Introduction
Breast cancer is the most frequently diagnosed
cancer and the leading cause of cancer-associated death among women
worldwide (1). In 2018, the
International Agency for Research on Cancer estimated a disease
incidence rate of 24.2% and mortality rate of 15.0% in women across
185 countries (1). Breast cancer is
divided into four subtypes according to the 2015 St. Gallen
consensus (2): Luminal A-like,
luminal B-like, human epidermal growth factor receptor 2
(HER2)-positive and triple-negative (TN) subtypes, which are based
on estrogen receptor (ER), progesterone receptor (PR), Ki-67 and
HER2 immunohistochemical status (2).
ERα is a nuclear transcription factor encoded by the estrogen
receptor 1 (ESR1) gene and activated by estrogen (3). ERα has different effects in normal
breast epithelial cells and breast cancer cells, and it serves a
predictive role in the response to endocrine therapies (4). Estrogen promotes cell proliferation and
breast cancer development in an ER-dependent manner (5); in turn, ERα promotes breast cancer
tumorigenesis and progression (6,7).
Therefore, estrogens serve an essential role in regulating breast
cancer cell proliferation, and estrogen-activated ERα is a crucial
factor for breast cancer development and therapy.
Anterior gradient protein 3 (AGR3) is a member of
the protein disulfide isomerase (PDI) gene family, which consists
of 21 members (http://www.genenames.org/cgi-bin/genefamilies/set/692),
and AGR3 also has two AGR subfamilies, AGR1 and AGR2 (8,9). AGR2
has been widely investigated in breast cancer and is known to
participate in numerous aspects of its development and therapy,
including cell proliferation and migration (10,11).
Although AGR3 and AGR2 are highly similar homologous genes, the
function of AGR3 in cancer may not be the same as that of the
metastasis-associated AGR2 (12).
AGR3 is upregulated in serous borderline ovarian tumor compared
with serous ovarian carcinoma, and high levels of AGR3 predict a
longer survival time in patients with serous ovarian carcinoma
(13). In prostate cancer cells,
AGR3 is upregulated by androgens and estrogen in an androgen
receptor dependent manner (14).
Additionally, AGR3 is highly expressed in intrahepatic
cholangiocarcinoma compared with its expression levels in
hepatocellular carcinoma (15). In
breast cancer, AGR3 is positively associated with low histological
grade breast tumors (16). Recent
studies have demonstrated that extracellular AGR3 can regulate
breast cancer cell migration via Src signaling (17), and that AGR3 can promote the
proliferative and invasive abilities of breast cancer cells, as
well as chemotherapy response (18).
Although differential expression of AGR3 has been
identified among different types of cancer, including ovarian,
prostate, liver and breast cancers (13–16), the
role of AGR3 in breast cancer oncogenesis and development remains
unclear. The present study aimed to investigate the association
between AGR3 and ER status, and the function of AGR3 in ER-positive
breast cancer. It was hypothesized that AGR3 may promote breast
cancer development in an ER-dependent manner, and AGR3 may serve as
a potential therapeutic target for patients with ER-positive breast
cancer.
Materials and methods
Tissue sample collection
A total of 72 breast tumor and paired adjacent
normal tissue samples were collected from 72 patients with breast
cancer (age range, 30–74 years; median age, 46 years) at The First
Affiliated Hospital of Chongqing Medical University (Chongqing,
China) between July 2017 and October 2017. Female patients with
primary breast cancer, normal cardiopulmonary function and
willingness to undergo breast surgery were included, while patients
with secondary breast cancer, intolerable or unwilling to undergo
breast surgery, and male patients with breast cancer were excluded
from the present study. All patients signed informed consent for
the retention and analysis of their tissues for research purposes.
Breast tumor and paired normal tissue samples (>3 cm from the
primary tumor boundary) were collected after surgery and
immediately transferred to liquid nitrogen for fast-frozen storage
and long-term storage at −80°C. The collection of tumor samples and
patient information was approved by the Ethics Committee of The
First Affiliated Hospital of Chongqing Medical University (approval
no. 2017-16).
Cell culture and drug treatment
MCF7, T47D, ZR75-1, BT549, MDA-MB-231, MDA-MB-468
and SK-BR-3 cell lines were obtained from the American Type Culture
Collection. T47D and ZR75-1 cells were cultured at 37°C with 5%
CO2 in RPMI-1640 medium, whereas the other cell lines
were grown in DMEM, both RPMI-1640 and DMEM medium were
supplemented with 10% FBS (all Gibco; Thermo Fisher Scientific,
Inc.). Before adding estrogen (MedChemExpress, Inc.), cells in the
logarithmic growth phase were seeded in 6-well plates at a
confluency of 60–70% and were serum-starved for 24 h in DMEM. RNA
was extracted from estrogen-treated cells at different time points
(0, 1, 3, 6, 12 and 24 h). For 4-hydroxytamoxifen (4-OH Tam;
MedChemExpress, Inc.) treatment, cells in the logarithmic growth
phase were seeded in 6-well plates at a confluency of 60–70% and
cultured in DMEM supplemented with 10% FBS. Different
concentrations of 4-OH Tam (0, 2, 4, 6, 8 and 10 µmol) were added,
and RNA and proteins were extracted after 24 h for RT-qPCR and
western blot analysis.
siRNA transfection
The siRNAs sequences used to target AGR3 and ESR1
(Shanghai GenePharma Co., Ltd.) were as follows: AGR3-siRNA1
forward, 5′-CAGAUUGUACACAUAUGAG-3′ and reverse,
5′-CUCAUAUGUGUACAAUCUG-3′; AGR3-siRNA2 forward,
5′-AGUUCAUCAUGCUAAACCU-3′ and reverse, 5′-AGGUUUAGCAUGAUGAACU-3′;
ESR1-siRNA1 forward, 5′-GGAGGAUGUUGAAACACAATT-3′ and reverse,
5′-UUGUGUUUCAACAUUCUCCTT-3′; and ESR1-siRNA2 forward,
5′-GGAUUUGACCCUCCAUGAUTT-3′ and reverse,
5′-AUCAUGGAGGGUCAAAUCCTT-3′. T47D cells (at a confluency of 60–70%)
were transfected with siRNAs (100 nM) using
Lipofectamine® RNAiMAX reagent (Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. The
culture medium was changed after 4–6 h, and knockdown efficiencies
were determined by RT-qPCR after 24 h.
RNA extraction and qPCR analysis
Total RNA was extracted from breast cancer tissues
and cells using TRIzol reagent (Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol, and the RNA concentration
was measured using a NanoDrop ND-2000 (Thermo Fisher Scientific,
Inc.). cDNA was synthesized from 1 µg total RNA with a PrimeScript
II cDNA Synthesis kit (37°C for 15 min, 85°C for 5 sec, 4°C for
short preservation (usually 30 min prior to qPCR); Takara Bio,
Inc.). Targeted genes were detected with a SYBR Premix Ex Taq II
qPCR kit (Takara Bio, Inc.) using a primer mixture, and cDNA was
amplified by CFX96 fluorescence qPCR (thermocycling conditions:
Initial denaturation at 95°C for 2 min; 40 of cycles of
denaturation at 95°C for 30 sec, annealing at 58°C for 30 sec,
elongation at 72°C for 20 sec; and a final extension at 65°C for 5
sec; Bio-Rad Laboratories, Inc.). Gene-specific primer sequences
were designed as follows: AGR3 forward, 5′-AGAGGCCTCCTCAGACACTC-3′
and reverse, 5′-GGCACATATTGCCCATCAGGT-3′; ESR1 forward,
5′-GGTCAGTGCCTTGTTGGATG-3′ and reverse, 5′-CAGGTTGGTGAGTAAGC-3′;
GAPDH forward, 5′-TTCCAGGAGCGAGATCCCT-3′ and reverse,
5′-GGCTGTTGTCATACCTTCTCATGG-3′; Growth regulation by estrogen in
breast cancer 1 (GREB1) forward, 5′-AAATCGAGGATGTGGAGTG-3′ and
reverse, 5′-TCTCACCAAGCAGGAGGA-3′. mRNA abundance was calculated
according to the 2−ΔΔCq method (19), relative to the internal control
GAPDH.
Protein extraction and western blot
analysis
Total protein from cell lines was extracted on ice
using RIPA lysis buffer (Thermo Fisher Scientific, Inc.). Protein
concentration was quantified using a Pierce™ BCA Protein assay kit
(Thermo Fisher Scientific, Inc.), mixed with 5X SDS-PAGE loading
buffer and double-distilled H2O, boiled at 95°C for 10
min and stored at −20°C. Proteins (30 ug/lane) were separated by
12% SDS-PAGE and transferred onto 0.2 µm immobilon-PSQ
polyvinylidene difluoride membranes (EMD Millipore). The membranes
were blocked with 5% skimmed powdered milk for 1 h at room
temperature and incubated overnight at 4°C with the following
primary antibodies: Antibodies against GAPDH (1:1,000; cat. no.
10494-1-AP) and ER (1:1,000; cat. no. 21244-1-AP) were purchased
from Wuhan Sanying Biotechnology, and the antibody against AGR3
(1:800; cat. no. AP9424b) was purchased from Abgent Biotech Co.,
Ltd. After rinsing three times with Tris-buffered saline containing
0.1% Tween-20 (TBST) for 5 min, the membranes were incubated with
HRP-conjugated anti-rabbit IgG (cat. no. NEF812001EA) or anti-mouse
IgG (cat. no. NEF822001EA) secondary antibodies (both at a dilution
ratio of 1:3,000 and from PerkinElmer, Inc.) for 90 min at room
temperature, rinsed 3 times with TBST for 5 min and soaked in an
enhanced chemiluminescence detection reagent (Bio-Rad Laboratories,
Inc.) for 1 min. The membranes were developed using a GeneGnome
Chemiluminescence Imaging system (Syngene) to automatically detect
protein levels, which were quantified according to the gray value
and normalized to that of GAPDH using ImageJ software (version
1.8.0; National Institutes of Health).
Cell Counting Kit-8 (CCK-8) cell
viability test
Cells were seeded into 96-well plates at a density
of 5×103 cells/well following siRNA transfection, or
following the addition of different concentrations of 4-OH Tam (0,
1, 2, 3, 4, 5, 6, 7, 8, 9 and 10 µmol) at 37°C for 24 h. CCK-8
solution (MedChemExpress, Inc.) was used according to the
manufacturer's protocol. The cells were incubated for 2 h at 37°C.
The mixture of CCK-8 solution and DMEM supplemented with 10% FBS
was used as a blank control, and absorbance was measured at 450 nm.
The experiment was repeated in triplicate.
Gene Expression Omnibus (GEO) and Gene
Set Enrichment Analysis (GSEA)
Datasets from patients with breast cancer were
obtained from the GEO data repository (https://www.ncbi.nlm.nih.gov/geo) and The Cancer
Genome Atlas (TCGA) database (https://portal.gdc.cancer.gov/). The GEO datasets used
in the current study were as follows: Richardson (GSE5460; n=129)
(20), Borresen-Dale (GSE19783;
n=115) (21) and Wang (GSE19615;
n=115) (22). Series matrix files
were downloaded from the online GEO datasets and used for
correlation analyses. GSEA was used to explore in which pathway
AGR3 may be involved. The dataset TCGA-BRCA was downloaded from
TCGA using the GDC Data Transfer Tool (23), processed and analyzed using R2
software (http://r2.amc.nl) and normalized with
the ‘DESeq2’ package (24),
according to the variance stabilizing transformation method
(25). A total of 186 Kyoto
Encyclopedia of Genes and Genomes (KEGG) gene sets from the
Molecular Signatures Database (MSigDB; http://www.broadinstitute.org/gsea/msigdb/) were used
in GSEA. GSEA requires ≥2 different phenotypes. TCGA data for 1,096
patients with breast cancer were divided into 2 groups according to
AGR3 expression. The top 10% (110) of the data with the highest
AGR3 expression were defined as the high expression group, and the
bottom 10% (110) of the date with the lowest AGR3 expression were
defined as the low expression group. The R script of GSEA from the
Broad Institute (https://www.gsea-msigdb.org/gsea) was used for the
GSEA analysis of the two groups.
Statistical analysis
All the statistical analyses were performed using
GraphPad Prism 7 (GraphPad Software, Inc.) and R 3.4.0 (https://www.r-project.org), for GSEA analysis. The
statistical significance of GSEA analysis was determined by
unpaired Student's t-test. The statistical significance of paired
groups was determined by Wilcoxon matched-pairs signed-rank test.
The statistical significance of unpaired two groups was determined
by Mann-Whitney test. Pearson's correlation analysis was performed
to analyze the correlation of target genes. Multiple groups were
statistically compared using one-way ANOVA and Dunnett's multiple
comparisons test. The results are presented as the mean ± standard
deviation (n≥3). P<0.05 was considered to indicate a
statistically significant difference.
Results
AGR3 expression is upregulated in
luminal breast cancer tissues and is positively correlated with
ESR1 expression
To investigate AGR3 expression in breast cancer,
AGR3 expression was analyzed in ER-positive and ER-negative paired
cancer tissues and adjacent normal tissues by RT-qPCR. AGR3
expression was significantly upregulated in luminal A breast cancer
tissues compared with that in their paired adjacent normal tissues
(n=21; Tumor, 0.3335±0.0719 vs. normal, 0.2704±0.1231; P=0.0421;
Fig. 1A), and was visibly increased
in luminal B breast cancer tissues compared with normal tissues
(n=23; Tumor, 0.3344±0.2599 vs. normal, 0.0279±0.0084; P=0.0605;
Fig. 1B). Among the 44 pairs of
ER-positive breast cancer and adjacent normal tissues, the RNA
levels of AGR3 were significantly upregulated in breast cancer
tissues compared with that in paired normal tissues (n=44; Tumor,
0.3340±0.1386 vs. normal, 0.1436±0.0610; P=0.0016; Fig. 1C). Additionally, AGR3 RNA expression
was detected in patients with TN breast cancer and was
significantly downregulated in ER-negative TN breast cancer tissues
compared with that in normal adjacent tissues (n=28; Tumor,
0.0017±0.0006 vs. normal, 0.1197±0.0514; P<0.0001; Fig. 1D).
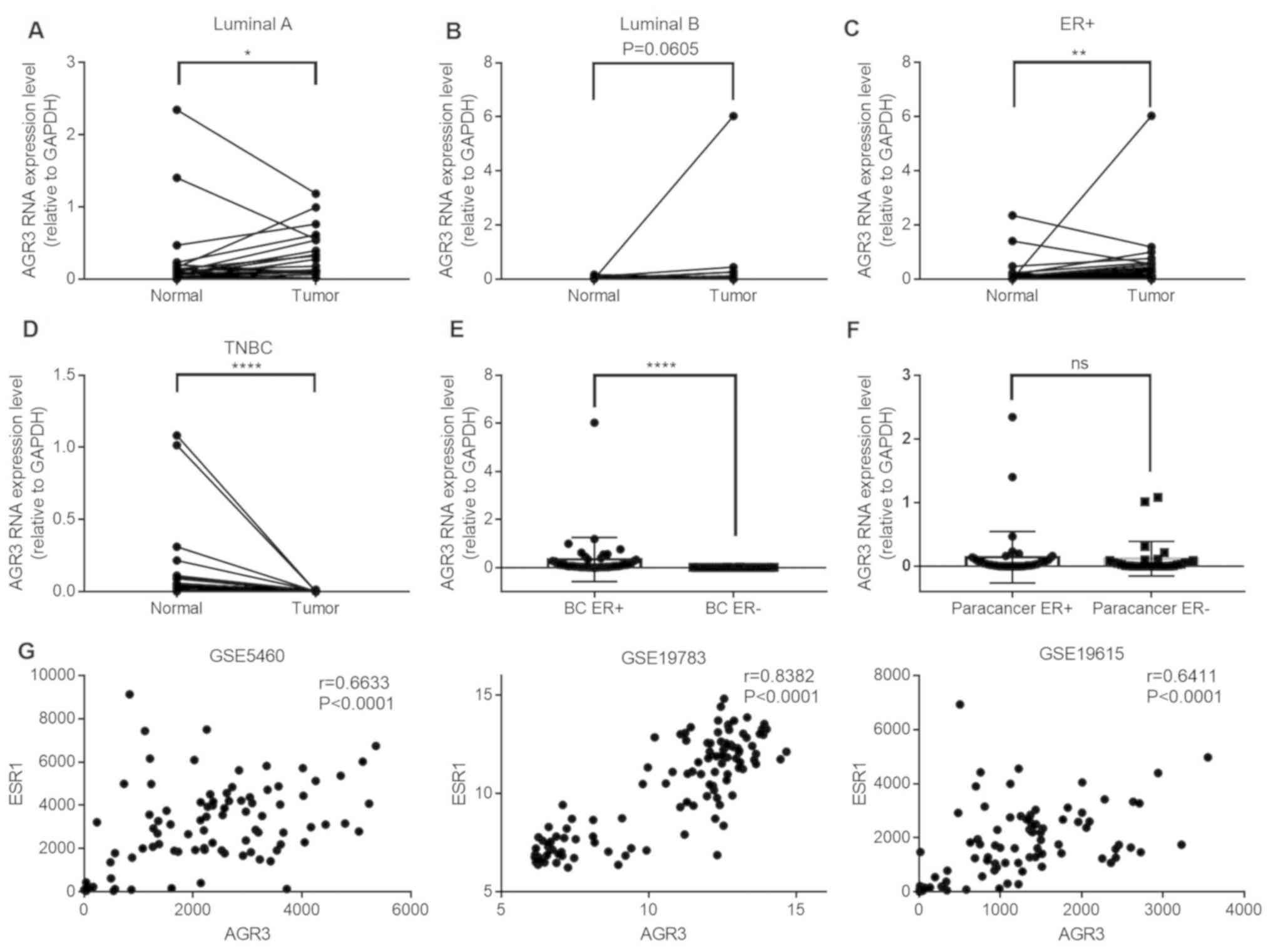 | Figure 1.AGR3 expression is upregulated in
ER-positive breast cancer and positively correlates with ESR1
expression. (A-C) AGR3 RNA expression was detected in (A) luminal
A, (B) luminal B and (C) ER-positive breast cancer tissues and
their paired normal tissues, and compared using the Wilcoxon
matched-pairs signed-rank test. (D) AGR3 RNA expression was
detected in TN breast cancer tissues and their paired normal
tissues, and compared using the Wilcoxon matched-pairs signed-rank
test. (E) AGR3 RNA expression was compared between ER-positive and
ER-negative breast cancer tissues using the Mann-Whitney test. (F)
AGR3 RNA expression was compared between ER-positive and
ER-negative adjacent normal tissues using the Mann-Whitney test.
(G) AGR3 and ESR1 expression values were extracted from GEO dataset
series matrix files (GSE5460, GSE19783 and GSE19615), and
correlation analyses of AGR3 and ESR1 from each GEO expression
dataset were conducted using the Pearson correlation coefficient.
*P<0.05, **P<0.01, ****P<0.0001. AGR3, anterior gradient
3; ER, estrogen receptor; ESR1, estrogen receptor 1; TNBC,
triple-negative breast cancer; ns, not significant; GEO, Gene
Expression Omnibus. |
AGR3 expression between ER-positive and ER-negative
breast cancer tissues, as well as between their paired ER-positive
and ER-negative adjacent normal tissues was analyzed. AGR3 RNA
levels were significantly upregulated in ER-positive (n=44)
compared with in ER-negative (n=28) breast cancer tissues (BC ER+,
0.3340±0.1386 vs. BC ER-, 0.0017±0.0006; P<0.0001; Fig. 1E), although this difference was not
observed between ER-positive (n=44) and ER-negative (n=28) adjacent
normal tissues (paracancer ER+, 0.1436±0.0610 vs. paracancer ER-,
0.1197±0.0514; P=0.5994; Fig. 1F),
suggesting that AGR3 may be a tumor-specific gene that is
upregulated in breast cancer depending on the ER status. To confirm
the present findings, the correlation between AGR3 and ESR1
expression was analyzed using the GEO datasets. The correlation
coefficients in three GEO datasets were analyzed, and all displayed
significant positive correlations between AGR3 and ESR1 expression
(GSE5460, r=0.6633; 95% CI: 0.554–0.7502; P<0.0001; GSE19783,
r=0.8382; 95% CI: 0.7739–0.8854; P<0.0001; GSE19615, r=0.6411;
95% CI: 0.5189–0.7376; P<0.0001; Fig.
1G). The present results suggested that ESR1 and AGR3 may act
in a similar manner and play similar roles in breast cancer.
AGR3 is associated with the estrogen
response and cancer-associated signaling pathways according to
GSEA
To comprehensively understand the role of AGR3 in
breast cancer, GSEA was performed to explore the potential pathways
associated with AGR3. GSEA is an analytical method that can reveal
the biological pathways of candidate genes from gene sets (26). A total of 186 KEGG gene sets from the
MSigDB were used for the GSEA, and a number of AGR3-associated gene
sets were identified (Fig. 2A and
Table I). AGR3 was positively
associated with the ‘HALLMARK_ESTROGEN_RESPONSE_EARLY’ [enrichment
score (ES)=0.6592; normalized ES (NES)=2.5579; nominal P-value (Nom
P<0.0001; false discovery rate q-value (FDR q-val)=0;
family-wise error rate P-value (FWER P<0.0001; Fig. 2B] and the
‘HALLMARK_ESTROGEN_RESPONSE_LATE’ (ES=0.57267; NES=2.2178; NOM
P<0.0001; FDR q-val=0; FWER P<0.0001; Fig. 2C) gene sets, indicating that AGR3 may
function as an estrogen response factor. Additionally, AGR3 was
negatively associated with ‘HALLMARK_E2F_TARGETS’ (ES=−0.64839;
NES=−2.4647; NOM P<0.0001; FDR q-val=0; FWER P<0.0001;
Fig. 2D), allograft rejection,
G2/M checkpoint, mitotic spindle, interferon γ response,
Myc targets, mTORc1 signaling and other gene sets (Fig. 2A and Table
I), indicating that AGR3 may function in different
cancer-associated signaling pathways in breast cancer.
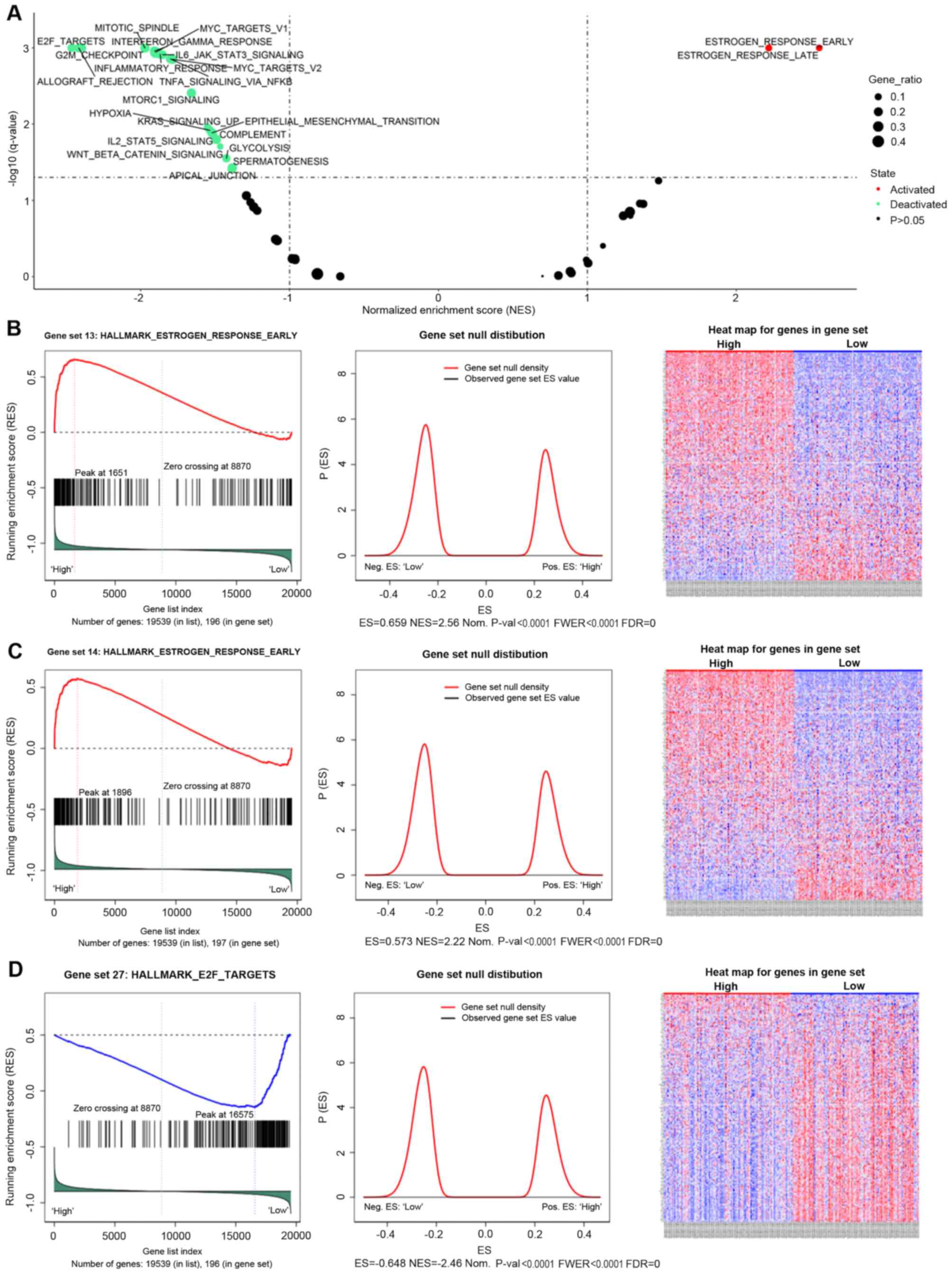 | Figure 2.GSEA results of hallmark gene set
analysis of AGR3 in breast cancer. (A) Volcano plot of the AGR3
GSEA results; enriched hallmark gene sets with an FDR q-value
<0.01 were considered statistically significant. GSEA enrichment
scores (left), gene set null distribution (middle) for gene sets
and heat maps (right) for genes in the hallmark gene sets: (B)
‘HALLMARK_ESTROGEN_RESPONSE_EARLY’, (C)
‘HALLMARK_ESTROGEN_RESPONSE_LATE’ and (D) ‘HALLMARK_E2F_TARGETS’.
The gene set null distribution was drawn using 1,000 permutations,
and the gene labels were rearranged each time. ES, enrichment
score; NES, normalized ES; NOM p-val, nominal P-value; FDR, false
discovery rate; FWER p-val: Familywise error rate P-value; GSEA,
Gene Set Enrichment Analysis; AGR3, anterior gradient 3. |
 | Table I.Gene Set Enrichment Analysis results
of hallmark gene sets of anterior gradient 3 in breast cancer. |
Table I.
Gene Set Enrichment Analysis results
of hallmark gene sets of anterior gradient 3 in breast cancer.
| Gene set | Size | ES | NES | NOM P-value | FDR q-value | FWER P-value | Gene ratio |
|---|
|
HALLMARK_ESTROGEN_RESPONSE_EARLY | 196 | 0.65919 | 2.5579 | <0.0001 | 0.00000000 | <0.0001 | 0.0845 |
|
HALLMARK_ESTROGEN_RESPONSE_LATE | 197 | 0.57267 | 2.2178 | <0.0001 | 0.00000000 | <0.0001 | 0.0970 |
|
HALLMARK_PEROXISOME | 103 | 0.41390 | 1.4776 | 0.0113100 | 0.05422300 | 0.1878 | 0.1120 |
|
HALLMARK_E2F_TARGETS | 196 | −0.64839 | −2.4647 | <0.0001 | 0.00000000 | <0.0001 | 0.1520 |
|
HALLMARK_ALLOGRAFT_REJECTION | 199 | −0.63245 | −2.4169 | <0.0001 | 0.00000000 | <0.0001 | 0.2010 |
|
HALLMARK_G2M_CHECKPOINT | 194 | −0.63038 | −2.3917 | <0.0001 | 0.00000000 | <0.0001 | 0.1380 |
|
HALLMARK_MITOTIC_SPINDLE | 198 | −0.51817 | −1.9722 | <0.0001 | 0.00000000 | <0.0001 | 0.2300 |
|
HALLMARK_INTERFERON_GAMMA_RESPONSE | 198 | −0.50047 | −1.9088 | <0.0001 | 0.00009480 | 0.0004 | 0.2280 |
|
HALLMARK_MYC_TARGETS_V1 | 197 | −0.49912 | −1.9013 | <0.0001 | 0.00013949 | 0.0007 | 0.3410 |
|
HALLMARK_INFLAMMATORY_RESPONSE | 196 | −0.49297 | −1.8694 | <0.0001 | 0.00020188 | 0.0012 | 0.2130 |
|
HALLMARK_IL6_JAK_STAT3_SIGNALING | 87 | −0.54467 | −1.8545 | <0.0001 | 0.00023175 | 0.0016 | 0.1880 |
|
HALLMARK_TNFA_SIGNALING_VIA_NFKB | 198 | −0.47236 | −1.7999 | <0.0001 | 0.00039442 | 0.0031 | 0.2300 |
|
HALLMARK_MYC_TARGETS_V2 | 58 | −0.56107 | −1.7859 | 0.0003642 | 0.00040505 | 0.0035 | 0.2310 |
|
HALLMARK_MTORC1_SIGNALING | 197 | −0.43572 | −1.6602 | <0.0001 | 0.00288310 | 0.0266 | 0.2200 |
|
HALLMARK_KRAS_SIGNALING_UP | 195 | −0.40799 | −1.5502 | 0.0003493 | 0.01011000 | 0.0989 | 0.1380 |
AGR3 is regulated by estrogen in T47D
cells
To explore the possible association between AGR3 and
ESR1, the RNA expression levels of AGR3 in different breast cancer
cell lines, including the ER-positive breast cancer MCF7, ZR75-1
and T47D cell lines and the ER-negative breast cancer MB231, MB468,
BT549 and SKBR-3 cell lines, were detected. AGR3 RNA expression was
markedly upregulated in T47D cells and was relatively abundant in
the ZR75-1 cell line, whereas very low levels were observed in the
other ER-negative breast cancer cell lines (MCF7, 0.0009±0.0001;
T47D, 6.8700±0.3437; ZR75-1, 1.0000±0.0000 (for the convenience of
comparing with other cell lines, ZR75-1 was normalized to 1);
BT549, NA (not available value that was too low to detect); MB231,
NA; MB468, 0.0013±0.0001; and SKBR-3,
5.443×10−5±5.862×10−6; Fig. 3A). To confirm AGR3 protein expression
in breast cancer cell lines, AGR3 protein levels were detected
using an AGR3 antibody. AGR3 protein was highly expressed in T47D
and ZR75-1 cells compared with the other cell lines (MCF7, BT549,
MB231, MB468 and SKBR-3). However, the AGR3 protein was lowly
expressed in ER-negative breast cancer cell lines, including BT549,
MB231, MB468 and SKBR-3 cells (Fig.
3B), consistent with the findings at the RNA level. ER protein
was also expressed in all the ER-positive breast cancer cell lines;
however, no ER protein was detected in ER-negative BT549, MB231,
MB468 and SKBR-3 breast cancer cell lines (Fig. 3B).
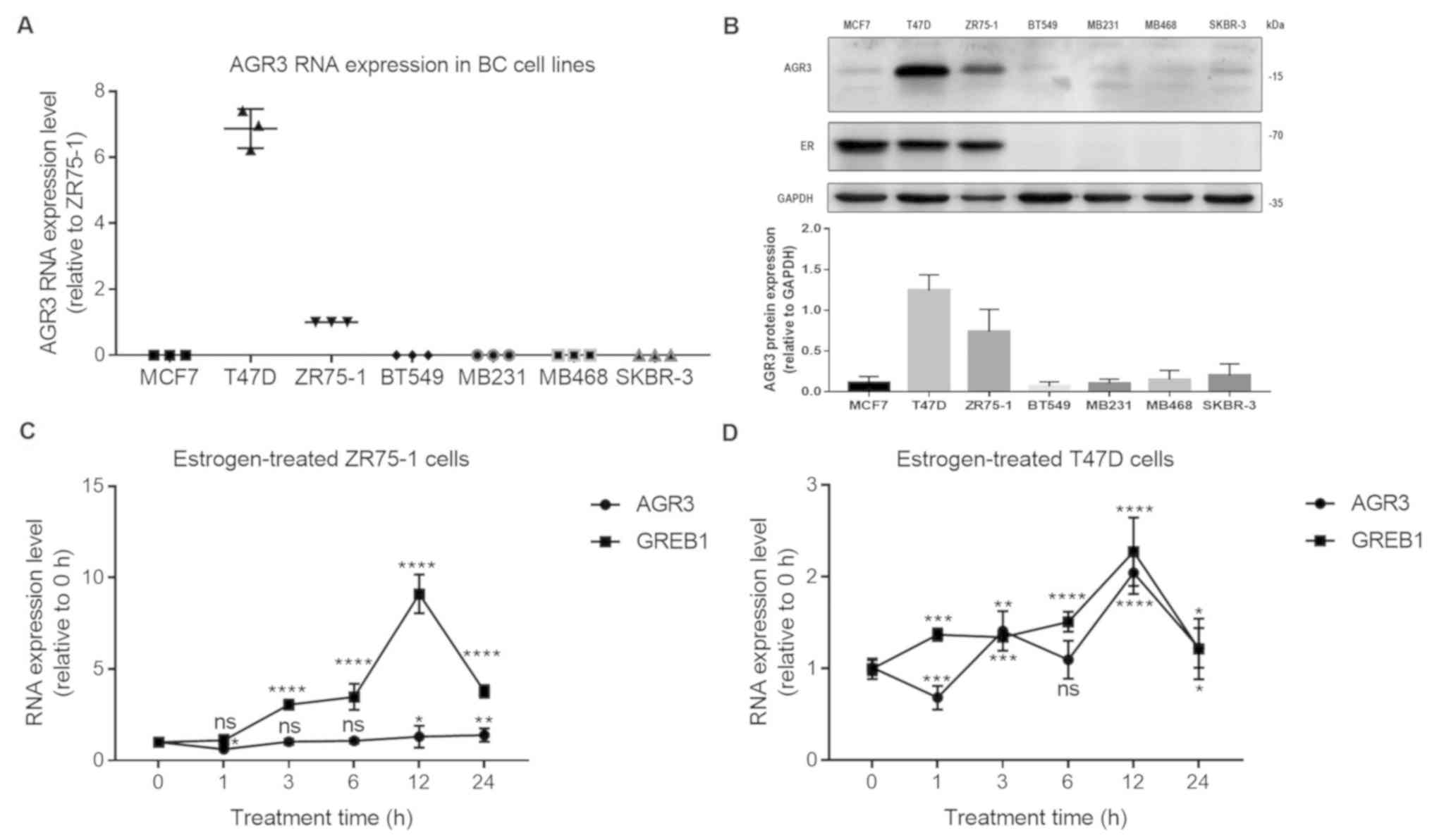 | Figure 3.AGR3 is upregulated in estrogen
receptor-positive breast cancer cell lines and regulated by
estrogen in T47D breast cancer cells. (A) Reverse
transcription-quantitative PCR analysis of AGR3 RNA expression in
breast cancer MCF7, T47D, ZR75-1 (normalized to 1 for the
convenience of comparing with other cell lines), BT549, MB231,
MB468 and SKBR-3 cell lines. (B) Western blot analysis of AGR3
protein levels in breast cancer MCF7, T47D, ZR75-1, BT549, MB231,
MB468 and SKBR-3 cell lines (top) and the bar graph of AGR3 protein
levels (bottom). AGR3 and GREB1 RNA expression was analyzed in (C)
ZR75-1 cells and (D) T47D cells after treatment with 10 nmol
estrogen for 0, 1, 3, 6, 12 and 24 h. The RNA expression level of 0
h was normalized to 1 in (C) and (D), and GREB1 was used as an
estrogen-regulated positive control. Each experiment was repeated
≥3 times. *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001
vs. 0 h. AGR3, anterior gradient 3; BC, breast cancer; GREB1,
Growth regulating estrogen receptor binding 1; ns, not
significant. |
Estrogen is an important factor in breast cancer,
and it can directly activate the ER (27). To examine whether estrogen regulates
AGR3 expression, estrogen was added to T47D and ZR75-1 cells at a
final concentration of 10 nmol, and the RNA level of AGR3 at
different time intervals after treatment was detected. Although the
RNA levels of GREB1, an estrogen response molecule (28), were gradually increased and reached
the highest level after 12 h of estrogen treatment in ZR75-1 cells
(12 h, 9.1120±0.2447 vs. 0 h, 1.0000±0.0000 (0 h was normalized to
1); P<0.0001; one-way ANOVA, F=512.4 and P<0.0001), AGR3 did
not appear to be significantly increased in ZR75-1 cells treated
with estrogen for 12 h (12 h, 1.3008±0.1390 vs. 0 h, 1.0000±0.0000
(0 h was normalized to 1); P=0.0368) or 24 h (24 h, 1.3900±0.08388
vs. 0 h, 1.0000±0.0000; P=0.0085; one-way ANOVA, F=14.8 and
P<0.0001; Fig. 3C). However, the
RNA expression levels of AGR3 and GREB1 were increased in a similar
manner in T47D cells, and both reached their highest levels after
estrogen treatment for 12 h (AGR3: 12 h, 2.043±0.05334 vs. 0 h,
1.0000±0.0000 (0 h was normalized to 1); P<0.0001; one-way
ANOVA, F=110.6 and P<0.0001; GREB1: 12 h, 2.274±0.08661 vs. 0 h,
1.0000±0.0000 (0 h was normalized to 1); P<0.0001; one-way
ANOVA, F=77.2 and P<0.0001; Fig.
3D), suggesting that AGR3 may be regulated by estrogen in
ER-positive breast cancer cells.
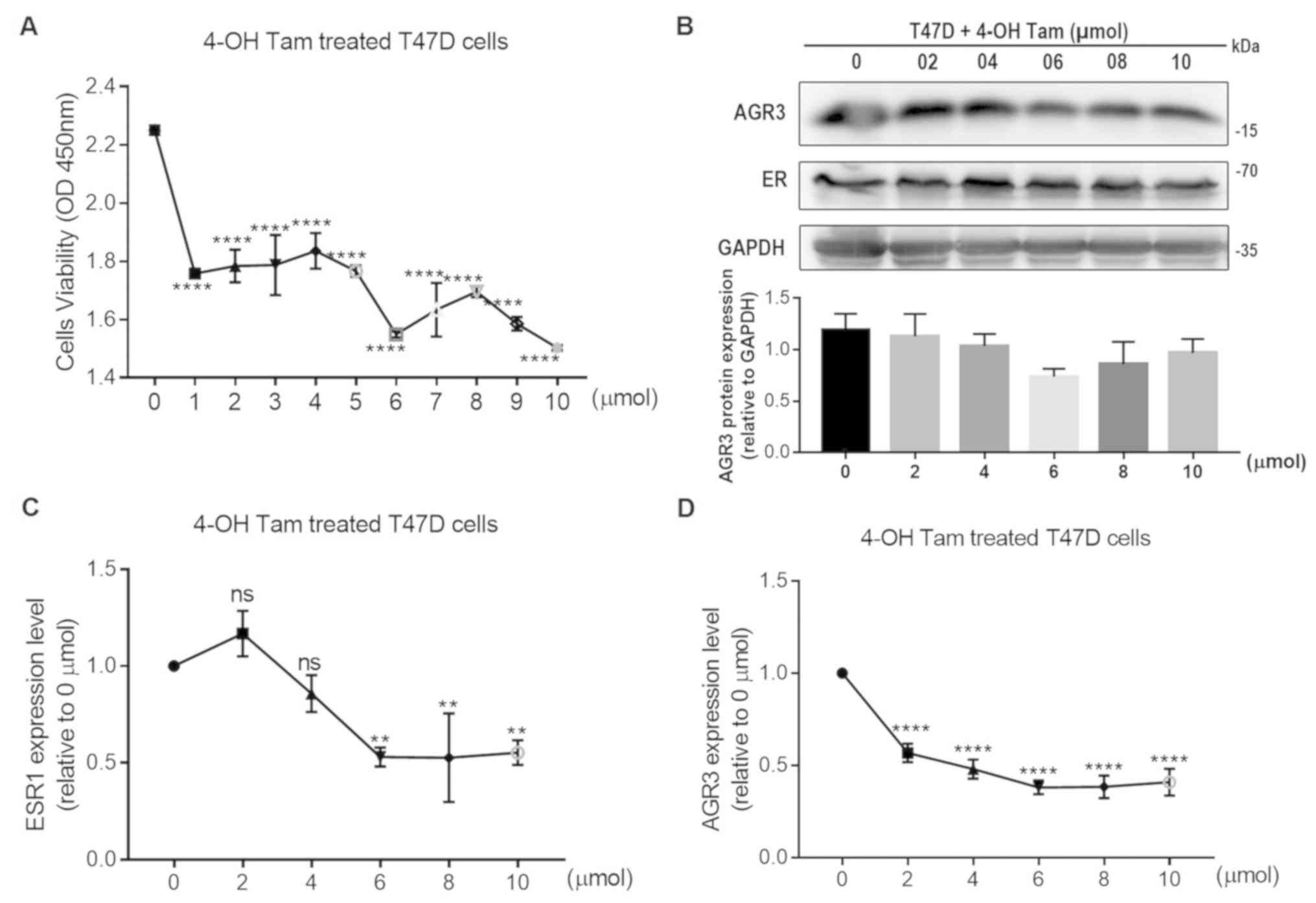
To further investigate the association between
estrogen and AGR3, the present study assessed whether 4-OH Tam may
influence the viability of T47D cells and the expression levels of
AGR3. When different concentrations of 4-OH Tam were added to T47D
cells, cell viability was significantly decreased with increasing
4-OH Tam concentrations, and was the lowest after the addition of
10 µmol 4-OH Tam (10 µmol, 1.5030±0.0052 vs. 0 µmol, 2.2510±0.0092;
P<0.0001; one-way ANOVA, F=48.2 and P<0.0001; Fig. 4A). Additionally, the protein levels
of AGR3 and ER were analyzed; these appeared to decrease following
4-OH Tam treatment, with protein levels of AGR3 in T47D cells
reaching their lowest level using 6 µmol 4-OH Tam, and ER protein
levels also had a visible decline following addition of 6 µmol 4-OH
Tam (Fig. 4B). Additionally, AGR3
and ESR1 RNA expression was significantly decreased in T47D cells
following treatment with different concentrations of 4-OH Tam. ESR1
and AGR3 RNA expression reached their lowest levels in T47D cells
using 6 µmol 4-OH Tam (ESR1: 6 µmol, 0.5301±0.0282 vs. 0 µmol,
1.0000±0.0000 (0 µmol was normalized to 1); P=0.0015; one-way
ANOVA, F=16.9 and P<0.0001; AGR3: 6 µmol, 0.3810±0.0212 vs. 0
µmol, 1.0000±0.0000 (0 µmol was normalized to 1); P<0.0001;
one-way ANOVA, F=65.8 and P<0.0001; Fig. 4C and D). Therefore, both ESR1 and
AGR3 expression may be regulated by 4-OH Tam, which further
confirmed the effect of estrogen on AGR3 RNA and protein
expression.
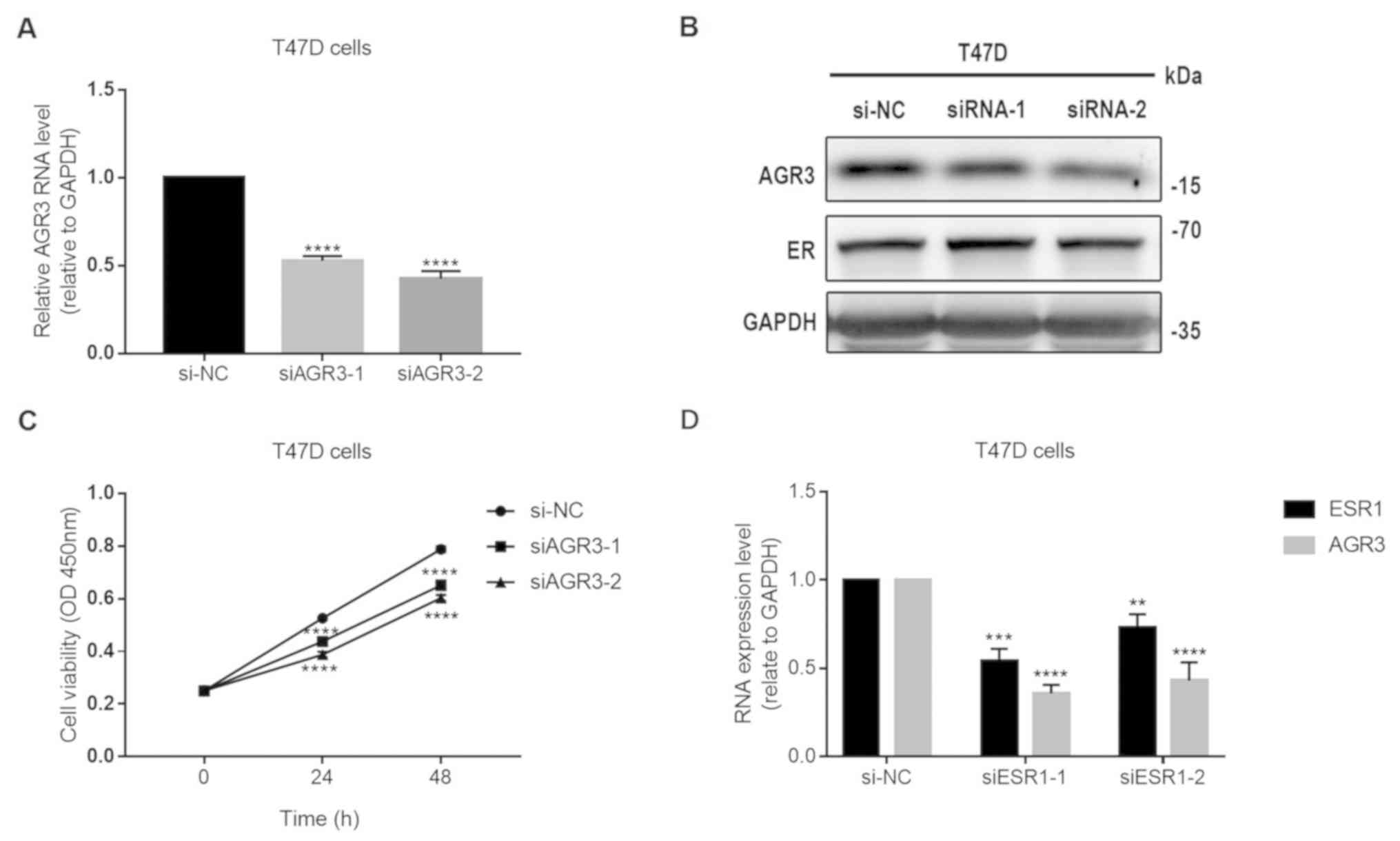
AGR3 promotes T47D cell proliferation
in an estrogen- dependent manner
To further explore the role of AGR3 in ER-positive
breast cancer, the present study analyzed whether AGR3 expression
may influence the ER status by knocking down AGR3 expression in
T47D cells using two siRNAs. Both siRNAs significantly decreased
AGR3 RNA levels (siAGR3-1, 0.5265±0.0150 vs. si-NC, 1.0000±0.0000;
P<0.0001; siAGR3-2, 0.4257±0.0243 vs. si-NC, 1.0000±0.0000;
P<0.0001; one-way ANOVA, F=345.2 and P<0.0001; Fig. 5A). The protein levels of AGR3
decreased after transfecting T47D cells with the AGR3 siRNAs, but
there was no notable change in ER protein expression (Fig. 5B). After knocking down AGR3 using
siRNAs, the viability of T47D cells was significantly decreased
compared with that of the negative control cells 24 h
post-transfection (siAGR3-1, 0.4383±0.007 vs. si-NC
0.5259±0.002963; P<0.0001; siAGR3-2, 0.3883±0.006245 vs. si-NC,
0.5259±0.002963; P<0.0001; one-way ANOVA, F=150.5 and
P<0.0001) and 48 h post-transfection (siAGR3-1, 0.6512±0.0103
vs. si-NC, 0.7879±0.0062; P<0.0001; siAGR3-2, 0.6032±0.0067 vs.
si-NC, 0.7879±0.0062; P<0.0001; one-way ANOVA, F=145.5 and
P<0.0001) (Fig. 5C).
Additionally, siRNAs against ESR1 significantly decreased the
expression levels of ESR1 (siESR1-1, 0.7283±0.0456 vs. si-NC,
1.0000±0.0000; P=0.0026; siESR1-2, 0.5435±0.0385 vs. si-NC,
1.0000±0.0000; P=0.0002; one-way ANOVA, F=44.4 and P=0.0003) and
AGR3 (siESR1-1, 0.4302±0.0604 vs. si-NC, 1.0000±0.0000;
P<0.0001; siESR1-2, 0.3576±0.0285 vs. si-NC, 1.0000±0.0000;
P<0.0001; one-way ANOVA, F=83.3 and P<0.0001) in T47D cells
(Fig. 5D). In conclusion, the
present results suggest that AGR3, which is regulated by ESR1, may
promote the proliferation of ER-positive breast cancer cells in an
estrogen-dependent manner.
Discussion
AGR2 and AGR3 both belong to the PDI family, and
they are similarly expressed in breast cancer due to the 71%
similarity in their protein sequences and adjacent positions at
7p21 (29). AGR2 is a widely
accepted oncogene in breast cancer and is regulated by estrogen in
normal breast tissue and ER-positive breast cancer cells (30,31).
Although AGR2 and AGR3 share sequence homology, they are not
identical in function, and AGR3 may serve a different role in
breast cancer (12). Recent studies
have revealed the oncogenic functions of AGR3 in breast cancer. For
example, extracellular AGR3 was demonstrated to regulate breast
cancer cell migration via Src signaling, which provided an
exogenous mode of expression (17),
whereas the present study focused on endogenous AGR3 and its
function in ER-positive breast cancer cells. In addition, AGR3
promotes the proliferative and invasive abilities of breast cancer
cells and chemotherapy response (18), which further confirmed the oncogenic
role of AGR3 in breast cancer. In order to fully understand the
function of AGR3 in breast cancer, the present study aimed to
explore the function of AGR3 in ER-positive breast cancer, and
focused on the direct association between AGR3 and ER.
In the present study, AGR3 expression was
significantly upregulated in ER-positive compared with ER-negative
breast cancer tissues. Considering the close association between
AGR3 and ER status, and that ER is a key molecule in distinguishing
between low- and high-grade breast cancer, AGR3 RNA expression was
detected in ER-positive and ER-negative breast cancer tissues and
their paired adjacent normal breast tissues. AGR3 expression was
significantly upregulated in luminal breast cancer tissues compared
with that in paired adjacent normal tissues, and was significantly
downregulated in TN breast cancer tissues compared with that in
paired adjacent normal tissues. The results of the present study
suggest that there may be different modes of AGR3 expression
between ER-positive and ER-negative breast cancer. However, further
studies are required to determine whether differential expression
tendencies of AGR3 have the same or different function in
ER-positive and ER-negative breast cancers. Of note, no differences
were observed in AGR3 expression between ER-positive and
ER-negative adjacent normal breast tissues, indicating that the
differential expression tendency only occurred in breast cancer
samples. Therefore, AGR3 may be a tumor-specific gene, and its
expression may depend on the ER status of the breast.
To fully understand the role of AGR3 in breast
cancer, GSEA of AGR3 was conducted using a breast cancer TCGA
dataset; the results revealed that AGR3 was positively associated
with estrogen response gene sets, including early and late steps.
Additionally, when estrogen was added to T47D cells, AGR3 RNA
expression was increased by indicating the peak time, and AGR3 was
confirmed to be an estrogen response molecule in ER-positive breast
cancer. Furthermore, AGR3 was also negatively associated with
several cancer-associated gene sets, such as ‘E2F_TARGETS’,
‘G2M_CHECKPOINT’ and ‘MITOTIC_SPINDLE’, which are associated with
the cell cycle regulation; as well as ‘ALLOGRAFT_REJECTION’,
‘IL6_JAK_STAT3_SIGNALING’, ‘TNFA_SIGNALING_VIA_NFKB’, which are
associated with immunity and inflammation;
‘EPITHELIAL_MESENCHYMAL_TRANSITION’ and
‘WNT_BETA_CATENIN_SIGNALING’, which are associated with
epithelial-mesenchymal transition. Therefore, the function of AGR3
in breast cancer requires further investigation.
In conclusion, the results of the present study
demonstrated that AGR3 was an estrogen response gene that was
closely associated with ER status in breast cancer. In ER-positive
breast cancer, AGR3 expression was positively correlated with ESR1
expression levels, and in ER-negative breast cancer, AGR3
expression was significantly downregulated compared with that in
paired adjacent normal tissues. The differential expression modes
of AGR3 according to ER status may lead to different functions in
breast cancer. In ER-positive breast cancer cells, AGR3 was
responsive to estrogen, and ESR1-regulated AGR3 promoted the
proliferation of ER-positive T47D breast cancer cells. Thus, AGR3
may serve an important function in estrogen-mediated cell
proliferation in breast cancer, whereas the role of AGR3 in
ER-negative breast cancer requires further investigation. Knockdown
of AGR3 may be a potential therapeutic strategy for ER-positive
breast cancer.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81772979).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LJ, JX, HY, KT, CY and KL performed the experiments
and analyzed the data. SL provided the reagents, designed the
experiments and critically revised the manuscript. LJ, SG and RC
designed the study and drafted the initial manuscript. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
The collection of tumor samples and patient
information was approved by the Ethics Committee of The First
Affiliated Hospital of Chongqing Medical University (approval no.
2017-16; Chongqing, China). All patients signed informed consent
for the retention and analysis of their tissues for research
purposes.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Coates AS, Winer EP, Goldhirsch A, Gelber
RD, Gnant M, Piccart-Gebhart M, Thürlimann B, Senn HJ and Panel M:
Tailoring therapies-improving the management of early breast
cancer: St gallen international expert consensus on the primary
therapy of early breast cancer 2015. Ann Oncol. 26:1533–1546. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Allred DC: Issues and updates: Evaluating
estrogen receptor- alpha, progesterone receptor, and HER2 in breast
cancer. Mod Pathol. 23 (Suppl 2):S52–S59. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bai Z and Gust R: Breast cancer, estrogen
receptor and ligands. Arch Pharm (Weinheim). 342:133–149. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yue W, Yager JD, Wang JP, Jupe ER and
Santen RJ: Estrogen receptor-dependent and independent mechanisms
of breast cancer carcinogenesis. Steroids. 78:161–170. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jia M, Dahlman-Wright K and Gustafsson JA:
Estrogen receptor alpha and beta in health and disease. Best Pract
Res Clin Endocrinol Metab. 29:557–568. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jia M, Andreassen T, Jensen L, Bathen TF,
Sinha I, Gao H, Zhao C, Haldosen LA, Cao Y, Girnita L, et al:
Estrogen receptor α promotes breast cancer by reprogramming choline
metabolism. Cancer Res. 76:5634–5646. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Persson S, Rosenquist M, Knoblach B,
Khosravi-Far R, Sommarin M and Michalak M: Diversity of the protein
disulfide isomerase family: Identification of breast tumor induced
Hag2 and Hag3 as novel members of the protein family. Mol
Phylogenet Evol. 36:734–740. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ivanova AS, Tereshina MB, Ermakova GV,
Belousov VV and Zaraisky AG: Agr genes, missing in amniotes, are
involved in the body appendages regeneration in frog tadpoles. Sci
Rep. 3:12792013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li Z, Wu Z, Chen H, Zhu Q, Gao G, Hu L,
Negi H, Kamle S and Li D: Induction of anterior gradient 2 (AGR2)
plays a key role in insulin-like growth factor-1 (IGF-1)-induced
breast cancer cell proliferation and migration. Med Oncol.
32:5772015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Verma S, Salmans ML, Geyfman M, Wang H, Yu
Z, Lu Z, Zhao F, Lipkin SM and Andersen B: The estrogen-responsive
Agr2 gene regulates mammary epithelial proliferation and
facilitates lobuloalveolar development. Dev Biol. 369:249–260.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Obacz J, Takacova M, Brychtova V, Dobes P,
Pastorekova S, Vojtesek B and Hrstka R: The role of AGR2 and AGR3
in cancer: Similar but not identical. Eur J Cell Biol. 94:139–147.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
King ER, Tung CS, Tsang YT, Zu Z, Lok GT,
Deavers MT, Malpica A, Wolf JK, Lu KH, Birrer MJ, et al: The
anterior gradient homolog 3 (AGR3) gene is associated with
differentiation and survival in ovarian cancer. Am J Surg Pathol.
35:904–912. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bu H, Schweiger MR, Manke T, Wunderlich A,
Timmermann B, Kerick M, Pasqualini L, Shehu E, Fuchsberger C, Cato
AC and Klocker H: Anterior gradient 2 and 3-two prototype
androgen-responsive genes transcriptionally upregulated by
androgens and by oestrogens in prostate cancer cells. FEBS J.
280:1249–1266. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Brychtova V, Zampachova V, Hrstka R,
Fabian P, Novak J, Hermanova M and Vojtesek B: Differential
expression of anterior gradient protein 3 in intrahepatic
cholangiocarcinoma and hepatocellular carcinoma. Exp Mol Pathol.
96:375–381. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Obacz J, Brychtova V, Podhorec J, Fabian
P, Dobes P, Vojtesek B and Hrstka R: Anterior gradient protein 3 is
associated with less aggressive tumors and better outcome of breast
cancer patients. Onco Targets Ther. 8:1523–1532. 2015.PubMed/NCBI
|
|
17
|
Obacz J, Sommerova L, Sicari D, Durech M,
Avril T, Iuliano F, Pastorekova S, Hrstka R, Chevet E, Delom F and
Fessart D: Extracellular AGR3 regulates breast cancer cells
migration via Src signaling. Oncol Lett. 18:4449–4456.
2019.PubMed/NCBI
|
|
18
|
Xu Q, Shao Y, Zhang J, Zhang H, Zhao Y,
Liu X, Guo Z, Chong W, Gu F and Ma Y: Anterior gradient 3 promotes
breast cancer development and chemotherapy response. Cancer Res
Treat. 52:218–245. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lu X, Lu X, Wang ZC, Iglehart JD, Zhang X
and Richardson AL: Predicting features of breast cancer with gene
expression patterns. Breast Cancer Res Treat. 108:191–201. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Enerly E, Steinfeld I, Kleivi K, Leivonen
SK, Aure MR, Russnes HG, Rønneberg JA, Johnsen H, Navon R, Rødland
E, et al: miRNA-mRNA integrated analysis reveals roles for miRNAs
in primary breast tumors. PLoS One. 6:e169152011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li Y, Zou L, Li Q, Haibe-Kains B, Tian R,
Li Y, Desmedt C, Sotiriou C, Szallasi Z, Iglehart JD, et al:
Amplification of LAPTM4B and YWHAZ contributes to chemotherapy
resistance and recurrence of breast cancer. Nat Med. 16:214–218.
2010. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Grossman RL, Heath AP, Ferretti V, Varmus
HE, Lowy DR, Kibbe WA and Staudt LM: Toward a shared vision for
cancer genomic data. N Engl J Med. 375:1109–1112. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Love MI, Huber W and Anders S: Moderated
estimation of fold change and dispersion for RNA-seq data with
DESeq2. Genome Biol. 15:5502014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Durbin BP, Hardin JS, Hawkins DM and Rocke
DM: A variance-stabilizing transformation for gene-expression
microarray data. Bioinformatics. 18 (Suppl 1):S105–S110. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Subramanian A, Tamayo P, Mootha VK,
Mukherjee S, Ebert BL, Gillette MA, Paulovich A, Pomeroy SL, Golub
TR, Lander ES and Mesirov JP: Gene set enrichment analysis: A
knowledge-based approach for interpreting genome-wide expression
profiles. Proc Natl Acad Sci USA. 102:15545–15550. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yager JD and Davidson NE: Estrogen
carcinogenesis in breast cancer. N Engl J Med. 354:270–282. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mohammed H, D'Santos C, Serandour AA, Ali
HR, Brown GD, Atkins A, Rueda OM, Holmes KA, Theodorou V, Robinson
JL, et al: Endogenous purification reveals GREB1 as a key estrogen
receptor regulatory factor. Cell Rep. 3:342–349. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fletcher GC, Patel S, Tyson K, Adam PJ,
Schenker M, Loader JA, Daviet L, Legrain P, Parekh R, Harris AL and
Terrett JA: hAG-2 and hAG-3, human homologues of genes involved in
differentiation, are associated with oestrogen receptor-positive
breast tumours and interact with metastasis gene C4.4a and
dystroglycan. Br J Cancer. 88:579–585. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hrstka R, Nenutil R, Fourtouna A, Maslon
MM, Naughton C, Langdon S, Murray E, Larionov A, Petrakova K,
Muller P, et al: The pro-metastatic protein anterior gradient-2
predicts poor prognosis in tamoxifen-treated breast cancers.
Oncogene. 29:4838–4847. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wilson CL, Sims AH, Howell A, Miller CJ
and Clarke RB: Effects of oestrogen on gene expression in
epithelium and stroma of normal human breast tissue. Endocr Relat
Cancer. 13:617–628. 2006. View Article : Google Scholar : PubMed/NCBI
|