leading cause of death in China
(1,2). Esophageal carcinoma mainly includes
esophageal squamous cell carcinoma (ESCC) and esophageal
adenocarcinoma; ESCC is the predominant subtype of esophageal
cancer in developing countries accounting for >90% of all
esophageal cancer subtypes in China (3). In 2018, the global cancer observatory
reported 572,034 new cases of esophageal cancer and 508,585 deaths
from esophageal cancer between males and females combined (2). Despite recent advances in surgical and
therapeutic techniques, such as chemotherapy and radiotherapy,
extensive metastasis has led to a poor 5-year survival rate of
~15-25% (4). Therefore, there is an
urgent need to explore the underlying molecular mechanisms that can
be used to improve the diagnosis and treatment of ESCC.
Vaccinia-related kinase (VRK) is a
member of the serine/threonine kinase family in mammals, which
performs pivotal functions by regulating a variety of cellular and
physiological activities through phosphorylation reactions
(5). VRK1 encodes a protein
of 396 amino acids in length and was originally discovered from a
cDNA library enriched in human fetal-specific liver genes (6). VRK1 is localized to the cell
nuclei and is one of the three subtypes of VRK; it has
exhibited important roles in cell cycle progression,
transcriptional activation, chromosome condensation, DNA repair and
histone modification (5,7). An increasing number of studies have
found that VRK1 expression regulates the proliferation and
survival of cells in normal or malignant tissues (7–9). During
embryonic development of hematopoiesis, the development of the
mouse liver is accompanied with high expression of VRK1. Similarly,
high expression of VRK1 has been demonstrated in regenerated liver
and liver cancer, which suggests that its expression is associated
with the increase of the number of cells in the early hematopoietic
process (10). In addition, VRK1 is
highly expressed in high-proliferating cells, such as those found
in the testis, thymus and fetal liver (6). Notably, a previous study has shown that
VRK1 induces the G1/S transition by promoting the
expression of cyclin D1 (CCND1) at the
G1/S phase (11). VRK1
expression has been shown to be upregulated in several types of
cancer, including glioma, lung carcinomas, hepatocellular
carcinoma, breast carcinomas and head and neck squamous cell
carcinoma (9,12–14).
Barrier-to-autointegration factor 1
(BANF1) is encoded by the BANF1 gene and is a small,
highly conserved DNA-binding protein of 10 kDa in size that is
located in the cytoplasm and nuclei of cells (15). BANF1 serves a crucial role in
mitotic nuclear recombination, regulation of the stability of the
pre-integration complex of retroviruses and in the regulation of
transcriptional function (16).
Margalit et al (15) reported
a linkage of genomic DNA with the nuclear envelope in the
interphase of mitosis through interactions with the nuclear
envelope components (lamin) and BANF1 protein. Previous
studies have also reported that phosphorylation regulates the DNA
binding activity of BANF1 and its subcellular localization and
dimerization (17,18). It is important to note that Ser-4 is
a major phosphorylation site of BANF1 during both the interphase
and the mitotic phase (19). The
phosphorylation of Ser-4 abrogates the interaction of BANF1 with
DNA and reduces its interaction with the LEM domain and thereby
disrupts the connection between the DNA and the nuclear envelope,
which in turn maintains the normal process of the cell cycle
(18). Previous studies (7,20,21) have
shown that VRK1 can catalyze the phosphorylation of
BANF1, which is a high affinity substrate for VRK1
protein kinase (22). Nichols et
al (22) demonstrated that VRK1
regulated the interaction between BANF1 and DNA by phosphorylation
of the N-terminus of BANF1. VRK1 participated directly in the
regulation of the binding of chromatin to membrane proteins and
BANF1 by facilitating the phosphorylation of the latter (7,17).
Results from the aforementioned studies led to the aim of the
present study, which was to investigate the interaction of
VRK1 and BANF1 and its association with the
physiology of ESCC cancer cells.
VRK1 and BANF1 expression levels were
found to be elevated in ESCC tissues compared with the
corresponding levels noted in adjacent non-tumor tissues. In
addition, the expression levels of VRK1 and BANF1
were significantly associated with the clinical characteristics of
patients with esophageal cancer (23). In the present study, the ESCC cell
lines EC109 and EC1 were used to examine the interaction between
VRK1 and BANF1 in ESCC. Small interfering (si) RNA
was utilized to downregulate the expression of VRK1 and the
changes in the expression levels of BANF1 were investigated
in ESCC cells. In addition, changes in proliferation and migration
of ESCC cells were assessed to explore the potential of this
protein in targeted therapy of ESCC. Taken collectively, the
evidence in the present study indicated that VRK1 and
BANF1 may have pivotal roles during ESCC development and
progression, and represent potential targets for novel ESCC
treatments.
Materials and methods
Cell lines and cell culture
The human ESCC cell lines EC109 and EC1 were
purchased from Sangon Biotech Co., Ltd. The cell lines were
cultured and maintained in RPMI-1640 (Sangon Biotech Co., Ltd.)
supplemented with 10% fetal bovine serum (Sangon Biotech Co., Ltd)
at 37°C in the presence of 5% CO2.
Cell transfection
The siRNA sequences targeting VRK1 were
constructed by Guangzhou RiboBio Co., Ltd. A total of three siRNA
sequences were designed against VRK1, and the efficacy of
transfection was assessed using western blot analysis. The
candidate VRK1 siRNA sequences were as follows: VRK1-176
(5′-GCAGUUGGAGAGAUAAUAATT-3′), VRK1-571
(5′-GCAGCUAAGCUUAAGAAUUTT-3′) and VRK1-862
(5′-CCAAUGGCUUACUGGCCAUTT-3′). The negative control siRNA (siNC)
sequence was 5′-UUCUCCGAACGUGUCACGUTT-3′. Downregulation of
VRK1 by siRNA in EC109 and EC1 cells was performed using the
riboFECT™ CP Reagent (Guangzhou RiboBio Co. Ltd.,) according to the
manufacturer's instructions. Cells underwent transfection when in
the logarithmic growth phase. EC109 and EC1 cells were cultured in
6-well plates at a density of 4×105 cells/well at 37°C,
in the presence of 5% CO2. The transfection was
performed at 37°C for 24 h when the confluency of the cells was
~30-50%. The concentration of siRNAs transfected was 50 nM.
Subsequent experimentation was performed 24 h after
transfection.
Western blot analysis
The protein expression levels of VRK1 and
BANF1 were detected by western blotting in ESCC cells
following siRNA transfection. Transfected EC109 and EC1 cells were
collected following transfection which lasted 24 h and washed three
times with ice-cold PBS. The proteins were extracted using RIPA
lysis buffer (Wuhan Boster Biological Technology, Ltd.) with PMSF
(Boster) and the extracts were centrifuged at 4°C, at 12,000 × g
for 15 min. The total protein concentration was determined using
the BCA Protein Assay kit (Boster). Following denaturation of the
proteins by boiling for 10 min, equal amount of protein samples (25
µg) were separated by 10% SDS-PAGE (Boster) and subsequently
transferred to PVDF membranes (Boster). Subsequently, the membranes
were blocked with TBS + Tween 20 (TBST; 0.1% Tween-20) containing
5% non-fat milk (Boster) at room temperature for 1 h and
subsequently incubated with primary antibodies against VRK1
(1:1,000; cat. no. ab211358; Abcam), BANF1 (1:1,000; cat.
no. ab231331; Abcam) and GAPDH (1:1,000; cat. no. ab9485;
Abcam) overnight at 4°C. The membranes were rinsed with TBST 3–5
times and incubated with horseradish peroxidase-conjugated
secondary antibodies (1:1,000; cat. no. bs-40295G-HRP; BIOSS) for 1
h at room temperature. Following an additional rinse with TBST, the
protein bands were visualized using the eECL Western Blot Kit
(Beyotime Institute of Biotechnology) and a UVP gel imaging
scanning analyzer (GENE). Protein expression analysis was performed
using Image J software v1.8.0 (National Institute of Health) to
calculate the relative protein expression. GAPDH was used to
normalize protein expression.
Cell proliferation assay
EC109 and EC1 cell proliferation was examined by the
Cell Counting Kit-8 (CCK-8; Sangon Biotech) assay. A total of
2×103 cells/well were seeded in 96-well plates. In the
following 3 days, 10 µl CCK-8 reagent was added in 100 µl fresh
medium after culturing for 12, 24, 36, 48, 60 and 72 h and the
cells were incubated at 37°C for 2 h. The absorbance was measured
at 450 nm to calculate the number of viable cells. All the assays
were repeated three times.
Flow cytometry and cell cycle
analysis
Cells were harvested by trypsinization without EDTA
and collected by centrifugation at 400 × g in 37°C for 5 min. The
cells were resuspended and washed twice with pre-cooled PBS to
obtain a single-cell suspension. The cells were fixed overnight at
4°C with 500 µl 70% alcohol. The cell suspension concentration was
estimated at 1×106 cells/ml. Cell cycle analysis was
performed using propidium iodide staining following treatment with
RNaseA (cat. no. R1030; Beijing Solarbio Science & Technology
Co., Ltd.) to remove contaminating RNA. The contents of DNA in each
phase (G0/G1, S and G2/M) were
detected using a FACSAria flow cytometer (BD Biosciences) and
ModFit LT software v.4.0 (Verity Software House, Inc.).
Transwell migration assays
The Transwell migration assay was performed to
detect the changes of the migratory ability of ESCC cells following
transfection with siRNAs. The migration assay was performed by a
Transwell system (8.0 µm pore size; 24-well insert). The cells were
seeded at a density of 5×104 cells in FBS-free DMEM
(Sangon Biotech Co., Ltd) to the permeable membrane of the insert
in the upper chamber. The lower chamber was filled with 600 µl DMEM
containing 10% FBS (Sangon Biotech Co., Ltd). Following incubation
for 24 h at 37°C, the cells left on the upper surface of the insert
were carefully removed with a cotton swab and the migrated tumor
cells on the lower surface were stained by crystal violet for cell
number determination.
Statistical analysis
All experimental data were analyzed using the SPSS
software package, v21.0 (IBM Corp.). The GraphPad Prism software
(v6.0; GraphPad Software, Inc.) was used for graph preparation.
Data were analyzed by Student's t-test, χ2 or ANOVA for
multiple groups followed by a Bonferroni's post hoc test. The
results are expressed as the mean ± standard deviation, and
P<0.05 was considered to indicate a statistically significant
difference.
Results
Depletion of VRK1 results in
downregulation of BANF1 expression
A total of three siRNA sequences including VRK1-176,
VRK1-571 and VRK1-862 were specifically designed and synthesized
for transfection into EC109 and EC1 cells, respectively. Following
culture for 24 h, siVRK1-571 exhibited the highest reduction
efficiency of VRK1 protein expression and was selected as
the siRNA to be used in the subsequent experiments (Fig. 1A). Western blot analysis indicated no
significant difference in VRK1 protein expression between
the blank control group and the siNC-transfected group of the EC109
and EC1 cell lines (Fig. 1C).
However, the expression levels of VRK1 in the siVRK1-571
interference group were significantly lower compared with those in
the siNC group, with a decrease of 62.40 and 52.14% in the EC109
and EC1 cell lines, respectively (P<0.05). These results
demonstrated that the expression of VRK1 was downregulated
by siRNA specifically and effectively.
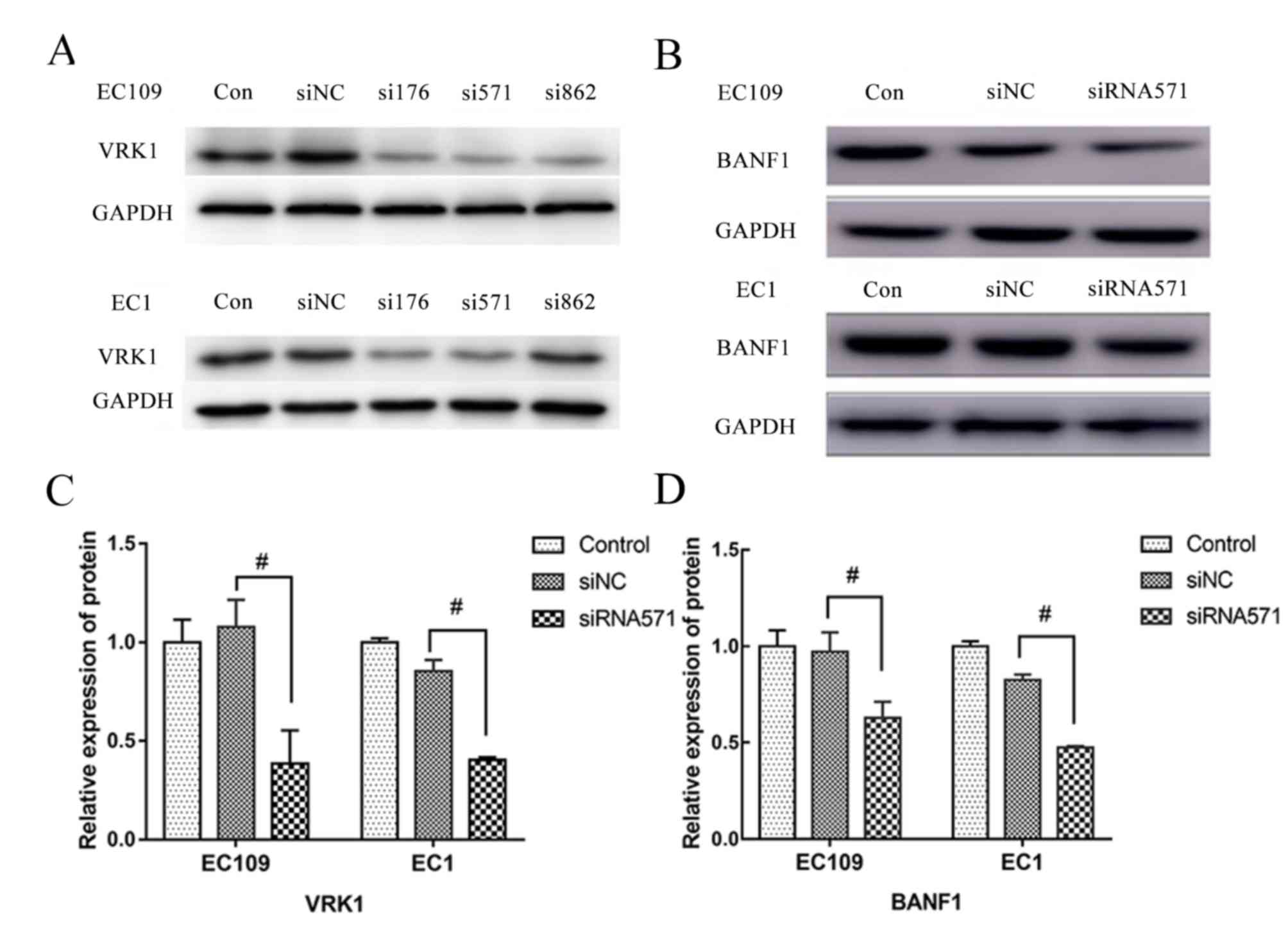 | Figure 1.BANF1 expression is reduced following
inhibition of the expression of VRK1 in ESCC cell lines EC109 and
EC1. (A) Expression of VRK1 following transfection with
gene-specific siRNAs, including VRK1-176, VRK1-571 and VRK1-862 in
ESCC cell lines EC109 and EC1. In order to select a siRNA with
stable transfection effect and eliminate the interference of other
factors, 2 siRNAs (VRK1-176 and VRK1-571) were selected for a
preliminary experiment. As the status of ESCC cells transfected
with siRNA-176 was unstable, VRK1-571 was selected. (B) Expression
of BANF1 was reduced after the expression of VRK1 was inhibited, as
shown by western blot. (C) Semi-quantification analysis of the
expression of VRK1 and BANF1 after transfection with VRK1-siRNA571
from (B). (D)Semi-quantification analysis of the expression of
BANF1 after transfection with VRK1-siRNA571 from (B).
#P<0.05. BANF1, barrier-to-autointegration factor 1;
con, non-transfected control; ESCC, esophageal squamous cell
carcinoma; NC, negative control; siRNA, small interfering RNA;
VRK1, vaccinia-related kinase. |
Furthermore, western blot analysis demonstrated that
the protein expression levels of BANF1 in
siVRK1-571-transfected cells were significantly lower compared with
those of the siNC-transfected control group, with a decrease of
24.51 and 52.87% in the EC109 and EC1 cell lines, respectively
(P<0.05; Fig. 1B and C). In
contrast to these findings, BANF1 protein expression levels
did not reveal a significant difference between the blank and the
negative control groups.
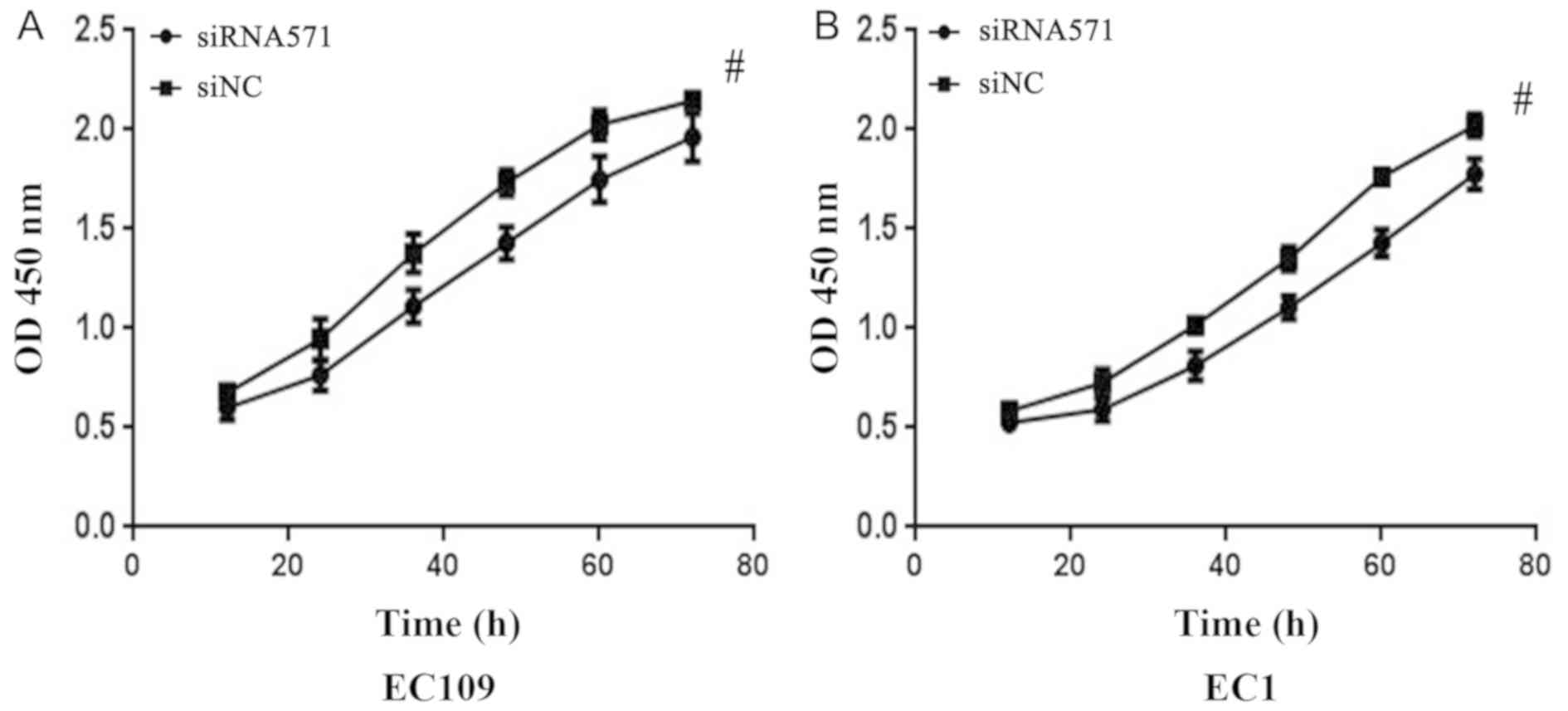
Depletion of VRK1 suppresses the
proliferation of ESCC cells
CCK-8 cell proliferation analysis was conducted on
the EC109 and EC1 cell lines to investigate the effects of
VRK1 on the proliferation of ESCC cells. Following 12 h of
transfection, the cells proliferated less effectively and the
inhibition rate of EC109 and EC1 cells reached the maximum effect
at 48 and 60 h after transfection, respectively (P<0.05;
Fig. 2). The results demonstrated
that downregulation of VRK1 significantly inhibited the
proliferative ability of EC109 and EC1 cells.
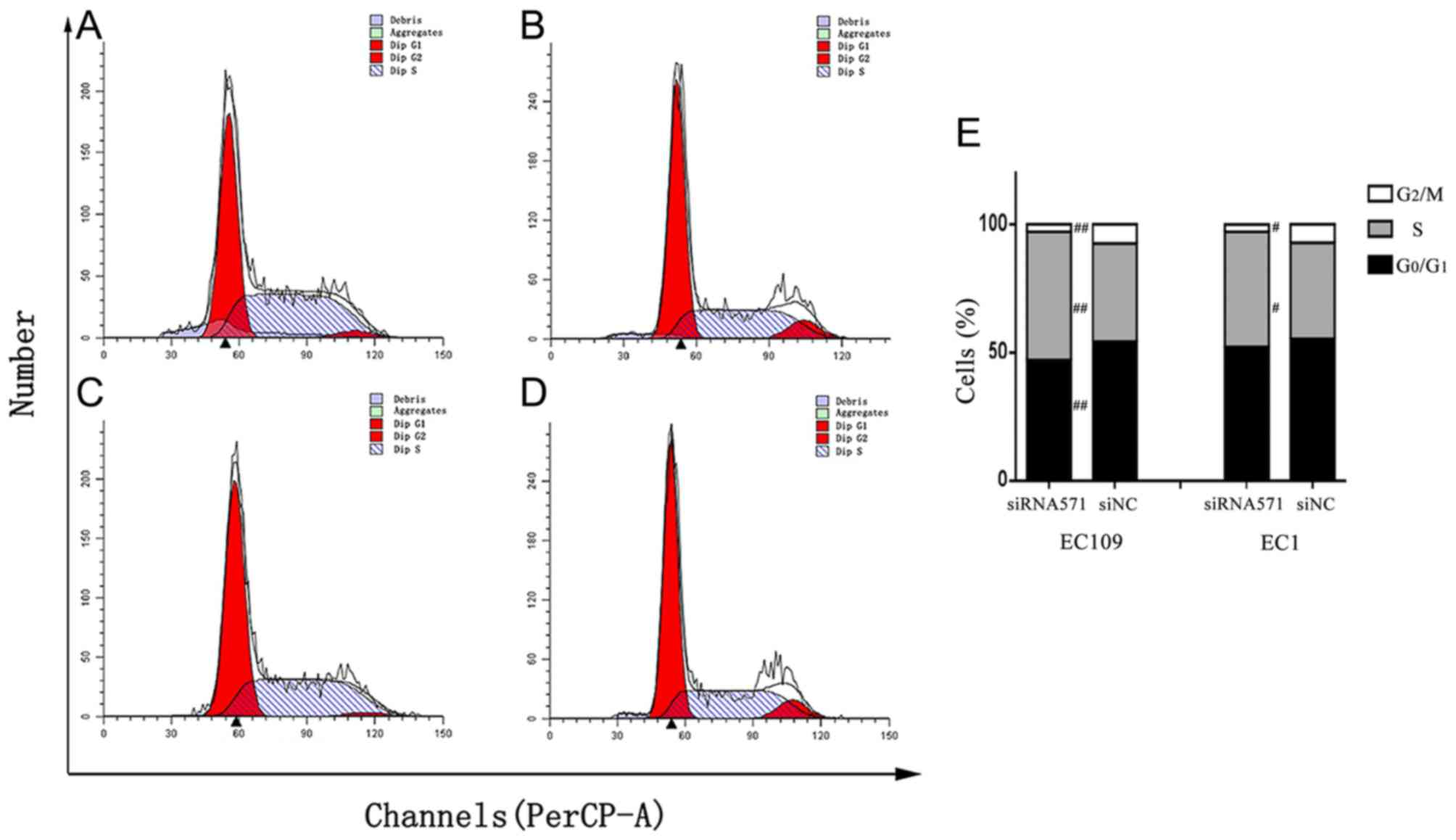
Depletion of VRK1 triggers cell cycle
arrest in ESCC cells
To further verify the effect of VRK1 on ESCC
cell proliferation, VRK1 was depleted and the effects on
cell cycle of ESCC cells were examined using flow cytometry. Flow
cytometric analysis of EC109 cells indicated that depletion of
VRK1 led to a significantly increased population of cells in
the S phase compared with that noted in the negative control group
(P=0.003; Fig. 3). The percentage of
cells treated with siVRK1-571 at the G0/G1
and G2/M phases was significantly lower compared with
that noted in the siNC group (PG0/G1=0.005;
PG2/M=0.001). Similarly, in EC1 cells, the percentage of
cells treated with siVRK1-571 at the G2/M phase was
significantly lower compared with that in the negative control
group (P=0.022). The percentage of cells treated with siVRK1-571 at
the S phase of the cell cycle was higher than that noted in the
negative control group (P=0.023; Fig.
3). The results further demonstrated that depletion of
VRK1 induced cell cycle arrest at the S phase, which in turn
resulted in inhibition of cell proliferation of ESCC cells.
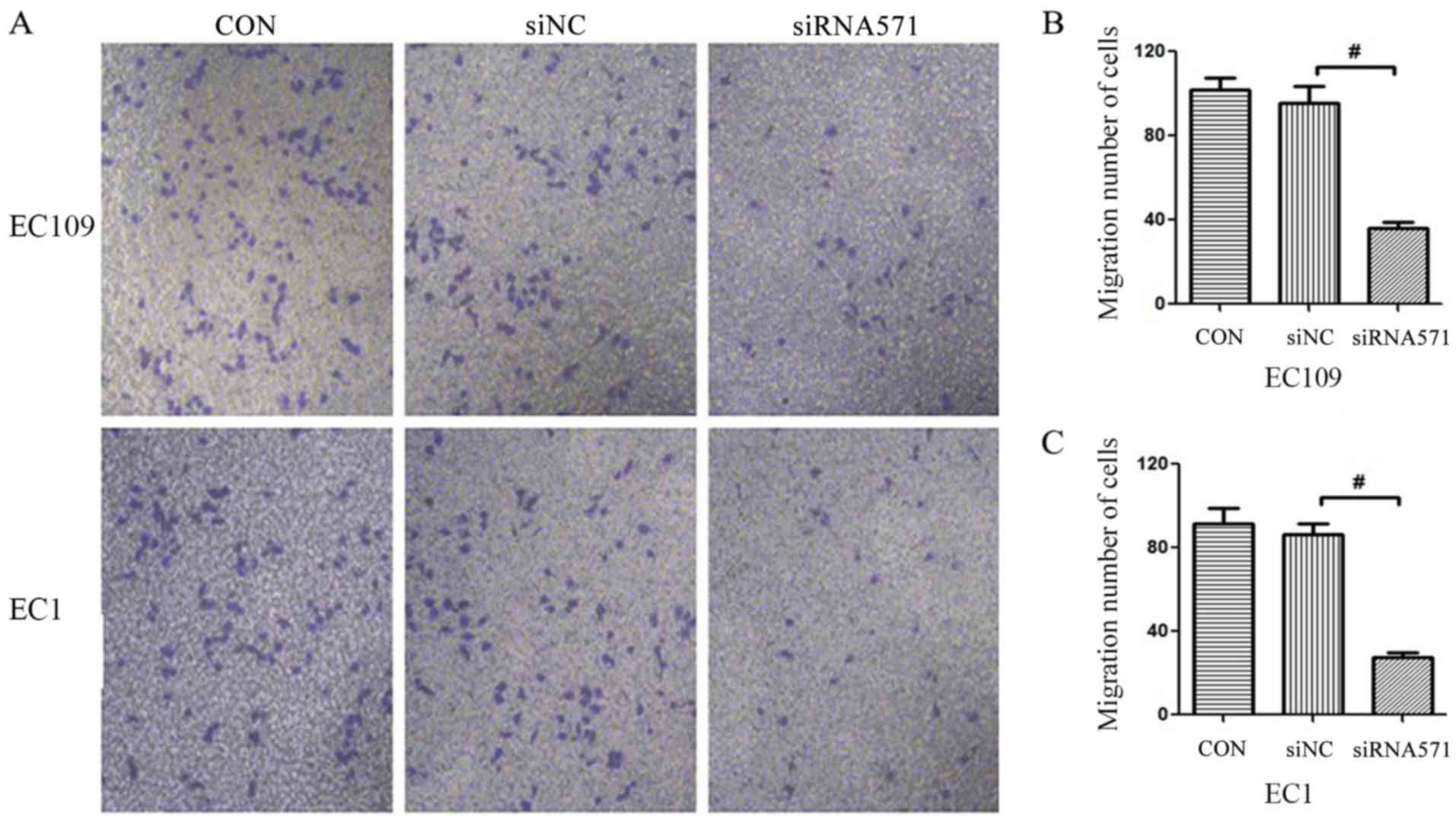
Depletion of VRK1 inhibits ESCC cell
migration
Metastasis is a critical problem during cancer
therapy. Therefore, the Transwell assay was used to assess whether
loss of VRK1 could affect tumor migratory activity. The
results demonstrated that siRNA-based depletion of VRK1
significantly reduced the number of migrating cells compared with
that noted in the blank control (CON) and the negative control
(siNC) groups (Fig. 4). These
results indicated that VRK1 may serve an essential role in
the migration of ESCC.
Discussion
Previous reports suggested that VRK1 has a
pivotal role in the regulation of a variety of cellular
physiological activities; in various cancers, the expression levels
of VRK1 have been significantly associated with cancer
progression and prognosis (9,12,13,24).
In addition, BANF1 has been proven to be a valuable member
in regulating the reassembly of the nuclear envelope and
maintaining appropriate nuclear architecture during mitosis
(15). Certain studies (20,21) have
suggested that BANF1 is an efficient substrate for
VRK1 as VRK1 phosphorylates the N-terminus of
BANF1 (22). These findings
suggested that VRK1 and BANF1 were closely associated
with cell cycle regulation, which led to the hypothesis that the
expression of BANF1 and VRK1 may contribute to the
development of ESCC. This hypothesis was examined in the present
study to provide additional information with regard to the
molecular mechanisms of action during ESCC pathogenesis.
In a previous study conducted by our group, the mRNA
and protein expression levels of VRK1 and BANF1 were
higher in tumor tissues compared with those noted in the adjacent
non-cancerous tissues, as determined by RT-qPCR and
immunohistochemical analyses (23).
Furthermore, the expression levels of VRK1 and BANF1
were associated with the tumor, node, metastasis (TNM) stage and
the differentiation degree of ESCC patients (23). In the present study, VRK1
expression was depleted by siRNA in ESCC cell lines and,
consequently, the expression levels of BANF1 were
significantly downregulated. In addition, in the present study it
was demonstrated that the proliferative activity of EC109 and EC1
cells were significantly reduced following inhibition of
VRK1 expression. Flow cytometric analysis further revealed
that the cell cycle was inhibited mainly at the S phase. Moreover,
Transwell assays indicated that VRK1 depletion significantly
suppressed the migratory ability of ESCC cells. In conclusion,
depletion of VRK1 resulted in the downregulation of
BANF1 and suppressed the proliferation and migration of ESCC
cells.
A growing body of evidence indicates that
VRK1 has an important role in the development of tumors. In
hepatocellular carcinoma, the expression levels of VRK1 in
cancer tissues were higher than those noted in normal and adjacent
tissues (12). In breast cancer,
although high expression levels of VRK1 exhibited a
protective effect on DNA damage, they also led to poorer disease
prognosis (25). Therefore,
VRK1 may represent a potential prognostic indicator for
breast cancer (24,25). Further experiments in mammary
epithelial cells demonstrated that depletion of VRK1
inhibits their proliferation and metastasis in vitro and
in vivo (8). VRK1 has
also been suggested as a proliferative marker in head and neck
squamous cell carcinoma and as a potential drug target in breast
cancer and lung adenocarcinomas (9,14,25). In
addition, VRK1 has an important role in the stress response
of DNA damage induced by ionizing radiation and ultraviolet
radiation (26,27). In the present study, the increased
VRK1 levels in ESCC were consistent with the results noted
in other types of cancer, such as hepatocellular carcinoma and
breast cancer (12,13). Therefore, the data suggested that
VRK1 may contribute to the progression of the ESCC. In the
present study, the CCK-8 cell proliferation assay demonstrated that
the proliferative activity was decreased in EC109 and EC1 cells
following depletion of the expression of VRK1. Moreover,
Transwell experiments indicated that the migratory ability of ESCC
cells was also reduced. Given that the reduction of VRK1
expression could inhibit the proliferation and migration of ESCC
cells, it is hypothesized that VRK1 could represent a potent
new target for the treatment of esophageal cancer.
BANF1 has been shown to bind to
double-stranded DNA at the LEM domain of transcriptional regulators
and at the histone H3 protein, which is localized in the nucleus
(16). Previous findings suggested
that lamin, LEM-domain proteins and BANF1 performed
essential functions in chromatin organization and cell division
(28). Lamin-A/C,
lamin-associated polypeptide 2α and BANF1 proteins
constitute protein complexes that regulate mitotic spindle assembly
and localization during mitosis (29). In addition, BANF1 was directly
involved in the formation of the nuclear envelope (NE). These
results revealed the essential role of BANF1 in regulating
the mitotic process and normal cell cycle progression. Moreover,
BANF1 has been reported to be a novel biomarker for gastric
cancer (30). In one of our previous
studies, it was shown that BANF1 was highly expressed in
ESCC and that its expression was associated with the TNM stage and
the tumor differentiation degree of ESCC patients (23). The data suggested that the highly
expressed levels of BANF1 may lead to an abnormal mitotic
process of ESCC cells through regulation of the interaction of DNA
with perinuclear proteins and the assembly and localization of the
spindle filaments.
Notably, BANF1 has also been established as a
high-affinity substrate of VRK1. In addition, it has been
shown that depletion of VRK1 affects the interaction between
BANF1 and DNA, thereby affecting nuclear membrane structure
and mitotic chromosomal dynamics (31). Together, these studies suggested that
VRK1 and BANF1 were closely associated with the
regulation of normal mitosis. The essential role of VRK1 and
BANF1 in regulating the mitotic nuclear reassembly indicates
that the abnormal expression of these two proteins in ESCC may
affect the normal cellular functions, such as nuclear organization
and cell cycle progression. Based on the high expression of
VRK1 and BANF1 in ESCC, the present study
demonstrated that the BANF1 protein was downregulated
following depletion of the expression of VRK1 in ESCC cells.
These findings indicated a putative interaction between VRK1
and BANF1 in promoting the development of ESCC.
The present study data indicated that the depletion
of VRK1 and the subsequent downregulation of BANF1
resulted in changes in the cell cycle distribution, which mainly
manifested as arrest of the cell cycle at the S phase. The results
indicated that the depletion of VRK1 may affect cell
proliferation by blocking cell cycle progression. These results
support previous studies (9,11,12).
Previous research in head and neck squamous cell carcinoma
suggested that VRK1 plays a role in cell cycle regulation and may
be a new control mechanism of cell cycle, particularly late in G1-S
phase (9). In addition, Valbuena
et al (11) demonstrated that
the elimination of VRK1 by siRNA results in a G1 block in cell
division. As a member of the novel VRK protein family,
VRK1 can phosphorylate the Thr-18 region of p53, a
vital tumor suppressor protein (32). The region that becomes phosphorylated
comprises the MDM-2 binding domain and is required for
maintaining p53 stability (33). VRK1 has been shown to be a key
regulator of p53 and to control cell proliferation. A study
by Waters et al (34) further
revealed that VRK1 promoted germ cell proliferation by
preventing p53 from triggering abnormal cell cycle arrest.
An additional study reported a newly formed autoregulatory loop
between p53 and VRK1 (35). Therefore, VRK1 has been
regarded as an upstream regulator of p53, which participates
in the integration of various cell signals by p53 (36). In addition, VRK1 can
phosphorylate other transcription factors, such as c-Jun and
ATF, which play essential roles in cell cycle regulation
(37,38). Valbuena et al (11) demonstrated that the loss of
VRK1 led to the block of the cell cycle. Similar findings
were presented in the current report highlighting that the
downregulation of VRK1 triggered cell cycle arrest at the
G1 phase. In addition, VRK1 is a regulator of
cyclin D1 (CCND1) expression in the DNA replication
period (39). VRK1 is also
known to phosphorylate histone H3 to regulate chromatin
condensation (40,41). Collectively, these results suggest
that further study of the interaction between VRK1 and
transcription factors such as p53 may be a meaningful direction for
exploring the mechanism of cell cycle regulation in ESCC. However,
further experiments are required to confirm this assumption.
BANF1 was also reported to perform crucial functions in both
the mitotic phase and the cell cycle interphase (42). The reduction or loss of BANF1
expression caused the aberrant cell cycle progression or phenotype.
For example, BANF1-null Drosophila flies present
various cell phenotypes that involve cell cycle arrest, chromatin
clumping, abnormal lamin distribution and nuclear lamina structure
(43). These findings indicated that
the depletion of BANF1 could affect cell cycle progression.
In the present study, BANF1 expression was decreased
following depletion of the expression of VRK1. Therefore, it
was hypothesized that VRK1 may regulate abnormal cell
proliferation by affecting BANF1 expression, which may be a
possible mechanism in the process of esophageal cancer
development.
In conclusion, results from the present study
indicated that downregulation of VRK1 suppressed the
proliferative and migratory ability of ESCC cells in vitro
and suggested that VRK1 may serve as a therapeutic target in
the treatment of ESCC. Furthermore, VRK1 depletion
suppressed BANF1 expression. Taken collectively, the
aforementioned findings suggested a potential connection between
VRK1 and BANF1 in the development of ESCC. The
results presented may be used to further examine the interaction
between VRK1 and BANF1 in the progression of
ESCC.
Acknowledgements
The authors would like to thank Professor Chunfeng
Ren (First Affiliated Hospital of Zhengzhou University) for
providing laboratory support.
Funding
The work was supported by The Henan Provincial
Department of Education Key Science and Technology Project (grant
no. 18A320007) and The Ministry of Health and Welfare Committee
(grant no. SBGJ2018013).
Availability of data and materials
The datasets used and/or analyzed during the
present study are available from the corresponding author on
reasonable request.
Authors' contributions
ZR, JG, CX and XL performed the experiments. ZR
wrote the manuscript. ZR, YL and JL analyzed and interpreted the
data and revised the manuscript for important intellectual content.
HL participated in the design of the research, and was responsible
for the guidance in the experimental process and gave the approval
for the final version of the manuscript. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Enzinger PC and Mayer RJ: Esophageal
cancer. N Engl J Med. 349:2241–2252. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wu PC and Posner MC: The role of surgery
in the management of oesophageal cancer. Lancet Oncol. 4:481–488.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nichols RJ and Traktman P:
Characterization of three paralogous members of the mammalian
vaccinia related kinase family. J Biol Chem. 279:7934–7946. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nezu J, Oku A, Jones MH and Shimane M:
Identification of two novel human putative serine/threonine
kinases, VRK1 and VRK2, with structural similarity to vaccinia
virus B1R kinase. Genomics. 45:327–331. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Valbuena A, Sanz-García M, López-Sánchez
I, Vega FM and Lazo PA: Roles of VRK1 as a new player in the
control of biological processes required for cell division. Cell
Signal. 23:1267–1272. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Molitor TP and Traktman P: Molecular
genetic analysis of VRK1 in mammary epithelial cells: Depletion
slows proliferation in vitro and tumor growth and metastasis in
vivo. Oncogenesis. 2:e482013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Santos CR, Rodríguez-Pinilla M, Vega FM,
Rodríguez-Peralto JL, Blanco S, Sevilla A, Valbuena A, Hernández T,
van Wijnen AJ, Li F, et al: VRK1 signaling pathway in the context
of the proliferation phenotype in head and neck squamous cell
carcinoma. Mol Cancer Res. 4:177–185. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Vega FM, Gonzalo P, Gaspar ML and Lazo PA:
Expression of the VRK (vaccinia-related kinase) gene family of p53
regulators in murine hematopoietic development. FEBS Lett.
544:176–180. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Valbuena A, López-Sánchez I and Lazo PA:
Human VRK1 is an early response gene and its loss causes a block in
cell cycle progression. PLoS One. 3:e16422008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lee N, Kwon JH, Kim YB, Kim SH, Park SJ,
Xu W, Jung HY, Kim KT, Wang HJ and Choi KY: Vaccinia-related kinase
1 promotes hepatocellular carcinoma by controlling the levels of
cell cycle regulators associated with G1/S transition. Oncotarget.
6:30130–30148. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mon AM, MacKinnon AC Jr and Traktman P:
Overexpression of the VRK1 kinase, which is associated with breast
cancer, induces a mesenchymal to epithelial transition in mammary
epithelial cells. PLoS One. 13:e02033972018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kim IJ, Quigley D, To MD, Pham P, Lin K,
Jo B, Jen KY, Raz D, Kim J, Mao JH, et al: Rewiring of human lung
cell lineage and mitotic networks in lung adenocarcinomas. Nat
Commun. 4:17012013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Margalit A, Brachner A, Gotzmann J,
Foisner R and Gruenbaum Y: Barrier-to-autointegration factor-a
BAFfling little protein. Trends Cell Biol. 17:202–208. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Margalit A, Neufeld E, Feinstein N, Wilson
KL, Podbilewicz B and Gruenbaum Y: Barrier to autointegration
factor blocks premature cell fusion and maintains adult muscle
integrity in C. elegans. J Cell Biol. 178:661–673. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gorjanacz M, Klerkx EP, Galy V, Santarella
R, López-Iglesias C, Askjaer P and Mattaj IW: Caenorhabditis
elegans BAF-1 and its kinase VRK-1 participate directly in
post-mitotic nuclear envelope assembly. EMBO J. 26:132–143. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jamin A, Wicklund A and Wiebe MS: Cell-
and virus-mediated regulation of the barrier-to-autointegration
factor's phosphorylation state controls its DNA binding,
dimerization, subcellular localization, and antipox viral activity.
J Virol. 88:5342–5355. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bengtsson L and Wilson KL:
Barrier-to-autointegration factor phosphorylation on ser-4
regulates emerin binding to lamin A in vitro and emerin
localization in vivo. Mol Biol Cell. 17:1154–1163. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lancaster OM, Cullen CF and Ohkura H:
NHK-1 phosphorylates BAF to allow karyosome formation in the
drosophila oocyte nucleus. J Cell Biol. 179:817–824. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kim W, Lyu HN, Kwon HS, Kim YS, Lee KH,
Kim DY, Chakraborty G, Choi KY, Yoon HS and Kim KT: Obtusilactone B
from machilus thunbergii targets barrier-to-autointegration factor
to treat cancer. Mol Pharmacol. 83:367–376. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nichols RJ, Wiebe MS and Traktman P: The
vaccinia-related kinases phosphorylate the N′ terminus of BAF,
regulating its interaction with DNA and its retention in the
nucleus. Mol Biol Cell. 17:2451–2464. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li J, Wang T, Pei L, Jing J, Hu W, Sun T
and Liu H: Expression of VRK1 and the downstream gene BANF1 in
esophageal cancer. Biomed Pharmacother. 89:1086–1091. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Martin KJ, Patrick DR, Bissell MJ and
Fournier MV: Prognostic breast cancer signature identified from 3D
culture model accurately predicts clinical outcome across
independent datasets. PLoS One. 3:e29942008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Salzano M, Vázquez-Cedeira M, Sanz-García
M, Valbuena A, Blanco S, Fernández IF and Lazo PA: Vaccinia-Related
kinase 1 (VRK1) confers resistance to DNA-damaging agents in human
breast cancer by affecting DNA damage response. Oncotarget.
5:1770–1778. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sanz-García M, Monsalve DM, Sevilla A and
Lazo PA: Vaccinia-Related kinase 1 (VRK1) is an upstream
nucleosomal kinase required for the assembly of 53BP1 foci in
response to ionizing radiation-induced DNA damage. J Biol Chem.
287:23757–23768. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
López-Sánchez I, Valbuena A,
Vázquez-Cedeira M, Khadake J, Sanz-García M, Carrillo-Jiménez A and
Lazo PA: VRK1 interacts with p53 forming a basal complex that is
activated by UV-induced DNA damage. FEBS Lett. 588:692–700. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Margalit A, Liu J, Fridkin A, Wilson KL
and Gruenbaum Y: A lamin-dependent pathway that regulates nuclear
organization, cell cycle progression and germ cell development.
Novartis Found Symp. 264:231–240. 2005.PubMed/NCBI
|
|
29
|
Qi R, Xu N, Wang G, Ren H, Li S, Lei J,
Lin Q, Wang L, Gu X, Zhang H, et al: The lamin-A/C-LAP2alpha-BAF1
protein complex regulates mitotic spindle assembly and positioning.
J Cell Sci. 128:2830–2841. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li J, Hu B, Fang L, Gao Y, Shi S, He H,
Liu X and Yuan C: Barrier-To-Autointegration factor 1: A novel
biomarker for gastric cancer. Oncol Lett. 16:6488–6494.
2018.PubMed/NCBI
|
|
31
|
Molitor TP and Traktman P: Depletion of
the protein kinase VRK1 disrupts nuclear envelope morphology and
leads to BAF retention on mitotic chromosomes. Mol Biol Cell.
25:891–903. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Reinhardt HC and Schumacher B: The p53
network: Cellular and systemic DNA damage responses in aging and
cancer. Trends Genet. 28:128–136. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Barcia R, López-Borges S, Vega FM and Lazo
PA: Kinetic properties of p53 phosphorylation by the human
vaccinia-related kinase 1. Arch Biochem Biophys. 399:1–5. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Waters K, Yang AZ and Reinke V:
Genome-Wide analysis of germ cell proliferation in C.elegans
identifies VRK-1 as a key regulator of CEP-1/p53. Dev Biol.
344:1011–1025. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Valbuena A, Vega FM, Blanco S and Lazo PA:
P53 downregulates its activating vaccinia-related kinase 1, forming
a new autoregulatory loop. Mol Cell Biol. 26:4782–4793. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lopez-Borges S and Lazo PA: The human
vaccinia-related kinase 1 (VRK1) phosphorylates threonine-18 within
the mdm-2 binding site of the p53 tumour suppressor protein.
Oncogene. 19:3656–3664. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sevilla A, Santos CR, Barcia R, Vega FM
and Lazo PA: C-Jun phosphorylation by the human vaccinia-related
kinase 1 (VRK1) and its cooperation with the N-terminal kinase of
c-Jun (JNK). Oncogene. 23:8950–8958. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sevilla A, Santos CR, Vega FM and Lazo PA:
Human vaccinia-related kinase 1 (VRK1) activates the ATF2
transcriptional activity by novel phosphorylation on Thr-73 and
Ser-62 and cooperates with JNK. J Biol Chem. 279:27458–27465. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kang TH, Park DY, Kim W and Kim KT: VRK1
phosphorylates CREB and mediates CCND1 expression. J Cell Sci.
121:3035–3041. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Shin J, Chakraborty G, Bharatham N, Kang
C, Tochio N, Koshiba S, Kigawa T, Kim W, Kim KT and Yoon HS: NMR
solution structure of human vaccinia-related kinase 1 (VRK1)
reveals the C-terminal tail essential for its structural stability
and autocatalytic activity. J Biol Chem. 286:22131–22138. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kim W, Chakraborty G, Kim S, Shin J, Park
CH, Jeong MW, Bharatham N, Yoon HS and Kim KT: Macro histone H2A1.2
(macroH2A1) protein suppresses mitotic kinase VRK1 during
interphase. J Biol Chem. 287:5278–5289. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Cox JL, Mallanna SK, Ormsbee BD, Desler M,
Wiebe MS and Rizzino A: Banf1 is required to maintain the
self-renewal of both mouse and human embryonic stem cells. J Cell
Sci. 124:2654–2665. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Furukawa K, Sugiyama S, Osouda S, Goto H,
Inagaki M, Horigome T, Omata S, McConnell M, Fisher PA and Nishida
Y: Barrier-to-autointegration factor plays crucial roles in cell
cycle progression and nuclear organization in drosophila. J Cell
Sci. 116:3811–3823. 2003. View Article : Google Scholar : PubMed/NCBI
|