Introduction
Globally, bladder cancer has the 9th highest and
14th highest rates of incidence and mortality of all types of
cancer (1). Approximately 900,000
patients are newly diagnosed each year, with 250,000 deaths
occurring in the same timeframe (2).
Of all newly diagnosed cases of bladder cancer, ~75% are non-muscle
invasive (NMIBC) and ~25% are muscle invasive (MIBC). The
transurethral resection of bladder tumors combined with
intravesical instillation has become the first-line treatment for
high-risk patients with NMIBC. Instillation therapy is usually
applied with Pirarubicin (THP), Epirubicin and Mitomycin (3), with THP being the most commonly
applied, clinically. It is generally accepted that THP kills tumor
cells by inducing cell apoptosis. However, >30% of patients with
NMIBC still exhibit recurrence or the development of MIBC within 5
years (4). Chemoresistance is a
serious challenge to bladder tumor therapy. Therefore, it is
necessary to identify more effective targets to better cope with
chemotherapeutic resistance.
Fibroblast growth factor receptor 3 (FGFR3) is a
member of the FGFR family and is implicated in various cellular
activities, including proliferation, migration, survival and death
(5). FGFR3 mutations are common in
bladder cancer, occurring in ~75% of patients with NMIBC. FGFR3 is
therefore an attractive target for the treatment of bladder cancer
(6,7). It has been determined that FGFR3
mutations are correlated with FGFR3 overexpression (8). Further study has revealed that 74% of
patients with bladder cancer exhibiting a high expression of FGFR3
also possessed an FGFR3 mutation, indicating that FGFR3 expression
is strongly associated with FGFR3 mutation (9). Clinically, chemoresistance occurs
frequently in patients with bladder cancer that exhibit an aberrant
activation of FGFR3. It has been found that chemoresistance may be
associated with FGFR3 mutations (10). It has also been demonstrated that
FGFR3 expression is increased in metastatic tumors compared with
primary bladder cancer (11). It can
therefore be hypothesized that FGFR3 overexpression may be involved
in chemoresistance. Furthermore, resistance commonly occurs as a
result of extensive phosphorylation, which is activated by FGFR3
downstream signaling. FGFR3 activates multiple downstream signaling
pathways, including MAPK, PI3K/AKT and STAT pathways (7,12,13).
MKP-1 is a member of the MKP family that negatively
regulates the MAPK pathway (14). It
also serves a significant role in proliferation, inflammation and
apoptosis (15,16), and is involved in the MAPK pathway.
Its dephosphorylation inactivates JNK, ERK and p38 (17,18).
MKP-1 is overexpressesed in various types of epithelial cancer at
early stages, including gastric, bladder, prostate and colon
carcinomas (19,20). However, MKP-1 expression also
decreases as tumor grades and stages advance (21,22),
indicating that MKP-1 is an essential factor for the determination
of benign and malignant tumors. An increasing number of studies
have revealed that MKP-1 correlates with drug resistance in lung
cancer, ovarian cancer, breast cancer, osteosarcoma and lymphoma
(23–26). MKP-1 has also been revealed to be
overexpressed in NMIBC. However, whether MKP-1 is associated with
chemoresistance in bladder cancer is yet to be fully elucidated.
Furthermore, whether an association exists between FGFR3 and MKP-1
in the regulation of chemoresistance in bladder cancer is yet to be
determined. The present study assessed FGFR3 and MKP-1 in patients
with primary and recurrent bladder cancer, and established a 3D
model to determine the role of MKP-1 in chemoresistance. The
results indicated that MKP-1 inhibited RT112 cell apoptosis, which
served an important role in the chemotherapeutic resistance of
bladder cancer.
Materials and methods
Tissue specimens
Bladder cancer tissues, which were resected by
urologists, were obtained from patients (age range, 47–83 years)
between March 2018 and February 2019. A total of 10 specimens (5
primary and 5 recurrent urothelial carcinoma specimens) were
diagnosed as urothelial carcinoma in accordance with the
histological criteria of the World Health Organization, and 5
recurrent patients all had a history of vesical chemotherapy
instillation of pirarubicin. Following resection, samples were
immediately and stored at −80°C until further analysis. The
clinical characteristics of the patients included in the current
study are presented in Table I. The
present study was approved by the Ethics Committee of The First
Affiliated Hospital of Jiaxing University, and informed consent was
obtained from each patient.
 | Table I.Characteristics of bladder cancer
tissue samples from 10 Chinese patients. |
Table I.
Characteristics of bladder cancer
tissue samples from 10 Chinese patients.
| Patient no. | Sex | Age, years | Histological typing
WHO | Grade | Primary or
recurrent | History of
chemotherapy |
|---|
| 1 | Female | 82 | Urothelial
carcinoma | Low grade | Primary | No |
| 2 | Male | 68 | Urothelial
carcinoma | Low grade | Primary | No |
| 3 | Female | 68 | Urothelial
carcinoma | Low grade | Primary | No |
| 4 | Male | 77 | Urothelial
carcinoma | Low grade | Primary | No |
| 5 | Female | 47 | Urothelial
carcinoma | Low grade | Primary | No |
| 6 | Female | 69 | Urothelial
carcinoma | Low grade | Recurrent | Pirarubicin |
| 7 | Male | 83 | Urothelial
carcinoma | Low grade | Recurrent | Pirarubicin |
| 8 | Female | 81 | Urothelial
carcinoma | Low grade | Recurrent | Pirarubicin |
| 9 | Male | 64 | Urothelial
carcinoma | Low grade | Recurrent | Pirarubicin |
| 10 | Male | 59 | Urothelial
carcinoma | Low grade | Recurrent | Pirarubicin |
Cells, culture and reagents
The human bladder cancer cell line RT112 was
purchased from the Leibniz Institute DSMZ and maintained in RPMI
1640 medium supplemented with 10% fetal bovine serum at 37°C in a
humidified atmosphere containing of 5% CO2. Tissue
culture media and serum were purchased from Hyclone (GE Healthcare
Life Sciences). 3D Cell Culture Gel (cat. no. P720M-10) was
purchased from Col-Tgel Med (http://www.101bio.com/P720_3D_cell_culture_gel.php).
Rabbit polyclonal antibodies against total and phosphorylated JNK,
ERK1/2 and p38 were purchased from Cell Signaling Technology, Inc.
The JNK inhibitor SP600125 (cat. no. T3109), the ERK inhibitor
PD98059 (cat. no. T2623), the p38 inhibitor SB202190 (cat. no.
T2301) and THP were purchased from Target Molecule Corp.
Small interfering RNA (siRNA)
transfection
MKP-1 siRNA and scrambled siRNA were purchased from
GE Healthcare Dharmacon, Inc. The sequences were as follows: MKP-1
siRNA forward, 5′-CACAAGGCAGACATCAGCTC-3′ and reverse,
5′-AGGTAAGCAAGGCAGATGGT-3′; scrambled siRNA forward,
5′-GGGTGTGAACCATGAGAAGT-3′ and reverse, 5′-GACTGTGGTCATGAGTCCT-3′.
RT112 cells were transiently transfected with 100 nM MKP-1 siRNA
and scrambled siRNA in six-well plates treated with TurboFect
reagent according to manufacturer's protocol. After 48 h, cells
were harvested for subsequent experimentation.
Immunohistochemistry
Fixed tissues were dehydrated, embedded, sliced and
dyed, and washed in triplicate. Tissue sections were subsequently
autoclaved in sodium citrate buffer (pH 6.0) for 30 min, after
which goat serum blocking solution was added and samples were
incubated. Primary rabbit polyclonal anti-MKP-1 antibodies
(dilution: 1:500, catlog: NBP2-67909, Novus Biologicals, LLC) were
then added and incubated overnight at 4°C. After being washed three
times, samples were further incubated with biotinylated secondary
antibodies (the Jackson laboratory, Bar Harbor, ME, USA) for 30 min
at room temperature. Sections were then developed using Dimethyl
benzidine for 2 min. Slides were counterstained with hematoxylin
and examined using light microscopy. The labeled substance appeared
yellow-brown under the microscope. Negative control sections were
not incubated with primary antibodies. However, the remaining steps
of the protocol were the same.
Reverse transcription-quantitative PCR
(RT-qPCR)
Cells were lysed and total RNA was isolated using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) according to manufacturer's protocol. RNA was reverse
transcribed using the M-MLV reverse transcription kit (Takara,
Tokyo, Japan). Quantitative PCR was performed using a SYBR Premix
Ex Taq kit (Roche, Basel, Switzerland). The sequences of the
primers obtained from Sangon Biotech Co., Ltd. were as follows:
MKP-1 forward, 5-CCTTTCTGTACCTGGGCAGT-3 and reverse,
5-GGTTGGGACAATTGGCTGAG-3; GAPDH forward, 5-GGGTGTGAACCATGAGAAGT-3
and reverse, 5-GACTGTGGTCATGAGTCCT-3. GAPDH was used as an internal
control. The thermocycling conditions for qPCR were as follows:
94°C for 2 min, followed by 40 cycles at 94°C for 15 sec, 60°C for
1 min and 72°C for 10 min. The 2−ΔΔCq analysis method
was used to calculate relative expression.
Flow cytometry (FACS)
Apoptosis was assessed using an Annexin V-APC kit
(BD Biosciences) in accordance with the manufacturer's protocol.
Treated cells were washed and centrifuged (1,000 rpm, 10 min, 4°C).
The supernatant was resuspended and 500 µl binding buffer, 2 µl
Annexin V-APC and 5 µl propidium iodide was added. Cells were
analyzed using a FACS Canto plus flow cytometer (BD Biosciences)
after 15 min of incubation.
Western blot analysis
Western blot analysis was performed as previously
described (27). Cells were
harvested, washed and centrifuged (1,500 rpm, 5 min, 4°C). An equal
quantity of protein was loaded on 10 or 12% SDS-PAGE gels and
transferred onto nitrocellulose membranes. Membranes were
subsequently blocked with 5% skimmed milk and then hybridized to
primary antibodies overnight at 4°C. Subsequently, samples were
incubated with secondary antibodies for 1 h at room temperature.
Bound antibodies was detected using an enhanced chemiluminescence
reagent in accordance with the to manufacturer's protocol. GAPDH
and actin were used as a loading control.
Cell viability assay
Cell viability was determined using a cell counting
kit 8 (CCK8) assay. Freshly isolated cells (5×103/well)
were seeded into 96-well plates. Cells were pretreated with
SP600125 (20 µmol/l), PD98059 (20 µmol/l) and SB202190 (25 µmol/l)
for 1 h, after which THP (80 nmol/l) was added to each group. After
48 h, 10 µl CCK8 (Beyotime Institute of Biotechnology) was added to
cells and the optical density of each well was measured at 450 nm
using an ELISA reader (BioTek, Winooski, Vermont, USA) according to
the manufacturer's protocol.
Statistical analysis
Statistical analysis was performed using the
Statistical Package of Social Sciences (IBM Corp.; version 20.0.).
Statistical significance was determined using One-way ANOVA and
followed by Tukey post-hoc tests across the groups. Differences
between the two groups were analyzed by Unpaired t-test. Data were
expressed as the mean ± standard deviation and P<0.05 was
considered to indicate a statistically significant difference.
Results
FGFR3 and MKP-1 expression is
increased in recurrent bladder cancer cells
FGFR3 mutations or its expression commonly occurs in
low grade, superficial bladder tumors (8,28). FGFR3
also regulates cellular processes primarily via the MAPK pathway
(13). MKP-1 has been proven to be
associated with chemoresistance in many different types of tumor,
and negatively regulates MAPKs. To determine if FGFR3 and MKP-1 are
associated with chemoresistance in bladder cancer, the current
study collected multiple specimens of primary and recurrent bladder
cancer tissue. FGFR3 and MKP-1 expression were subsequently
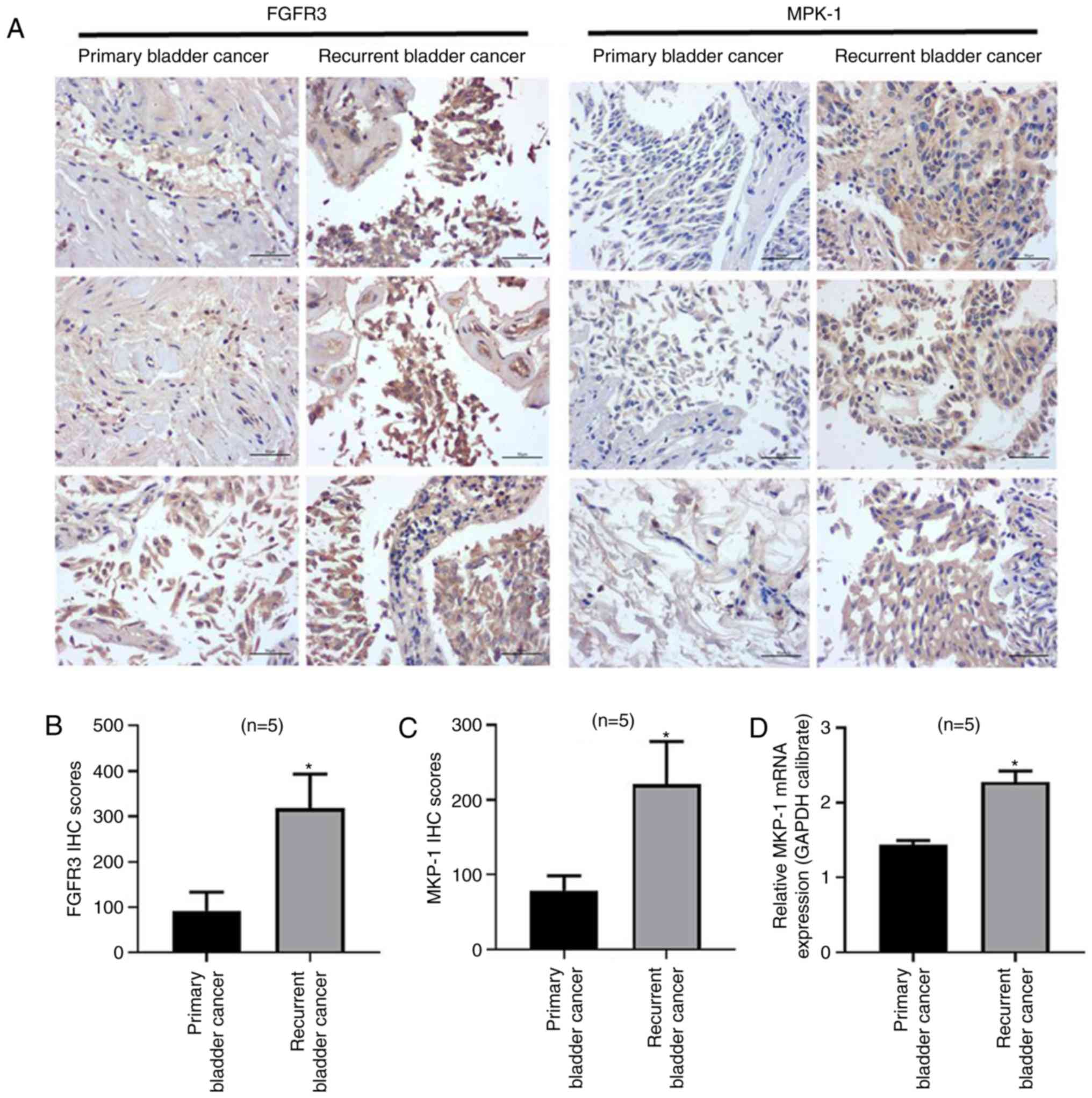
detected using immunohistochemistry (Fig. 1A). the results revealed that FGFR3
and MKP-1 expressions were increased in patients with recurrent
cancer (Fig. 1B and C). Furthermore,
MKP-1 mRNA levels (Fig. 1D) were
increased in patients with recurrent bladder cancer. The results
indicated that MKP-1 and FGFR3 expressions were upregulated in
recurrent tissue, leading to the hypothesis that FGFR3 and MKP-1
may be closely associated with chemoresistance in human bladder
cancer. FGFR3 and MKP-1 may therefore serve as potential targets
for resistance therapy in patients with bladder cancer.
Establishment of human 3D bladder
cancer model and role of MKP-1 in MAPK pathway
To better mimic the development of human bladder
cancer, a novel 3D cellular model was established in the current
study using RT112 cells in accordance with a previously published
method (29). To explore the effect
of MKP-1 in bladder cancer, RT112 cells were transiently
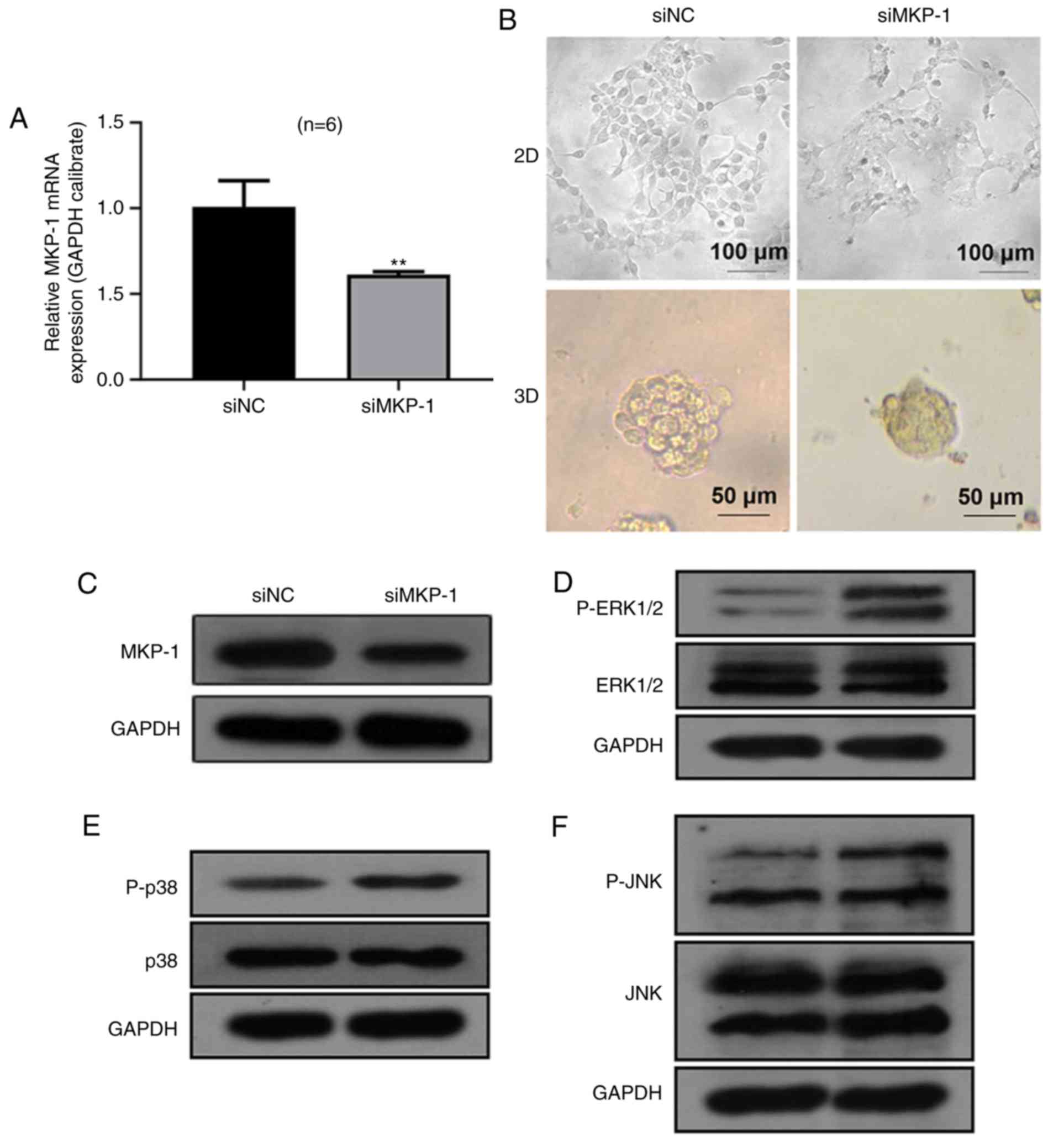
transfected with scrambled siRNA (siNC) and MKP-1 siRNA (siMKP-1).
The results revealed that MKP-1 mRNA levels (Fig. 2A) were significantly decreased in the
siMKP-1 group, demonstrating that MKP-1 was effectively inhibited
by MKP-1 siRNA. RT112 cells of the 2D and 3D model appeared to have
different morphological characteristics (Fig. 2B). Specifically, 3D cells generated
spheroids with a smooth surface and rounded shape. However, 2D
cells appeared irregularly shaped with an obscure margin. The
successful establishment of the 3D model was used for the
subsequent 3D culture of remaining experiments.
MKP-1 serves an important role in the regulation of
MAPK signaling by inactivating JNK, ERK and p38 (30). Thus, the current study hypothesized
that MKP-1 knockdown may activate MAPK. As presented in Fig. 2C, no significant differences were
identified between total levels of JNK, ERK1/2 and p38.
Furthermore, levels of JNK, ERK1/2 and p38 phosphorylation were
markedly increased in the siMKP-1 group, indicating that MKP-1
inhibited the phosphorylation of JNK, ERK1/2 and p38. The results
indicated that MKP-1 serves an essential role in the negative
regulation of JNK, ERK1/2 and p38 in RT112 cells.
MKP-1 affects drug susceptibility and
protects RT112 cells from cell apoptosis
MKP-1 has been implicated in proliferation,
survival, apoptosis and other cellular processes, and has also been
correlated with chemoresistance (23). The current study aimed to establish
the effects of MKP-1 knockdown on chemotherapeutics and to
ascertain whether MKP-1 serves a role in bladder cancer drug
resistance. The current study therefore treated siNC and siMKP-1
cells with gradually increasing doses of THP for 24 h (Fig. 3A). The results revealed that the
viability of siMKP-1 cells was less than siNC cells, indicating
that siMKP-1 cells were more sensitive to THP. As presented in
Fig. 3A, the viability of siNC cells
following THP (80 nmol/l) treatment was significantly higher when
compared with siMKP-1 cells. Extensive exposure of THP for 48 and
72 h (Fig. 3B) demonstrated that
viability was visibly decreased in siMKP-1 cells. Two groups of
cells were then selected and treated with 80 nmol/l THP for 48 h to
detect cell apoptosis via FCAS. The results revealed that cell
apoptosis sharply increased in siMKP-1 cells (Fig. 3C and D), indicating that MKP-1
effectively inhibited RT112 cell apoptosis. Taken together, the
results suggested that MKP-1 protected RT112 cells from cell
apoptosis and served an important role in chemoresistance.
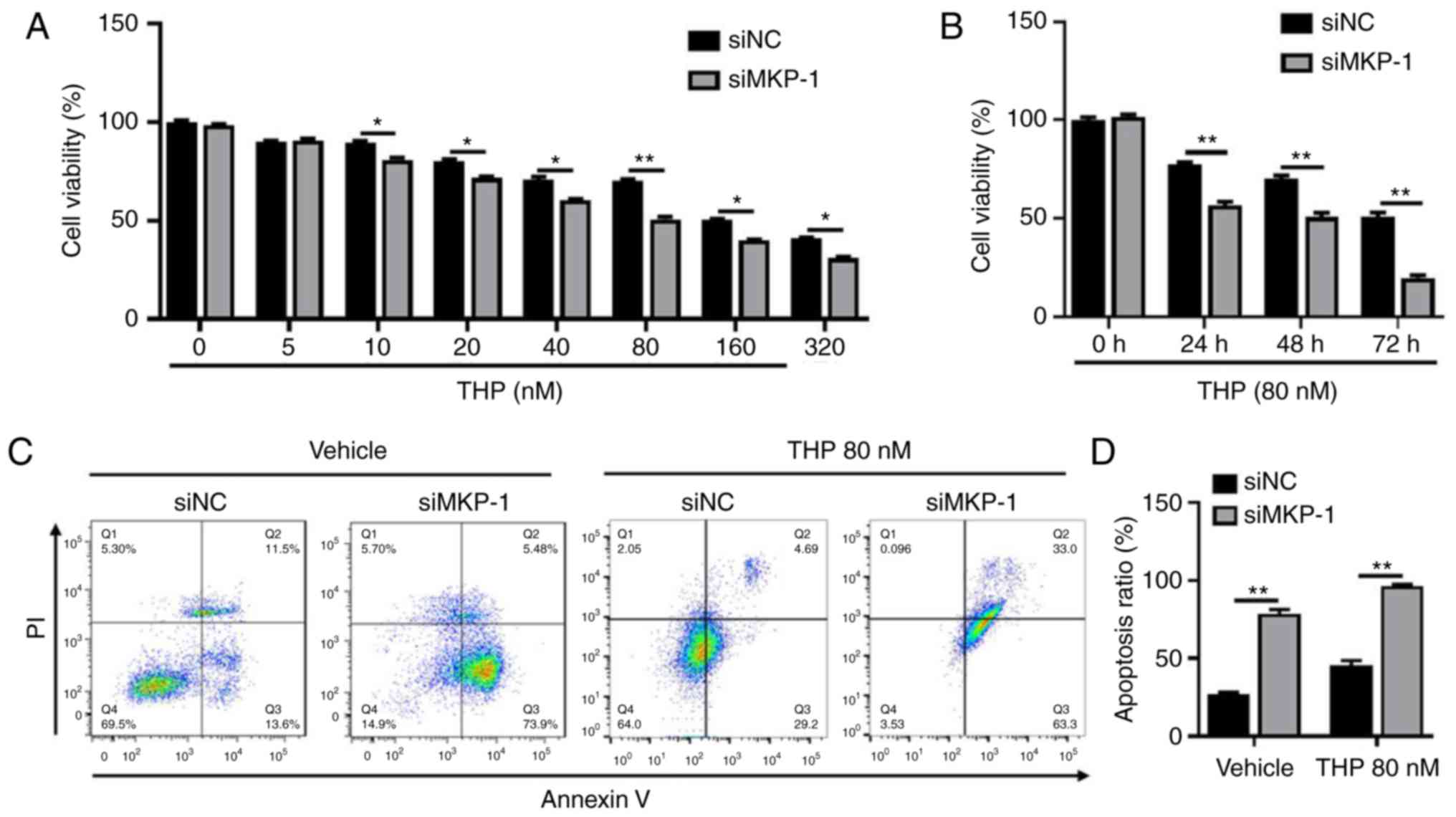 | Figure 3.MKP-1 knockdown increases
drug-susceptibility and induces cell apoptosis. Dose responses and
the timed course of cell death in siNC and siMKP-1 treated cells is
presented. (A) Two groups of cells were treated with increasing
concentrations (0, 5, 10, 20, 40, 80, 160 and 320 nmol/l) of THP
for 24 h. (B) Two groups of cells were treated with 80 nmol/l THP
and incubated for different durations (24, 48 and 72 h). Cell
viability was determined via Cell Counting Kit-8 assays. *P<0.05
and **P<0.01. (C) Cell apoptosis of the two groups was measured
via flow cytometry with or without 80 nmol/l THP treatment. (D) A
comparison of apoptosis rates between the two groups is presented.
**P<0.01. MKP-1, mitogen activated protein kinase phosphatase-1;
siNC, small interfering negative control; siMKP-1, MKP-1 small
interfering RNA; THP, pirarubicin. |
MKP-1 is involved in chemoresistance
via JNK, ERK and p38 pathways
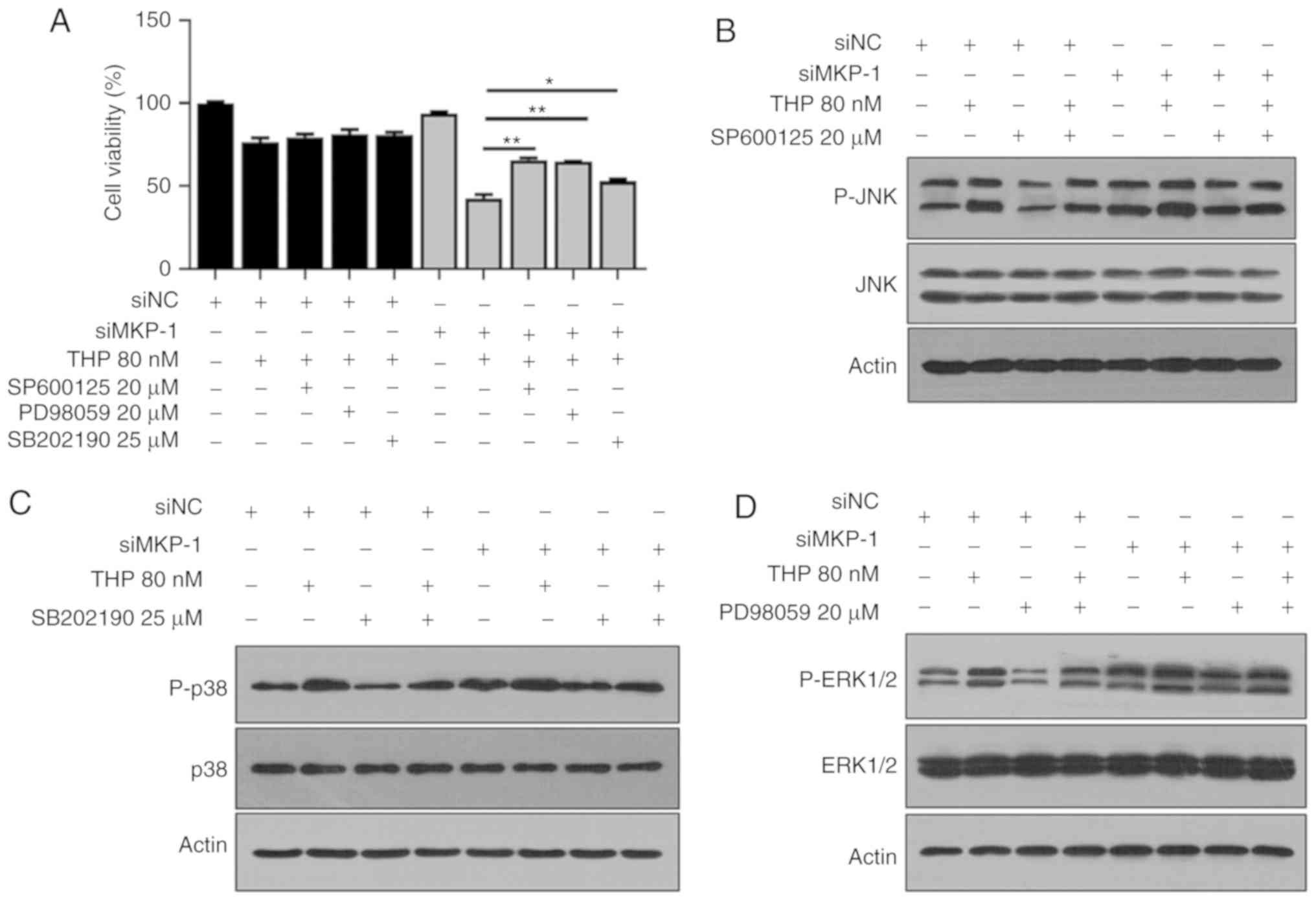
MKP-1 knockdown induced cell apoptosis (Fig. 3C and D). However, the underlying
mechanism of this process remains unclear. As aforementioned, the
current study determined that JNK, ERK and p38 were upregulated in
siMKP-1 cells (Fig. 2C), leading to
the further hypothesis that JNK, ERK and p38 were responsible for
cell apoptosis. To test this hypothesis, siNC and siMKP-1 groups
were individually treated with JNK, ERK and p38 inhibitors
(SP600125, PD98059 and SB202190, respectively). As presented in
Fig. 4A, cell viability markedly
increased in siMKP-1 cells treated with SP600125 and THP when
compared with THP treatment alone. This indicated that SP600125
served a role in protecting siMKP-1 cells from cell death. PD98059
and SB202190 exerted similar effects following treatment with
SP600125. The results also revealed that SP600125 treatment
inhibited JNK phosphorylation in siNC and siMKP-1 cells (Fig. 4B). Treatment with ERK or p38
inhibitors exerted similar effects. The results of Fig. 4 indicated that JNK, ERK and p38 serve
significant roles in RT112 cell death. In conclusion, the results
suggested that MKP-1 may serve a role in chemoresistance through
JNK, ERK and p38 pathways in bladder cancer.
Discussion
Chemotherapy is a primary treatment of bladder
cancer, and patients exhibiting chemoresistance have high
recurrence rates, making it a major challenge to the treatment and
prognosis of patients with bladder cancer (31). There are various mechanisms that have
led to the emergence of chemoresistance, which involve diverse cell
signaling pathways that are often interconnected, hindering bladder
cancer therapy. At present, more studies aiming to identify various
molecular targets have been intitated to create more effective
methods of treating bladder cancer chemoresistance.
Many cell-based resistance assays are performed
using conventional 2D models (32).
However, in vivo cells exist in a 3D environment, which
comprises the extracellular matrix and other neighboring cells. 3D
models also mimic natural cellular responses in vivo,
supporting angiogenesis, slowing proliferation, enhancing
metastatic potential and increasing chemotherapeutic resistance
(33,34). Thus, the current study established a
novel 3D bladder cancer cell model in vitro using
gel-embedding methods.
FGFR3 is expressed on different cells and regulates
biological processes (5) by
triggering multiple signal transduction pathways, including MAPK,
PI3K/AKT and JAK/STAT pathways (5).
It has been demonstrated that FGFR3 mutations occur in myeloma,
cervix and bladder cancer (35,36), and
is the most frequently mutated oncogene in NMIBC, primarily
occurring in exons 7, 10 and 15 (37). Further studies have elucidate the
FGFR3 overexpression and mutations are correlated (8,9). It has
been demonstrated that FGFR3 is highly expressed in patients with
recurrent metastatic bladder cancer (11,38).
Similar results were obtained in the current study, indicating that
results may be associated with chemoresistance (39). The dysregulated activity of FGFR3 may
also mediate chemoresistance via gene mutations (40).
MKP-1, negatively regulates MAPKs through threonine
or tyrosine residues. It has been demonstrated that MKP-1
expression is increased in lung, ovarian and breast cancer
(23–25) after chemotherapy, which indicates
that MKP-1 is closely associated with chemoresistance. MKP-1 has
been demonstrated to exert sustaining tamoxifen resistance in
breast cancer (41) and may induce
resistance by inhibiting cell apoptosis (26,42).
FGFR3 is thought to regulate cell growth and survival by activating
ERK (43), which in turn increases
the expression of MKP-1 (44). MKP-1
is therefore regulated by FGFR3 to a certain extent. In the current
study, it was determined that FGFR3 and MKP-1 expressions increased
in patients with recurrent bladder cancer, and it was inferred that
MKP-1 may serve as a novel target for the treatment of patients
with resistant bladder cancer with FGFR3 overexpression. We must
confess that such a small sample size may become a potential
limitation of the study, but more samples will be collected for
verification later, and future research will be carried out.
MKP-1 regulates the MAPK pathway by inactivating
JNK, ERK and p38. However, previous studies have revealed that p38
and JNK are preferred substrates during cellular responses to
stress (45,46). A second study revealed that MKP-1
mediates cisplatin-induced apoptosis via the JNK pathway in lung
cancer, but not via ERK or p38 pathways. Therefore, JNK, ERK and
p38 may be regulated by MKP-1 depending on the type of cell and
stimulus. The present study determined that JNK, ERK and p38
expression were increased in siMKP-1 cells, indicating that MKP-1
knockdown markedly activates JNK, ERK and p38 expression.
MKP-1 sensitizes RT112 cells to drugs and MKP-1
knockdown enhances THP-susceptibility. The results of the current
study indicated that MKP-1 protected RT112 cells from apoptosis and
that MKP-1 knockdown induced cell apoptosis, which indicated that
MKP-1 was closely associated with chemoresistance. Corresponding
JNK, ERK and p38 inhibitors were subsequently selected to block
corresponding signal expression. The results revealed that JNK,
ERK, p38 knockdown protects siMKP-1 cells from death, which
strongly suggested that MKP-1 may inhibit RT112 cell death.
In conclusion, overexpression of MKP-1 protected
cells from death and the knockdown of MKP-1 induced RT112 cell
death. It was further revealed that the activation of JNK, ERK and
p38 serve important roles in the regulation of cell apoptosis.
MKP-1 may be involved in chemoresistance by inactivating JNK, ERK
and p38, and may therefore lead to the inhibition of apoptosis in
bladder cancer. Therefore, MKP-1 may serve as an effective
therapeutic target for overcoming resistance in bladder cancer.
However, further study is required to determine which upstream
signaling pathways are involved and their specific roles in MAPK
pathway, VEGFR and ROS (47) may
become potential and great future research directions.
Acknowledgements
Not applicable.
Funding
The present study was supported by Jiaxing Science
and Technology Project (grant. nos. 2017AY33004 and 2018AD32083)
and Medical Scientific Research Foundation of Zhejiang Province,
China (grant. no. 2019KY694).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
SL, YH and JJ conceived and designed the study. HX,
NC and WX collected tissue samples and patient data. SL, HX, NC, WX
and HP performed the experiments and the statistical analyses. SL
and XH analyzed the data and wrote the manuscript. YH and JJ
reviewed and revised the manuscript. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of The First Hospital of The First Affiliated Hospital of
Jiaxing University (approval no. 2016-056), and informed consent
was obtained from all patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
MKP-1
|
mitogen activated protein kinase
phosphatase-1
|
|
THP
|
Pirarubicin
|
|
FGFR3
|
fibroblast growth factor 3
|
|
NMIBC
|
non-muscle invasive bladder cancer
|
|
MIBC
|
muscle invasive bladder cancer
|
|
TURBT
|
transurethral resection of the bladder
tumor
|
|
ERK
|
extracellular signal-regulated
kinase
|
|
JNK
|
c-Jun NH2-terminal kinase
|
|
MAPK
|
mitogen activated protein kinases
|
|
MKPs
|
MAPK phosphatases
|
References
|
1
|
Mahdavifar N, Ghoncheh M, Pakzad R,
Momenimovahed Z and Salehiniya H: Epidemiology, incidence and
mortality of bladder cancer and their relationship with the
development index in the world. Asian Pac J Cancer Prev.
17:381–386. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Klotz L and Brausi MA: World urologic
oncology federation bladder cancer prevention program: A global
initiative. Urol Oncol. 33:25–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Liu S, Hou J, Zhang H, Wu Y, Hu M, Zhang
L, Xu J, Na R, Jiang H and Ding Q: The evaluation of the risk
factors for non-muscle invasive bladder cancer (NMIBC) recurrence
after transurethral resection (TURBt) in Chinese population. PLoS
One. 10:e01236172014. View Article : Google Scholar
|
|
4
|
Miki T, Nonomura N, Kojima Y, Okuyama A,
Nakano E, Kiyohara H, Fujioka H, Koide T, Wakatsuki A, Kuroda H, et
al: A randomized study on intravesical pirarubicin (THP)
chemoprophylaxis of recurrence after transurethral resection of
superficial bladder cancer. Hinyokika Kiyo. 43:907–912. 1997.(In
Japanese). PubMed/NCBI
|
|
5
|
Turner N and Grose R: Fibroblast growth
factor signalling: From development to cancer. Nat Rev Cancer.
10:116–129. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
di Martino E, Tomlinson DC and Knowles MA:
A decade of FGF receptor research in bladder cancer: Past, present,
and future challenges. Adv Urol. 2012:4292132012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Acquaviva J, He S, Zhang C, Jimenez JP,
Nagai M, Sang J, Sequeira M, Smith DL, Ogawa LS, Inoue T, et al:
FGFR3 translocations in bladder cancer: Differential sensitivity to
HSP90 inhibition based on drug metabolism. Mol Cancer Res.
12:1042–1054. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bodoor K, Ghabkari A, Jaradat Z, Alkhateeb
A, Jaradat S, Al-Ghazo MA, Matalka I, Musleh H and Haddad Y: FGFR3
mutational status and protein expression in patients with bladder
cancer in a Jordanian population. Cancer Epidemiol. 34:724–732.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Poyet C, Hermanns T, Zhong Q, Drescher E,
Eberli D, Burger M, Hofstaedter F, Hartmann A, Stöhr R, Zwarthoff
EC, et al: Positive fibroblast growth factor receptor 3
immunoreactivity is associated with low-grade non-invasive
urothelial bladder cancer. Oncol Lett. 10:2753–2760. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
McConkey DJ, Choi W, Marquis L, Martin F,
Williams MB, Shah J, Svatek R, Das A, Adam L, Kamat A, et al: Role
of epithelial-to-mesenchymal transition (EMT) in drug sensitivity
and metastasis in bladder cancer. Cancer Metastasis Rev.
28:335–344. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Guancial EA, Werner L, Bellmunt J, Bamias
A, Choueiri TK, Ross R, Schutz FA, Park RS, O'Brien RJ, Hirsch MS,
et al: FGFR3 expression in primary and metastatic urothelial
carcinoma of the bladder. Cancer Med. 3:835–844. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
L'Hote CG and Knowles MA: Cell responses
to FGFR3 signalling: Growth, differentiation and apoptosis. Exp
Cell Res. 304:417–431. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Katoh M and Nakagama H: FGF receptors:
Cancer biology and therapeutics. Med Res Rev. 34:280–300. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wu GS: Role of mitogen-activated protein
kinase phosphatases (MKPs) in cancer. Cancer Metastasis Rev.
26:579–585. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Owens DM and Keyse SM: Differential
regulation of MAP kinase signalling by dual-specificity protein
phosphatases. Oncogene. 26:3203–3213. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Boutros T, Chevet E and Metrakos P:
Mitogen-activated protein (MAP) kinase/MAP kinase phosphatase
regulation: Roles in cell growth, death, and cancer. Pharmacol Rev.
60:261–310. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sun H, Charles CH, Lau LF and Tonks NK:
MKP-1 (3CH134), an immediate early gene product, is a dual
specificity phosphatase that dephosphorylates MAP kinase in vivo.
Cell. 75:487–493. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li J, Gorospe M, Hutter D, Barnes J, Keyse
SM and Liu Y: Transcriptional induction of MKP-1 in response to
stress is associated with histone H3 phosphorylation-acetylation.
Mol Cell Biol. 21:8213–8224. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bang YJ, Jin HK, Kang SH, Kim JW and Yun
CY: Increased MAPK activity and MKP-1 overexpression in human
gastric adenocarcinoma. Biochem Biophys Res Commun. 250:43–47.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Haagenson KK and Wu GS: Mitogen activated
protein kinase phosphatases and cancer. Cancer Biol Ther.
9:337–340. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Loda M, Capodieci P, Mishra R, Yao H,
Corless C, Grigioni W, Wang Y, Magi-Galluzzi C and Stork PJ:
Expression of mitogen-activated protein kinase phosphatase-1 in the
early phases of human epithelial carcinogenesis. Am J Pathol.
149:1553–1564. 1996.PubMed/NCBI
|
|
22
|
Manzano RG, Montuenga LM, Dayton M, Dent
P, Kinoshita I, Vicent S, Gardner GJ, Nguyen P, Choi YH, Trepel J,
et al: CL100 expression is down-regulated in advanced epithelial
ovarian cancer and its re-expression decreases its malignant
potential. Oncogene. 21:4435–4447. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang Z, Xu J, Zhou JY, Liu Y and Wu GS:
Mitogen-activated protein kinase phosphatase-1 is required for
cisplatin resistance. Cancer Res. 66:8870–8877. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang J, Zhou JY and Wu GS: ERK-dependent
MKP-1-mediated cisplatin resistance in human ovarian cancer cells.
Cancer Res. 67:11933–11941. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Haagenson KK and Wu GS: The role of MAP
kinases and MAP kinase phosphatase-1 in resistance to breast cancer
treatment. Cancer Metastasis Rev. 29:143–149. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang Z, Zhou JY, Kanakapalli D, Buck S, Wu
GS and Ravindranath Y: High level of mitogen-activated protein
kinase phosphatase-1 expression is associated with cisplatin
resistance in osteosarcoma. Pediatr Blood Cancer. 51:754–759. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zheng L, Chen J, Ma Z, Liu W, Yang F, Yang
Z, Wang K, Wang X, He D, Li L and Zeng J: Capsaicin enhances
anti-proliferation efficacy of pirarubicin via activating TRPV1 and
inhibiting PCNA nuclear translocation in 5637 cells. Mol Med Rep.
13:881–887. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kimura T, Suzuki H, Ohashi T, Asano K,
Kiyota H and Eto Y: The incidence of thanatophoric dysplasia
mutations in FGFR3 gene is higher in low-grade or superficial
bladder carcinomas. Cancer. 92:2555–2561. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ringuette Goulet C, Bernard G, Chabaud S,
Couture A, Langlois A, Neveu B, Pouliot F and Bolduc S:
Tissue-engineered human 3D model of bladder cancer for invasion
study and drug discovery. Biomaterials. 145:233–241. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Camps M, Nichols A and Arkinstall S: Dual
specificity phosphatases: A gene family for control of MAP kinase
function. FASEB J. 14:6–16. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Arakawa M, Nakamura K, Yamada Y, Kato K,
Katsuda R, Tobiume M, Zennami K, Watanabe M, Kato Y, Nishikawa G,
et al: Intravesical administration of pirarubicin against
superficial bladder cancer: Relationship between tumor tissue
concentration and exposure time in the bladder or therapeutic
effect. Exp Ther Med. 2:901–905. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Edmondson R, Broglie JJ, Adcock AF and
Yang L: Three-dimensional cell culture systems and their
applications in drug discovery and cell-based biosensors. Assay
Drug Dev Technol. 12:207–218. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Imamura Y, Mukohara T, Shimono Y,
Funakoshi Y, Chayahara N, Toyoda M, Kiyota N, Takao S, Kono S,
Nakatsura T and Minami H: Comparison of 2D- and 3D-culture models
as drug-testing platforms in breast cancer. Oncol Rep.
33:1837–1843. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li C, Singh B, Graves-Deal R, Ma H,
Starchenko A, Fry WH, Lu Y, Wang Y, Bogatcheva G, Khan MP, et al:
Three-dimensional culture system identifies a new mode of cetuximab
resistance and disease-relevant genes in colorectal cancer. Proc
Natl Acad Sci USA. 114:E2852–E2861. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chesi M, Nardini E, Brents LA, Schröck E,
Ried T, Kuehl WM and Bergsagel PL: Frequent translocation
t(4;14)(p16.3;q32.3) in multiple myeloma is associated with
increased expression and activating mutations of fibroblast growth
factor receptor 3. Nat Genet. 16:260–264. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Cappellen D, De Oliveira C, Ricol D, de
Medina S, Bourdin J, Sastre-Garau X, Chopin D, Thiery JP and
Radvanyi F: Frequent activating mutations of FGFR3 in human bladder
and cervix carcinomas. Nat Genet. 23:18–20. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hernández S, López-Knowles E, Lloreta J,
Kogevinas M, Amorós A, Tardón A, Carrato A, Serra C, Malats N and
Real FX: Prospective study of FGFR3 mutations as a prognostic
factor in nonmuscle invasive urothelial bladder carcinomas. J Clin
Oncol. 24:3664–3671. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Rosenberg JE, Werner L, Bamias A, Choueiri
TK, Schutz FAB, O'Brien RSP, Guancial EA, Ross RW, Berman DM,
Riester M, et al: FGFR3 protein expression and gene mutation in
primary and metastatic urothelial carcinoma (UC) tumors. J Clin
Oncol 30 (15 Suppl). S4572012.
|
|
39
|
Chell V, Balmanno K, Little AS, Wilson M,
Andrews S, Blockley L, Hampson M, Gavine PR and Cook SJ: Tumour
cell responses to new fibroblast growth factor receptor tyrosine
kinase inhibitors and identification of a gatekeeper mutation in
FGFR3 as a mechanism of acquired resistance. Oncogene.
32:3059–3070. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wu K, Wang B, Chen Y, Zhou J, Huang J, Hui
K, Zeng J, Zhu J, Zhang K, Li L, et al: DAB2IP regulates the
chemoresistance to pirarubicin and tumor recurrence of non-muscle
invasive bladder cancer through STAT3/Twist1/P-glycoprotein
signaling. Cell Signal. 27:2515–2523. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ma G, Pan Y, Zhou C, Sun R, Bai J, Liu P,
Ren Y and He J: Mitogen-activated protein kinase phosphatase 1 is
involved in tamoxifen resistance in MCF7 cells. Oncol Rep.
34:2423–2430. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Rojo F, González-Navarrete I, Bragado R,
Dalmases A, Menéndez S, Cortes-Sempere M, Suárez C, Oliva C,
Servitja S, Rodriguez-Fanjul V, et al: Mitogen-activated protein
kinase phosphatase-1 in human breast cancer independently predicts
prognosis and is repressed by doxorubicin. Clin Cancer Res.
15:3530–3539. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Hu Y, Mintz A, Shah SR, Quinones-Hinojosa
A and Hsu W: The FGFR/MEK/ERK/brachyury pathway is critical for
chordoma cell growth and survival. Carcinogenesis. 35:1491–1499.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Brondello JM, Brunet A, Pouysségur J and
Mckenzie FR: The dual specificity mitogen-activated protein kinase
phosphatase-1 and −2 are induced by the p42/p44MAPK cascade. J Biol
Chem. 272:1368–1376. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Franklin CC and Kraft AS: Conditional
expression of the mitogen-activated protein kinase (MAPK)
phosphatase MKP-1 preferentially inhibits p38 MAPK and
stress-activated protein kinase in U937 cells. J Biol Chem.
272:16917–16923. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhao Q, Shepherd EG, Manson ME, Nelin LD,
Sorokin A and Liu Y: The role of mitogen-activated protein kinase
phosphatase-1 in the response of alveolar macrophages to
lipopolysaccharide: Attenuation of proinflammatory cytokine
biosynthesis via feedback control of p38. J Biol Chem.
280:8101–8108. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Takeuchi H, Taoka R, Mmeje CO, Jinesh GG,
Safe S and Kamat AM: CDODA-Me decreases specificity protein
transcription factors and induces apoptosis in bladder cancer cells
through induction of reactive oxygen species. Urol Oncol.
34:337.e11–e18. 2016. View Article : Google Scholar
|