Introduction
Colorectal cancer (CRC) is one of the most common
malignancies and the second most common cause of cancer-associated
mortality globally (1). The
incidence and mortality of CRC has increased in previous years,
particularly in developing Asian countries, including China
(2). Despite advances in the
prevention, diagnosis and treatment of CRC, to the best of our
knowledge, no effective treatment strategy for this disease has yet
been developed. Metastasis and recurrence are the main reasons why
CRC treatment may fail (3,4). Therefore, it is vital to elucidate the
molecular mechanisms governing CRC progression and migration and
invasion.
γ-glutamylcyclotransferase (GGCT) is an important
enzyme in glutathione metabolism, which catalyzes the reaction of
the γ-glutamyl peptide to produce 5-oxyproline and free amino acids
(5). The GGCT gene is highly
preserved in a number of species, including in bacteria, plants and
nematodes, and a number of other higher organisms (6). GGCT has been reported to accumulate in
a variety of cancer types, including breast, ovarian, cervical,
lung, bladder, prostate, colon, osteosarcoma and glioma, indicating
that it may serve as an oncogene in these tumor types (7–12). The
depletion of GGCT has been indicated to inhibit the proliferation
of bladder cancer cells and induce the cytotoxicity of McF-7/ADR
cells in vitro and in vivo (13). In previous years, one study has
identified the suppression of cancer cell proliferation by
disrupting GGCT, highlighting the potential for treatment of
malignant tumor types through inhibiting GGCT (14). However, the underlying mechanism as
to how GGCT regulates CRC progression remains yet to be determined.
Therefore, investigation into the precise function of GGCT in human
CRC is urgently required.
The present study demonstrated the association
between GGCT expression level and the prognosis of patients with
CRC. Further results indicated that GGCT promoted CRC cells
migration and invasion via the epithelial to mesenchymal transition
(EMT). In summary, these data support that GGCT may be a novel
therapeutic target for use in the treatment of CRC.
Materials and methods
Microarray and database analysis
A microarray of mRNA from 286 colorectal tumor
samples was downloaded from The Cancer Genome Atlas (TCGA)
Colorectal Cancer database (https://www.cancer.gov/tcga). These samples were
separated into a low and high group based on the cut-off value of
GGCT median expression (cut-off value=39.1), which was obtained
from the online TCGA Colorectal Cancer database. Kyoto Encyclopedia
of Genes and Genomes (KEGG) pathway analysis of this microarray was
performed using KEGG PathwayFinder with a gene correlation that was
based on GGCT expression using the R2 web application (15–17). The
absolute r cutoff was set at r=0.5, P<0.05. Gene Set Enrichment
Analysis (GSEA; http://www.broadinstitute.org/gsea/index.jsp) was
conducted using the KEGG_CELL_ADHENSION_MOLECULES_CAMS, Gene
Ontology (GO)_EXTRACELLULAR_MATRIX, KEGG_FOCAL_ADHENSION and
HALLMARK_EPITHELIAL_MESENCHYMAL_TRANSITION gene sets (18,19). A
group of 594 patient specimens from the TCGA Colorectal Cancer
database (http://www.cbioportal.org/datasets) was used to
evaluate the expression and prognostic value of GGCT in patients
with CRC.
Cell culture and patient
specimens
CRC cell lines HCT-116 (ATCC® CCL-247)
and SW620 (ATCC® CCL-227) were purchased from American
Type Culture Collection. These cells were cultured in Dulbecco's
modified Eagle's medium (DMEM) containing 10% fetal bovine serum
(FBS), digested using trypsin every 2 days and cultured at 37°C in
a humidified incubator with 5% CO2. A total of 6 paired
CRC surgical specimens and corresponding adjacent normal colon
specimens were obtained from the Department of General Surgery,
Renhe Hospital (Shanghai, China) once written informed consent was
obtained from patients or their guardians. The present study was
ethically approved by the Ethic Committee of Renhe Hospital.
Western blot analysis
Western blot analysis was performed as previously
described (20). A total of
5×105 CRC cells were washed three times in cold
phosphate buffered saline (PBS), and total protein extracts were
obtained using RIPA buffer [50 mM Tris (pH 8.0), 150 mM NaCl, 0.5%
sodium deoxycholate, 0.1% SDS, 1% NP40, 1 mM EDTA and a mix of
protease inhibitors]. Total proteins were quantified using a
bicinchoninic acid assay (Pierce; Thermo Fisher Scientific, Inc.).
Lysates were subsequently collected using centrifugation at 12,000
× g for 10 min at room temperature, and 20 µg proteins were
separated by 10% SDS-PAGE and transferred to polyvinylidene
difluoride membranes. The membranes were blocked with 5% non-fat
dried milk in TBS and Tween-20 for 1 h at room temperature. The
membranes were then incubated overnight at 4°C with the following
specific primary antibodies: Anti-GGCT (1:1,000; cat. no.
PA5-54263; Invitrogen; Thermo Fisher Scientific), anti-N-Cadherin
(1:1,000; cat. no. 13116; Cell Signaling Technology, Inc.),
anti-vimentin (1:1,000; cat. no. 5741; Cell Signaling Technology,
Inc.), anti-snail family transcriptional repressor 2 (Slug;
1:1,000; cat. no. 9585; Cell Signaling Technology, Inc.),
anti-snail family transcriptional repressor 1 (Snail; 1:1,000; cat.
no. 3879; Cell Signaling Technology, Inc.) and anti-β-Actin
(1:1,000; cat. no. 3700; Cell Signaling Technology, Inc.).
Following the primary incubation, membranes were incubated for 2 h
at room temperature with either of the following horseradish
peroxidase-conjugated secondary antibodies: Goat anti-mouse
(1:2,000; cat. no. sc-2005) and goat anti-rabbit IgG (1-,000; cat.
no. sc-2004) (all from Santa Cruz Biotechnlogy).
Lentiviral vectors construction
GGCT-short-hairpin (sh)RNA lentiviral vectors and a
non-targeting control shRNA (shNT; SHC002) vector were purchased
from Hanbio Biotechnology Co., Ltd. The shRNAs used were as
follows; shCtrl forward,
5′-GCTCAGCGGGAGGAGGCTATATATTGCAAGAGAACGTTT-3′ and reverse,
5′-ACTTGAACAAAGCACGCGGAGGCACAGTGTCCTCT-3′; shGGCT#1 forward,
5′-GACACGACGTCCCGTCTAGGCTGTCAGTCAAGCGGCCTCGGCAT-3′ and reverse,
5′-ACGCGCACCCGTGCGACTTGTGCATACACAC-3′; shGGCT#2 forward,
5′-GCTCTGTATCAGATGTGTCTCGTATACATGAGCTT-3′; and reverse,
5′-ACTTGGATAAGTTACGACCTATCCGTCATCTCTTGAAT-3′. A total of
8×104 CRC cells were plated overnight in 500 µl growth
medium, in a single well of a 24-well plate. Cells were transiently
transfected with 15 pmol shRNAs against GGCT or a non-targeting
control using 1.5 µl Lipofectamine 3000 reagent for 48 h at 37°C in
a humidified incubator with 5% CO2 according to the
manufacturer's protocol (cat. no. L3000015; Thermo Fisher
Scientific, Inc.).
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
assay
Total RNA of GGCT-knockdown and control HCT-116 and
SW620 cells were extracted using a total RNA extracting kit
purchased from Fastagen Biotechnology according to the
manufacturer's protocol. For RT-qPCR, mRNAs mixture was incubated
at 42°C for 60 min and 70°C for 15 min to be reverse transcribed to
cDNA using the Reverse Transcription kit (Takara Biotechnology Co.,
Ltd.). RT-qPCR was performed using SYBR Premix Ex Taq (Takara
Biotechnology Co., Ltd.). The thermocycling conditions were as
follows: Pre-denaturation at 94°C for 5 min; 33 cycles of 94°C for
30 sec, 64°C for 30 sec and 72°C for 45 sec; and 72°C for 10 min.
Results were normalized to the expression of β-actin using the
2−ΔΔCq method (21). The
sequence of the primers used were as follows: β-actin forward,
5′-CATGTACGTTGCTATCCAGGC-3′ and reverse,
5′-CTCCTTAATGTCACGCACGAT-3′; GGCT forward,
5′-TGGCAATTCCCAAGGCAAAAC-3′ and reverse,
5′-CCCCTTCTTGCTCATCCAGAG-3′; N-cadherin forward,
5′-TCAGGCGTCTGTAGAGGCTT-3′ and reverse,
5′-ATGCACATCCTTCGATAAGACTG-3′; Vimentin forward
5′-GACGCCATCAACACCGAGTT-3′ and reverse,
5′-CTTTGTCGTTGGTTAGCTGGT-3′; Slug forward,
5′-CGAACTGGACACACATACAGTG-3′ and reverse,
5′-CTGAGGATCTCTGGTTGTGGT-3′; Snail forward,
5′-TCGGAAGCCTAACTACAGCGA-3′ and reverse,
5′-AGATGAGCATTGGCAGCGAG-3′.
Transwell invasion and migration
assay
For the transwell invasion assay, GGCT-knockdown and
control HCT-116 cells were trypsinized and resuspended with
serum-free DMEM. Cells were then seeded into suspension cell
culture inserts (8.0 µm; EMD Millipore), which were firstly coated
with 1 mg/ml Matrigel (BD Biosciences), at a density of
4×104 cells/well in 200 µl DMEM with 6 replicate wells,
and 500 µl DMEM with 10% FBS was added to the lower chambers of the
24-well plate. Subsequent to incubation at 37°C with 5%
CO2 for 24 h, inserts were washed with PBS and fixed
with 4% paraformaldehyde for 15 min at room temperature. The
migration ability of cells was examined by using 8.0 µm cell
culture inserts without coating Matrigel. Following 12 h incubation
at 37°C, the upper inserts were removed and then washed and fixed
as described above. All inserts were subsequently wiped and cells
were stained using a crystal violet solution (Beyotime Institute of
Biotechnology) for 15 min at room temperature. Stained cells were
counted and images were acquired using a microscope (Olympus
Corporation).
Wound-healing assay
GGCT-knockdown and control HCT-116 cells were seeded
at a density of 2×103 cells/well in 24-well plates with
serum-free DMEM, and a wound was created using a 10 µl plastic
pipette tip in three replicate wells. The migration distance was
observed and measured after 24 h incubation at 37°C. The images
were acquired using a light microscope (Olympus) at a magnification
of ×100. A total of 9 areas were selected randomly in each well at
×100 magnification and the migration distance was measured using
the software program HMIAS-2000 version 2.0 (Wuhan Qianping Imaging
Technology Co., Ltd.) as previously described (22).
Statistical analysis
All experiments were performed at least three times
with triplicate samples. The survival curves were created using
Kaplan-Meier analysis, and P values were calculated using a
Log-rank test. Statistical analysis was performed using SPSS
statistical software (SPSS16.0; SPSS, Inc.) and GraphPad Prism 6
software (GraphPad Software, Inc.) The unpaired two-group
comparison and multiple comparisons were made using the unpaired
Student's t-test or one-way analysis of variance with
Student-Keuls-Neuman post hoc test, respectively. Multiple
comparison between the groups was performed using method. Data were
presented as the mean ± standard deviation. P<0.05 was
considered to indicate a statistically significant difference.
Results
GGCT is upregulated in human CRC
tissues and is associated with a poor prognosis
As GGCT has been previously demonstrated to be
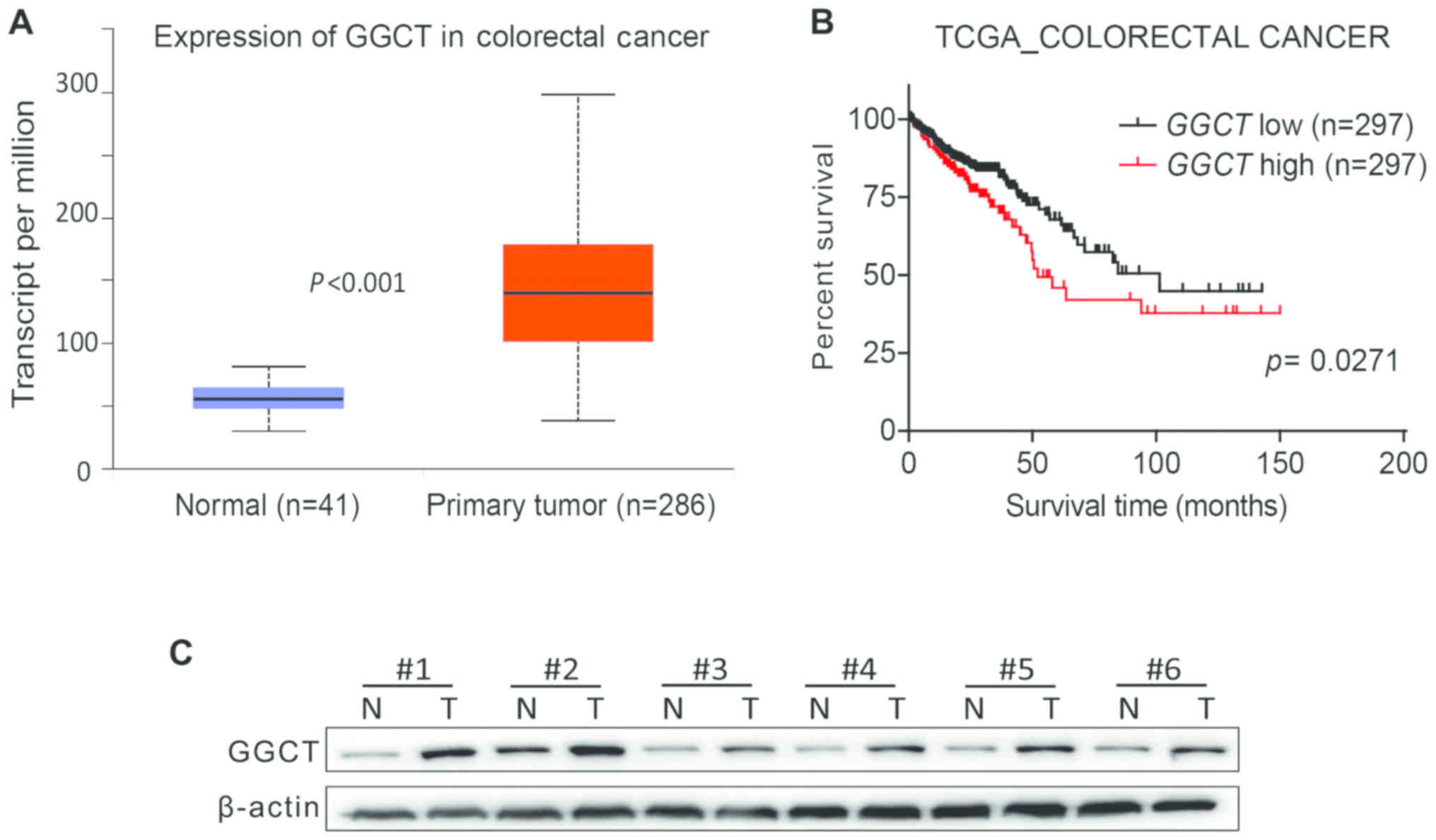
overexpressed in a variety of cancer types (5), the effect of GGCT expression on mRNA
levels in human CRC was assessed by analyzing TCGA Colorectal
Cancer database, and the results indicated that GGCT expression was
increased significantly in CRC tissues compared with normal colon
tissues (P<0.001; Fig. 1A).
Furthermore, the prognostic value of GGCT was evaluated, and the
Kaplan-Meier survival curve revealed that the overall survival time
for patients with CRC with a higher GGCT expression was
significantly decreased compared with patients exhibiting a lower
GGCT expression (P=0.0271; Fig. 1B).
To evaluate the expression of GGCT on the protein expression in
human CRC, six pairs of tissue samples were examined using western
blot analysis. The pathological features of the CRC specimens were
summarized in Table I. The results
demonstrated that GGCT expression was increased in the tissues of
patients with CRC compared with corresponding normal colon tissues
(Fig. 1C). In conclusion, the
results revealed that GGCT was upregulated in human CRC tissues and
was associated with a poor patient prognosis.
 | Table I.Basic clinicopathological
characteristics of the colorectal cancer specimens used in the
present study. |
Table I.
Basic clinicopathological
characteristics of the colorectal cancer specimens used in the
present study.
| Characteristic | Number (%) |
|---|
| Sex |
|
| Male | 4 (66.7) |
|
Female | 2 (33.3) |
| Tumor size, cm |
|
|
<5 | 3 (50.0) |
| ≥5 | 3 (50.0) |
| Age, years |
|
|
<65 | 4 (66.7) |
| ≥65 | 2 (33.3) |
| Lymphatic
metastasis |
|
| No | 4 (66.7) |
| Yes | 2 (33.3) |
GGCT is associated with migration and
invasion according to GSEA enrichment analysis
As aforementioned, GGCT was demonstrated to be
upregulated in CRC tissues and was associated with a poor
prognosis. Therefore, the function of GGCT in CRC was further
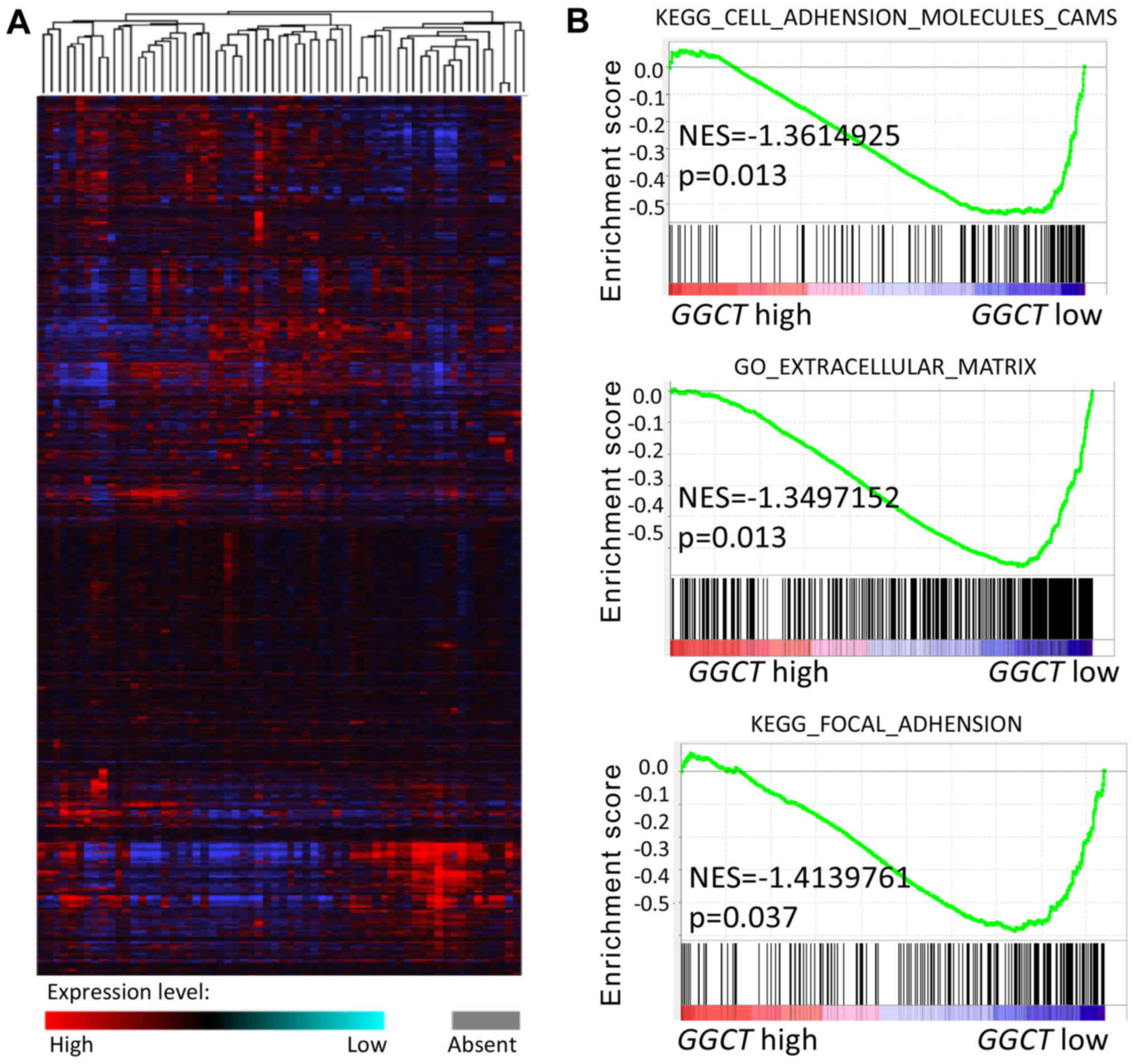
examined. A microarray of mRNA from 286 colorectal tumor samples
was downloaded from the TCGA Colorectal Cancer database (Fig. 2A). GSEA enrichment analysis was
subsequently performed and the results indicated that GGCT was
significantly associated with CRC migration and invasion in the
context of the KEGG_CELL_ADHENSION_MOLECULES_CAMS,
GO_EXTRACELLULAR_MATRIX and KEGG_FOCAL_ADHENSION gene sets
(P<0.05). Patients with CRC, which exhibited a high GGCT
expression, were indicated to exhibit an increased number of
EMT-associated genes in the context of the
HALLMARK_EPITHELIAL_MESENCHYMAL_TRANSITION gene set (Fig. 2B). These results revealed that GGCT
may regulate the migration and invasion of CRC cells via EMT.
GGCT promotes the migration and
invasion of CRC cells in vitro
GSEA enrichment analysis revealed that GGCT may
regulate the migration and invasion of CRC; therefore, to assess
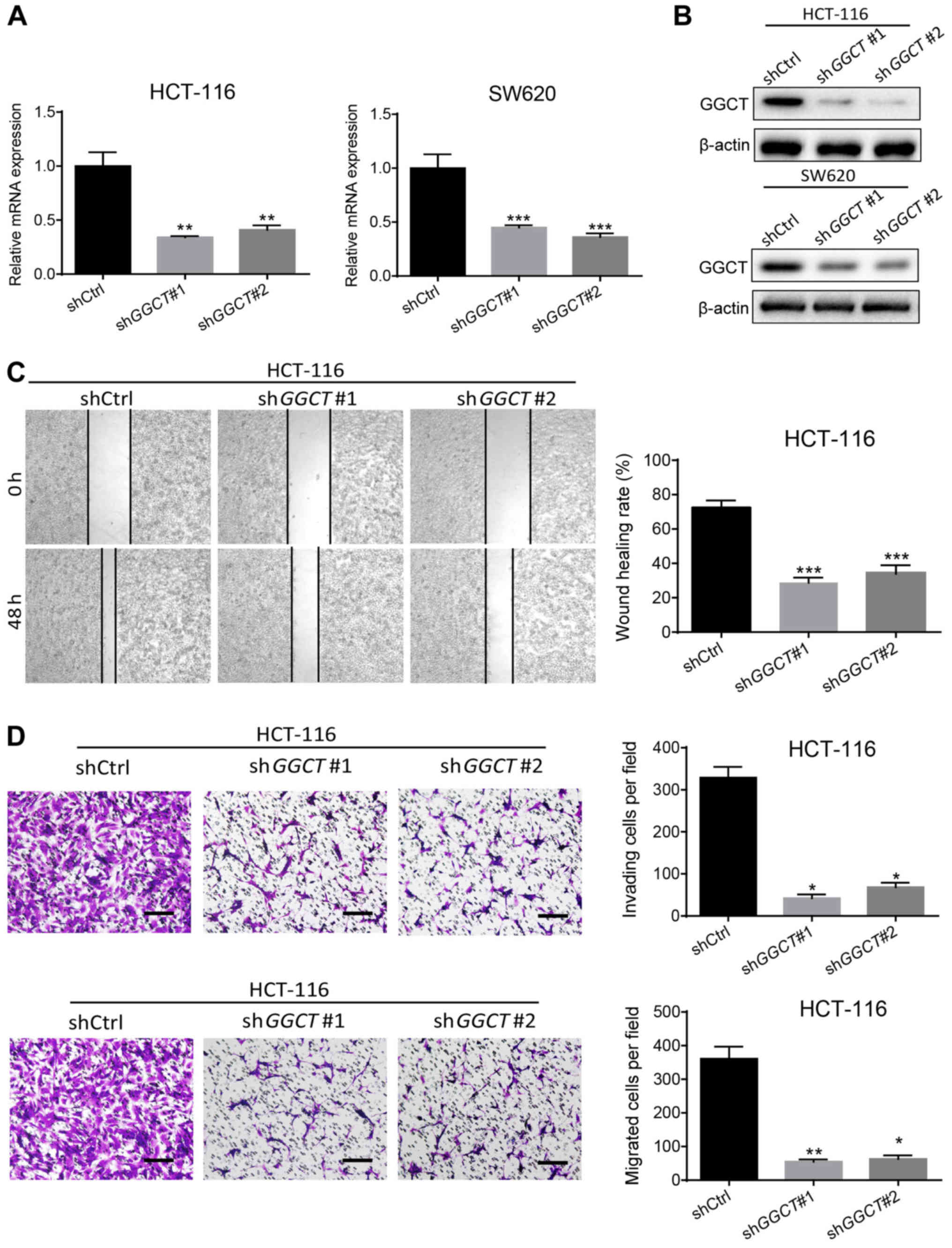
the function of GGCT in CRC malignancy, GGCT-knockdown colon cancer
cell lines HCT-116 and SW620 were established using a lentiviral
system. RT-qPCR and western blot analysis demonstrated that the
expression of GGCT significantly decreased in infected HCT-116 and
SW620 cells compared with corresponding control cells (P<0.01),
indicating that the interference effects of knocking down GGCT were
consistent (Fig. 3A and B). The
migratory ability of GGCT-knockdown cells was detected and analyzed
using a wound healing assay. In the GGCT-knockdown groups, a
significantly lower wound healing rate was observed compared with
the control cells (P<0.001; Fig.
3C). Furthermore, a transwell invasion and migration assay was
performed, which indicated that the disruption of GGCT
significantly inhibited the invasion and migration capability of
GGCT-knockdown HCT-116 cells compared with the control cells
(P<0.05; Fig. 3D). In conclusion,
the results demonstrated that GGCT promoted CRC migration and
invasion in vitro.
GGCT induces EMT in the colon cancer
cell line HCT-116
To determine the mechanism in which GGCT promoted
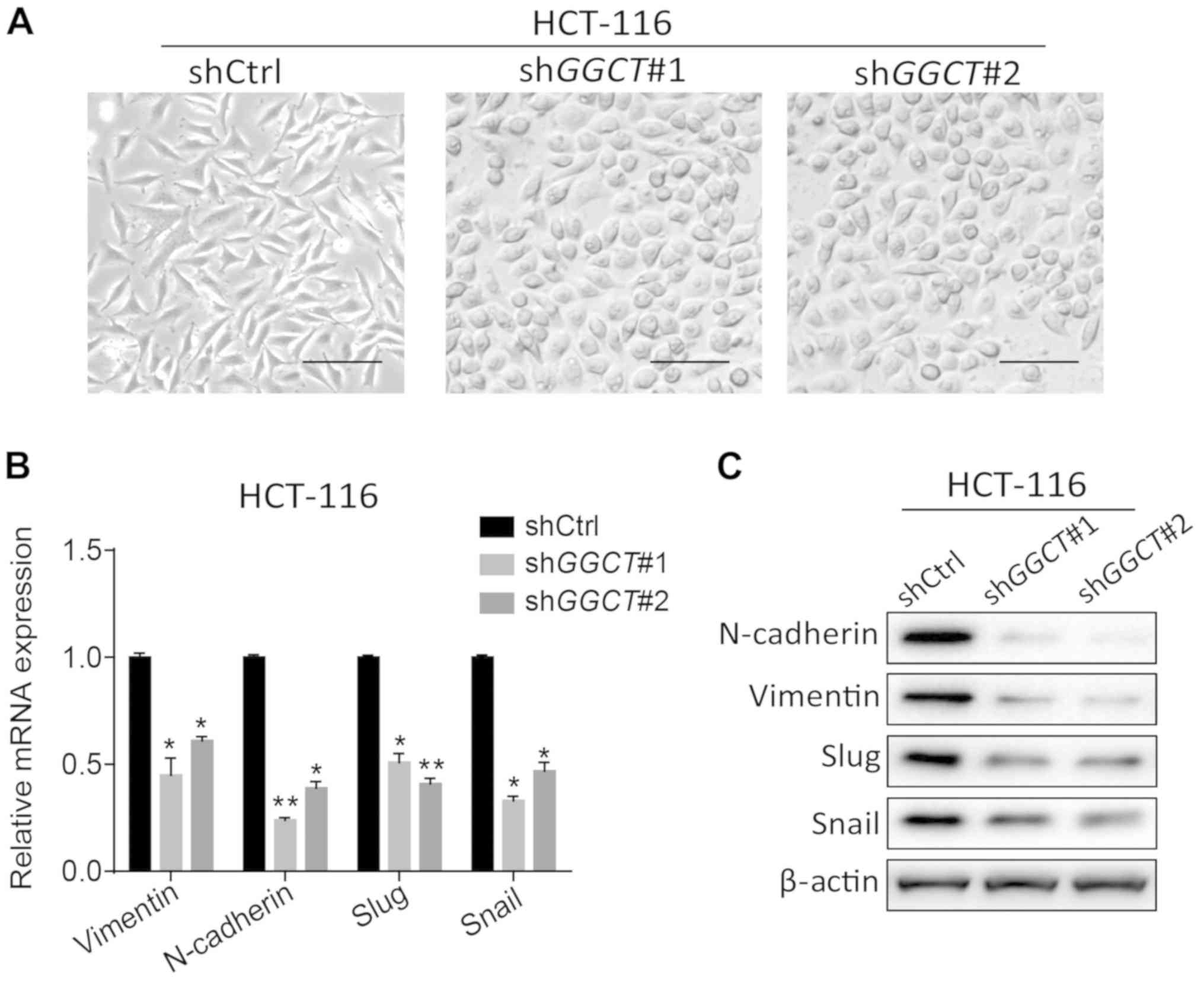
the migration and invasion of CRC cells, the present study assessed
whether the depletion of GGCT inhibited EMT in HCT-116 cells. The
cellular morphology was observed using a light microscope, and the
cell morphology of GGCT-knockdown HCT-116 cells was indicated to
have notably changed into the epithelial cell type from the
original HCT-116 mesenchymal-like cell type (Fig. 4A). EMT-associated biomarkers were
subsequently assessed in GGCT-knockdown HCT-116 cells, and it was
revealed that the depletion of GGCT significantly decreased the
expression of N-cadherin, Vimentin, Slug and Snail compared with
control cells (P<0.05; Fig. 4B and
C). Overall, these data demonstrated that GGCT induced EMT in
the colon cancer cell line HCT-116.
Discussion
CRC is the third most common malignancy, and the
fourth most common cause of cancer-associated mortality globally
(2). Despite substantial advances in
modern medicine, including the development of novel treatment
methods, the mortality of patients with CRC remains high, which may
be due to the lack of specific biomarkers and effective treatments
for the disease (23,24). Therefore the identification of novel
prognostic markers and therapeutic targets for use in CRC treatment
is urgently required.
EMT is essential for metastatic dissemination
(25). During EMT, tumor cells with
epithelial characteristics are transformed into tumor cells with
mesenchymal characteristics, which contributes to cancer
progression (26). It has recently
been accepted that EMT is an important and complex phenomenon that
determines the aggressiveness of colon cancer (27). Zhang et al (20) demonstrated that Prostate
Transmembrane Protein, Androgen Induced 1 induced EMT via a
non-canonical transforming growth factor-β signaling pathway in
CRC, and Wei et al (28)
indicated that FAT Atypical Cadherin 4 modulated CRC tumorigenesis
by regulating the status of EMT and autophagy (28). Sun et al (29) demonstrated that Tripartite Motif
Containing 29 facilitated the EMT of CRC via the activation of the
Wnt/β-catenin signaling pathway. The aforementioned results
indicated that elucidating the molecular mechanisms of EMT may aid
in the progression of CRC research and the identification of novel
therapeutic targets.
In the present study, GGCT was revealed to be
increased in CRC tissues compared with normal colon tissues, and
patients with CRC that had higher GGCT expression exhibited a worse
prognosis compared with patients with a lower expression of GGCT,
indicating the prognostic value of GGCT. The dysregulation of GGCT
has been reported in certain cancer types, for example Li et
al (13) demonstrated that GGCT
suppressed the progression of prostate cancer, indicating that GGCT
may serve as a tumor suppressor gene. However, Zhang et al
(30) indicated that the disruption
of GGCT inhibited proliferation and induced late apoptosis in human
gastric cancer, demonstrating that GGCT may serve as an oncogene.
The present study demonstrated that GGCT was highly expressed in
CRC and may function as a novel prognostic marker for the disease.
Furthermore, GGCT was indicated to promote CRC cell migration and
invasion through regulating EMT. The disruption of GGCT in HCT-116
cells markedly decreased the EMT associated markers and changed the
morphology of GGCT-knockdown HCT-116 cells from mesenchymal-like
cell type to the epithelial cell type, compared with the control
cells. Disrupting GGCT was also observed to notably decrease the
cell migration and invasion of HCT-116 cells, which was consistent
with previous reports that indicated that GGCT influenced the
migration and invasion of cancer cells (5,31).
GGCT has previously been described to serve an
important function in the proliferation of gastric, breast and lung
cancer cells (12,30,32). In
the present study, the proliferation of GGCT-knockdown CRC cells
was not assessed, nevertheless this part is worthy of further
work.
As GGCT has been reported to regulate the
penultimate step in glutathione catabolism and serve a critical
function in glutathione homeostasis, biochemical factors were not
assessed following the disruption of GGCT in CRC cells. However,
assessing how the depletion of GGCT influences EMT in CRC cells
requires investigation in future studies.
In the present study, GGCT was demonstrated to serve
a novel function as an oncogene in CRC, was demonstrated to promote
CRC migration and invasion via EMT and was identified as a novel
prognostic marker for this disease. In conclusion, the present
study provides novel insight into the mechanisms responsible for
CRC migration and invasion and identifies GGCT as a promising
therapeutic target for use in the treatment of CRC.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Science and
Technology Committee of Baoshan District, Shanghai City Special
Funds for Science, Technology and Innovation (grant no.
16-E-21).
Availability of data and materials
All data generated and/or analyzed during this study
are included in this published article.
Authors' contributions
QH and YZ completed all trial procedures and data
analysis in writing the manuscript. QH, YZ and YL performed the
experiments. ZL designed the overall idea of the experiment and
provided theoretical guidance throughout the process. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was ethically approved by the
Ethic Committee of Renhe Hospital (Shanghai, China). All subjects
provided written informed consent to participate in the present
study.
Patient consent for publication
Not applicable.
Competing interests
The authors have declared that they have no
competing interests.
References
|
1
|
Braillon A: Screening for colorectal
cancer at earlier ages: Putting the cart before the horse.
Gastroenterology. 156:15322019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Young GP, Rabeneck L and Winawer SJ: The
global paradigm shift in screening for colorectal cancer.
Gastroenterology. 156:843–851. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
de Andrea CE, Schalper KA, Sanmamed MF and
Melero I: Immunodivergence in metastatic colorectal cancer. Cancer
Cell. 34:876–878. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Huyghe JR, Bien SA, Harrison TA, Kang HM,
Chen S, Schmit SL, Conti DV, Qu C, Jeon J, Edlund CK, et al:
Discovery of common and rare genetic risk variants for colorectal
cancer. Nat Genet. 51:76–87. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kageyama S, Hanada E, Ii H, Tomita K,
Yoshiki T and Kawauchi A: Gamma-glutamylcyclotransferase: A novel
target molecule for cancer diagnosis and treatment. Biomed Res Int.
2015:3452192015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liu Y, Hyde AS, Simpson MA and Barycki JJ:
Emerging regulatory paradigms in glutathione metabolism. Adv Cancer
Res. 122:69–101. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Amano T, Eishi Y, Yamada T, Uchida K,
Minegishi K, Tamura T, Kobayashi D, Hiroshi K, Suzuki T and Board
PG: Widespread expression of gamma-glutamyl cyclotransferase
suggests it is not a general tumor marker. J Histochem Cytochem.
60:76–86. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kageyama S, Isono T, Iwaki H, Wakabayashi
Y, Okada Y, Kontani K, Yoshimura K, Terai A, Arai Y and Yoshiki T:
Identification by proteomic analysis of calreticulin as a marker
for bladder cancer and evaluation of the diagnostic accuracy of its
detection in urine. Clin Chem. 50:857–866. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Uejima D, Nishijo K, Kajita Y, Ishibe T,
Aoyama T, Kageyama S, Iwaki H, Nakamura T, Iida H, Yoshiki T and
Toguchida J: Involvement of cancer biomarker C7orf24 in the growth
of human osteosarcoma. Anticancer Res. 31:1297–1305.
2011.PubMed/NCBI
|
|
10
|
Takemura K, Kawachi H, Eishi Y, Kitagaki
K, Negi M, Kobayashi M, Uchida K, Inoue J, Inazawa J, Kawano T and
Board P: γ-Glutamylcyclotransferase as a novel immunohistochemical
biomarker for the malignancy of esophageal squamous tumors. Hum
Pathol. 45:331–341. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shen SH, Yu N, Liu XY, Tan GW and Wang ZX:
Gamma-glutamylcyclotransferase promotes the growth of human glioma
cells by activating Notch-Akt signaling. Biochem Biophys Res
Commun. 471:616–620. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Matsumura K, Nakata S, Taniguchi K, Ii H,
Ashihara E, Kageyama S, Kawauchi A and Yoshiki T: Depletion of
gamma-glutamylcyclotransferase inhibits breast cancer cell growth
via cellular senescence induction mediated by CDK inhibitor
upregulation. BMC Cancer. 16:7482016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li H, Yoshiya T, Nakata S, Taniguchi K,
Hidaka K, Tsuda S, Mochizuki M, Nishiuchi Y, Tsuda Y, Ito K, et al:
A Novel Prodrug of a γ-Glutamylcyclotransferase Inhibitor
Suppresses Cancer Cell Proliferation in vitro and Inhibits Tumor
Growth in a Xenograft Mouse Model of Prostate Cancer. ChemMedChem.
13:155–163. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ii H, Yoshiya T, Nakata S, Taniguchi K,
Hidaka K, Tsuda S, Mochizuki M, Nishiuchi Y, Tsuda Y, Ito K, et al:
A novel prodrug of a γ-glutamylcyclotransferase inhibitor
suppresses cancer cell proliferation in vitro and inhibits tumor
growth in a xenograft mouse model of prostate cancer. Chemmedchem.
13:155–163. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kanehisa M and Goto S: KEGG: Kyoto
encyclopedia of genes and genomes. Nucleic Acids Res. 28:27–30.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kanehisa M, Sato Y, Furumichi M, Morishima
K and Tanabe M: New approach for understanding genome variations in
KEGG. Nucleic Acids Res. 47:D590–D595. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kanehisa M: Toward understanding the
origin and evolution of cellular organisms. Protein Sci.
28:1947–1951. 2019. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ashburner M, Ball CA, Blake JA, Botstein
D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT,
et al: Gene ontology: Tool for the unification of biology. The Gene
Ontology Consortium. Nat Genet. 25:25–29. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
The Gene Ontology Consortium, . The Gene
Ontology resource: 20 years and still GOing strong. Nucleic Acids
Res. 47:D330–D338. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang L, Wang X, Lai C, Zhang H and Lai M:
PMEPA1 induces EMT via a non-canonical TGF-β signalling in
colorectal cancer. J Cell Mol Med. 23:3603–3615. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang R, Hoang BH, Kubo T, Kawano H, Chou
A, Sowers R, Huvos AG, Meyers PA, Healey JH and Gorlick R:
Over-expression of parathyroid hormone Type 1 receptor confers an
aggressive phenotype in osteosarcoma. Int J Cancer. 121:943–954.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ganesh K, Stadler ZK, Cercek A, Mendelsohn
RB, Shia J, Segal NH and Diaz LJ Jr: Immunotherapy in colorectal
cancer: Rationale, challenges and potential. Nat Rev Gastroenterol
Hepatol. 16:361–375. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Valle L, de Voer RM, Goldberg Y, Sjursen
W, Försti A, Ruiz-Ponte C, Caldés T, Garré P, Olsen MF, Nordling M,
et al: Update on genetic predisposition to colorectal cancer and
polyposis. Mol Aspects Med. 69:10–26. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pastushenko I and Blanpain C: EMT
transition states during tumor progression and metastasis. Trends
Cell Biol. 29:212–226. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Singh M, Yelle N, Venugopal C and Singh
SK: EMT: Mechanisms and therapeutic implications. Pharmacol Ther.
182:80–94. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhu Y, Wang C, Becker SA, Hurst K,
Nogueira LM, Findlay VJ and Camp ER: miR-145 antagonizes
SNAI1-mediated stemness and radiation resistance in colorectal
cancer. Mol Ther. 26:744–754. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wei R, Xiao Y, Song Y, Yuan H, Luo J and
Xu W: FAT4 regulates the EMT and autophagy in colorectal cancer
cells in part via the PI3K-AKT signaling axis. J Exp Clin Cancer
Res. 38:1122019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sun J, Zhang T, Cheng M, Hong L, Zhang C,
Xie M, Sun P, Fan R, Wang Z, Wang L and Zhong J: TRIM29 facilitates
the epithelial-to-mesenchymal transition and the progression of
colorectal cancer via the activation of the Wnt/β-catenin signaling
pathway. J Exp Clin Cancer Res. 38:1042019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang W, Chen L, Xiang H, Hu C, Shi W,
Dong P and Lv W: Knockdown of GGCT inhibits cell proliferation and
induces late apoptosis in human gastric cancer. BMC Biochem.
17:192016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kageyama S, Ii H, Taniguchi K, Kubota S,
Yoshida T, Isono T, Chano T, Yoshiya T, Ito K, Yoshiki T, et al:
Mechanisms of tumor growth inhibition by depletion of
γ-glutamylcyclotransferase (GGCT): A novel molecular target for
anticancer therapy. Int J Mol Sci. 19:E20542018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li Y, Zhao W, Zhao Z, Wu J, Chen L, Ma Y,
Li Q, Lu D, Jin L and Wang J: IL1B gene polymorphisms, age and the
risk of non-small cell lung cancer in a Chinese population. Lung
Cancer. 89:232–237. 2015. View Article : Google Scholar : PubMed/NCBI
|