Introduction
Lung cancer includes non-small cell lung cancer
(NSCLC) and small cell lung cancer (SCLC). The difference between
NSCLC and SCLC is not defined by size, but by pathology (1). The incidence of NSCLC is higher and
more common than SCLC. In addition, patients with NSCLC have a
lower degree of malignancy and better treatment outcome (2). Currently, the treatment of NSCLC is
based on the clinical stage of lung cancer. The NSCLC patients in
stage I, II, and IIIA are primarily surgically resected. For
patients with significant lymphatic metastasis, chemotherapy or
radiotherapy can be performed prior to surgery (3). However, the disease is prone to
recurrence after surgery, and the prognosis of patients with
advanced NSCLC is poor (4).
Therefore, NSCLC still faces enormous challenges in diagnosis and
treatment.
A large number of studies have shown that microRNAs
(miRNAs) are involved in organ development or pathological
processes in humans and animals (5).
miRNA exerts its effects by binding to the 3′- untranslated region
of the target mRNA, thereby inhibiting gene expression or degrading
proteins. In various human cancers, the difference in miRNA
function depends on the type of cancer or the difference in
downstream genes involved (6). At
present, the dysregulation of miR-147 has been widely investigated
in several cancers. It has been reported that miR-147 was
upregulated in gastric cancer and played a carcinogenic role
(7). Sui et al (8) found that miR-147 suppressed
proliferation and migration of human hepatocellular carcinoma cells
by inhibiting homeobox C6 (HOXC6). These studies suggested that
miR-147 has tissue specificity in human cancers. In addition,
miR-147 was found to be a diagnostic biomarker for human NSCLC
(9). However, the biological
function and corresponding mechanism of miR-147 remains unclear in
NSCLC and needs to be explored. In addition, few studies have shown
that miRNAs are involved in NSCLC by regulating phosphoinositide
3-kinase (PI3K)/protein kinase B (AKT) pathway (10). In other cancers, miR-383 suppressed
the development of cervical cancer via downregulating
poly(ADP-ribose) polymerase-2 (PARP-2) and regulating the PI3K/AKT
signaling pathway (11).
Furthermore, miR-147 was found to inhibit AKT phosphorylation in
HCT116 colon carcinoma cells (12).
Therefore, we suspected that miR-147 may regulate the PI3K/AKT
pathway in NSCLC cells.
As a member of the neurotrophin family,
brain-derived neurotrophic factor (BDNF) is involved in
neurotrophins and neuronal differentiation (13). Furthermore, it has been reported that
the PI3K/AKT pathway is activated by the BDNF/tyrosine kinase B
(TRKB) and BDNF/p75NTR signaling system (14). The function of BDNF has been found in
various cancers. For example, BDNF promoted migration and survival
of clear cell renal cell carcinoma cells (15). In addition, BDNF was found to promote
cell metastasis in human colon cancer (16). In particular, BDNF was associated
with poor prognosis in NSCLC patients (17). Upregulation of BDNF has been reported
to promote proliferation and invasion of lung squamous cell
carcinoma cells (18). However, the
relationship between miR-147 and BDNF has not been reported in
NSCLC.
In this study, we focused on the effects of miR-147
on NSCLC cell viability and metastasis. In addition, how miR-147
regulates BDNF and the PI3K/AKT pathway was also explored in NSCLC.
miR-147 may have diagnostic and therapeutic value for NSCLC.
Patients and methods
Experimental samples
The tissues used in this study were obtained from 79
patients with NSCLC in People's Hospital of Rizhao (Rizhao, China).
All NSCLC patients enrolled in this study were treated only with
surgery. Participants provided written informed consent and the
study was approved by the Institutional Ethics Committee of
People's Hospital of Rizhao.
Cell culture and transfection
Human bronchial epithelial cells (16HBE) and A549
NSCLC cells were obtained from the American Type Culture Collection
(ATCC). Next, these cells were incubated in RPMI-1640 medium
(Gibco; Thermo Fisher Scientific, Inc.) with 10% fetal bovine serum
(FBS). A549 cells were then transfected with miR-147 mimics or
inhibitor as well as BDNF vector (Genechem), respectively, using
Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific,
Inc.).
Real-time quantitative polymerase
chain reaction (RT-qPCR)
Total RNA isolation was performed using TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc). The cDNA
solution was synthesized using PrimeScript RT reagent (Takara).
RT-qPCR was performed on ABI 7500 thermocycler (Applied Biosystems)
using SYBR-Green Premix Ex Taq II (Takara). miR-147 or BDNF was
normalized by U6 or GAPDH as an endogenous control. Their
expression levels were calculated using the 2−ΔΔct
method. The primers used were: miR-147 forward,
5′-CCCCTATCACGATTAGCATTAA-3′ and reverse,
5′-CCCAAGCTTTTATGTGGTTGTTACTATGC-3′; U6 forward,
5′-CTCGCTTCGGCAGCACA-3′ and reverse, 5′-AACGCTTCACGAATTTGCGT-3′;
GAPDH forward, 5′-GAAGGTGAAGGTCGGAGTC-3′ and reverse,
5′-GAAGATGGTGATGGGATTTC-3′; and BDNF forward,
5′-CTACGAGACCAAGTGCAATCC-3′ and reverse,
5′-AATCGCCAGCCAATTCTCTTT-3′.
Methyl thiazolyl tetrazolium (MTT)
assay
First, transfected A549 cells (4×103
cells/well) were prepared in 96-well plates. Then, A549 cells were
incubated in fresh medium for 24, 48, 72 or 96 h, respectively.
Next, 10 µl MTT solution was added. The cells were cultured for 4
h. The MTT solution was aspirated and the Formazan solution was
added to fully dissolve the crystals. The absorbance at 490 nm was
examined by a microscope (Olympus Corp.).
Transwell assay
Invasion assay was performed in the upper chamber
with Matrigel (BD Biosciences). Transfected cells (4×103
cells/well) were put in the upper chamber, and lower chamber was
filled with 10% FBS. Next, the cells were fixed and stained. The
cell migration assay does not require Matrigel and other procedures
are identical to the cell invasion assay. Finally, the migrated and
invaded cells were counted using a microscope (Olympus Corp.).
Luciferase reporter assay
First, we constructed a pcDNA3.1 plasmid vector
(Promega) containing wild-type or mutant 3′-UTR of BDNF. Next, A549
cells were transfected with the above plasmid and miR-147 mimics
and were incubated for 48 h. Finally, luciferase activity was
measured using a dual luciferase assay system (Promega).
Western blot analysis
Protein samples were lysed using Radio
immunoprecipitation assay (RIPA) lysis buffer (Beyotime). Next, 10%
sodium salt-polyacrylamide gel electrophoresis (SDS-PAGE) protein
loading buffer was added to the collected protein samples. After
denaturation of the protein, the protein sample was directly loaded
into SDS-PAGE gel and transferred into polyvinylidene difluoride
(PVDF) membranes. Next, primary antibodies (Vimentin, N-cadherin,
E-cadherin, PI3K, AKT, p-PI3K, p-AKT and GAPDH) were added, and the
membrane was incubated overnight at 4°C. The wash solution was
added for 5–10 min. Next, the diluted secondary antibody was added
and incubated for 1 h at room temperature. Finally, the protein was
examined using an Enhanced chemiluminescence reagent (ECL; Pierce;
Thermo Fisher Scientific, Inc.).
Statistical analysis
Data are shown as mean ± SD and analyzed using
Statistical Product and Service Solutions (SPSS) 18.0 (SPSS, Inc.)
or GraphPad Prism 6 (GraphPad Systems). Statistical analysis was
performed using Chi-square test or one-way analysis of variance
(ANOVA) followed by Fisher's least significant difference. The
correlation between miR-147 and BDNF expression was examined by
Pearson's correlation analysis. The overall survival rates and
survival differences were analyzed using the Univariate
Kaplan-Meier method followed by log-rank test. P<0.05 was
considered to be a statistically significant difference.
Results
miR-147 expression is decreased in
NSCLC tissues
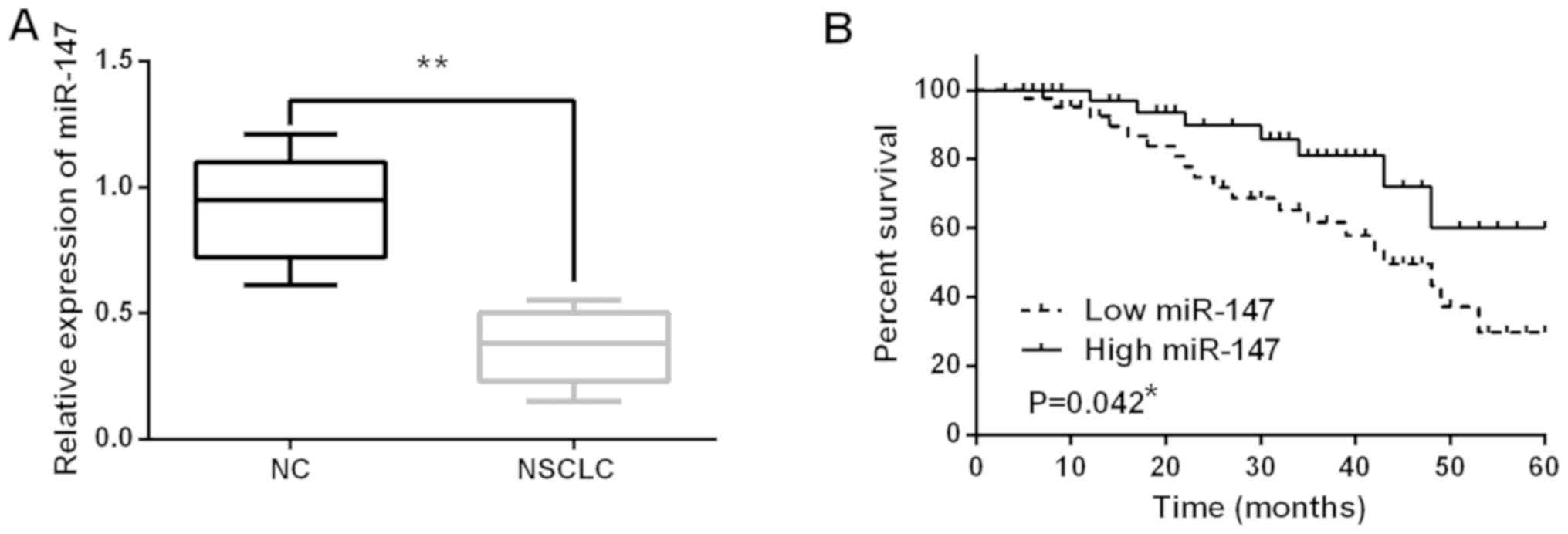
The alteration of miR-147 expression was observed in
NSCLC tissues using RT-qPCR. miR-147 expression was decreased in
NSCLC tissues compared to normal tissues (P<0.01; Fig. 1A). Next, the association between
miR-147 expression and clinical features in NSCLC tissues was
analyzed. It was found that low expression of miR-147 was
associated with poor clinical features (including lymph nodes
metastasis or tumor stage) in NSCLC patients (Table I). In addition, poor prognosis in
NSCLC patients was found to be associated with low miR-147
expression (P=0.042; Fig. 1B). These
results indicate that miR-147 is involved in pathogenesis of
NSCLC.
 | Table I.Relationship between miR-147
expression and clinico-pathological characteristics of NSCLC
patients. |
Table I.
Relationship between miR-147
expression and clinico-pathological characteristics of NSCLC
patients.
|
|
| miR-147 |
|
|---|
|
|
|
|
|
|---|
| Characteristics | Cases | High | Low | P-value |
|---|
| Age (years) |
|
|
| 0.06 |
| ≥60 | 44 | 15 | 29 |
|
|
<60 | 35 | 14 | 21 |
|
| Sex |
|
|
| 0.13 |
| Male | 43 | 12 | 31 |
|
|
Female | 36 | 17 | 19 |
|
| Tumor size (mm) |
|
|
| 0.22 |
| ≤3 | 52 | 20 | 32 |
|
|
>3 | 27 | 9 | 18 |
|
| Lymph nodes
metastasis |
|
|
| 0.03a |
|
Yes | 17 | 5 | 12 |
|
| No | 62 | 24 | 38 |
|
| Tumor stage |
|
|
| 0.02a |
|
I–II | 57 | 21 | 36 |
|
|
III–IV | 22 | 8 | 14 |
|
Overexpression of miR-147 suppresses
NSCLC cell viability and metastasis
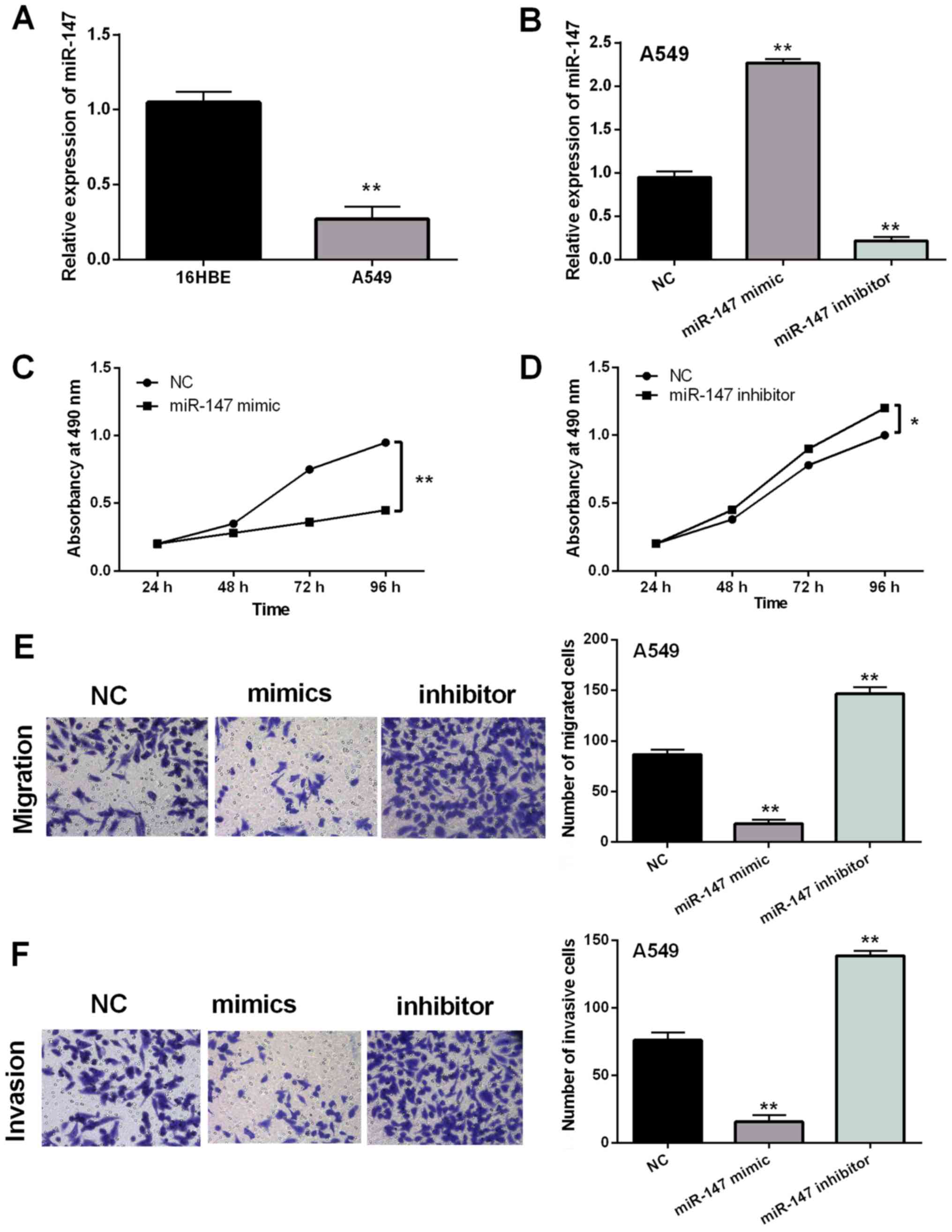
Next, the expression of miR-147 was examined in A549
and 16HBE cell lines. Similarly, downregulation of miR-147 was also
assessed in A549 cells compared to 16HBE cells (P<0.01; Fig. 2A). The role of miR-147 in NSCLC was
explored in A549 cells with miR-147 mimics or inhibitor. It was
found that expression of miR-147 was significantly regulated by
miR-147 mimics or inhibitor (P<0.01; Fig. 2B). MTT assay indicated that
overexpression of miR-147 suppressed proliferation of A549 cells
(P<0.01; Fig. 2C). In contrast,
downregulation of miR-147 was found to promote cell proliferation
(P<0.05; Fig. 2D). In addition,
overexpression of miR-147 inhibited cell migration, whereas miR-147
silencing promoted cell migration in A549 cells (P<0.01;
Fig. 2E). Similar results on cell
invasion were also detected in A549 cells with miR-147 mimics or
inhibitor (P<0.01; Fig. 2F).
Based on these results, it was considered that overexpression of
miR-147 suppressed the viability and metastasis of NSCLC cells.
miR-147 blocks epithelial-mesenchymal
transition (EMT) and inactivates the PI3K/AKT pathway in NSCLC
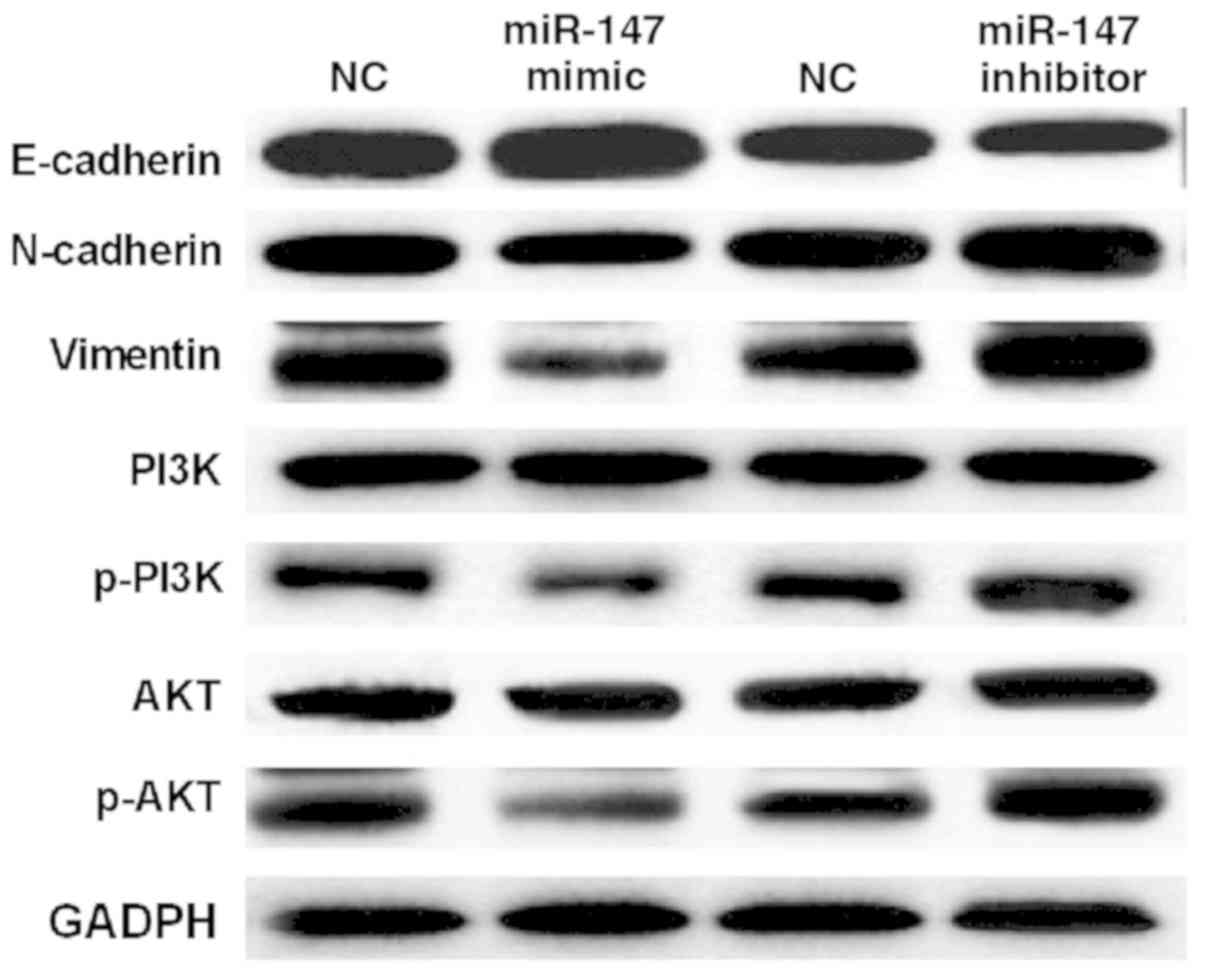
To further explain the above conclusion, how miR-147
regulates EMT and the PI3K/AKT pathway was investigated. It was
found that miR-147 mimics suppressed the expression of Vimentin and
N-cadherin and enhanced E-cadherin expression level in A549 cells
(Fig. 3). In contrast, Vimentin and
N-cadherin expressions were improved by downregulation of miR-147.
E-cadherin expression was suppressed by miR-147 silencing in A549
cells (Fig. 3). Next, it was
investigated how miR-147 regulates the PI3K/AKT pathway in A549
cells. Western blot analysis showed that miR-147 mimics
downregulated p-PI3K and p-AKT expression in A549 cells (Fig. 3). miR-147 silencing was found to
promote expression of p-PI3K and p-AKT. However, miR-147 had no
effect on the expression of PI3K and AKT in A549 cells (Fig. 3). These results demonstrate that
miR-147 blocks EMT and inactivates the PI3K/AKT pathway in
NSCLC.
BDNF is a direct target of
miR-147
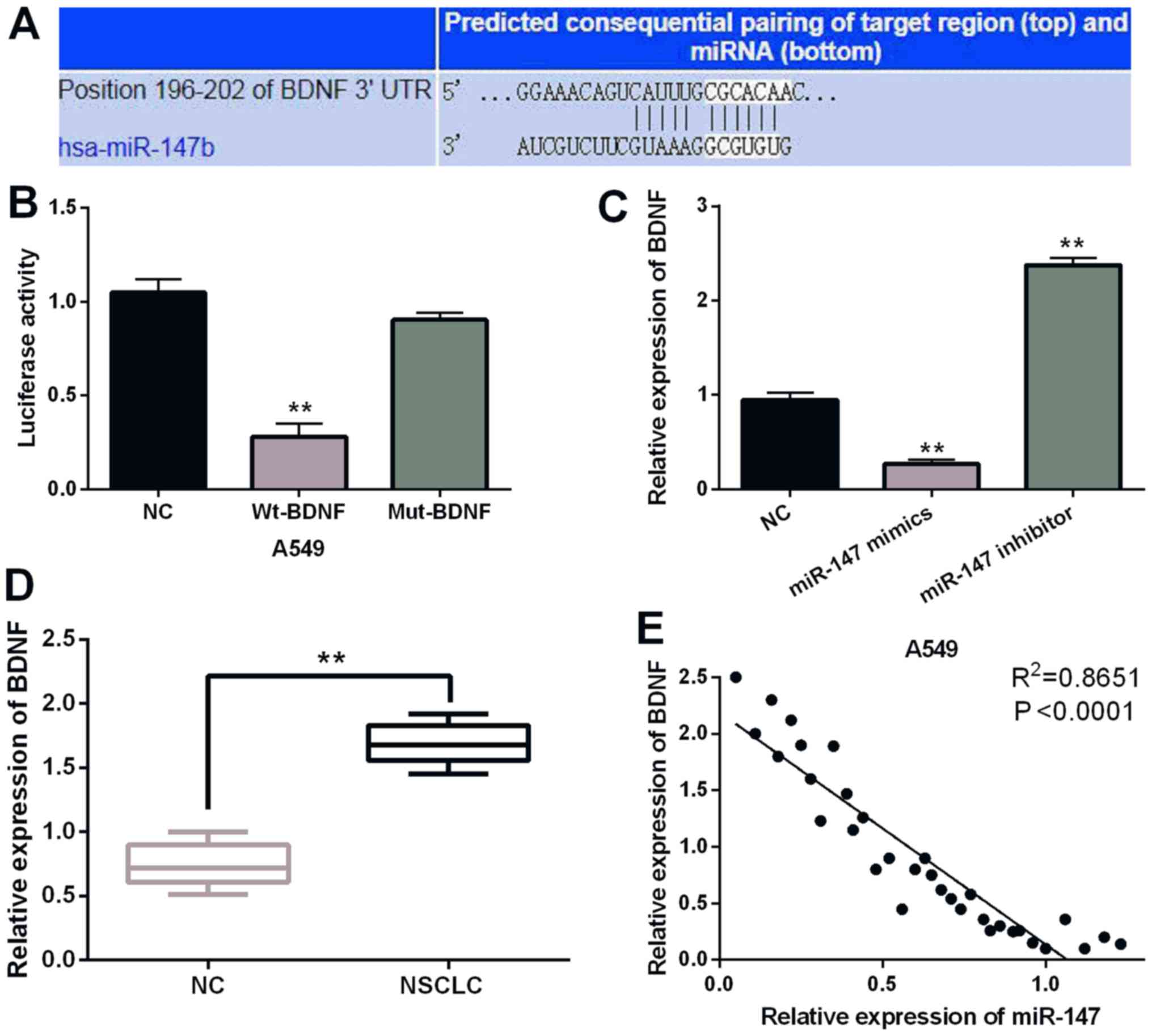
To further disclose the molecular mechanism of
miR-147 in NSCLC, the target gene of miR-147 was searched in the
TargetScan (http://www.targetscan.org/) databases. It is predicted
that miR-147 has a binding site with the 3′-UTR of BDNF (Fig. 4A). It was found that miR-147 mimics
significantly inhibited Wt-BDNF luciferase activity, whereas the
luciferase activity of Mut-BDNF was not regulated by miR-147 mimics
(P<0.01; Fig. 4B). Then, BDNF
expression was observed in A549 cells containing miR-147 mimics or
inhibitor. Overexpression of miR-147 suppressed the expression of
BDNF. miR-147 silencing promoted BDNF expression (P<0.01;
Fig. 4C). BDNF expression was
detected in NSCLC tissues. RT-qPCR indicated that BDNF was
upregulated in NSCLC tissues compared to normal expression
(P<0.01; Fig. 4D). Furthermore, a
negative correlation between expression of miR-147 and BDNF was
identified in NSCLC tissues (P<0.0001; R2=0.8651;
Fig. 4E). Collectively, miR-147
directly targets BDNF and negatively regulates BDNF expression in
NSCLC.
Upregulation of BDNF impairs the
inhibitory effect of miR-147 in NSCLC
Finally, BDNF vectors were transfected into A549
cells with miR-147 mimics to explore their interactions. RT-qPCR
showed that BDNF vector restored the decreased expression of BDNF
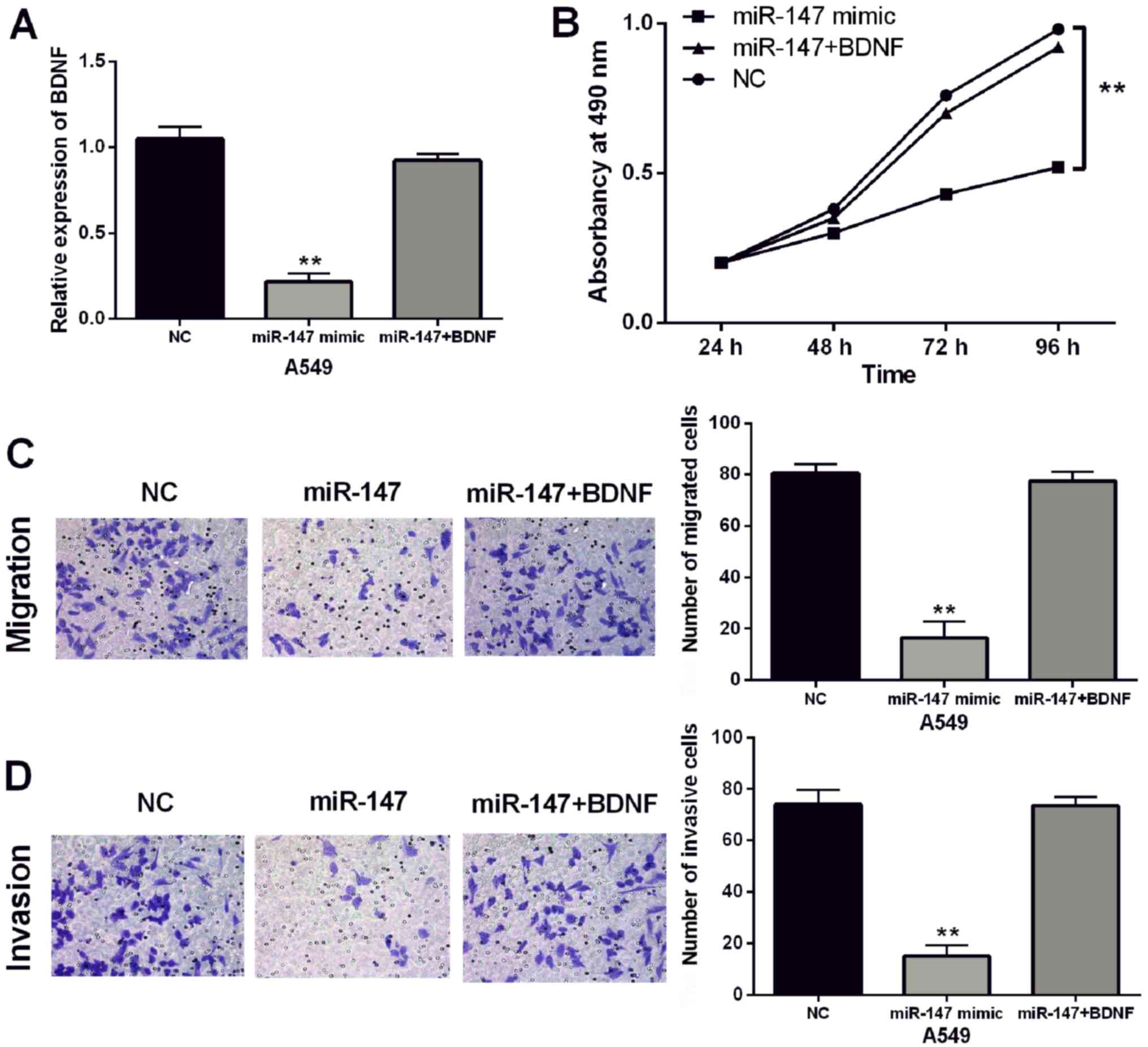
induced by miR-147 mimics in A549 cells (P<0.01; Fig. 5A). Furthermore, BDNF vectors
attenuated miR-147-induced inhibition of cell proliferation
(P<0.01; Fig. 5B). In addition,
BDNF vectors also abolished the inhibitory effect of miR-147 on
cell migration and invasion in A549 cells (P<0.01; Fig. 5C and D). Taken together, upregulation
of BDNF attenuated the inhibitory effect of miR-147 in NSCLC.
Discussion
In this study, it was found that miR-147 expression
was decreased in NSCLC tissues, which was associated with poor
prognosis in NSCLC patients. In addition, low expression of miR-147
was associated with lymph node metastasis or tumor stage in NSCLC
patients. Chu et al (9) also
found similar results. However, miR-147 was upregulated in gastric
cancer, and downregulation of miR-147 inhibited cell proliferation
(19), which is different from the
present results. Upregulation of miR-147 suppressed NSCLC cell
viability and metastasis. The difference may be due to the
different tumor tissues. Moreover, ectopic expression of miR-147
was found to inhibit epithelial-mesenchymal transition
(EMT)-related protein expression in colon cancer cells (20), which is similarly to our results. Our
study also found that miR-147 suppressed Vimentin and N-cadherin
expression and promoted E-cadherin expression to block EMT in
NSCLC. Moreover, miR-147 has been reported to suppress breast
cancer cell proliferation, invasion and migration through the
PI3K/AKT/mTOR signaling pathway (21). In NSCLC cells, miR-147 was also
involved in the PI3K/AKT pathway by suppressing expression of
p-PI3K and p-AKT.
EMT and the PI3K/AKT pathway have been reported to
be involved in the pathogenesis of NSCLC. For example, miR-149
promoted the expression of E-cadherin in NSCLC (22). Herein, E-cadherin expression was also
promoted by miR-147 in NSCLC cells. Furthermore, it has been
reported that miR-3188 inhibited NSCLC cell proliferation by
regulating p-PI3K and p-AKT expression (23). In addition, PI3K and AKT are key
regulators of cell survival, which are critical for cancer
development (24,25). Therefore, the expression of PI3K and
AKT was detected in A549 cells with miR-147 mimics or inhibitor. It
was found that miR-147 suppressed p-PI3K and p-AKT expression to
inactivate the PI3K/AKT pathway in NSCLC, which agrees with
previous studies. For example, miR-107 has been found to inhibit
cell metastasis and tumor growth by inhibiting BDNF and mediating
the PI3K/AKT pathway in NSCLC (26).
The results suggest that BDNF may be involved in the PI3K/AKT
pathway in NSCLC. In the current study, miR-147 was found to
directly target BDNF and to negatively regulate BDNF expression in
NSCLC. Furthermore, overexpression of miR-147 suppressed p-PI3K and
p-AKT expression in NSCLC. Based on these results, we suspect that
BDNF can regulate the PI3K/AKT pathway by mediating miR-147.
However, this regulatory mechanism is complex. Therefore, further
exploration of the regulatory mechanism of miR-147/BDNF/EMT/
PI3K/AKT pathway in NSCLC is required.
Numerous studies have shown that BDNF exerts
carcinogenic effects in a variety of cancers. Here, upregulation of
BDNF was detected in NSCLC tissues. Similarly, upregulation of BDNF
has also been found in breast cancer and colorectal cancer
(27,28). As a target gene, BDNF was found to be
regulated by several miRNAs, such as miR-15a and miR-613 (29,30). In
the present study, miR-147 was also found to negatively regulate
BDNF expression in NSCLC. Upregulation of BDNF attenuated the
inhibitory effect of miR-147 on cell proliferation, invasion and
migration in NSCLC. Similarly, BDNF has been reported to impair the
inhibitory effect of miR-497 on cell invasion and tumor growth in
thyroid cancer (31). Furthermore,
miR-613 inhibited carcinogenesis of gastric cancer by inhibiting
BDNF expression (32). miR-147 also
inhibited the progression of NSCLC by targeting BDNF.
In conclusion, miR-147 exerts an inhibitory effect
in NSCLC. Moreover, miR-147 is involved in tumorigenesis of NSCLC
by targeting BDNF or blocking EMT and PI3K/AKT pathway. This study
initially indicates that miR-147 may be a promising therapeutic
option for NSCLC.
Acknowledgements
Not applicable.
Funding
Not funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
FL designed the study and wrote the manuscript; XW
performed the data analyses; LY contributed to the conception of
the study. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The study was approved by the Institutional Ethics
Committee of People's Hospital of Rizhao (Rizhao, China).
Participants provided written informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Oser MG, Niederst MJ, Sequist LV and
Engelman JA: Transformation from non-small-cell lung cancer to
small-cell lung cancer: Molecular drivers and cells of origin.
Lancet Oncol. 16:e165–e172. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mornex F and Girard N: Gemcitabine and
radiation therapy in non-small cell lung cancer: State of the art.
Ann Oncol. 17:1743–1747. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Osugi J, Muto S, Matsumura Y, Higuchi M,
Suzuki H and Gotoh M: Prognostic impact of the high-sensitivity
modified Glasgow prognostic score in patients with resectable
non-small cell lung cancer. J Cancer Res Ther. 12:945–951. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Novaes FT, Cataneo DC, Ruiz Junior RL,
Defaveri J, Michelin OC and Cataneo AJ: Lung cancer: Histology,
staging, treatment and survival. J Bras Pneumol. 34:595–600. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kloosterman WP and Plasterk RH: The
diverse functions of microRNAs in animal development and disease.
Dev Cell. 11:441–450. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shenouda SK and Alahari SK: MicroRNA
function in cancer: Oncogene or a tumor suppressor? Cancer
Metastasis Rev. 28:369–378. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yao Y, Suo AL, Li ZF, Liu LY, Tian T, Ni
L, Zhang WG, Nan KJ, Song TS and Huang C: MicroRNA profiling of
human gastric cancer. Mol Med Rep. 2:963–970. 2009.PubMed/NCBI
|
|
8
|
Sui CJ, Xu F, Shen WF, Dai BH, Lu JJ,
Zhang MF and Yang JM: MicroRNA-147 suppresses human hepatocellular
carcinoma proliferation migration and chemosensitivity by
inhibiting HOXC6. Am J Cancer Res. 6:2787–2798. 2016.PubMed/NCBI
|
|
9
|
Chu G, Zhang J and Chen X: Serum level of
microRNA-147 as diagnostic biomarker in human non-small cell lung
cancer. J Drug Target. 24:613–617. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang Y, Zhao M, Liu J, Sun Z, Ni J and Liu
H: miRNA-125b regulates apoptosis of human non-small cell lung
cancer via the PI3K/Akt/GSK3β signaling pathway. Oncol Rep.
38:1715–1723. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Teng P, Jiao Y, Hao M and Tang X:
microRNA-383 suppresses the PI3K-AKT-MTOR signaling pathway to
inhibit development of cervical cancer via down-regulating PARP2. J
Cell Biochem. 119:5243–5252. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lee CG, McCarthy S, Gruidl M, Timme C and
Yeatman TJ: MicroRNA-147 induces a mesenchymal-to-epithelial
transition (MET) and reverses EGFR inhibitor resistance. PLoS One.
9:e845972014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
McAllister AK: Neurotrophins and neuronal
differentiation in the central nervous system. Cell Mol Life Sci.
58:1054–1060. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sandhya VK, Raju R, Verma R, Advani J,
Sharma R, Radhakrishnan A, Nanjappa V, Narayana J, Somani BL,
Mukherjee KK, et al: A network map of BDNF/TRKB and BDNF/p75NTR
signaling system. J Cell Commun Signal. 7:301–307. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
De la Cruz-Morcillo MA, Berger J,
Sánchez-Prieto R, Saada S, Naves T, Guillaudeau A, Perraud A,
Sindou P, Lacroix A, Descazeaud A, et al: p75 neurotrophin receptor
and pro-BDNF promote cell survival and migration in clear cell
renal cell carcinoma. Oncotarget. 7:34480–34497. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Huang SM, Lin C, Lin HY, Chiu CM, Fang CW,
Liao KF, Chen DR and Yeh WL: Brain-derived neurotrophic factor
regulates cell motility in human colon cancer. Endocr Relat Cancer.
22:455–464. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Okamura K, Harada T, Wang S, Ijichi K,
Furuyama K, Koga T, Okamoto T, Takayama K, Yano T and Nakanishi Y:
Expression of TrkB and BDNF is associated with poor prognosis in
non-small cell lung cancer. Lung Cancer. 78:100–106. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang SY, Hui LP, Li CY, Gao J, Cui ZS and
Qiu XS: More expression of BDNF associates with lung squamous cell
carcinoma and is critical to the proliferation and invasion of lung
cancer cells. BMC Cancer. 16:1712016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shen J, Niu W and Zhang H, Jun M and Zhang
H: Downregulation of microRNA-147 inhibits cell proliferation and
increases the chemosensitivity of gastric cancer cells to
5-fluorouracil by directly targeting PTEN. Oncol Res. 26:901–911.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ning X, Wang C, Zhang M and Wang K:
Ectopic expression of miR-147 inhibits stem cell marker and
epithelial-mesenchymal transition (EMT)-related protein expression
in colon cancer cells. Oncol Res. 27:399–406. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang Y, Zhang HE and Liu Z: MicroRNA-147
suppresses proliferation, invasion and migration through the
AKT/mTOR signaling pathway in breast cancer. Oncol Lett.
11:405–410. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ke Y, Zhao W, Xiong J and Cao R: miR-149
inhibits non-small-cell lung cancer cells EMT by targeting FOXM1.
Biochem Res Int. 2013:5067312013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang C, Liu E, Li W, Cui J and Li T:
miR-3188 inhibits non-small cell lung cancer cell proliferation
through FOXO1-mediated mTOR-p-PI3K/AKT-c-JUN signaling pathway.
Front Pharmacol. 9:13622018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Samuels Y, Wang Z, Bardelli A, Silliman N,
Ptak J, Szabo S, Yan H, Gazdar A, Powell SM, Riggins GJ, et al:
High frequency of mutations of the PIK3CA gene in human cancers.
Science. 304:5542004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Datta SR, Brunet A and Greenberg ME:
Cellular survival: A play in three Akts. Genes Dev. 13:2905–2927.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xia H, Li Y and Lv X: MicroRNA-107
inhibits tumor growth and metastasis by targeting the BDNF-mediated
PI3K/AKT pathway in human non-small lung cancer. Int J Oncol.
49:1325–1333. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kang HJ, Kim JM, Kim SY, Kim SW, Shin IS,
Kim HR, Park MH, Shin MG, Yoon JH and Yoon JS: A longitudinal study
of BDNF promoter methylation and depression in breast cancer.
Psychiatry Investig. 12:523–531. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tanaka K, Okugawa Y, Toiyama Y, Inoue Y,
Saigusa S, Kawamura M, Araki T, Uchida K, Mohri Y and Kusunoki M:
Brain-derived neurotrophic factor (BDNF)-induced
tropomyosin-related kinase B (Trk B) signaling is a potential
therapeutic target for peritoneal carcinomatosis arising from
colorectal cancer. PLoS One. 9:e964102014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Long J, Jiang C, Liu B, Fang S and Kuang
M: MicroRNA-15a-5p suppresses cancer proliferation and division in
human hepatocellular carcinoma by targeting BDNF. Tumour Biol.
37:5821–5828. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li W, Li X, Xin X, Kan PC and Yan Y:
MicroRNA-613 regulates the expression of brain-derived neurotrophic
factor in Alzheimer's disease. Biosci Trends. 10:372–377. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang P, Meng X, Huang Y, Lv Z, Liu J, Wang
G, Meng W, Xue S, Zhang Q, Zhang P, et al: MicroRNA-497 inhibits
thyroid cancer tumor growth and invasion by suppressing BDNF.
Oncotarget. 8:2825–2834. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ding D, Hou R, Gao Y and Feng Y: miR-613
inhibits gastric cancer progression through repressing brain
derived neurotrophic factor. Exp Ther Med. 15:1735–1741.
2018.PubMed/NCBI
|