Introduction
Gastric cancer (GC) is the fourth most common
malignancy worldwide and is the second leading cause of
cancer-associated mortality (1).
Patients with chronic inflammation due to Helicobacter
pylori infection, heavy alcohol drinking, heavy smoking and
excessive salt intake are at high risks of developing GC (2). In particular, the rate of
Helicobacter pylori infection in Japanese has been reported
to be high among developed countries (3). Although patients with early GC are
curable by endoscopic surgery, patients with advanced stages are
usually treated with systemic chemotherapy (4). Despite advances in anticancer agents,
the overall 5-year survival rate of patients with GC remains low
(20%) (5).
Programmed cell death protein 1 (PD-1) and its
ligand programmed death-ligand 1 (PD-L1) are important immune
checkpoints in the tumor (6), and it
was reported that the PD-1/PD-L1 pathway functions as adaptive
immune escape machinery (7,8). Therefore, blockade of the PD-1/PD-L1
pathway by immune checkpoint inhibitors (ICIs), including
pembrolizumab and nivolumab, has already been clinically applied
for a variety of cancers, including GC (9). PD-L1 is expressed at the surface of
tumor cells, tumor-associated macrophages (TAMs) and T lymphocytes
and its expression can be induced by cytokines, such as interferons
(IFNs) and tumor necrosis factors (TNFs) (10,11).
Recent studies reported that the soluble form of PD-L1 (sPD-L1) is
detected in the blood of patients with tumors (12,13).
However, the underlying mechanisms remain unknown.
The present study aimed to evaluate PD-L1 expression
and sPD-L1 secretion in GC cells following treatment with IFN-γ.
Furthermore, ELISA was used to examine the serum level of sPD-L1 in
patients with GC to examine its utility as a candidate
biomarker.
Materials and methods
Cell culture and reagents
The human GC cell lines MKN1, MKN74, KATO III,
OCUM-1 were obtained from the Health Science Research Resources
Bank. MKN1, MKN74, and OCUM-1 cells were cultured in Dulbecco's
modified Eagle's medium (Invitrogen; Thermo Fisher Scientific,
Inc.) and KATO III cells were cultured in RPMI-1640 medium
(Invitrogen; Thermo Fisher Scientific, Inc.) supplemented with 10%
fetal calf serum (Invitrogen; Thermo Fisher Scientific, Inc.) and
1% penicillin/streptomycin (Invitrogen; Thermo Fisher Scientific,
Inc.) placed at 37°C in a humidified incubator containing 5%
CO2. Recombinant human interferon (IFN)-γ and TNF-α were
obtained from PeproTech, Inc.
Reverse transcription-quantitative
(RT-q) PCR
Total RNA was extracted from all GC cells treated
with 1, 10 or 100 ng/ml TNF-α or IFN-γ for 24 h using RNeasy Mini
Kit (Qiagen, Inc.) according to the manufacturer's instructions.
Quantitative real-time PCR was performed with an MX3000P qPCR
system (Stratagene; Agilent) using the Universal Probe Library
System (Roche Diagnostics GmbH) according to the manufacturer's
protocol. The thermocycling conditions were: Initial denaturation
at 95°C for 10 min, followed by 40 cycles of 95°C for 30 sec, 55°C
for 60 sec, and 72°C for 60 sec. The sequences of the primers were
as follows: PD-L1, forward 5′-AAATGGAACCTGGCGAAAG-3′, reverse
5′-GCTCCCTGTTTGACTCCATC-3′; and GAPDH, forward
5′-CTGACTTCAACAGCGACACC-3′ and reverse
5′-TAGCCAAATTCGTTGTCATACC-3′. Calculation of relative gene
expression was performed using 2−ΔΔCq method (14).
Immunocytochemistry
All GC cells were fixed with 2% paraformaldehyde for
10 min at room temperature and blocked with 10% normal goat serum
(Invitrogen; Thermo Fisher Scientific, Inc.) for 30 min at room
temperature. Cells were subsequently stained with anti-PD-L1
antibody (1:100; cat. no. 13684; Cell Signaling Technology, Inc.)
at 4°C overnight, followed by incubation with Alexa 555-conjugated
immunoglobulin G secondary antibody (1:400; cat. no. A27039;
Molecular Probes; Thermo Fisher Scientific, Inc.) for 60 min at
room temperature. Subsequently, the cover slip was mounted on each
well with the help of a mounting medium containing
4′,6-diamidino-2-phenylindole (DAPI) (Vector Laboratories, Inc.).
The stained cells were then observed by fluorescence microscopy
(IX70; Olympus Corp.) at ×200 magnification.
Flow cytometric analysis
GC cells non-treated (Mock) or treated with 1, 10 or
100 ng/ml IFN-γ for 24 h were subjected to flow cytometric
analysis. Single GC cell suspensions were stained with
allophycocyanin (APC)-conjugated anti-PD-L1 antibody (BioLegend,
Inc.) at a final concentration of 1 µg/ml for 30 min at 4°C.
Subsequently, 1 µg/ml propidium iodide (PI) was added to eliminate
dead cells for 10 min at room temperature. Flow cytometry analyses
were performed using FACSCanto II (BD Biosciences). After gating
for PI-negative cells (viable cells), PD-L1 expression was
analyzed. Data were analyzed using Flow Jo software (version
10.5.2; TreeStar Inc.).
Western blotting
GC cells non-treated (Mock) or treated with IFN-γ
(100 ng/ml) for 24 h were subjected to western blotting according
to a previous study (15).
Quantification of total protein in samples was performed by
bicinchoninic acid protein assay (Thermo Fisher Scientific, Inc.).
Briefly, whole cell lysates were prepared using 0.1% NP-40 lysis
buffer (20 mM sodium phosphate, Ph 7.0; 300 mM NaCl, 5 mM EDTA and
0.1% NP40) supplemented with protease and phosphatase inhibitor
cocktails (Roche Diagnostics) or sodium dodecyl sulfate
(SDS)-sample buffer (25 mM Tris, pH 6.8; 1% SDS, 5% glycerol, 0.05%
bromophenol blue and 1% β-mercaptoethanol). These lysates (50 µg of
protein/ lane) were separated by 8% SDS-PAGE gels (Bio-Rad
Laboratories, Inc.) and then transferred to a poly vinylidene
di-fluoride (PVDF) membrane (Bio-Rad Laboratories, Inc.). The
membranes were blocked with 5% bovine serum albumin (Sigma-Aldrich,
Merck KGaA) for 1 h at room temperature. Subsequently, they were
blotted with primary antibodies against PD-L1 (1:1,000; cat. no.
13684; Cell Signaling Technology, Inc.) and heat shock protein 90
(HSP90; 1:1,000; cat. no. 610418; BD Biosciences) at 4°C overnight.
After washing, the membranes were incubated with anti-rabbit
horseradish peroxidase (HRP)-conjugated secondary antibody
(1:4,000; cat. no. NA934; GE Healthcare Life Sciences) or
anti-mouse HRP-conjugated secondary antibody (1:4,000; cat. no.
NA931; GE Healthcare Life Sciences) for 1 h at room temperature.
The membranes were washed again and developed with Immobilon
Western Chemiluminescent HRP substrate (EMD Millipore) and the
signals were detected using ChemiDoc XRS systems (Bio-Rad
Laboratories, Inc.). Hsp90 was used as the loading control. To
quantify band intensity, densitometry was performed using Image Lab
4.1 software (Bio-Rad Laboratories, Inc.).
Patients and blood samples
Blood samples (10 ml) were collected before
treatment initiation from 40 patients with GC and 10 healthy
controls. Patients with GC (median age, 71.5 years; age range, 47-
94 years) who were treated at Chiba University Hospital between
June 2018 and December 2019 were analyzed. Tumor size, lymph node
metastasis, distant metastasis and tumor stage were determined
according to the Tumor-Node-Metastasis classification for GC
(16). Healthy controls (median age,
72.5 years; age range, 37–86 years) who visited Chiba University
Hospital from February to March in 2020 were studied. After
centrifuging for 5 min at 1,600 × g, the supernatant serum was
removed and stored at −80°C. After obtaining written informed
consent, blood samples were analyzed for measurement of serum
sPD-L1 concentration and data were acquired from the medical record
of each participant. This study was approved by the Research Ethics
Committees of the Graduate School of Medicine, Chiba University
(approval. no. 3552 and 3671).
ELISA
sPD-L1 levels in the culture supernatant of GC
cells, and the serum of patients with GC and healthy controls were
determined using a sandwich ELISA kit (cat. no. DB7H10; R&D
Systems, Inc.) according to the manufacturer's instructions. sPD-L1
level in the supernatant of GC cells treated with IFN-γ (100 ng/ml)
was determined using 100 µl culture supernatants collected 24 and
48 h after seeding 100,000 cells in 60 mm dishes. Serum sPD-L1
concentration in patients with GC and healthy controls were also
measured before treatment initiation.
Statistical analysis
Data were presented as the means ± standard error of
the mean. The significance of differences between two and multiple
groups were analyzed using Mann-Whitney U test and two ways-ANOVA
followed by post-hoc Tukey's test, respectively. P<0.05 was
considered to indicate a statistically significant difference.
Results
PD-L1 expression in GC cells treated
with TNF-α and IFN-γ
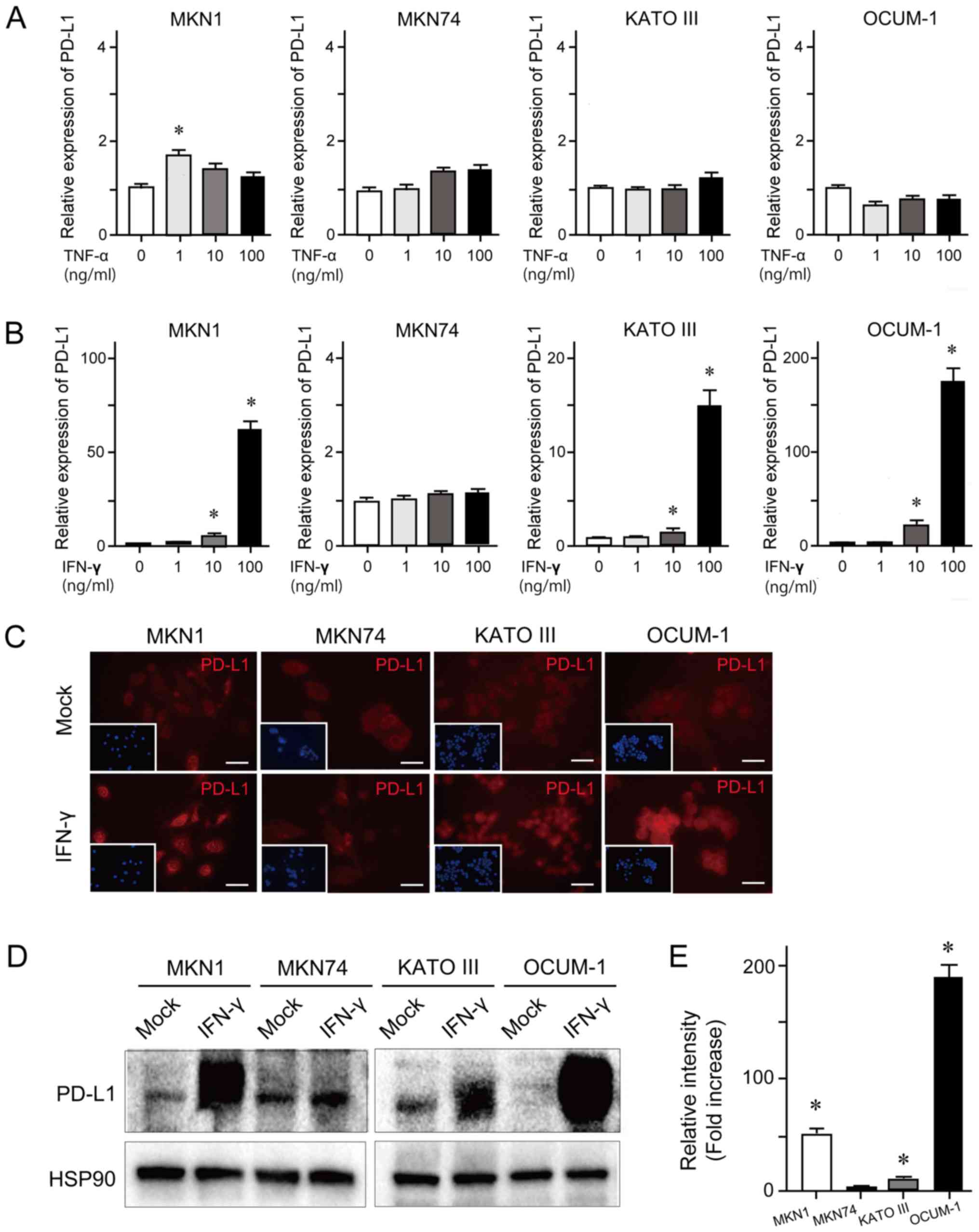
To examine cytokine-induced PD-L1 expression,
RT-qPCR was performed in the GC cell lines MKN1, MKN74, KATO III
and OCUM-1. These cells were treated with 1, 10 or 100 ng/ml TNF-α
or IFN-γ for 24 h. The results demonstrated that TNF-α treatment at
the various concentrations had no effect on PD-L1 expression level
(Fig. 1A). However, IFN-γ treatment
induced an increased expression level of PD-L1 in a dose-dependent
manner in MKN1, KATO III and OCUM-1 cell lines (Fig. 1B). Concordant with these findings,
the results from immunocytochemistry and western blotting
demonstrated that IFN-γ treatment (100 ng/ml) for 24 h enhanced
PD-L1 expression in MKN1, KATO III and OCUM-1 cell lines (Fig. 1C and D). The results from western
blotting quantification revealed that PD-L1 expression was
significantly upregulated in MKN1, KATO III and OCUM-1 cells
treated with IFN-γ compared with mock-treated cells (Fig. 1E).
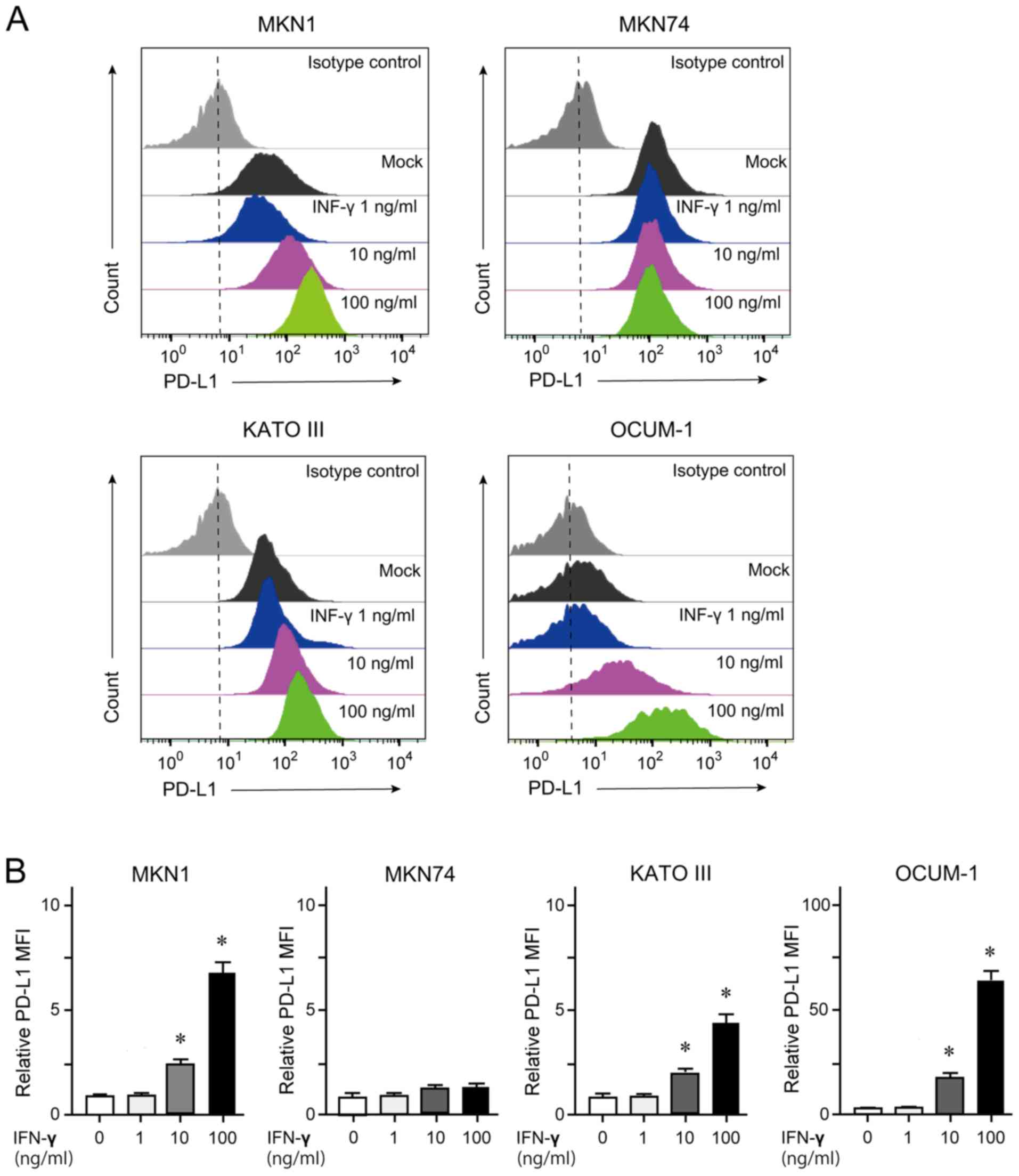
Quantification of membrane-bound PD-L1
by flow cytometry
GC cells treated with 100 ng/ml IFN-γ for 24 h were
subjected to flow cytometry analyses. The results demonstrated that
MKN1, KATO III and OCUM-1 cells, but not MKN74 cells, showed an
increase in membrane-bound PD-L1 expression in a dose-dependent
manner (Fig. 2).
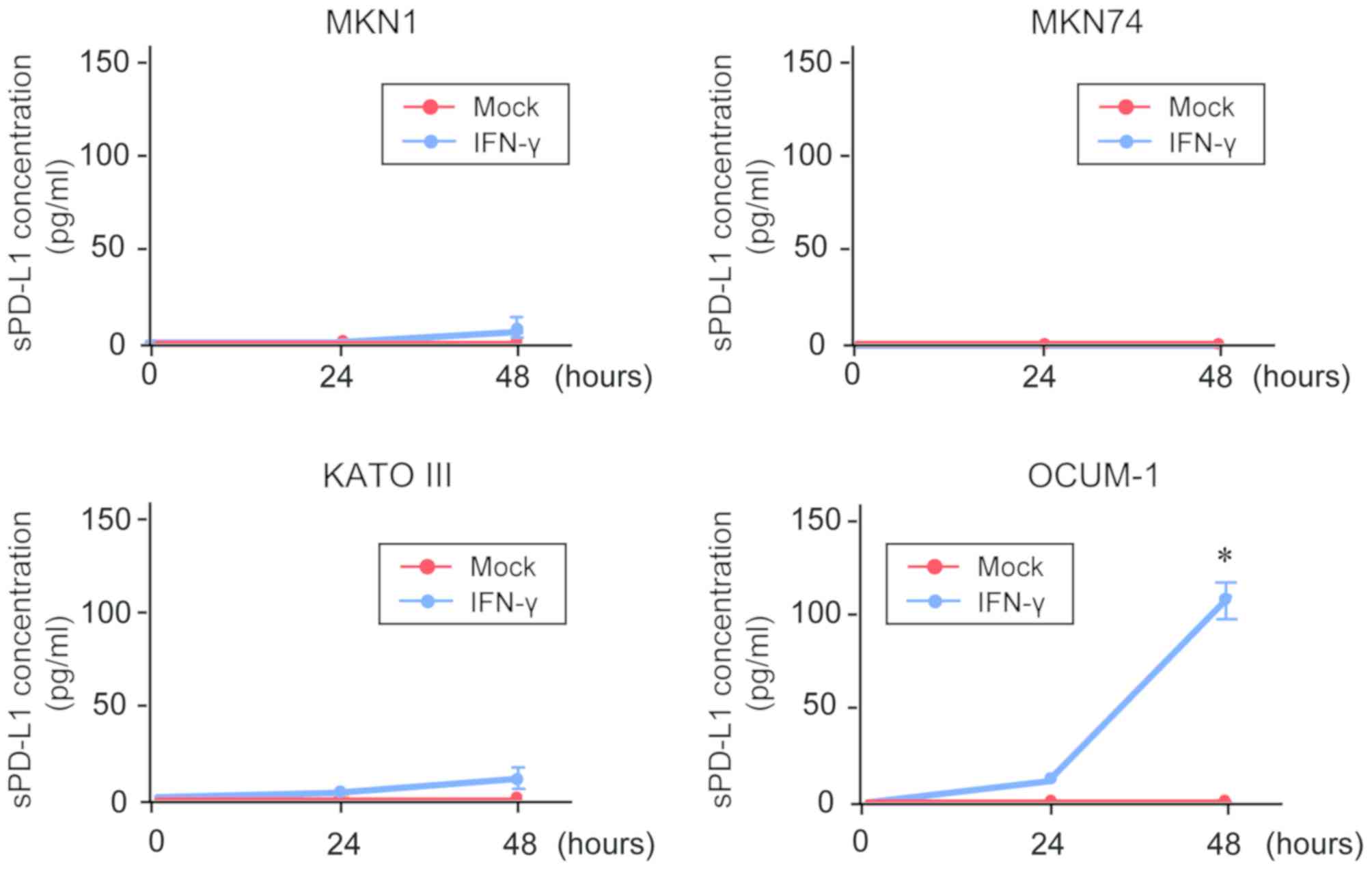
sPD-L1 detection in the culture
supernatant of GC cells
To investigate the association between
membrane-bound PD-L1 expression and sPD-L1 production, an ELISA was
conducted for the measurement of sPD-L1 in the culture supernatant
of GC cells (Fig. 3). The results
demonstrated that sPD-L1 was not detected in the supernatant of
untreated-GC cells. In addition, only sPD-L1 level in the
supernatant of OCUM-1cells was significantly increased following
IFN-γ treatment in a time-dependent manner.
Serum sPD-L1 concentrations in GC
patients
ELISA was conducted to measure the serum PD-L1
concentration in patients with GC and healthy controls (Table I). The serum PD-L1 level in patients
with GC was significantly increased compared with controls
(P<0.05; Fig. 4A). The median
sPD-L1 levels in controls and patients with GC were 20.8 and 33.8
pg/ml, respectively. Furthermore, the serum PD-L1 level in patients
with GC was evaluated according to stage progression. The results
demonstrated that the serum PD-L1 level in patients with stage IV
GC was significantly increased compared with that in patients with
stages I, II or III GC (P<0.05; Fig.
4B).
 | Table I.Clinicopathological characteristics
of patients with gastric cancer. |
Table I.
Clinicopathological characteristics
of patients with gastric cancer.
|
Characteristics | Value (n=40) |
|---|
| Age, years
(range) | 71.5 (47–94) |
| Sex, n
(male/female) | 26/14 |
| BMI,
kg/m2 (range) | 21.2
(15.6–32.0) |
| Smoking, n |
|
|
Yes | 25 |
| No | 15 |
| Alcohol intake,
n |
|
|
Yes | 24 |
| No | 16 |
| Helicobacter
pylori, n |
|
|
Positive | 20 |
|
Negative | 20 |
| Tumor markers,
ng/ml (range) |
|
|
CEA | 2.85
(0.5–86.9) |
|
CA19-9 | 19.1 (0–719.0) |
| Stagea, n (%) |
|
| I | 10 (25) |
| II | 10 (25) |
|
III | 10 (25) |
| IV | 10 (25) |
| Tumora, n (%) |
|
| T1 | 8 (20) |
| T2 | 4 (10) |
| T3 | 14 (35) |
| T4 | 14 (35) |
| Lymph node
metastasisa, n (%) |
|
| N1 | 14 (35) |
| N2 | 5 (12.5) |
| N3 | 8 (20) |
| N4 | 13 (32.5) |
| Distant
metastasisa, n (%) |
|
| M0 | 30 (75.0) |
| M1 | 10 (25.0) |
| Histological
finding, n (%) |
|
|
Diffuse | 19 (47.5) |
|
Intestinal | 17 (42.5) |
|
Mix | 4 (10.0) |
Discussion
PD-1 is a single-pass type I membrane protein that
belongs to the CD28/CTLA-4 family (17). PD-1 is mainly expressed at the
surface of immunocompetent cells, including T lymphocytes, B
lymphocytes and natural killer cells (18). In addition, high expression level of
PD-1 is associated with T-cell exhaustion (19). PD-L1 has been determined as B7
homolog 1 (B7-H1) and functions as a ligand for PD-1 (20). In normal tissues, PD-1/PD-L1 binding
prevents an excessive immune response and protects tissues from
damage through the induction of immune tolerance (21). However, PD-L1/PD-1-mediated tumor
immune escape attenuates the immune response in cancer tissues and
makes the elimination of cancer cells difficult (22). Furthermore, PD-L1 is a target for
hypoxia-inducible factor-1, and PD-L1 expression is further
upregulated under hypoxia (23).
Overall, PD-L1 expression is closely associated with cancer
development and progression.
Both aberrant expression of PD-L1 and uncontrolled
PD-L1/PD-1 signaling are observed with variable frequency in
various types of cancer, such as lung cancer and GC (24). A previous study demonstrated by
immunohistochemistry that PD-L1 expression is detected in ~40% of
GC tissues analyzed and is correlated with both aggressiveness and
unfavorable prognosis (25). PD-L1
expression is regulated by inflammatory signaling, oncogenic
signaling and genetic and epigenetic alterations (26,27). In
addition, the co-existence of PD-L1-positive cancer cells and tumor
infiltrating lymphocytes (TILs) has been reported to be associated
with a poor prognosis in patients with GC (28).
Inflammatory cytokines, including IFNs, TNFs and
interleukins, are mainly released form TILs and upregulate PD-L1
expression in various types of cancer cells, such as lung cancer,
breast cancer, and GC (29).
Similarly, the present study demonstrated that IFN-γ treatment
induced a significant increase in PD-L1 expression in the GC cell
lines MKN1, KATO III and OCUM-1. Furthermore, TNF-α treatment
modestly increased PD-L1 expression in these cell lines. Although
IFN-γ induces PD-L1 expression by stimulating the janus kinase
(JAK)/signal transducer and activator of transcription (STAT)
signaling pathway, scarce expression of STAT1 have been reported in
some GC cells (30). This might
contribute to the lack of response to IFN treatment in MKN74 cells.
Subsequently, we examined whether an increase in membranous PD-L1
expression was accompanied by an upregulation of PD-L1 induced by
IFN-γ treatment. The results from flow cytometry demonstrated that
IFN-γ treatment induced an increase in membrane-bound PD-L1
expression in a dose-dependent manner. It is well known that
membranous PD-L1 expression is closely associated with
PD-1/PD-L1-mediated tumor immune escape (31). In addition, intracellular PD-L1, but
not membranous PD-L1, serves a crucial role in the proliferation
and migration of melanoma and ovarian cancer cells (30,32).
A recent study demonstrated that sPD-L1 is detected
not only in human serum but also in culture supernatants of
PD-L1-expressing cell lines (33).
The present study aimed therefore to detect sPD-L1 in the
supernatant of GC cell lines using ELISA. Although both
intracellular and membrane-bound PD-L1 was upregulated in three
cell lines treated with IFN-γ, sPD-L1 overproduction was only
observed in IFN-γ-treated OCUM-1 cells. Although the accurate
source of sPD-L1 remains unclear, sPD-L1 might be released or shed
from PD-L1-positive tumor cells. Considering that some of
disintegrin and metalloproteinase (ADAM) proteases, transmembrane
protein shedding enzymes, are overexpressed in undifferentiated GC
tissues (34), enhanced ADAM
activity might contribute to sPD-L1 production in OCUM-1 cells. It
is also possible that matrix metalloproteinases might partly be
associated with sPD-L1 release (35). Alternatively, sPD-L1 has been
reported to originate from its splicing variants lacking the
transmembrane domain (36). Further
investigation is required to elucidate the underlying mechanisms of
sPD-L1 production in GC cells.
sPD-L1 is used as a prognostic biomarker in various
types of cancer (37). It has been
reported that sPD-L1 functions as a lure and attenuates the effect
of ICI in lung cancer (38).
Furthermore, the exposure of CD4+ and CD8+
lymphocytes to sPD-L1 induces their apoptosis (39). Previous studies reported that a high
sPD-L1 level is closely associated with an unfavorable prognosis in
many types of cancer, including GC (28,40–43).
However, Zheng et al (44)
reported opposite results, where patients with GC and high sPD-L1
levels have a better prognosis than those with low sPD-L1 levels.
Concordant with these findings, the present study demonstrated that
sPD-L1 level in the serum of patients with stage IV GC was
significantly higher than in those with stages I–III GC. Further
investigation using a larger number of patients is required to
determine the role of sPD-L1 in GC.
In conclusion, the present study demonstrated that
IFN-γ treatment simultaneously enhanced the intracellular and
membranous PD-L1 expression in some GC cells. In addition, a
significantly high concentration of sPD-L1 was also detected in the
serum of patients with GC. Further investigation on the underlying
mechanism of the regulation of PD-L1 expression and sPD-L1
production is required and would serve the development of novel
therapeutic approaches in GC.
Acknowledgements
Not applicable.
Funding
This work was partly funded by the Japan Society for
the Promotion of Science (grant no. 16K09340) and the Program for
Basic and Clinical Research on Hepatitis from Japan Agency for
Medical Research and Development (AMED; grant no.
JP20fk0210054).
Availability of data and materials
All results and data obtained from the present study
are available from the corresponding author on reasonable
request.
Authors' contributions
YI and TC designed this study. TK, HK, KK, JA, RK,
YK and MN performed experiments and analyzed data. TS, RN, ES, SN,
RM, AT, TM, TN and JK checked all data and analyzed data. YI and HM
collected samples and data. AK and NK contributed to data
interpretation and writing of the manuscript. The manuscript was
written by YI and TC. All authors read and approved the final the
manuscript.
Ethics approval and consent to
participate
This study was approved by the Research Ethics
Committees of the Graduate School of Medicine, Chiba University
(approval no. 3552 and 3671). Informed consent was obtained from
all participants.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interest.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
Statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Khasag O, Boldbaatar G, Tegshee T, Duger
D, Dashdorj A, Uchida T, Matsuhisa T and Yamaoka Y: The prevalence
of helicobacter pylori infection and other risk factors among
mongolian dyspeptic patients who have a high incidence and
mortality rate of gastric cancer. Gut Pathog. 10:142018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Shiota S, Murakawi K, Suzuki R, Fujioka T
and Yamaoka Y: Helicobacter pylori infection in Japan. Expert Rev
Gastroenterol Hepatol. 7:35–40. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Selim JH, Shaheen S, Sheu WC and Hsueh CT:
Targeted and novel therapy in advanced gastric cancer. Exp Hematol
Oncol. 8:252019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Karimi P, Islami F, Anandasabapathy S,
Freedman ND and Kamangar F: Gastric cancer: Descriptive
epidemiology, risk factors, screening, and prevention. Cancer
Epidemiol Biomarkers Prev. 23:700–713. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wei SC, Duffy CR and Allison JP:
Fundamental mechanisms of immune checkpoint blockade therapy.
Cancer Discov. 8:1069–1086. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Schreiber RD, Old LJ and Smyth MJ: Cancer
immunoediting: Integrating immunity's roles in cancer suppression
and promotion. Science. 331:1565–1570. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Postow MA, Callahan MK and Wolchok JD:
Immune checkpoint blockade in cancer therapy. J Clin Oncol.
33:1974–1982. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kang YK, Boku N, Satoh T, Ryu MH, Chao Y,
Kato K, Chung HC, Chen JS, Muro K, Kang WK, et al: Nivolumab in
patients with advanced gastric or gastro-oesophageal junction
cancer refractory to, or intolerant of, at least two previous
chemotherapy regimens (ONO-4538-12, ATTRACTION-2): A randomised,
double-blind, placebo-controlled, phase 3 trial. Lancet.
390:2461–2471. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Keir ME, Francisco LM and Sharpe AH: PD-1
and its ligands in T-cell immunity. Curr Opin Immunol. 19:309–314.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Guan J, Lim KS, Mekhail T and Chang CC:
Programmed death ligand-1 (PD-L1) expression in the programmed
death receptor-1 (PD-1)/PD-L1 blockade: A key player against
various cancers. Arch Pathol Lab Med. 141:851–861. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nagato T, Ohkuri T, Ohara K, Hirata Y,
Kishibe K, Komabayashi Y, Ueda S, Takahara M, Kumai T, Ishibashi K,
et al: Programmed death-ligand 1 and its soluble form are highly
expressed in nasal natural killer/T-cell lymphoma: A potential
rationale for immunotherapy. Cancer Immunol Immunother. 66:877–890.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chang B, Huang T, Wei H, Shen L, Zhu D, He
W, Chen Q, Zhang H, Li Y, Huang R, et al: The correlation and
prognostic value of serum levels of soluble programmed death
protein 1 (sPD-1) and soluble programmed death-ligand 1 (sPD-L1) in
patients with hepatocellular carcinoma. Cancer Immunol Immunother.
68:353–363. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Deng R, Zhang P, Liu W, Zeng X, Ma X, Shi
L, Wang T, Yin Y, Chang W, Zhang P, et al: HDAC is indispensable
for IFN-γ-induced B7-H1 expression in gastric cancer. Clin
Epigenetics. 10:1532018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Union for International Cancer Control, .
Briely JD, Gospodarowicz MK and Wittekind CH: TNM Classification of
Malignant Tumors. 8th. Wiley-Blackwell; Hoboken, NJ: 2017
|
|
17
|
Intlekofer AM and Thompson CB: At the
bench: Preclinical rationale for CTLA-4 and PD-1 blockade as cancer
immunotherapy. J Leukoc Biol. 94:25–39. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ishida Y, Agata Y, Shibahara K and Honjo
T: Induced expression of PD-1, a novel member of the immunoglobulin
gene superfamily, upon programmed cell death. EMBO J. 11:3887–3895.
1992. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ma J, Zheng B, Goswami S, Meng L, Zhang D,
Cao C, Li T, Zhu F, Ma L, Zhang Z, et al: PD1Hi
CD8+ T cells correlate with exhausted signature and poor
clinical outcome in hepatocellular carcinoma. J Immunother Cancer.
7:3312019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ishida M, Iwai Y, Tanaka Y, Okazaki T,
Freeman GJ, Minato N and Honjo T: Differential expression of PD-L1
and PD-L2, ligands for an inhibitory receptor PD-1, in the cells of
lymphohematopoietic tissues. Immunol Lett. 84:57–62. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Francisco LM, Sage PT and Sharpe AH: The
PD-1 pathway in tolerance and autoimmunity. Immunol Rev.
236:219–242. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Teng MW, Ngiow SF, Ribas A and Smyth MJ:
Classifying cancers based on t-cell infiltration and PD-L1. Cancer
Res. 75:2139–2145. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Noman MZ, Desantis G, Janji B, Hasmim M,
Karray S, Dessen P, Bronte V and Chouaib S: PD-L1 is a novel direct
target of HIF-1α, and its blockade under hypoxia enhanced
MDSC-mediated T cell activation. J Exp Med. 211:781–790. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Darvin P, Toor SM, Sasidharan Nair V and
Elkord E: Immune checkpoint inhibitors: Recent progress and
potential biomarkers. Exp Mol Med. 50:1–11. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wu C, Zhu Y, Jiang J, Zhao J, Zhang XG and
Xu N: Immunohistochemical localization of programmed death-1
ligand-1 (PD-L1) in gastric carcinoma and its clinical
significance. Acta Histochem. 108:19–24. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sun C, Mezzadra R and Schumacher TN:
Regulation and function of the PD-L1 checkpoint. Immunity.
48:434–452. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cha JH, Chan LC, Li CW, Hsu JL and Hung
MC: Mechanisms controlling PD-L1 expression in cancer. Mol Cell.
76:359–370. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shigemori T, Toiyama Y, Okugawa Y,
Yamamoto A, Yin C, Narumi A, Ichikawa T, Ide S, Shimura T, Fujikawa
H, et al: Soluble PD-L1 expression in circulation as a predictive
marker for recurrence and prognosis in gastric cancer: Direct
comparison of the clinical burden between tissue and serum PD-L1
expression. Ann Surg Oncol. 26:876–883. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jiang X, Wang J, Deng X, Xiong F, Ge J,
Xiang B, Wu X, Ma J, Zhou M, Li X, et al: Role of the tumor
microenvironment in PD-L1/PD-1-mediated tumor immune escape. Mol
Cancer. 18:102019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mimura K, Teh JL, Okayama H, Shiraishi K,
Kua LF, Koh V, Smoot DT, Ashktorab H, Oike T, Suzuki Y, et al:
PD-L1 expression is mainly regulated by interferon gamma associated
with JAK-STAT pathway in gastric cancer. Cancer Sci. 109:43–53.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen J, Feng Y, Lu L, Wang H, Dai L, Li Y
and Zhang P: Interferon-gamma-induced PD-L1 surface expression on
human oral squamous carcinoma via PKD2 signal pathway.
Immunobiology. 217:385–393. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Clark CA, Gupta HB and Curiel TJ: Tumor
cell-intrinsic CD274/PD-L1: A novel metabolic balancing act with
clinical potential. Autophagy. 13:987–988. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Qu QX, Xie F, Huang Q and Zhang XG:
Membranous and cytoplasmic expression of PD-L1 in ovarian cancer
cells. Cell Physiol Biochem. 43:1893–1906. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Aydin D, Bilici A, Yavuzer D, Kefeli U,
Tan A, Ercelep O, Mert A, Yuksel S, Ozcelik M, Isik D, et al:
Prognostic significance of ADAM17 expression in patients with
gastric cancer who underwent curative gastrectomy. Clin Transl
Oncol. 17:604–611. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chen Y, Wang Q, Shi B, Xu P, Hu Z, Bai L
and Zhang X: Development of a sandwich ELISA for evaluating soluble
PD-L1 (CD274) in human sera of different ages as well as
supernatants of PD-L1+ cell lines. Cytokine. 56:231–238. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhou J, Mahoney KM, Giobbie-Hurder A, Zhao
F, Lee S, Liao X, Rodig S, Li J, Wu X, Butterfield LH, et al:
Soluble PD-L1 as a biomarker in malignant melanoma treated with
checkpoint blockade. Cancer Immunol Res. 5:480–492. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhu X and Lang J: Soluble PD-1 and PD-L1:
Predictive and prognostic significance in cancer. Oncotarget.
8:97671–97682. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Gong B, Kiyotani K, Sakata S, Nagano S,
Kumehara S, Baba S, Besse B, Yanagitani N, Friboulet L, Nishio M,
et al: Secreted PD-L1 variants mediate resistance to PD-L1 blockade
therapy in non-small cell lung cancer. J Exp Med. 216:982–1000.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Frigola X, Inman BA, Krco CJ, Liu X,
Harrington SM, Bulur PA, Dietz AB, Dong H and Kwon ED: Soluble
B7-H1: Differences in production between dendritic cells and T
cells. Immunol Lett. 142:78–82. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Frigola X, Inman BA, Lohse CM, Krco CJ,
Cheville JC, Thompson RH, Leibovich B, Blute ML, Dong H and Kwon
ED: Identification of a soluble form of B7-H1 that retains
immunosuppressive activity and is associated with aggressive renal
cell carcinoma. Clin Cancer Res. 17:1915–1923. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Rossille D, Azzaoui I, Feldman AL, Maurer
MJ, Labouré G, Parrens M, Pangault C, Habermann TM, Ansell SM, Link
BK, et al: Soluble programmed death-ligand 1 as a prognostic
biomarker for overall survival in patients with diffuse large
B-cell lymphoma: A replication study and combined analysis of 508
patients. Leukemia. 31:988–991. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Finkelmeier F, Canli Ö, Tal A, Pleli T,
Trojan J, Schmidt M, Kronenberger B, Zeuzem S, Piiper A, Greten FR
and Waidmann O: High levels of the soluble programmed death-ligand
(sPD-L1) identify hepatocellular carcinoma patients with a poor
prognosis. Eur J Cancer. 59:152–159. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ito M, Oshima Y, Yajima S, Suzuki T,
Nanami T, Shiratori F, Funahashi K, Nemoto T and Shimada H: Is high
serum programmed death ligand 1 level a risk factor for poor
survival in patients with gastric cancer? Ann Gastroenterol Surg.
2:313–318. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zheng Z, Bu Z, Liu X, Zhang L, Li Z, Wu A,
Wu X, Cheng X, Xing X, Du H, et al: Level of circulating PD-L1
expression in patients with advanced gastric cancer and its
clinical implications. Chin J Cancer Res. 26:104–111.
2014.PubMed/NCBI
|