Introduction
Colorectal cancer (CRC) is the third most common
cancer and the fourth highest for cancer-associated deaths,
accounting for 700,000 deaths worldwide in 2012 (1). Incidence rates of CRC are increasingly
reported in industrialized countries with a moderate and high Human
Development Index (2). There is no
effective early detection method for CRC and most of the cases are
diagnosed at advanced stages of the disease. Conventional
radiotherapy and chemotherapy regimens for CRC have a low cure rate
and cause adverse side effects. CRC incidence could be potentially
preventable by the consumption of a healthy diet (3). Therefore, diet modification and
education on healthy lifestyle are necessary to reduce the incident
rate of CRC. CRC risk is inversely associated with the total intake
of fruit and vegetables in the diet, which are excellent sources of
phytochemicals (4). Several in
vitro and in vivo studies have demonstrated that
phytochemicals, such as sulforaphane, diferuloylmethane and
resveratrol, serve antioxidant, anti-inflammatory and anticancer
roles by modulating specific signaling pathways including
Bax/Bcl-2-dependent apoptosis and p53- and p21-mediated cell cycle
arrest, to inhibit the progression and promotion of CRC (3).
Moringa oleifera Lam. (MO) belongs to the
Moringaceae family. Various parts of this vegetable, including
leaves and pods with seeds, are commonly consumed as part of the
regular diet in different parts of the world. MO has a wide range
of health benefits, including anti-inflammatory, anticancer,
hepatoprotective and neuroprotective functions (5). It has been reported that MO has the
medicinal potential including anti-bacterial, anti-inflammatory,
anti-oxidant and antitumor roles (6). Due to these reported functions, the
understanding of the pharmacological functions and underlying
mechanisms of MO action need to be further explored. Bioactive
compounds have been isolated from MO leaves, pods and seeds, such
as vitamins, carotenoids, polyphenol, phenolic acids, flavonoids,
alkaloids, niazirin, niazimicin, niazin A, benzyl-isothiocyanate,
benzyl-thiocarbamate and glucomoringin (7–9).
Extracts from MO leaves and bark have efficacy as anticancer agents
as demonstrated by them inducing apoptosis in a HCT-8 colorectal
cancer cell line (10). The
essential oil from MO seeds and MO dichloromethane extract had
significant cytotoxic, anti-oxidant and anti-proliferative effects
in the Caco-2 colorectal cell line (11,12).
Recent studies have also indicated that MO lam extracts have
cytotoxic and apoptotic effects on human lymphoid and monocytoid
cell lines potentially due to the cross-kingdom regulation by miRNA
present in MO (13,14).
Although various parts of MO exhibit biological
functions, MO pods have drawn considerable attention (15–17) as
they are common ingredients in traditional Asian soups from
Thailand, India and Philippines. Boiled MO (bMO) pods exhibited
strong anti-inflammatory effects in LPS-induced murine macrophages
(15). The chemopreventive and
therapeutic potential of bMO pods were demonstrated in azoxymethane
(AOM)/dextran sodium sulfate (DSS)-induced mouse colon cancer. The
results indicated that tumor formation and multiple tumors were
significantly decreased in the AOM/DSS-induced mice receiving bMO
compared with AOM/DSS-induced mice (16,17). The
therapeutic efficacy of boiled MO was partially due to its ability
to induce apoptotic cell death in colon tissues, by upregulating
the pro-apoptotic protein Bax and decreasing the anti-apoptotic
protein Bcl-2 expression (17).
Preventive treatment with bMO in mice reduced the expression of
inflammatory mediators, inducible nitric oxide synthase (iNOS) and
cyclooxygenase 2 (COX-2) (16).
However, the underlying protective and therapeutic mechanisms of
bMO in exerting anticancer effects remains unclear and requires
further elucidation.
The present study aimed to investigate the molecular
mechanisms for the suppressive effect of bMO supplementation on
azoxymethane (AOM)/dextran sodium sulfate (DSS)-induced mouse colon
carcinogenesis. The proteomic analysis of colon tissues from
AOM/DSS-induced mice receiving preventive and therapeutic bMO diets
was performed, in order to explore the protein expression profiles
in response to bMO consumption. A group of candidate proteins were
identified by Venn diagram analysis and their protein-protein
interaction network and association with apoptotic and inflammatory
signaling pathways were predicted based on the STITCH database.
These data provide an insight into the molecular mechanisms that
underpin the suppressive effects of bMO in a CRC mouse model.
Materials and methods
Chemicals
AOM was purchased from Merck KGaA and dextran sodium
sulfate (molecular weight, 36,000-50,000) was purchased from MP
Biomedicals, LLC. For animal diet preparation, AIN-76 mineral and
vitamin mixtures were purchased from MP Biomedicals, LLC and CLEA
Japan, Inc. Vitamin K1 was kindly provided by DSM Nutritional
Products Ltd. and cellulose (SOLKA-FLOC® 200 FCC) from
FS&D Corp. Sodium caseinate was the product of Erie Foods
International, Inc.
Animal diet preparation
MO boiled pods were prepared as previously described
(16). In brief, MO pods were boiled
in boiling water at ratio 1:1 for 7 min and left them cool down at
25°C. The boiled MO and water were blended, grinded in an
electronic grinder and freeze-dried by lyophilization at −30°C and
termed as bMO. The bMO was packed in vacuum bags and stored at
−20°C until use. The experimental diets were prepared by mixing bMO
at concentrations of 1.5, 3.0 and 6.0% with the basal AIN-76 diet
(Table SI). Animal diet preparation
was prepared according to the protocol previously described
(16,17). The bMO content in animal diets was
equivalent to 10, 20 and 40 times of human consumption (Food
consumption survey data of Thai population, 2006) (18).
Animals and experimental design
A total of 112 male ICR mice [age, 3 weeks old and
weight, 15±3 g (mean ± SD)] were maintained at the Laboratory
Animal Facility of the National Cancer Institute according to the
Institute Care Guidelines as previously described (16,17). The
study protocol was approved by the Animal Ethics Committees of the
National Cancer Institute and Mahidol University (Thailand). Mice
were acclimatized and fed with the modified AIN-76 semi-purified
diet (basal diet), described in Table
SI, and had access to water ad libitum for 5 days. After
acclimatization, mice were randomly divided into 14 groups
(n=8/group) and fed with basal AIN-76 or AIN-76 diet containing
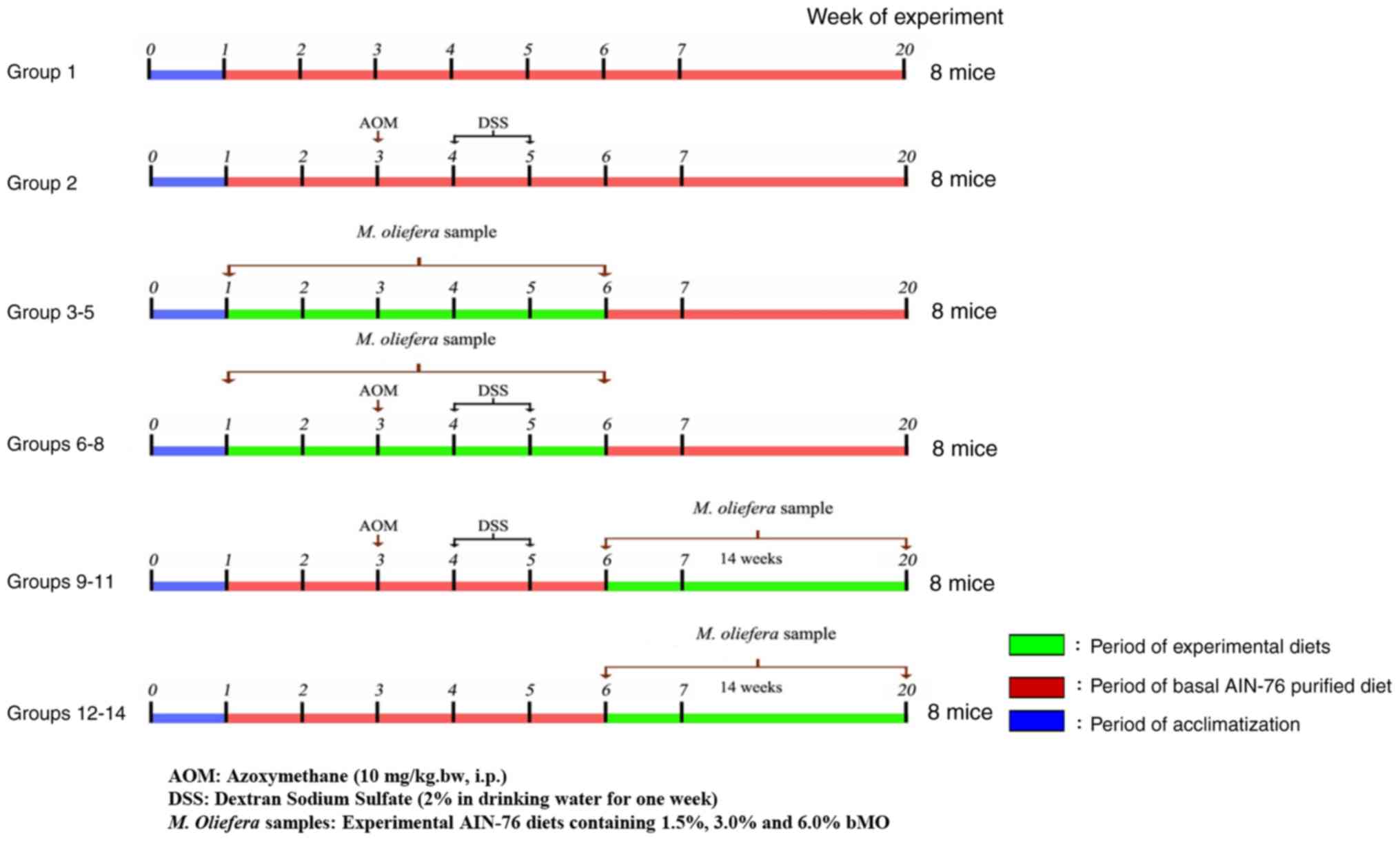
bMO. The experimental design is presented in Fig. 1. In brief, group 1, negative control
was fed the basal diet throughout the 20-week experimental period.
Group 2, positive control for colitis-associated colon cancer
(CACC), was intraperitoneally injected with AOM [10 mg/kg body
weight (BW)] at the 3rd week, and this was followed by
administration of 2% DSS in drinking water at the 4th
week for one week. The mice in Group 2 were fed with basal diets
for the 20-week experimental period. Group 3–5 mice were fed with
basal diet supplemented with bMO at 1.5, 3.0 and 6.0%, respectively
for 5 weeks (from week 1 to 5) and then continued with the basal
diet until the end of experiment. Group 6–8 mice, designated to
evaluate the preventive effect of bMO in CACC mice, were fed with
basal diet containing bMO at 1.5, 3.0 and 6.0%, respectively for 2
weeks prior to giving AOM at week 3 of experimentation; this was
followed by administration with 2% DSS in drinking water in the
4th week for 1 week. Group 6–8 mice were fed with
experimental diets for 5 weeks, followed by basal diet until the
end of the experiment. Group 9–11 mice, designated to assess the
therapeutic effect of bMO in CACC mice, were intraperitoneally
injected with AOM (10 mg/kg BW) at week 3, followed by the
administration of 2% DSS in drinking water at week 4 for a week.
Mice were then fed with the basal diets containing 1.5, 3.0 and
6.0% bMO, respectively at week 6 until the end of the experiment.
Group 12–14 mice were fed with basal diet until week 6, followed by
experimental diets containing 1.5, 3.0 and 6.0% bMO, respectively,
until the end of the experiment. At the end of the study, mice were
euthanized by replacing 30% of the chamber volume per minute with
carbon dioxide. Colons were excised and examined for the presence
of tumors. As described previously, colons from group 1, 2 and 3–8
mice were examined for the preventive effect of bMO (16), and those from group 1, 2 and 9–14
mice were examined for the therapeutic effect (17) on the AOM/DSS-induced carcinogenesis.
The data on tumor nodule incidence in 14 groups of mice described
in the previous reports are summarized in Table I. The portion of colons was frozen
until use for the present proteomic analysis.
 | Table I.Tumor nodule incidence in colons of
experimental mice (8 mice/group). |
Table I.
Tumor nodule incidence in colons of
experimental mice (8 mice/group).
| Group | Treatment | No. of mice
developing tumor nodules | Incidence of mice
with tumor nodule (%) |
|---|
| 1 | Basal diet
(negative control) | 0 | 0 |
| 2 | Basal diet with
AOM/DSS (positive control) | 7 | 87.5 |
| 3 | 1.5% bMO for 5
weeks | 0 | 0 |
| 4 | 3.0% bMO for 5
weeks | 0 | 0 |
| 5 | 6.0% bMO for 5
weeks | 0 | 0 |
| 6 | 1.5% bMO 2 weeks
prior to, during, 1 week after AOM/DSS | 5 | 62.5 |
| 7 | 3.0% bMO 2 weeks
prior to, during, 1 week after AOM/DSS | 4 | 50.0 |
| 8 | 6.0% bMO 2 weeks
prior to, during, 1 week after AOM/DSS | 3 | 37.5a |
| 9 | AOM/DSS before
feeding with 1.5% bMO for 14 weeks | 2 | 25.0a |
| 10 | AOM/DSS before
feeding with 3.0% bMO for 14 weeks | 4 | 50.0 |
| 11 | AOM/DSS before
feeding with 6.0% bMO for 14 weeks | 6 | 75.0 |
| 12 | 1.5% bMO for 14
weeks | 0 | 0 |
| 13 | 3.0% bMO for 14
weeks | 0 | 0 |
| 14 | 6.0% bMO for 14
weeks | 0 | 0 |
Tissue protein extraction
Frozen colon tissue was washed with phosphate buffer
saline (PBS; pH 7.4) and centrifuged (3,000 × g, 5 min, 4°C).
Ice-cold RIPA buffer [50 mM Tris-HCl pH 7.4, 150 mM NaCl, 1 mM
EDTA, 0.25% sodium deoxycholate, 1% (v/v) NP-40, 1 mM DTT and 1X
protease inhibitor cocktail (Roche Diagnostics) was added to the
tissue and was placed on ice for 30 min. After centrifugation at
10,000 × g for 10 min, the supernatant was transferred into a new
microtube and the protein concentration was determined by Lowry
method at 690 nm. Total protein lysate was freshly used or kept at
−80°C until further use.
In-gel digestion
Gel plugs were placed in 96 well-plates and washed
with sterile distilled water. The plate was shaken at room
temperature for 5 min. The gel plugs were destained at room
temperature with 25 mM ammonium bicarbonate
(NH4HCO3) in 50% methanol until they became
transparent. Destaining solution was removed and sterile distilled
water was added for washing three times. Then the 100% acetonitrite
(ACN), 10 mM DTT in 10 mM NH4HCO3, was
applied to the samples and incubated for 1 h at room temperature
for dehydration and reduction. Alkylation was performed by adding
100 mM iodoacetamide in 10 mM NH4HCO3 and
incubating at room temperature for 1 h in the dark. Before
digestion, the gel slices were washed twice with 100% ACN for 5
min. Subsequently, 10 ng/ml trypsin in 50% ACN/10 mM
NH4HCO3 was added to the gels. The gels were
incubated at room temperature for 20 min and kept immersed
throughout the digestion with 30% ACN. Next, the gels were
incubated at 37°C overnight. To extract the digested peptides, 50%
ACN in 0.1% formic acid (FA) was added to the gels and incubated
for 10 min whilst shaking. The peptide products were vacuum-dried
and stored at −80°C until used for mass spectrometric analysis.
Liquid chromatography-mass
spectrometry (LC-MS/MS)
Tryptic peptides were analyzed using the HCT ultra
PTM Discovery System (Bruker Corporation) and UltiMate 3000 LC
System (Thermo Fisher Scientific, Inc.). The peptide samples were
dissolved in 0.1% FA and the separation was performed on a
nanocolumn (Onyx monolithic HDC18; 0.2×150 mm) with electrospray at
a flow rate of 300 nl/min. 0.1% FA and 80% ACN with 0.1% FA were
used as mobile phase A and mobile phase B, respectively. A
multistep gradient was used to elute peptides. The first step
involved the increase of B from 10 to 70% for 13 min, to 90% B at
13–15 min, followed by decreasing to 10% for 15–20 min. Mass
spectra were recorded in the positive-ion mode. A scan range of
300-1,500 m/z in AutoMS mode was used for determining the peptide
fragment mass spectra. The three averages and five precursor ions
were selected from the MS scan 50-3,000 m/z
Protein quantitation and
identification
The DeCyder MS 2.0 Differential analysis software
(DeCyderMS, GE Healthcare) was used for protein quantitation. The
peptide was detected by the PepDetect module and the peptide ion
signals were quantified. The Mascot software version 2.2 (Matrix
Science, Ltd.) was used for analyzing MS/MS data and protein
identification was obtained by searching against NCBI database
(https://www.ncbi.nlm.nih.gov). The
biological function of each protein was obtained from Uniprot
(https://www.uniprot.org) and STITCH 5.0
(http://stitch.embl.de/) databases. A Venn diagram
was generated by the jvenn web application (https://xcmsonline.scripps.edu/lib/jvenn-1.8).
Protein interaction networks were determined using the STITCH 5.0
database.
Statistical analysis
All data except tumor incidence were expressed as
mean ± SD (n=8). Differences between tumor incidence in
experimental and positive control groups were evaluated by the
χ2 or Fisher's exact probability test. P<0.05 was
considered to indicate a statistically significant difference. All
statistical analyses were analyzed by SPSS19.0 software (IBM).
Results
Effects of bMO on AOM/DSS-induced
mouse colon carcinogenesis
The effects of bMO on AOM/DSS-induced CACC were
studied in an experimental mouse model, as presented in Fig. 1. Incidence of tumor nodules were
examined in 14 experimental groups of mice. No tumor was observed
in the negative control (group 1), while the positive control mice
(group 2) fed with a basal diet with AOM/DSS demonstrated the
highest incidence of tumor nodules (87.5%) classified as tubular
adenoma (Table I). Group 6–8 mice
fed with basal diets containing 1.5, 3.0 and 6.0% bMO respectively
prior to induction with AOM/DSS had lower incidences of tumor
nodules compared with the positive control mice (Table I). Notably, mice fed with 6.0% bMO
diet (group 8) had a significantly lower tumor nodule incidence
compared with the positive control mice (37.5%, P<0.05), while
tumor nodules were not observed in group 3–5 mice fed with 1.5, 3.0
and 6.0% bMO diets without AOM/DSS induction, respectively
(Table I). In addition, group 9–11
mice treated with AOM/DSS and subsequently fed with 1.5, 3 and 6.0%
bMO diets, respectively from week 6 until the end of the experiment
also demonstrated lower incidences of tumor nodules compared with
positive control mice (group 2). In particular, mice fed with a
1.5%-bMO diet (group 9) had a significantly lower tumor nodule
incidence (25.0%) compared with the positive control mice
(P<0.05; Table I). No tumor
nodule was observed in group 12–14 mice fed with the same feeding
regimen without AOM/DSS induction (Table
I). Based on the significant reduction of tumor incidence
observed in mice receiving 6.0% bMO prior to induction with
AOM/DSS, designated as the preventive consumption, and 1.5% bMO
diets after inducing with AOM/DSS, assigned as therapeutic
consumption, colon samples from 6 experimental groups (group 1, 2,
5, 8, 9 and 12) were chosen for further comparative analysis of
protein expression profiles expressed in response to bMO
consumption in the mouse model of CACC.
Protein expression profiles associated
with the effects of bMO on AOM/DSS-induced mouse colon
carcinogenesis
To explore the protein expression profiles that
underlie the effects of bMO consumption on AOM/DSS-induced mouse
colon carcinogenesis, total proteins were extracted from the colons
of mice and subjected to analysis using GeLC-MS/MS. The analysis
demonstrated a total of 125 proteins that were differentially
expressed in tissue samples from all 14 groups of mice. Functional
annotations of the proteins, obtained from online Uniprot and
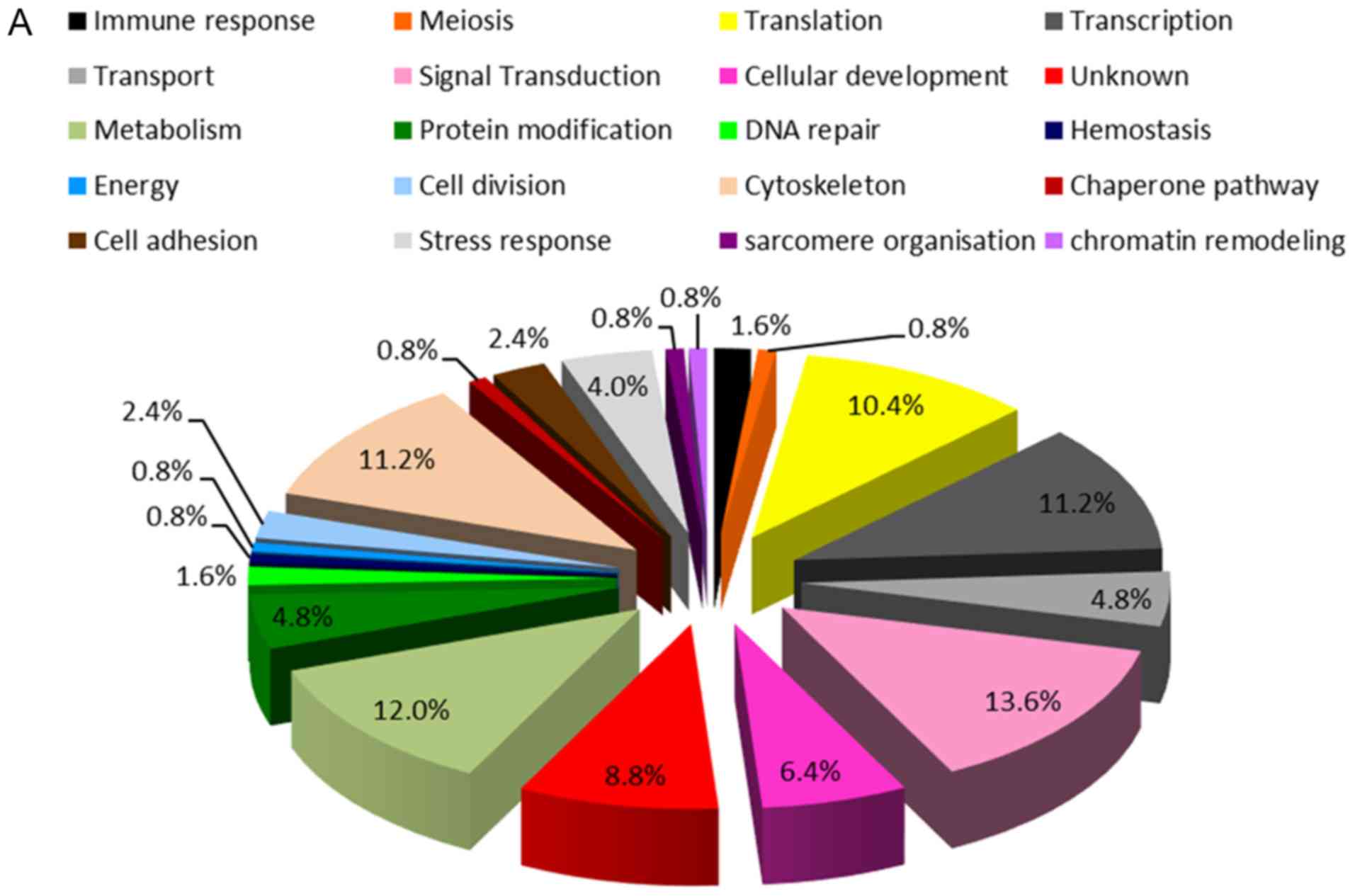
STITCH 5.0 databases, are presented in Fig. 2A. These proteins were associated with
various biological processes, including signal transduction
(13.6%), metabolism (12.0%), cytoskeleton (11.2%), transcription
(11.2%), translation (10.4%), cellular development (6.4%),
transport (4.8%), protein modification (4.8%), stress response
(4.0%), cell division (2.4%), cell adhesion (2.4%), immune response
(1.6%), DNA repair (1.6%), meiosis (0.8%), hemostasis (0.8%),
energy (0.8%), chaperone pathway (0.8%), sarcomere organization
(0.8%), chromatin remodeling (0.8%) and unknown (8.8%).
A Venn diagram was used to demonstrate the numbers
of proteins differentially expressed in 6 selected experimental
groups of mice designed to assess preventive and therapeutic
effects of bMO, as presented in Fig.
2B. There were 99 proteins identified in the negative control
group, 119 proteins in AOM/DSS-treated mice, 112 proteins in mice
supplemented with 1.5% bMO, 115 proteins in the AOM/DSS-induced
mice receiving therapeutic 1.5% bMO diets, 117 proteins in mice
supplemented with 6.0% bMO and 113 proteins in the AOM/DSS-induced
mice receiving preventive 6.0% bMO diets. There were seven proteins
potentially associated with the preventive and therapeutic effects
of bMO on AOM/DSS-induced CACC, as indicated by red arrows in
Fig. 2B. These proteins were
classified into three groups, based on their expression patterns
described in Table II. A total of
three proteins in group 1, 60S acidic ribosomal protein P1, fragile
X mental retardation 1 (FMR1) and cystatin 9, were detectable in
control mice and downregulated in AOM/DSS-induced mice. Notably,
all three proteins were upregulated in response to both preventive
and therapeutic supplementation with bMO. In contrast, two proteins
(group 2), STDP2 and zinc finger protein 638, were undetectable in
the control group and were expressed in response to the AOM/DSS
induction of CACC. STDP2 appeared to be undetectable in all mice
treated with preventive and therapeutic bMO, while zinc finger
protein 638 proteins were not found in preventive bMO-treated and
control therapeutic 1.5% bMO groups and remained detectable in
therapeutic 1.5% bMO-supplemented mice. Additionally,
magnesium-dependent protein phosphatase 2C and unnamed protein
product (group 3) were undetectable in control and AOM/DSS-induced
mice. These two proteins were upregulated in response to both
preventive and therapeutic bMO supplementation, however unnamed
protein product was undetectable in the AOM/DSS induced mice fed
with therapeutic 1.5% bMO. These data indicated a potential
association of these seven proteins in the preventive and
therapeutic effects of bMO on AOM/DSS-induced mouse CACC.
| Table II.Expression patterns of seven
candidate proteins in response to the preventive and therapeutic
consumption of bMO in AOM/DSS-induced mouse colon
carcinogenesis. |
Table II.
Expression patterns of seven
candidate proteins in response to the preventive and therapeutic
consumption of bMO in AOM/DSS-induced mouse colon
carcinogenesis.
|
|
|
|
|
|
|
|
| Preventive
treatment | Therapeutic
treatment |
|---|
|
|
|
|
|
|
|
|
|
|
|
|---|
| Protein group | Protein name | Accession
number | Organisms | Peptides | Function | Control | AOM/DSS | 6.0% b MO | 6.0% bMO + AOM | 1.5% bMO | AOM + 1.5% bMO |
|---|
| 1 | 60S acidic
ribosomal protein P1 | gi|109462386 | Rattus
norvegicus | MSSAAYV TVLWR | Translation | Yes | No | Yes | Yes | Yes | Yes |
|
| FMR1-tm1Cgr
(FMR1) | gi|46242307 | Mus
musculus | TVGVAPSRVK | Transcription | Yes | No | Yes | Yes | Yes | Yes |
|
| Cystatin 9 | gi|6753546 | Mus
musculus | MEHIMSSWR | Immune
response | Yes | No | Yes | Yes | Yes | Yes |
| 2 | Round spermatids
protein STDP2 | gi|51599286 | Mus
musculus | IPEGSLGKDL
EAVSETR | Meiosis | No | Yes | No | No | No | No |
|
| Zinc finger protein
638 | gi|6679098 | Mus
musculus | GIPHRFPGHGS
YQNMGPQR | Transcription | No | Yes | No | No | No | Yes |
| 3 | Protein phosphatase
2C, magnesium dependent, catalytic subunit | gi|12585293 | Mus
musculus | MSSVFEDQNA
ATHLIR | Protein
modification | No | No | Yes | Yes | Yes | Yes |
|
| Unnamed protein
product | gi|26380444 | Mus musculus | SSRSGPILGPP
GGSGHR | Unknown | No | No | Yes | Yes | Yes | No |
STITCH analysis for protein-protein
interaction networks of candidate proteins
In order to identify the molecular mechanisms
underlying the preventive and therapeutic effects of bMO on
AOM/DSS-induced carcinogenesis, protein-protein interaction
networks of the seven proteins obtained from Venn diagram analysis
were analyzed. Using the online STITCH 5.0 resource, the networks
were predicted with parameters limited to Mus musculus,
medium confidence score (0.400) and no more than 10 interactors.
All the candidate proteins except for FMR1, round spermatids
protein STDP2 and unnamed protein product were found in the
analysis. The analysis included ribosomal protein large P1 (Rplp1),
cystatin 9 (Cst9), zinc finger protein 638 or zinc finger
matrin-like protein (ZfmL), and magnesium dependent protein
phosphatase 1G (formerly 2C) (Ppm1g). As our previous studies
indicated that bMO has anticancer properties, as it induces
apoptosis and is anti-inflammatory (15,17), the
associations of these candidate proteins with apoptotic and
inflammatory signaling pathways were determined. The
protein-protein interaction with key regulators of apoptosis,
BCL2-associated X protein (Bax), B cell leukemia/lymphoma 2 (BCL-2)
and caspases (Casp) were analyzed (Fig.
3A). The data demonstrated that only Rplp1 protein was
associated with this apoptotic signaling network, whereas the other
proteins, Cst9, Zfml and Ppm1g, did not show any associations
(Fig. 3A). Analysis of the
interaction between the aforementioned proteins and important
inflammatory molecules, tumor necrosis factor-α (TNF-α), Fas ligand
(Fasl), inducible nitric oxide synthase (NOS2), endothelial nitric
oxide synthase (NOS3) and interleukin-6 (IL-6), signal transducer
and activator of transcription 3 (STAT3) is presented in Fig. 3B. The data indicated that Rplp1 and
Ppm1g were involved in inflammatory pathways, while Cst9 and Zfml
proteins did not demonstrate any interaction. The STITCH analysis
indicated that of a total of seven candidate proteins potentially
associated with the preventive and therapeutic effects of bMO in
AOM/DSS-induced carcinogenesis, Rplp1 and Ppm1g proteins had
potential interactions with apoptotic and inflammatory signaling
pathways.
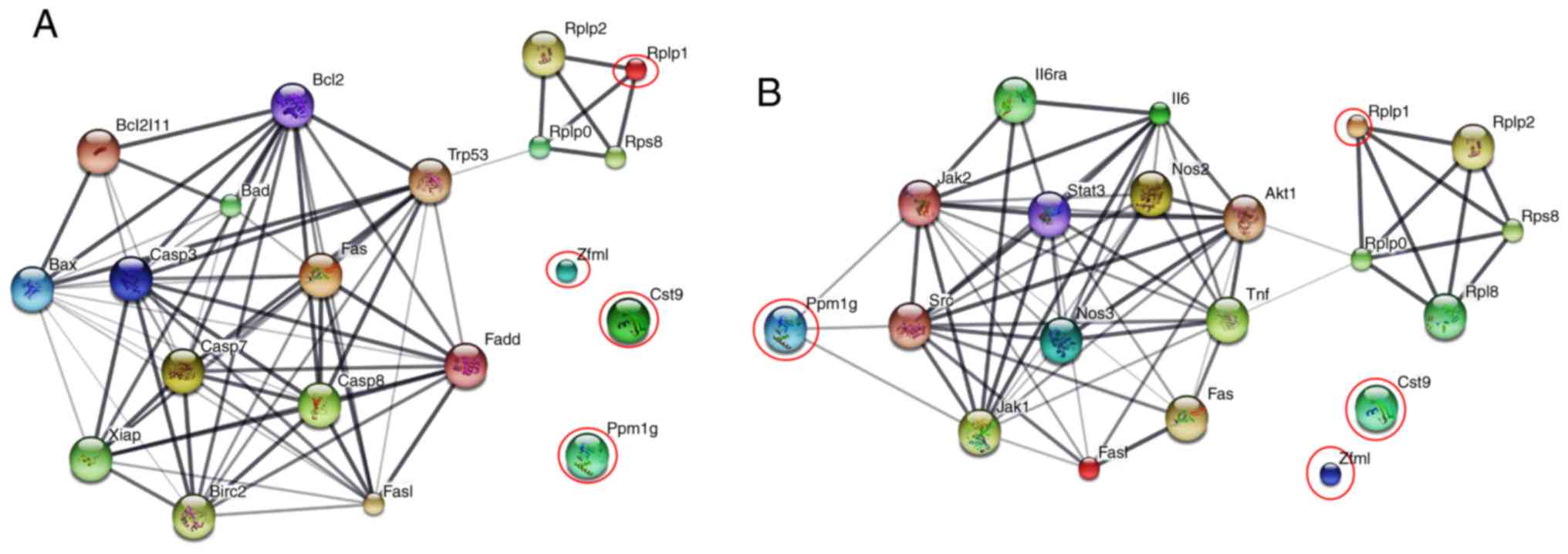 | Figure 3.Protein interaction networks of
candidate proteins and their association with apoptotic and
inflammatory signaling pathways. (A) and (B) present the
associations of candidate proteins with apoptotic and inflammatory
signaling networks, respectively. Red circles indicate the input
proteins identified in this study. Edges represent different levels
of meaningful protein-protein associations. Stronger associations
are presented by thicker lines and weaker associations by thinner
lines. Bax, Bcl-2-associated X protein; Bcl2 (Bcl-2), B-cell
lymphoma 2; casp3, caspase 3; casp7, caspase 7; TNF, tumor necrosis
factor; NOS2, inducible nitric oxide synthase; NOS3, endothelial
nitric oxide synthase; IL-6, interleukin-6; STAT 3, signal
transducer and activator of transcription 3. |
Discussion
Previous studies in an experimental mouse model have
demonstrated the preventive and therapeutic effects of bMO on the
AOM/DSS-induced CACC (16,17). In the present study investigating the
effects of 1.5, 3.0 and 6.0% bMO consumption on colon tumor
incidence, a significant preventive effect of bMO was observed only
in mice receiving 6.0% bMO diets, while mice fed with therapeutic
diets containing 1.5% bMO demonstrated a significant decrease in
the number of tumor nodules. Basically, vitamins, minerals and
dietary phytochemicals exhibited both antioxidant and prooxidant
activities. The lower efficacy of high-dose bMO in the present
study was possibly due to the activity of bioactive compounds in MO
pods including glucosinolates, isothiocyanates and β-carotene
(19). These results are consistent
with those of a previous study in AOM-induced rats. Rats that
received 100 or 200 ppm of β-carotene inhibited the aberrant
cryptic foci (ACF) formation, whereas administration of higher
doses, 1,000 or 2,000 ppm of β-carotene increased colonic ACF
formation (20).
In the present study, the comparative analysis of
protein expression using the GeLC-MS/MS technique in mouse colon
tissues from all 14 groups of mice revealed that 125 proteins were
differentially expressed. These proteins were identified to be
involved in various biological activities, including signal
transduction, metabolism, cytoskeleton, transcription, translation,
cellular development, transport, protein modification, stress
response, cell division, cell adhesion, immune response, DNA
repair, meiosis, hemostasis, energy, the chaperone pathway,
sarcomere organization and chromatin remodeling. Notably, there
were seven candidate proteins found that were potentially expressed
in response to bMO consumption in AOM/DSS-induced CACC and serve
key roles in the suppression associated with bMO. Using the online
STITCH 5.0 database, all the candidate proteins were found except
for FMR1, STDP2 and unnamed protein product and the association of
Rplp1 and Ppm1g proteins with inflammatory and apoptotic signaling
networks were demonstrated. The data obtained in the present study
is consistent with previous reports demonstrating that preventive
and therapeutic efficacies of bMO were partially based on the
induction of apoptosis in colon tissues and reduction of
proinflammatory cytokines and mediators in the mouse CACC model
(15–17).
In the present study, ribosomal protein P1
expression was downregulated in mouse AOM/DSS-induced CACC, and was
detectable in mice receiving both preventive and therapeutic bMO
supplementation, indicating their potential roles in mouse CACC,
and the preventive and therapeutic potentials of bMO in colon
cancer. Rplp1 is a member of the acidic ribosomal proteins (RPs)
found in the ribosomal stalk (21,22). The
stalk is known as a pentameric P protein complex comprising RP
large P0 (Rplp0) and two heterodimers formed by RP large P1 (Rplp1)
and RP large P2 (Rplp2), which play important roles in the
translation process (21,22). Alterations of Rplp protein expression
have been reported in human cancer, such as colorectal and
pancreatic cancer (23,24). In contrast to the findings of the
present study that demonstrated the downregulation of Rplp1
expression in mice with CACC compared with normal mouse control, a
previous study indicated a significant increase of Rplp0, Rplp1 and
Rplp2 protein expression in breast, skin, colon, lung and ovarian
tumors with respect to corresponding normal tissues and the
presence of metastasis in gynecological cancer (25). Furthermore, co-expression of Rplp1
with mutant rasVal12 caused the transformation of murine NIH3T3
cells (26). However, Rplp1 was
demonstrated to be essential for cell proliferation, mouse
embryonic and brain development (27). Downregulation of Rplp0, Rplp1 and
Rplp2 proteins led to reactive oxygen species (ROS) accumulation,
endoplasmic reticulum (ER) stress/unfolded protein response (UPR)
activation, and induced autophagy, and inhibition of ROS production
resulted in restoring of proliferative capacity in breast and
ovarian cancer cells (28). The
STITCH analysis performed in the present study demonstrated
relationships between Rplp proteins and apoptotic and inflammatory
signaling networks. This data is consistent with previous studies
demonstrating that there is a link between the expression of Rplp1
and the tumor suppressor gene p53 in vitro and in vivo
(26–28), and that a deficiency of Rplp resulted
in the reduction of mTOR and Akt phosphorylation in MCF-7 breast
cancer cells (28). To the best of
our knowledge at present there is no direct evidence indicating a
functional role of Rplp1 in apoptosis and inflammation involving
colorectal carcinogenesis. While precise biological and
physiological functions of Rplp remain largely unclear, the
downregulation of Rplp1 expression observed in the present study
suggested that Rplp1 has a protective role in AOM/DSS-induced mouse
CACC and Rplp1 is probably one of the preventive and therapeutic
targets of bMO supplementation.
Similarly, FMR1 and Cst9 proteins were downregulated
in mouse AOM/DSS-induced CACC and appeared to be detectable in mice
receiving both preventive and therapeutic bMO supplementation. FMR1
proteins are encoded by the FMR1 gene whose inactivation causes
fragile X syndrome (FXS) characterized phenotypically and
molecularly in a knockout mouse model (29). The fragile X mental retardation
protein (FMRP), an RNA-binding protein that serves roles in RNA
metabolism (30), has been
demonstrated directly or indirectly to influence cancer
progression. Previous studies indicated overexpression of FMR1 mRNA
in hepatocellular carcinoma cells (31,32) and
patients with FXS had a decreased risk of cancer (33). The FMRP gene also modulated several
malignant phenotypes required for the metastasis and invasion of
melanoma and breast cancer (34,35).
Cystatin 9 (Cst9) proteins, also known as testatin,
are characterized as a member of the cysteine protease inhibitors
named Cystatin family, and are expressed in a restricted manner in
germ cells and somatic cells in reproductive tissues (36). A few studies have indicated that Cst9
proteins are involved in testis development and have
immunomodulatory and antimicrobial functions (36–38).
Although the functional properties of Cst9 remain unclear,
accumulating evidence implicates cystatins and their target
lysosomal proteases, termed the cathepsins, in tumorigenesis and
metastasis of various cancer types, such as brain tumors (39,40). The
cystatins appear to exert their tumor suppressor activity by
counter-balancing the overexpressed tumor-associated proteolytic
activity of the cathepsins (39,40). In
the present study, Cst9, but not FMR1, was found in the STITCH 5.0
database and was not found to be associated with apoptosis and
inflammatory signaling pathways. At present, there is a lack of
direct evidence supporting the roles of FMR1 and Cst9 proteins in
colorectal carcinogenesis, and the results of the proteomic
analysis performed in the present study may suggest the potential
implication of FMR1 and Cst9 in mouse AOM/DSS-induced
carcinogenesis and response to preventive and therapeutic bMO
treatments.
The proteomic analysis performed in the present
study found that STDP2 and ZNF638 were expressed in response to
AOM/DSS treatment in AOM/DSS-induced mouse CACC. The STDP2 protein
is encoded by identified novel genes with evident testis-specific
expression, characterized by in silico and in vitro
approaches in spermatogenetic cells (41). Presently, there is a lack of evidence
demonstrating the precise function of STDP2, consistent with an
absence of STDP2 in STITCH resources. ZNF638 alternatively named as
zinc finger matrin-like (Zfml), is one of the zinc-finger domain
containing proteins participating in a wide range of molecular and
cellular activities (42). While
there are several other zinc-finger proteins such as ZNF281, ZNF750
and ZNF185, having oncogenic and tumor suppressor functions, the
ZNF638 protein has been reported as an early regulator of
adipogenesis (42). The protein
appears to act as a transcriptional cofactor of
CCAAT/enhancer-binding protein, leading to the expression of
peroxisome proliferator-activated receptor γ, the key regulator of
adipocyte differentiation (43). The
STITCH analysis performed in the present study indicated that there
was no association between ZNF638 protein and apoptosis or
inflammation. In the present study, while both preventive and
therapeutic bMO consumption were able to downregulate STDP2
expression, ZNF638 was downregulated in response to only the
preventive supplementation. To the best of our knowledge, the
present proteomic study is the first report suggesting the
potential involvement of STDP2 and ZNF638 in mouse CACC and the
suppressive potentials of bMO in colorectal cancer.
In the experimental mouse model used in the present
study, Ppm1g and unnamed protein product (gi|26380444) were not
involved in the AOM/DSS-induced mouse CACC. They were likely to be
upregulated in response to both preventive and therapeutic bMO
consumption, although expression of unnamed protein product was not
observed in the AOM/DSS-induced mice fed with therapeutic bMO diet.
Whereas there is no information of the unnamed protein product,
Ppm1g is well-characterized. Ppm1g is a member of Type 2C
phosphatase family (PP2C or PPM) of Ser/Thr protein phosphatases
that possess unique patterns of tissue and subcellular distribution
associated with diverse functionalities (44). PP2C isoforms have been implicated in
signaling networks controlling cell differentiation, proliferation,
growth, survival and metabolism (44). Previous studies indicated that the
dephosphorylation of a cyclin-CDK inhibitor p27 by Ppm1g maintained
the proper level of the p27 proteins that basically induce cell
cycle arrest at G1 phase to prevent the improper entry to cell
cycle (45). Ppm1g has also been
demonstrated to participate in the DNA damage signal leading to
tumor suppressor p53-dependent response (46). In the present study, STITCH analysis
suggested the association of Ppm1g protein with inflammatory, but
not with apoptotic signaling networks. These results are contrary
to previous studies that demonstrated the anti-apoptotic activity
of Ppm1g in mouse neural development (47) and found that Ppm1g is a
transcriptional coactivator of NF-κB in the tumor necrosis
factor-α-mediated inflammatory pathways (48). Therefore, the upregulation of Ppm1g
in response to the preventive and therapeutic supplementation with
bMO observed in the present study may explain the potential of bMO
in this mouse colon carcinogenesis model.
The data generated in the present study provide
insight into the molecular mechanisms of the suppressive potential
of MO consumption in mouse colon carcinogenesis, based on the
expression of the seven candidate proteins in response to MO in
colon tissues. Nevertheless, further in vitro and in
vivo studies are required to investigate expression and
function of the candidate proteins in colon cancers and the lack of
experimental validation of protein expression may be noted as a
limitation of the present study. The proteomic findings of this
study support the potential of MO-supplemented diets in cancer
treatment and care.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank Associate Professor
Dr Anudep Rangsripipat from the Faculty of Veterinary Science,
Chulalongkorn University, Thailand for his assistance with the
pathological examination of mouse tissue. The authors would also
like to thank Professor Jeong-Sang Lee from Jeonju University,
Korea for his helpful guidance on constructing the AOM/DSS-induced
mouse colon carcinogenesis model.
Funding
This work was supported by grants from the National
Center for Genetic Engineering and Biotechnology (grant no.
P-09-00762), National Science and Technology Development Agency and
Ministry of Science and Technology, Thailand.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
PP performed data analysis and drafted the
manuscript. PK, SB and CB performed the animal experiments and data
analysis. SR and NP contributed to proteomic analysis. ST designed
the experiments and edited the manuscript for important
intellectual content. CA conducted proteomic analysis, interpreted
data and wrote the manuscript. All authors have read and approved
the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Animal Ethics
Committee of the National Cancer Institute (approval no. ACUC
2009/003) and Animal Care and Use Protocol of Mahidol University
(Nakhon Pathom, Thailand).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
AOM
|
azoxymethane
|
|
CACC
|
colitis-associated colon cancer
|
|
DSS
|
dextran sodium sulfate
|
|
MO
|
Moringa oleifera
|
|
GeLC-MS/MS
|
gel electrophoresis and liquid
chromatography-tandem mass spectrometry
|
References
|
1
|
Arnold M, Sierra MS, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global patterns and trends in
colorectal cancer incidence and mortality. Gut. 66:683–691. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dolatkhah R, Somi MH, Kermani IA,
Ghojazadeh M, Jafarabadi MA, Farassati F and Dastgiri S: Increased
colorectal cancer incidence in Iran: A systematic review and
meta-analysis. BMC Public Health. 15:9972015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yin TF, Wang M, Qing Y, Lin YM and Wu D:
Research progress on chemopreventive effects of phytochemicals on
colorectal cancer and their mechanisms. World J Gastroenterol.
22:7058–7068. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bradbury KE, Appleby PN and Key TJ: Fruit,
vegetable, and fiber intake in relation to cancer risk: Findings
from the European prospective investigation into cancer and
nutrition (EPIC). Am J Clin Nutr. 100 (Suppl 1):S394–S398. 2014.
View Article : Google Scholar
|
|
5
|
Abdull Razis AF, Ibrahim MD and Kntayya
SB: Health benefits of moringa oleifera. Asian Pac J Cancer Prev.
15:8571–8576. 2014. View Article : Google Scholar
|
|
6
|
Karim NA, Ibrahim MD, Kntayya SB, Rukayadi
Y, Hamid HA and Razis AFA: Moringa oleifera Lam: Targeting
chemoprevention. Asian Pac J Cancer Prev. 17:3675–3686.
2016.PubMed/NCBI
|
|
7
|
Leone A, Spada A, Battezzati A, Schiraldi
A, Aristil J and Bertoli S: Cultivation, genetic,
ethnopharmacology, phytochemistry and pharmacology of Moringa
oleifera leaves: An overview. Int J Mol Sci. 16:12791–12835. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Francis JA, Jayaprakasam B, Olson LK and
Nair MG: Insulin secretagogues from Moringa oleifera with
cyclooxygenase enzyme and lipid peroxidation inhibitory activities.
Helv Chim Acta. 87:317–326. 2004. View Article : Google Scholar
|
|
9
|
Cheenpracha S, Park EJ, Yoshida WY, Barit
C, Wall M, Pezzuto JM and Chang LC: Potential anti-inflammatory
phenolic glycosides from the medicinal plant Moringa oleifera
fruits. Bioorg Med Chem. 18:6598–6602. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Al-Asmari AK, Albalawi SM, Athar MT, Khan
AQ, Al-Shahrani H and Islam M: Moringa oleifera as an anti-cancer
agent against breast and colorectal cancer cell lines. PLoS One.
10:e01358142015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Elsayed EA, Sharaf-Eldin MA and Wadaan M:
In vitro evaluation of cytotoxic activities of essential oil from
moringa oleifera seeds on HeLa, HepG2, MCF-7, CACO-2 and L929 cell
lines. Asian Pac J Cancer Prev. 16:4671–4675. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Suphachai C: Antioxidant and anticancer
activities of Moringa oleifera leaves. J Med Plants Res. 8:318–325.
2014. View Article : Google Scholar
|
|
13
|
Potestà M, Minutolo A, Gismondi A, Canuti
L, Kenzo M, Roglia V, Macchi F, Grelli S, Canini A, Colizzi V and
Montesano C: Cytotoxic and apoptotic effects of different extracts
of Moringa oleifera Lam on lymphoid and monocytoid cells. Exp Ther
Med. 18:5–17. 2019.PubMed/NCBI
|
|
14
|
Pirrò S, Matic I, Guidi A, Zanella L,
Gismondi A, Cicconi R, Bernardini R, Colizzi V, Canini A, Mattei M
and Galgani A: Identification of microRNAs and relative target
genes in Moringa oleifera leaf and callus. Sci Rep. 9:151452019.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Muangnoi C, Chingsuwanrote P,
Praengamthanachoti P, Svasti S and Tuntipopipat S: Moringa oleifera
pod inhibits inflammatory mediator production by
lipopolysaccharide-stimulated RAW 264.7 murine macrophage cell
lines. Inflammation. 35:445–455. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Budda S, Butryee C, Tuntipopipat S,
Rungsipipat A, Wangnaithum S, Lee JS and Kupradinun P: Suppressive
effects of Moringa oleifera Lam pod against mouse colon
carcinogenesis induced by azoxymethane and dextran sodium sulfate.
Asian Pac J Cancer Prev. 12:3221–3228. 2011.PubMed/NCBI
|
|
17
|
Kraiphet S, Butryee C, Rungsipipat A,
Budda S, Rattanapinyopitak K and Tuntipopipat S: Apoptosis induced
by Moringa oleifera Lam. pod in mouse colon carcinoma model. Comp
Clin Pathol. 27:21–30. 2018. View Article : Google Scholar
|
|
18
|
Kosulwat V, Rojroongwasinkul N,
Boonpraderm A, Viriyapanich T, Jitnarin N, Sornkaew N and
Vanicchakul C: Food consumption data of Thailand. Bangkok: National
Bureau of Agricultural Commodity and Food Standards, Ministry of
Agriculture and Cooperatives; Bangkok, Thailand: 2006, (In
Thai).
|
|
19
|
Fahey JW: Moringa oleifera: A review of
the medical evidence for its nutritional, therapeutic, and
prophylactic properties. Part 1. Trees Life J. 1:52005.
|
|
20
|
Raju J, Swamy MV, Cooma I, Patlolla JM,
Pittman B, Reddy BS, Steele VE and Rao CV: Low doses of
beta-carotene and lutein inhibit AOM-induced rat colonic ACF
formation but high doses augment ACF incidence. Int J Cancer.
113:798–802. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Naganuma T, Shiogama K and Uchiumi T: The
N-terminal regions of eukaryotic acidic phosphoproteins P1 and P2
are crucial for heterodimerization and assembly into the ribosomal
GTPase-associated center. Genes Cells. 12:501–510. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xu X, Xiong X and Sun Y: The role of
ribosomal proteins in the regulation of cell proliferation,
tumorigenesis, and genomic integrity. Sci China Life Sci.
59:656–672. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang L, Zhou W, Velculescu VE, Kern SE,
Hruban RH, Hamilton SR, Vogelstein B and Kinzler KW: Gene
expression profiles in normal and cancer cells. Science.
276:1268–1272. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Loging WT and Reisman D: Elevated
expression of ribosomal protein genes L37, RPP-1, and S2 in the
presence of mutant p53. Cancer Epidemiol Biomarkers Prev.
8:1011–1016. 1999.PubMed/NCBI
|
|
25
|
Artero-Castro A, Castellvi J, García A,
Hernández J, Ramón y Cajal S and Lleonart ME: Expression of the
ribosomal proteins Rplp0, Rplp1, and Rplp2 in gynecologic tumors.
Hum Pathol. 42:194–203. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Artero-Castro A, Kondoh H,
Fernández-Marcos PJ, Serrano M, Ramón y Cajal S and Lleonart ME:
Rplp1 bypasses replicative senescence and contributes to
transformation. Exp Cell Res. 315:1372–1383. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Perucho L, Artero-Castro A, Guerrero S,
Ramón y Cajal S, LLeonart ME and Wang ZQ: RPLP1, a crucial
ribosomal protein for embryonic development of the nervous system.
PLoS One. 9:e999562014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Artero-Castro A, Perez-Alea M, Feliciano
A, Leal JA, Genestar M, Castellvi J, Peg V, Ramón Y Cajal S and
Lleonart ME: Disruption of the ribosomal P complex leads to
stress-induced autophagy. Autophagy. 11:1499–1519. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yan QJ, Asafo-Adjei PK, Arnold HM, Brown
RE and Bauchwitz RP: A phenotypic and molecular characterization of
the fmr1-tm1Cgr fragile X mouse. Genes Brain Behav. 3:337–359.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bagni C, Tassone F, Neri G and Hagerman R:
Fragile X syndrome: Causes, diagnosis, mechanisms, and
therapeutics. J Clin Invest. 122:4314–4322. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li Y, Tang Y, Ye L, Liu B, Liu K, Chen J
and Xue Q: Establishment of a hepatocellular carcinoma cell line
with unique metastatic characteristics through in vivo selection
and screening for metastasis-related genes through cDNA microarray.
J Cancer Res Clin Oncol. 129:43–51. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu Y, Zhu X, Zhu J, Liao S, Tang Q, Liu
K, Guan X, Zhang J and Feng Z: Identification of differential
expression of genes in hepatocellular carcinoma by suppression
subtractive hybridization combined cDNA microarray. Oncol Rep.
18:943–951. 2007.PubMed/NCBI
|
|
33
|
Schultz-Pedersen S, Hasle H, Olsen JH and
Friedrich U: Evidence of decreased risk of cancer in individuals
with fragile X. Am J Med Genet. 103:226–230. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lucá R, Averna M, Zalfa F, Vecchi M,
Bianchi F, La Fata G, Del Nonno F, Nardacci R, Bianchi M, Nuciforo
P, et al: The fragile X protein binds mRNAs involved in cancer
progression and modulates metastasis formation. EMBO Mol Med.
5:1523–1536. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zalfa F, Panasiti V, Carotti S,
Zingariello M, Perrone G, Sancillo L, Pacini L, Luciani F, Roberti
V, D'Amico S, et al: The fragile X mental retardation protein
regulates tumor invasiveness-related pathways in melanoma cells.
Cell Death Dis. 8:e31692017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Töhönen V, Osterlund C and Nordqvist K:
Testatin: A cystatin-related gene expressed during early testis
development. Proc Natl Acad Sci USA. 95:14208–14213. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hasegawa K, Chuma S, Tada T, Sakurai T,
Tamura M, Suemori H and Nakatsuji N: Testatin transgenic and
knockout mice exhibit normal sex-differentiation. Biochem Biophys
Res Commun. 341:369–375. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Eaves-Pyles T, Patel J, Arigi E, Cong Y,
Cao A, Garg N, Dhiman M, Pyles RB, Arulanandam B, Miller AL, et al:
Immunomodulatory and antibacterial effects of cystatin 9 against
Francisella tularensis. Mol Med. 19:263–275. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Magister S and Kos J: Cystatins in immune
system. J Cancer. 4:45–56. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Strojnik T, Zajc I, Bervar A, Zidanik B,
Golouh R, Kos J, Dolenc V and Lah T: Cathepsin B and its inhibitor
stefin A in brain tumors. Pflugers Arch. 439 (3 Suppl):R122S–R123S.
2000. View Article : Google Scholar
|
|
41
|
Hong S, Choi I, Woo JM, Oh J, Kim T, Choi
E, Kim TW, Jung YK, Kim DH, Sun CH, et al: Identification and
integrative analysis of 28 novel genes specifically expressed and
developmentally regulated in murine spermatogenic cells. J Biol
Chem. 280:7685–7693. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Cassandri M, Smirnov A, Novelli F, Pitolli
C, Agostini M, Malewicz M, Melino G and Raschellà G: Zinc-finger
proteins in health and disease. Cell Death Discov. 3:170712017.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Meruvu S, Hugendubler L and Mueller E:
Regulation of adipocyte differentiation by the zinc finger protein
ZNF638. J Biol Chem. 286:26516–26523. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lu G and Wang Y: Functional diversity of
mammalian type 2C protein phosphatase isoforms: New tales from an
old family. Clin Exp Pharmacol Physiol. 35:107–112. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Sun C, Wang G, Wrighton KH, Lin H,
Songyang Z, Feng XH and Lin X: Regulation of p27Kip1
phosphorylation and G1 cell cycle progression by protein
phosphatase PPM1G. Am J Cancer Res. 6:2207–2220. 2016.PubMed/NCBI
|
|
46
|
Khoronenkova SV, Dianova II, Ternette N,
Kessler BM, Parsons JL and Dianov GL: ATM-dependent downregulation
of USP7/HAUSP by PPM1G activates p53 response to DNA damage. Mol
Cell. 45:801–813. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Foster WH, Langenbacher A, Gao C, Chen J
and Wang Y: Nuclear phosphatase PPM1G in cellular survival and
neural development. Dev Dyn. 242:1101–1109. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
McNamara RP, McCann JL, Gudipaty SA and
D'Orso I: Transcription factors mediate the enzymatic disassembly
of promoter-bound 7SK snRNP to locally recruit P-TEFb for
transcription elongation. Cell Rep. 5:1256–1268. 2013. View Article : Google Scholar : PubMed/NCBI
|