Introduction
MicroRNAs (miRNAs/miRs) are endogenous non-coding
small RNAs that are 19–24 nucleotides in length and are widely
found in eukaryotes (1). In general,
they bind to complementary sites on the 3′-untranslated regions of
mRNAs, which are consequently destabilized or translationally
suppressed (1). This
post-transcriptional regulation serves essential roles
physiologically and pathologically. miRNAs regulate important
biological processes, including proliferation and apoptosis
(2,3). Numerous miRNAs are involved in the
tumorigenic transformation or migration of cells (4). Recent advancements in molecular biology
technologies have introduced novel approaches to study the role of
miRNAs in cancer and have thereby led to novel findings regarding
cancer pathogenesis, prognosis and treatment (5,6).
Cancer is one of the leading causes of mortality and
morbidity globally. According to the latest data published in the
GLOBOCAN database in 2018, 18.1 million cancer cases and 9.6
million cancer-associated deaths were reported worldwide (7). Asia has the highest number of patients
with cancer in the world, which accounts for 48.4% of the global
cancer population (7). The five most
frequent types of cancer worldwide in descending order are lung
cancer, breast carcinoma, colorectal cancer, prostate carcinoma and
gastric cancer (7). The five types
of cancer with the highest mortality rates worldwide are lung
cancer, colorectal carcinoma, gastric cancer, liver carcinoma and
breast cancer (7). The types of
cancer with the highest cancer-associated morbidity and mortality
in men and women worldwide are lung and breast cancer, respectively
(7). It has been estimated that
there will be 22 million new cancer cases and 13 million cancer
deaths per year by 2030 (8).
Therefore, the identification of effective approaches for the
prevention, diagnosis and treatment of cancer is required, in
addition to the traditional approaches of surgery, chemotherapy and
radiotherapy.
Abnormal miRNA expression is associated with
tumorigenesis, and more than half of all miRNAs in the human genome
are located in cancer-associated regions or fragile sites (9). In 2006, Calin et al (10) reported that the miRNA cluster
miR-15/miR-16-1 was eliminated or repressed in 69% of patients with
chronic lymphocytic leukemia. The aforementioned study was the
first to demonstrated that miRNAs are closely associated with
cancer (10). Abnormally expressed
miRNAs in cancer can be functionally classified as oncogenic, such
as miR-17-92 (11), or tumor
suppressors, such as let-7 (12),
which are upregulated and downregulated in tumor cells,
respectively. Accordingly, the upregulation of the oncogenic miRNAs
and the downregulation of the tumor suppressor miRNAs may promote
tumor development. However, a miRNA may serve different roles in
different types of cancer. For example, miR-221 acts as an oncogene
in hepatocellular carcinoma (13),
but it is a tumor suppressor in erythroblastic leukemia (14). In addition to serving as biomarkers
for tumorigenesis, miRNAs are associated with tumor
chemoresistance. Upregulation of some miRNAs, such as miR-19
(15) and miR-21 (16), can enhance tumor chemoresistance,
whereas other miRNAs, such as miR-130a (17) and miR-298 (18), are downregulated in chemoresistant
tumor cells. Therefore, it may be possible to use miRNA expression
profiles of tumors for diagnostic, prognostic and therapeutic
purposes. However, this requires an in-depth analysis of
miRNA-associated mechanisms in various types of tumor.
miR-33a is located in the 16th intron of the human
sterol regulatory element binding transcription factor 2 (SREBF-2)
gene on chromosome 22 (19). This
highly conserved intronic miRNA regulates the genes that control
cholesterol uptake or synthesis (20). SREBFs activate the expression of
>30 genes involved in the synthesis or cellular uptake of
cholesterol, triglycerides, phospholipids and fatty acids, along
with the NADPH cofactors required to synthesize these biomolecules
(21). In addition, the SREBF
signaling pathway regulates various cellular processes, including
phagocytosis and cell cycle progression (22). In healthy tissues, transcriptional
activation of SREBF-2 upregulates miR-33a, consequently enhancing
cellular lipid metabolism, including that of cholesterol (22). Studies have demonstrated that
cholesterol is involved in apoptosis, whereas the SREBP/miR-33 axis
participates in cell growth and cell cycle progression (23,24).
Therefore, dysregulation of miR-33a levels may contribute to
tumorigenesis by affecting cholesterol levels. In this case, tumors
may be treated by correcting the miR-33a levels or associated
mechanisms.
The present review describes the available
literature regarding miR-33a and cancer and provides an overview of
miR-33a-associated mechanisms in hepatocellular, gastric,
colorectal, pancreatic and gallbladder carcinomas, as well as
osteosarcoma, melanoma, glioma, esophageal and tongue squamous cell
cancer, and lung, laryngeal, renal, thyroid, prostate and breast
cancer. Additionally, 38 target genes of miR-33a and the
possibility of their involvement in cancer are examined, providing
a basis for future research investigating the role of miR-33a in
cancer.
miR-33a and its role in cancer
Digestive system cancers
Hepatocellular carcinoma
Hepatocellular carcinoma is the third most common
type of cancer globally (25–27).
Hepatitis B and C virus infections are considered to be the main
causes of hepatocellular carcinoma, which can also be caused by
non-alcoholic liver disease or alcohol abuse (28).
In this type of cancer, miR-33a can act as a tumor
suppressor gene. Liu et al (29) have demonstrated that miR-33a-5p is
downregulated, while its target gene paraneoplastic antigen Ma1
(PNMA1) is upregulated in hepatoma cells. Consequently, the
downstream proteins of PNMA1, such as C-MYC, β-catenin and cyclin
D1, are upregulated; these proteins activate the Wnt signaling
pathway, which facilitates the proliferation of hepatoma cells
(29). Additionally, PNMA1
upregulates N-cadherin, fibronectin and vimentin expression, but
downregulates E-cadherin and claudin-1 expression, thereby
promoting the epithelial-mesenchymal transition (EMT) of the cells,
while also promoting their migration through the upregulation of
matrix metalloproteinases-2/9 (Fig.
1A) (29). Similarly, Guo et
al (30) and Li et al
(31) have demonstrated that
hypoxia-inducible factors (HIFs) and long non-coding RNA (lncRNA)
cancer susceptibility 15 (CASC15) decrease the miR-33a level in
hepatoma cells. Additionally, the miR-33a target gene Twist basic
helix-loop-helix transcription factor 1 (TWIST1) is consequently
upregulated, which subsequently downregulates E-cadherin and
upregulates N-cadherin and vimentin expression, thereby further
promoting EMT and invasion of hepatoma cells (Fig. 1B) (30,31).
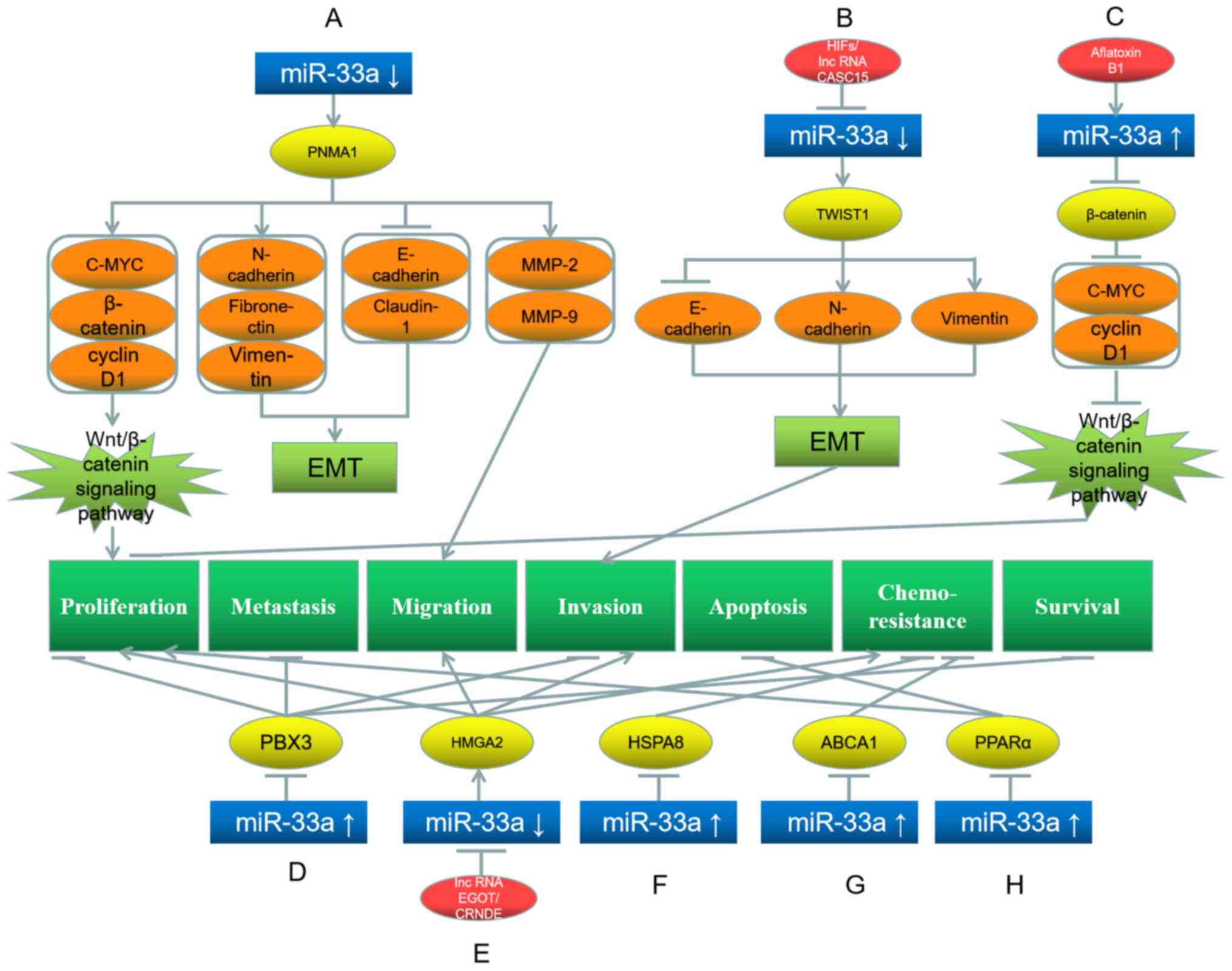 | Figure 1.Roles of miR-33a and its target genes
in hepatocellular carcinoma, and the underlying mechanisms. (A)
miR-33a downregulation upregulates its target gene PNMA1, which
further upregulates C-MYC, β-catenin, N-cadherin, fibronectin,
vimentin, MMP-2 and MMP-9, while downregulates E-cadherin and
claudin-1, promoting EMT, proliferation and migration. (B) HIFs and
lncRNA CASC15 downregulate miR-33a and further upregulate the
target gene TWIST1, which upregulates N-cadherin and vimentin,
while downregulates E-cadherin, promoting EMT and invasion. (C)
Aflatoxin B1 upregulates miR-33a, which inhibits its target gene
β-catenin and further downstream proteins C-MYC and cyclin D1,
leading to proliferation inhibition. (D) miR-33a upregulation
downregulates its target gene PBX3, leading to the inhibition of
proliferation, metastasis, invasion and survival. (E) LncRNA EGOT
and lncRNA CRNDE downregulate miR-33a, which further upregulates
its target gene HMGA2, promoting proliferation, migration, invasion
and chemoresistance. (F) miR-33a upregulation downregulates its
target gene HSPA8, inhibiting chemoresistance. (G) miR-33a
upregulation downregulates its target gene ABCA1, inhibiting
chemoresistance. (H) miR-33a upregulation downregulates its target
gene PPARα, promoting proliferation and inhibiting apoptosis. Red
represents the factor causing miR-33a dysregulation; blue
represents miR-33a; yellow represents the target genes of miR-33a;
orange represents the downstream proteins; light green represents
the signaling pathways and EMT processes; dark green represents the
biological processes of cancer cells. miR, microRNA; PNMA1,
paraneoplastic antigen Ma1; MMP, matrix metalloproteinase; TWIST,
Twist basic helix-loop-helix transcription factor 1; EMT,
epithelial-mesenchymal transition; PBX3, pre-B-cell leukemia
transcription factor 3; HSPA8, heat shock protein family A (Hsp70)
member 8; ABCA1, ATP-binding cassette transporter A1; PPARα,
peroxisome proliferator activated receptor α. |
By contrast, Fang et al (32) have revealed that aflatoxin B1
upregulates miR-33a in the hepatoma Chang (IC50, 40
µg/ml) and HepG2 (IC50, 77 µg/ml) cell lines.
Consequently, β-catenin and its downstream proteins C-MYC and
cyclin D1 are downregulated, whereby the Wnt signaling pathway is
suppressed, and so is the proliferation of hepatoma cells (Fig. 1C) (32). Similarly, Han et al (33) have observed that miR-33a-3p is
downregulated in hepatoma cells, and its forced expression
suppresses the proliferation, metastasis and invasion of these
cells by downregulating the target gene pre-B-cell leukemia
transcription factor 3 (Fig. 1D).
Furthermore, low miR-33a expression is associated with a low
survival rate in patients with hepatocellular carcinoma (33). Similarly, Wu et al (34) and Han et al (35) have illustrated that lncRNA eosinophil
granule ontogeny transcript and lncRNA colorectal neoplasia
differentially expressed in hepatoma cells inhibit miR-33a
expression, which upregulates the target gene high mobility group
AT-hook 2 (HMGA2), thereby promoting the proliferation, migration,
invasion and chemoresistance of hepatoma cells (Fig. 1E).
Furthermore, miR-33a can affect the chemoresistance
of hepatoma cells. Meng et al (36) have demonstrated that hepatoma cells
with high miR-33a-5p levels are more sensitive to cisplatin than
those with low expression levels, and this cisplatin resistance can
be decreased by downregulating heat shock protein family A (Hsp70)
member 8 through miR-33a-5p expression (Fig. 1F). Similarly, Hou et al
(37) have reported that miR-33a
overexpression in Lgr5+ hepatoma-cancer stem cells decreases
ATP-binding cassette transporter A1 (ABCA1) expression, thereby
reducing the resistance of these cells to doxorubicin (Fig. 1G). Accordingly, the aforementioned
study indicated that increasing the levels of miR-33a in hepatoma
cells may reduce their resistance to doxorubicin (37).
Although most studies have illustrated that miR-33a
serves as a tumor suppressor in hepatoma cells, Chang et al
(38) have observed the opposite
effect, reporting that miR-33a upregulation and the consequent
downregulation of its target peroxisome proliferator activated
receptor α promote proliferation, while inhibiting the apoptosis of
hepatoma cells (Fig. 1H). The reason
underlying these opposing observations is unclear.
Gastric cancer
Gastric cancer is one of the most common digestive
system cancers and causes major public health problems worldwide
(39). Dietary factors are the main
cause of this type of cancer, although genetic or environmental
factors, such as stress, are also closely associated with it
(40).
miR-33a serves a role in suppressing oncogenes in
gastric cancer. Wang et al (41), have illustrated that miR-33a
expression is decreased in gastric tumors. Increased miR-33a
downregulates Pim-1 proto-oncogene (PIM1), cyclin D1 and
cyclin-dependent kinase (CDK6) to impede the proliferation of
gastric cancer cells, causing them to be arrested at the
G1 stage (Fig. 2A)
(41). Among these three target
genes, the interaction of miR-33a with PIM1 has the greatest impact
on gastric cancer (41).
Additionally, Chen et al (42) have demonstrated that miR-33a is
downregulated in gastric tumor cells. miR-33a overexpression
inhibits Snail family transcriptional repressor 2 (SNAI2), thereby
upregulating the downstream protein E-cadherin and downregulating
N-cadherin, vimentin and β-catenin, which suppresses EMT in gastric
tumor cells (42). In addition,
miR-33a downregulates Snail and Slug levels, and inhibition of the
Snail/Slug signaling pathway hinders migration and invasion of
these cells (Fig. 2B) (42). Wu et al (43) have revealed that lncRNA CASC15 is a
miRNA sponge in gastric cancer cells, as in hepatoma cells. CASC15
reduces miR-33a-5p expression, thereby upregulating the miR-33a
target gene zinc finger E-Box binding homeobox 1 (ZEB1), which
subsequently upregulates N-cadherin and downregulates E-cadherin,
promoting the EMT and migration of gastric cancer cells (Fig. 2C) (43).
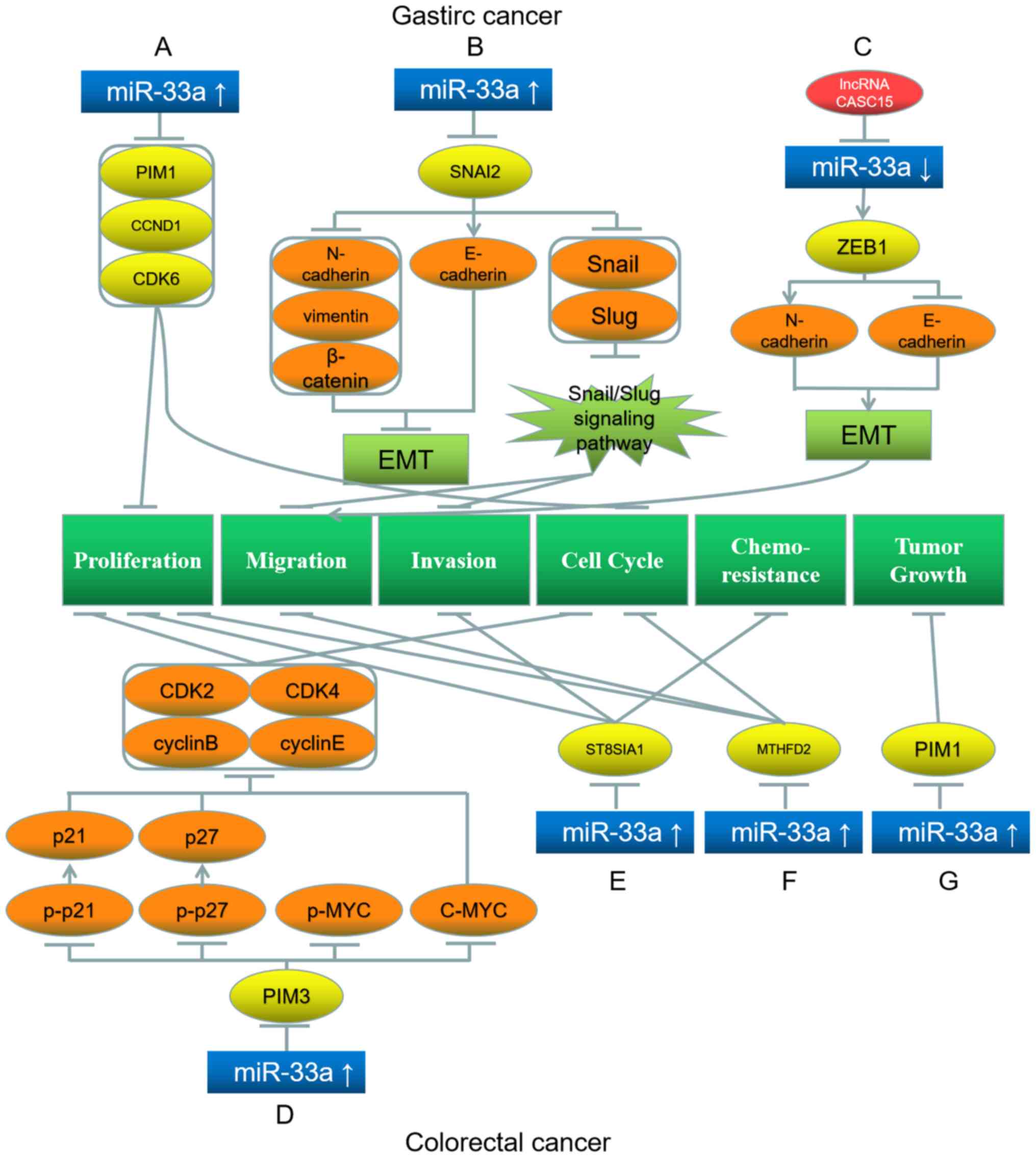 | Figure 2.Roles of miR-33a and its target genes
in gastric (top) and colorectal (bottom) cancer, and the underlying
mechanisms. (A) miR-33a upregulation downregulates its target genes
PIM1, CCND1 and CDK6, which inhibit proliferation and cell cycle.
(B) miR-33a upregulation inhibits its target gene SNAI2, which
further downregulates N-cadherin, vimentin, β-catenin, Snail and
Slug, while upregulates E-cadherin, leading to the inhibition of
EMT, migration and invasion. (C) LncRNA CASC15 inhibits miR-33a and
further upregulates its target gene ZEB1, which upregulates
N-cadherin and downregulates E-cadherin, promoting EMT and
proliferation. (D) miR-33a upregulation inhibits its target gene
PIM3, which further downregulates the downstream proteins p-p21
(upregulating p21), p-p27 (upregulating p27), p-MYC, C-MYC, CDK2,
CDK4, cyclin B and cyclin E, leading to proliferation and cell
cycle inhibition. (E) miR-33a upregulation downregulates its target
gene ST8SIA1, inhibiting proliferation, invasion and
chemoresistance. (F) miR-33a upregulation downregulates its target
gene MTHFD2, inhibiting proliferation, migration and cell cycle.
(G) miR-33a upregulation downregulates its target gene PIM1,
inhibiting tumor growth. Red represents the factor causing miR-33a
dysregulation; blue represents miR-33a; yellow represents the
target genes of miR-33a; orange represents the downstream proteins;
light green represents the signaling pathways and EMT processes;
dark green represents the biological processes of cancer cells.
miR, microRNA; EMT, epithelial-mesenchymal transition; p,
phosphorylated; PIM1/3, Pim-1/3 proto-oncogene; CCND1, cyclin D1;
CDK, cyclin-dependent kinase; SNAI2, Snail family transcriptional
repressor 2; ZEB1, Zinc finger E-Box binding homeobox 1; ST8SIA1,
ST8α-N-acetyl-neuraminide-α-2,8-sialyltransferase 1; MTHFD2,
methylenetetrahydrofolate dehydrogenase 2. |
Colorectal cancer
Colorectal cancer is the third most common
malignancy and the fourth most common principal cause of tumor
mortality in the world (44).
According to a cancer research carried out in 2019, a considerable
proportion (21%) of colorectal tumor cases is detected at the
advanced stage (45). The occurrence
of colorectal cancer is strongly associated with a high fat-diet,
especially with a cholesterol-rich diet (46–49).
miR-33a activates the transcription of genes
associated with cholesterol metabolism. Accordingly, it serves as a
tumor suppressor gene in colorectal carcinoma. Wang et al
(50) have reported that miR-33a can
neutralize the effect of excessive cholesterol intake on the
proliferation of colorectal cancer cells. While cholesterol
downregulates miR-33a and consequently upregulates the miR-33a
target PIM3, downregulation of PIM3 by miR-33a inhibits the
phosphorylation of p21, p27 and p-MYC, and C-MYC expression, which
deactivate the cell cycle-associated proteins CDK2, CDK4, cyclin B
and cyclin E (50). Consequently,
the proliferation and cell cycle progression of colorectal cancer
cells are inhibited (Fig. 2D)
(50). Shan et al (51) have demonstrated that miR-33a and
let-7e are downregulated in colorectal cancer cells. These two
miRNAs can bind to ST8α-N-acetyl-neuraminide-α-2,8-
sialyltransferase 1 (ST8SIA1) and consequently further inhibit the
proliferation, invasion and chemoresistance of colorectal cancer
cells (Fig. 2E) (51). ST8SIA1 overexpression suppresses
these inhibitory effects of miR-33a and let-7e (51). Therefore, these two miRNAs and
ST8SIA1 affect colorectal cancer cells through mutual restrictions
(51). Similarly, Yan et al
(52) have illustrated that
miR-33a-5p is downregulated, while its target gene
methylenetetrahydrofolate dehydrogenase 2 is upregulated in
colorectal carcinoma (52).
Additionally, the proliferation, migration and cell cycle
progression of colorectal carcinoma cells can be repressed by
overexpressed miR-33a-5p (Fig. 2F)
(52).
Notably, Ibrahim et al (53) have demonstrated that miRNAs and
polyethyleneimine (PEI) can form biocompatible complexes that can
be used in miRNA replacement therapies. Delivery of the miR-33a/PEI
complex into colorectal cancer cells suppresses tumor growth via
the downregulation of PIM1 (Fig. 2G)
(53). This observation indicates
the efficacy of such a biocomplex, which may be used as a novel
treatment in colorectal cancer (53).
Pancreatic cancer
Pancreatic ductal adenocarcinoma (PDAC), also known
as pancreatic cancer, is one of the most lethal malignancies in the
world (54). In 2018, PDAC exhibited
the lowest 5-year relative survival rate compared with all other
solid tumors, and caused 432,242 deaths worldwide (55). Early-stage PDAC is asymptomatic;
therefore, the manifestation of the illness is delayed (54). Consequently, ~50% patients are
already at the metastatic stage at initial diagnosis (54).
miR-33a is a tumor suppressor in PDAC. Liang et
al (56) have revealed that
increasing the expression of downregulated miR-33a in pancreatic
cancer inhibits PIM3 expression and downregulates the downstream
proteins phosphorylated (p)-AKT, p-glycogen synthase kinase
(GSK)-3β and β-catenin. Consequently, the AKT/GSK-3β/β-catenin
signaling pathway is repressed, and the proliferation and
gemcitabine-resistance of pancreatic cancer cells are inhibited
(Fig. 3A) (56). In another study, Liang et al
(57) demonstrated that high levels
of miR-33a suppress the nuclear translocation of β-catenin, thereby
repressing the expression levels of downstream proteins, such as
slug, vimentin, N-cadherin, survivin, cyclin D1 and multidrug
resistance 1. Consequently, the gemcitabine-sensitivity of
pancreatic cancer cells is increased, and the proliferation and EMT
of pancreatic tumor cells are suppressed (Fig. 3B) (57).
 | Figure 3.Roles of miR-33a and its target genes
in pancreatic cancer (top), and gallbladder (bottom left) and
esophageal squamous (bottom middle and right) carcinoma, and the
underlying mechanisms. (A) miR-33a upregulation inhibits its target
gene PIM3; this further downregulates p-AKT, p-GSK-3β and
β-catenin, leading to proliferation inhibition and chemosensitivity
promotion. (B) miR-33a upregulation inhibits its target gene
β-catenin, further downregulating slug, vimentin, N-cadherin,
survivin, cyclin D1 and MDR-1, leading to EMT inhibition and
chemosensitivity promotion. (C) miR-33a upregulation inhibits its
target gene TWIST1, further downregulating N-cadherin and
upregulating E-cadherin, leading to inhibition of EMT,
proliferation, migration and invasion. (D) miR-33a upregulation
downregulates its target gene ZEB1, inhibiting proliferation and
migration. (E) Linc-ROR downregulates miR-33a, further upregulating
its target gene SOX9, which promotes proliferation. Red represents
the factor causing miR-33a dysregulation; blue represents miR-33a;
yellow represents the target genes of miR-33a; orange represents
the downstream proteins; light green represents the signaling
pathways and EMT processes; dark green represents the biological
processes of cancer cells. miR, microRNA; EMT,
epithelial-mesenchymal transition; p-, phosphorylated; PIM3, Pim-3
proto-oncogene; GSK, glycogen synthase kinase; MDR-1, multidrug
resistance 1; SOX9, SRY-box 9; TWIST1, Twist basic helix-loop-helix
transcription factor 1; ZEB1, Zinc finger E-Box binding homeobox
1. |
Gallbladder cancer
Gallbladder cancer is a rare malignancy. The
incidence of gallbladder cancer is higher in developing countries
compared with developed countries (58).
miR-33a represses carcinogenesis in gallbladder
cancer. Zhang et al (59)
have demonstrated that upregulating miR-33a in IL-6-induced
gallbladder carcinoma cells downregulates the downstream protein
N-cadherin and upregulates E-cadherin via the downregulation of the
miR-33a target TWIST1. Consequently, the EMT, proliferation,
migration and invasion of gallbladder tumor cells are inhibited
(Fig. 3C) (59).
Esophageal squamous cell carcinoma (ESCC)
ESCC accounts for ~90% of esophageal cancer cases
(60). The causes of ESCC are
associated with economic status and unhealthy habits, such as
smoking and alcohol abuse (61).
miR-33a acts as a tumor suppressor in ESCC. It
targets and downregulates ZEB1 and thereby suppresses ESCC
proliferation and migration (Fig.
3D) (62). Notably, lncRNA
differentiation antagonizing non-protein coding RNA (DANCR)
downregulates miR-33a-5p levels through miRNA sponging in ESCC
(62). Wang et al (63) have detected the regulatory network
linc-ROR-miRNA-SRY-box 9 (SOX9) in ESCC. Linc-ROR inhibits the
expression of multiple miRNAs, including miR-33a, and consequently
upregulates their target gene SOX9, which confers stem cell-like
properties and promotes cell proliferation (Fig. 3E) (63).
Respiratory system cancers
As one of the most common types of cancer in the
world, lung cancer causes the majority of tumor-associated deaths;
in 2018, 2.1 million patients with lung cancer were diagnosed, and
lung cancer caused 1.8 million deaths worldwide (7). In China, the main causes of lung
carcinoma are air pollution and smoking (64). There are two types of lung cancer,
small cell lung carcinoma and non-small cell lung carcinoma
(NSCLC), and the latter can be further divided into three subtypes:
Squamous cell cancer, adenocarcinoma and large-cell carcinoma. Most
cases of lung cancer (80-85%) are NSCLC (64).
NSCLC
Du et al (65)
have indicated that miR-33a is downregulated in NSCLC cells, and
that the proliferation of NSCLC cells is subdued by upregulating
miR-33a, which downregulates its target gene methyltransferase-like
3 and the subsequent downstream proteins epidermal growth factor
receptor, tafazzin, mitogen-activated protein kinase-activated
protein kinase 2 and DNA methyltransferase 3A (Fig. 4A). Additionally, Kang et al
(66) have observed that miR-33a is
downregulated in NSCLC cells, and its upregulation represses the
proliferation, migration and cell cycle progression of these cells
by downregulating its target gene Cullin associated and neddylation
dissociated 1 and subsequent downstream proteins cyclin D1, c-Jun
and S-phase kinase associated protein 2 (Fig. 4B). Amaar and Reeves (67) have demonstrated that the oncogene Ras
association domain family member 1 (RAFFS1C) downregulates
miR-33a-5p, whereby the miR-33a-5p target ABCA1 is upregulated,
enhancing the chemoresistance of NSCLC cells. Additionally, the
aforementioned study has indicated that RAFFS1C promotes the EMT,
proliferation and migration of NSCLC cells by downregulating
miR-33a-5p and upregulating β-catenin, SNAIL and vimentin (Fig. 4C) (67). Furthermore, Yang et al
(68) have observed that miR-33a is
downregulated in NSCLC. miR-33a targets TWIST1, whereby the
downstream proteins vimentin and cadherin-1 are downregulated, and
the EMT and migration of NSCLC cells are suppressed (Fig. 4D) (68). Kang et al (69) have demonstrated that the oncogene
transcription factor AP-2 γ is overexpressed in NSCLC cells and
downregulates miR-33a. Consequently, the miR-33a target CDK6 is
upregulated, whereby the cell cycle progression and tumor
development of NSCLC cells are enhanced (Fig. 4E) (69). Wang et al (70) have revealed that overexpression of
circular-CCDC66 downregulates miR-33a-5p in NSCLC cells,
consequently upregulating the miR-33a-5p target karyopherin subunit
α4 and promoting tumorigenesis and development of NSCLC (Fig. 4F).
 | Figure 4.Roles of miR-33a and its target genes
in non-small cell lung cancer (top) and untyped lung cancer
(bottom), and the underlying mechanisms. (A) miR-33a upregulation
inhibits its target gene METTL3, further downregulating EGFR, TAZ,
MAPKAPK2 and DNMT3A, leading to proliferation inhibition. (B)
miR-33a upregulation inhibits its target gene CAND1, further
downregulating cyclin D1, c-Jun and Skp2, leading to inhibition of
proliferation, migration and cell cycle. (C) RAFFS1C inhibits
miR-33a and further upregulates its target gene ABCA1, as well as
β-catenin, SNAIL and vimentin, leading to promotion of EMT,
proliferation, migration and chemoresistance. (D) miR-33a
upregulation inhibits its target gene TWIST1, further
downregulating vimentin and CDH1, leading to inhibition of EMT and
migration. (E) TFAP2C inhibits miR-33a and further upregulates its
target gene CDK6, which promotes cell cycle progression and tumor
development. (F) circular-CCDC66 inhibits miR-33a and further
upregulates its target gene KPNA4, which promotes tumor
development. (G) miR-33a upregulation inhibits its target gene
mTOR, downregulating p-p70S6K and p-4EBP1, leading to inhibition of
proliferation and chemoresistance. (H) miR-33a upregulation
inhibits its target gene PTHrP, downregulating IL-8 and leading to
metastasis inhibition. (I) lncRNA JPX inhibits miR-33a and further
upregulates its target gene TWIST1, leading to promotion of
proliferation and metastasis. (J) TTF-1 upregulates miR-33a and
further downregulates its target gene HMGA2, which inhibits
migration. (K) miR-33a upregulation downregulates its target gene
β-catenin, which inhibits proliferation and cell cycle. (L) miR-33a
upregulation downregulates its target gene PD-1, resulting in a
better prognosis. Red represents the factor causing miR-33a
dysregulation; blue represents miR-33a; yellow represents the
target genes of miR-33a; orange represents the downstream proteins;
light green represents the signaling pathways and EMT processes;
dark green represents the biological processes of cancer cells.
miR, microRNA; EMT, epithelial-mesenchymal transition; p-,
phosphorylated; DNMT3A, DNA methyltransferase 3A; MAPKAPK2,
mitogen-activated protein kinase-activated protein kinase 2; PTHrP,
parathyroid hormone-related protein; TAZ, tafazzin; Skp2, S-phase
kinase associated protein 2; METTL3, methyltransferase-like 3;
EGFR, epidermal growth factor receptor; CAND1, Cullin associated
and neddylation dissociated 1; ABCA1, ATP-binding cassette
transporter A1; TWIST1, Twist basic helix-loop-helix transcription
factor 1; CDH1, cadherin 1; CDK6, cyclin-dependent kinase 6; KPNA4,
karyopherin subunit α4; IL-8, interleukin-8; HMGA2, high mobility
group AT-hook 2; PD-1, programmed cell death protein 1. |
Untyped lung cancer
Li et al (71)
have confirmed that miR-33a-5p is downregulated in lung tumor
cells. By acting on its target gene mTOR and downstream proteins
p-p70S6K and p-eukaryotic translation initiation factor 4E binding
protein 1 (4EBP1), miR-33a-5p inhibits the mTOR signaling pathway,
inhibits proliferation and reduces celastrol resistance in lung
cancer cells (Fig. 4G) (71). Additionally, Kuo et al
(72) have demonstrated that miR-33a
is downregulated in lung cancer and that its overexpression
represses bone metastasis by downregulating parathyroid
hormone-related protein and its downstream protein interleukin
(IL)-8 (Fig. 4H). Pan et al
(73) have illustrated that lncRNA
JPX transcript, XIST activator downregulates miR-33a-5p in lung
cancer cells through miRNA sponging. This downregulation
upregulates the target gene TWIST1 and activates the Wnt/β-catenin
signaling pathway, subsequently promoting lung cancer cell
proliferation and metastasis (Fig.
4I) (73). Rice et al
(74) have observed that thyroid
transcription factor-1 binds to the promoter of SREBF2, which is
the host gene of miR-33a, and activates miR-33a transcription.
miR-33a subsequently downregulates its target, HMGA2, whereby the
migration of lung cancer cells is suppressed (Fig. 4J) (74). Additionally, Zhu et al
(75) have demonstrated that miR-33a
expression is decreased in lung cancer cells, while increasing
miR-33a expression downregulates β-catenin and causes
G1/S phase arrest, thereby suppressing cancer cell
proliferation (Fig. 4K). Boldrini
et al (76) have indicated
that patients with lung cancer with high miR-33a expression and low
programmed cell death protein 1 expression in the tumor tissues
have an improved prognosis and a higher survival rate (Fig. 4L). Therefore, high levels of miR-33a
may be a favorable prognostic indicator for lung cancer (76).
Head and neck cancer
Laryngeal carcinoma
Laryngeal cancer is one of the most frequent head
and neck malignancies (77). Most
patients with laryngeal cancer (95%) have the habit of smoking or
consuming alcohol (78). Karatas
(79) demonstrated that high levels
of miR-33a inhibit the proliferation of laryngeal carcinoma cells
and promote apoptosis via downregulation of PIM1 (Fig. 5A).
 | Figure 5.Roles of miR-33a and its target genes
in laryngeal (top left) and tongue squamous (top right) carcinoma,
melanoma (bottom left and middle), renal cancer (bottom middle) and
thyroid cancer (bottom right), and the underlying mechanisms. (A)
miR-33a upregulation downregulates its target gene PIM1, inhibiting
proliferation and promotes apoptosis. (B) LncRNA CASC15
downregulates miR-33a and further upregulates its target gene ZEB1,
which promotes proliferation, migration and cell cycle. (C) miR-33a
upregulation inhibits its target gene HIF1A, downregulating LDHA,
HK1 and HK2, leading to inhibition of glycolysis, proliferation,
migration, invasion and promotion of apoptosis. (D) miR-33a
upregulation inhibits its target gene CDK16, downregulating p27,
leading to inhibition of proliferation. (E) miR-33a downregulation
upregulates its target gene MDM4, which promotes proliferation and
results in poor survival. (F) XB130 downregulates miR-33a and
further upregulates its target gene MYC, which promotes
proliferation. Red represents the factor causing miR-33a
dysregulation; blue represents miR-33a; yellow represents the
target genes of miR-33a; orange represents the downstream proteins;
light green represents the signaling pathways and EMT processes;
dark green represents the biological processes of cancer cells.
miR, microRNA; PIM1, Pim-1 proto-oncogene; ZEB1, Zinc finger E-Box
binding homeobox 1; HIF1A, hypoxia inducible factor 1 subunit α;
LDHA, lactate dehydrogenase A; HK, hexokinase; CDK,
cyclin-dependent kinase; MDM4, mouse double minute 4. |
Tongue squamous carcinoma (TSCC)
TSCC is one of the most aggressive oral cancers,
accounting for ~41% of oral cancer cases (80). miR-33a acts as a tumor suppressor in
TSCC. Zuo et al (81) have
reported that lncRNA CASC15 downregulates miR-33a-5p by miRNA
sponging, thereby upregulating the miR-33a-5p target ZEB1 and
enhancing the proliferation, migration and cell cycle of TSCC cells
(Fig. 5B).
Skin cancer
Melanoma
Melanoma is the fifth and seventh most common
malignant tumor in males and females, respectively (82). Melanoma-associated mortality accounts
for 80% of skin cancer-associated deaths, and the occurrence has
notably increased in previous years (83). The cause of this disease is
associated with both environmental and genetic factors, such as
sex, family history, geographical location and sun exposure
(84).
miR-33a suppresses melanoma. Zhou et al
(83) have illustrated that miR-33a
overexpression inhibits the proliferation, invasion, migration and
promotes apoptosis of malignant melanoma cells by downregulating
its target gene HIF1A (Fig. 5C). Cao
et al (85) have indicated
that miR-33a-5p expression is decreased in melanoma cells. High
miR-33a-5p expression downregulates HIF1A and the downstream
proteins lactate dehydrogenase A, hexokinase 1 (HK1) and HK2,
thereby repressing the glycolysis process and increasing the
radiation sensitivity of melanoma cells (Fig. 5C) (85). Tian et al (86) have revealed that miR-33a targets and
downregulates CDK16 in melanoma, consequently subduing the
phosphorylation of p27. As a result, the degradation of p27 is
reduced, and the proliferation of melanoma cells is inhibited
(Fig. 5D) (86). Lv et al (87) have demonstrated that lncRNA colon
cancer associated transcript 1 downregulates miR-33a through miRNA
sponging to promote the proliferation, migration and invasion of
melanoma cells. Additionally, Fu et al (88) have illustrated that lncRNA paternally
expressed 10 (PEG10) inhibits melanoma cell proliferation,
migration and invasion by downregulating miR-33a and activating the
PI3K/AKT and mTOR signaling pathways.
Urinary system cancer
Renal cancer
Renal cancer incidence in males is double as that in
females (89). The morbidity of
renal cancer has been increasing in recent years (90). Jiang et al (91) have demonstrated that miR-33a serves
as a tumor suppressor in renal cancer. Low miR-33a levels in renal
cancer cells lead to the upregulation of its target gene mouse
double minute 4 (Fig. 5E) (91). Consequently, the proliferation of the
cancer cells increases, causing poor overall survival in patients
with renal cancer (91).
Endocrine system cancer
Thyroid cancer
Thyroid cancer incidence has notably increased in
the past few years, and according to a study published in 2020, it
has been predicted that thyroid cancer will be the fourth leading
type of cancer in the world (92).
Takeshita et al (93) have
illustrated that actin filament associated protein 1 like 2, a
novel protein, suppresses miR-33a expression in thyroid cancer
cells, thereby upregulating the miR-33a target MYC, which
facilitates the proliferation of thyroid cancer cells (Fig. 5F).
Nervous system cancer
Glioma
Glioma is the most common and deadly malignancy of
the central nervous system (94).
The main causes of glioma are congenital factors and carcinogenic
environmental exposure (94).
Studies have indicated that miR-33a acts as an oncogene in glioma.
Chang et al (95) have
revealed that miR-33a is upregulated in glioma cells. Consequently,
the miR-33a target sirtuin 6 (SIRT6) is downregulated, reducing the
levels of cleaved caspase 8 and BAX, and promoting Bcl-2 expression
(95). Downregulation of SIRT6
increases the phosphorylation level of Janus kinase 2 (JAK2) and
STAT3, enhancing the JAK2/STAT3 pathway, which attenuates
H2O2-induced oxidative stress and suppresses
apoptosis in glioma cells, and thus miR-33a overexpression inhibits
apoptosis in glioma (Fig. 6A)
(95). Wang et al (96) have demonstrated that miR-33a
overexpression downregulates phosphodiesterase 8A (PDE8A) and UV
radiation resistance associated (UVRAG), and upregulates their
downstream proteins p-CREB and Notch intracellular domain,
respectively, in glioma cells. Consequently, the cAMP/protein
kinase A (PKA) and NOTCH signaling pathways are promoted, which
enhance the self-renewal of glioma-initiating cells (Fig. 6B) (96).
 | Figure 6.Roles of miR-33a and its target genes
in glioma (top) and osteosarcoma (bottom), and the underlying
mechanisms. (A) miR-33a upregulation inhibits its target gene
SIRT6, upregulating Bcl-2, p-JAK2 and p-STAT3, while downregulating
cleaved caspase 8 and BAX, leading to apoptosis inhibition. (B)
miR-33a upregulation downregulates its target genes PDE8A and
UVRAG, upregulating p-CREB and NICD, leading to self-renewal
promotion. (C) LncRNA DANCR downregulates miR-33a and further
upregulates its target gene AXL, which upregulates AKT, mTOR, S6K
and 4EBP1, leading to proliferation promotion. (D) Lovastatin
upregulates miR-33a and inhibits its target gene CYR61,
downregulating N-cadherin and upregulating E-cadherin, leading to
inhibition of EMT and invasion. (E) miR-33a upregulation
downregulates TWIST1, promoting chemoresistance and inhibiting
apoptosis. Red represents the factor causing miR-33a dysregulation;
blue represents miR-33a; yellow represents the target genes of
miR-33a; orange represents the downstream proteins; light green
represents the signaling pathways and EMT processes; dark green
represents the biological processes of cancer cells. miR, microRNA;
EMT, epithelial-mesenchymal transition; SIRT6, sirtuin 6; p-,
phosphorylated; JAK2, Janus kinase 2; PDE8A, phosphodiesterase 8A;
PKA, protein kinase A; TWIST1, Twist basic helix-loop-helix
transcription factor 1; UVRAG, UV radiation resistance associated;
NICD, Notch intracellular domain; AXL, AXL receptor tyrosine
kinase; 4EBP1, eukaryotic translation initiation factor 4E binding
protein 1; S6K, ribosomal protein S6K; CYR61, cysteine-rich
angiogenic inducer 61. |
Musculoskeletal system cancer
Osteosarcoma
Osteosarcoma is a primary bone tumor, which
accounts for 60% of all patients with sarcoma (97), and is more common in children and
young adults than in adults (98).
The role of miR-33a in osteosarcoma is controversial. Jiang et
al (99) have observed that
lncRNA DANCR is a miRNA sponge in osteosarcoma cells. DANCR targets
and downregulates miR-33a-5p, resulting in the upregulation of AXL
receptor tyrosine kinase (AXL), a target gene of miR-33a-5p
(99). Consequently, the PI3K/AKT
signaling pathway is activated, with the upregulation of the
downstream genes AKT, mTOR, ribosomal protein S6K (S6K) and 4EBP1
(99). These molecular changes
eventually promote cell proliferation and osteosarcoma development
(Fig. 6C) (99). Huang et al (100) have illustrated that lovastatin
enhances miR-33a expression in osteosarcoma. Therefore, the miR-33a
target cysteine-rich angiogenic inducer 61 and downstream protein
N-cadherin are downregulated, while E-cadherin is upregulated,
whereby the EMT and invasion of tumor cells are suppressed
(Fig. 6D) (100). Zhang et al (101) have illustrated that miR-33a-5p
expression is downregulated in osteosarcoma cells compared with in
normal tissues, and overexpressing miR-33a-5p suppresses cell
proliferation in osteosarcoma.
In contrast to the aforementioned studies, Zhou
et al (102) have observed
miR-33a upregulation in osteosarcoma. Consequently, the miR-33a
target TWIST1 is downregulated, whereby the cisplatin resistance of
osteosarcoma cells is enhanced, while their apoptosis is suppressed
(Fig. 6E) (102).
Male and female cancer
Prostate cancer (PC)
PC is the second most common cause of
male-cancer-associated deaths worldwide (103). Androgen is generally considered as
the main inducer of PC. High androgen levels are accordingly
associated with a high risk of PC. In addition, PC is associated
with carcinogenic environmental exposure, an unhealthy diet and
genetic factors (104,105).
Abnormal downregulation of miR-33a is observed in
PC. Karatas et al (106)
have revealed that miR-33a downregulation increases the expression
levels of PIM1 and other lipid β-oxidation pathway proteins, such
as carnitine palmitoyltransferase 1A and hydroxyacyl-CoA
dehydrogenase trifunctional multienzyme complex subunit β, thereby
enhancing the proliferation of PC cells (Fig. 7A). Li et al (107) have demonstrated that miR-33a is a
tumor suppressor in PC, and high levels of miR-33a downregulate its
target gene engrailed homeobox 2, thereby suppressing the
proliferation, migration and invasion of PC cells (Fig. 7B). Dai et al (108) have demonstrated that miR-33a-5p is
downregulated in PC cells. miR-33a-5p overexpression in PC cells
downregulates its target gene transforming growth factor β receptor
1 (TGFBRI), leading to the repression of the TGF-β signaling
pathway and suppressing PC invasion and migration (108). Additionally, ZEB1, a downstream
protein of the TGF-β signaling pathway, is consequently
downregulated, suppressing bone metastasis (108). Furthermore, ZEB1 downregulation
inhibits EMT by downregulating vimentin and upregulating
fibronectin in PC cells (Fig. 7C)
(108).
 | Figure 7.Roles of miR-33a and its target genes
in prostate (top) and breast cancer (bottom), and the underlying
mechanisms. (A) miR-33a downregulation upregulates its target gene
PIM1, as well as CPT1A and HADHB, which promote β-oxidation pathway
and proliferation. (B) miR-33a upregulation downregulates EN-2,
inhibiting proliferation, migration and invasion. (C) miR-33a
upregulation inhibits its target gene TGFBRI, which further
downregulates ZEB1, downregulating vimentin and upregulating
fibronectin, leading to inhibition of EMT, metastasis, invasion and
migration. (D) miR-33a upregulation downregulates its target genes
ROS1 and ADAM9, inhibiting proliferation and migration. (E) miR-33a
upregulation downregulates its target gene ABCA1, inhibiting
radiosensitivity. (F) miR-33a upregulation downregulates its target
gene EZH2, inhibiting proliferation and migration. (G) miR-33a
upregulation inhibits its target gene eIF5A2, downregulating
vimentin and upregulating E-cadherin, leading to inhibition of EMT
and chemoresistance. Blue represents miR-33a; yellow represents the
target genes of miR-33a; orange represents the downstream proteins;
light green represents the signaling pathways and EMT processes;
dark green represents the biological processes of cancer cells.
miR, microRNA; EMT, epithelial-mesenchymal transition; PIM1, Pim-1
proto-oncogene; EN-2, engrailed homeobox 2; TFGBRI, transforming
growth factor β receptor 1; ZEB1, Zinc finger E-Box binding
homeobox 1; ADAM9, ADAM metallopeptidase domain 9; ABCA1,
ATP-binding cassette transporter A1; EZH2, enhancer of zeste
homolog 2; eIF5A2, eukaryotic translation initiation factor 5A2;
CPT1A, carnitine palmitoyltransferase 1A; HADHB, hydroxyacyl-CoA
dehydrogenase trifunctional multienzyme complex subunit β; ROS1,
ROS proto-oncogene 1, receptor tyrosine kinase. |
Breast cancer
Breast carcinoma is the leading cause of
female-cancer-associated deaths worldwide (109). It is mainly associated with age,
long-term excessive alcohol consumption, genetic factors and
long-term use of exogenous estrogen (110,111).
miR-33a acts as a tumor suppressor in breast
carcinoma. Zhang et al (112) have demonstrated that miR-33a is
downregulated in breast cancer cells. Increased miR-33a levels
downregulate its target genes ROS1 and ADAM metallopeptidase domain
9, and thus inhibits tumor cell proliferation and migration
(Fig. 7D) (112). Additionally, Wolfe et al
(113) have demonstrated that
miR-33a is downregulated in inflammatory breast tumor cells.
Notably, miR-33a can decrease high-density-lipoprotein
(HDL)-induced radio-sensitivity by downregulating ABCA1 in breast
cancer cells (Fig. 7E) (113). Zeng et al (114) have demonstrated that miR-33a
expression is downregulated in triple-negative breast cancer (TNBC)
cells, whereas the miR-33a target enhancer of zeste homolog 2 is
upregulated. The high levels of miR-33a in TNBC cells can inhibit
the proliferation and migration of these cells (Fig. 7F) (114). Guan et al (115) have illustrated that compared with
non-TNBC cells, miR-33a-5p expression is significantly lower in
TNBC cells, and miR-33a-5p overexpression downregulates its target
gene eukaryotic translation initiation factor 5A2 (eIF5A2) and
increases the sensitivity of TNBC cells to doxorubicin.
Additionally, eIF5A2 downregulation decreases vimentin expression,
while increasing that of E-cadherin, thereby inhibiting the EMT of
TNBC cells (Fig. 7G) (115).
Discussion
The involvement of miRNAs in cancer is an exciting
research field, but the association and mechanism of interaction
between miRNAs and cancer remain at the preliminary stages of
research. Studies have indicated that miR-33a affects cell
proliferation, migration, metastasis, invasion and other biological
processes by acting on 38 different target genes, thereby affecting
the cell cycle, apoptosis, self-renewal and resistance to
traditional chemotherapy and radiation therapy in cancer cells.
Therefore, the prognosis and survival rate of patients with cancer
may be predicted based on the levels of miR-33a.
Diverse roles of miR-33a in
cancer
To date, studies have revealed that miR-33a is a
tumor suppressor gene and is downregulated in numerous types of
cancer, such as liver carcinoma, lung cancer, colorectal carcinoma,
PC and melanoma, whereas it is upregulated in glioma cells and acts
as an oncogene. In addition, it is important to note that miR-33a
serves different roles in osteosarcoma. Three studies (99–101)
have illustrated that miR-33a is downregulated in osteosarcoma, and
that its overexpression suppresses the proliferation, invasion and
migration of osteosarcoma cells. By contrast, Zhou et al
(102) reported that miR-33a is
upregulated in osteosarcoma and inhibits the apoptosis of
osteosarcoma cells with cisplatin-resistance. However, these
opposing observations in osteosarcoma cells are not specific to
miR-33a, and there are studies presenting contrasting expression
patterns in the same types of cancer for other miRNAs. For example,
Jang et al (116) have
revealed that miR-708 is an oncogene in NSCLC, and its increased
expression in NSCLC is associated with a low survival rate in
non-smoker patients with lung cancer. By contrast, Wu et al
(117) have demonstrated that
miR-708-5p is a tumor suppressor gene in NSCLC, and that miR-708-5p
overexpression inhibits the invasion and metastasis of NSCLC cells.
Abnormal miRNA expression in cancer may occur via various
mechanisms, but the precise mechanistic details of aberrant miR-33a
expression in tumors have not been clearly determined, and further
research is required. Furthermore, it remains unclear why there are
opposing observations regarding the expression patterns of miR-33a
in osteosarcoma cells.
Target genes of miR-33a in
malignancies
Among the 38 target genes mentioned in the
literature (Table I), 8 have been
verified in different types of cancer: TWIST1 in hepatocellular
carcinoma, gallbladder cancer, lung cancer and osteosarcoma;
β-catenin in hepatocellular carcinoma, pancreatic cancer and lung
cancer; HMGA2 in hepatocellular carcinoma and lung cancer; ABCA1 in
hepatocellular carcinoma, NSCLC and breast carcinoma; CDK6 in
gastric and lung cancer; PIM1 in gastric, colorectal, laryngeal and
prostate cancer; PIM3 in pancreatic and colorectal cancer; and ZEB1
in gastric cancer, esophageal squamous carcinoma and tongue
squamous cell carcinoma. These target genes may be reliable
therapeutic targets as the target genes of miR-33a. In addition,
future studies should continue to investigate this miRNA and its
eight target genes in other types of cancer that have not been
analyzed yet, to explore whether these genes have similar
interactions and mechanisms affecting these other types of cancer.
The remaining 30 target genes are currently only validated in one
type of cancer each, and thus, these target genes have great
research potential. The expression levels, mechanism and
reliability of miR-33a and its association with these targets in
other types of cancer should be further explored. The reliability
of their interactions and underlying mechanistic details in cancer
should also be assessed.
 | Table I.miR-33a expression and roles in
different types of cancer. |
Table I.
miR-33a expression and roles in
different types of cancer.
| A, Digestive
system: Hepatocellular carcinoma |
|---|
|
|---|
| First author,
year | miR-33a
expression | Target gene | Target gene
expression | Function | (Refs.) |
|---|
| Liu et al,
2018 | Down | PNMA1 | Up | EMT, proliferation,
migration | (29) |
| Guo et al,
2016; Li et al, 2019 | Down | TWIST1 | Up | EMT, invasion,
proliferation | (30,31) |
| Fang et al,
2013 | Down | β-catenin | Up | Proliferation | (32) |
| Han et al,
2016 | Down | PBX3 | Up | Proliferation,
metastasis, invasion, low survival | (33) |
| Wu et al,
2019 | Down | HMGA2 | Up | Proliferation,
migration, invasion, drug resistance | (34,35) |
| Han et al,
2019 |
| Meng et al,
2017 | Down | HSPA8 | Up | Cisplatin
resistance | (36) |
| Hou et al,
2017 | Down | ABCA1 | Up | Doxorubicin
resistance | (37) |
| Chang et al,
2017 | Up | PPARα | Down | Proliferation,
apoptosis | (38) |
|
| B, Digestive
system: Gastric cancer |
| First author,
year | miR-33a
expression | Target
gene | Target gene
expression |
Function | (Refs.) |
|
| Wang et al,
2015 | Down | CDK6 | Up | Proliferation, cell
cycle | (41) |
|
|
| CCND1 |
|
|
|
|
|
| PIM1 |
|
|
|
| Chen et al,
2019 | Down | SNAI2 | Up | EMT, invasion,
migration | (42) |
| Wu et al,
2018 | Down | ZEB1 | Up | EMT, migration | (43) |
|
| C, Digestive
system: Colorectal cancer |
| First author,
year | miR-33a
expression | Target
gene | Target gene
expression |
Function | (Refs.) |
|
| Wang et al,
2019 | Down | PIM3 | Up | Proliferation, cell
cycle | (50) |
| Shan et al,
2017 | Down | ST8SIA1 | Up | Proliferation,
invasion, drug resistance | (51) |
| Yan et al,
2019 | Down | MTHFD2 | Up | Proliferation,
migration, cell cycle | (52) |
| Ibrahim et
al, 2011 | Down | PIM1 | Up | Tumor growth | (53) |
|
| D, Digestive
system: Pancreatic cancer |
| First author,
year | miR-33a
expression | Target
gene | Target gene
expression |
Function | (Refs.) |
|
| Liang et al,
2015 | Down | PIM3 | Up | Proliferation,
gemcitabine sensitivity | (56) |
| Liang et al,
2015 | Down | β-catenin | Up | EMT, proliferation,
gemcitabine sensitivity | (57) |
|
| E, Digestive
system: Gallbladder cancer |
|
| First author,
year | miR-33a
expression | Target
gene | Target gene
expression |
Function | (Refs.) |
|
| Zhang et al,
2016 | Down | TWIST1 | Up | EMT, proliferation,
migration, invasion | (59) |
|
| F, Digestive
system: Esophageal squamous carcinoma |
|
| First author,
year | miR-33a
expression | Target
gene | Target gene
expression |
Function | (Refs.) |
|
| Zhang et al,
2019 | Down | ZEB1 | Up | Proliferation,
migration | (62) |
| Wang et al,
2017 | Down | SOX9 | Up | Proliferation | (63) |
|
| G, Respiratory
system: Non-small cell lung cancer |
|
| First author,
year | miR-33a
expression | Target
gene | Target gene
expression |
Function | (Refs.) |
|
| Du et al,
2017 | Down | METTL3 | Up | Proliferation | (65) |
| Kang et al,
2018 | Down | CAND1 | Up | Proliferation,
migration Cell cycle | (66) |
| Amaar and Reeves
et al, 2019 | Down | ABCA1 | Up | EMT, proliferation,
migration, chemoresistance | (67) |
| Yang et al,
2015 | Down | TWIST1 | Up | Migration, EMT | (68) |
| Kang et al,
2017 | Down | CDK6 | Up | Cell cycle, tumor
development | (69) |
| Wang et al,
2020 | Down | KPNA4 | Up | Tumorigenesis,
tumor development | (70) |
|
| H, Respiratory
system: Untyped lung cancer |
|
| First author,
year | miR-33a
expression | Target
gene | Target gene
expression |
Function | (Refs.) |
|
| Li et al,
2018 | Down | mTOR | Up | Proliferation,
celastrol resistance | (71) |
| Kuo et al,
2013 | Down | CDK6 | Up | Bone
metastasis | (72) |
| Pan et al,
2020 | Down | TWIST1 | Up | Proliferation,
metastasis | (73) |
| Rice et al,
2013 | Down | HMGA2 | Up | Migration | (74) |
| Zhu et al,
2015 | Down | β-catenin | Up | Proliferation, cell
cycle | (75) |
| Boldrini et
al, 2017 | Down | PD-1 | Up | Prognosis | (76) |
|
| I, Head and
neck: Laryngeal carcinoma |
|
| First author,
year | miR-33a
expression | Target
gene | Target gene
expression |
Function | (Refs.) |
|
| Karatas et
al, 2018 | Down | PIM1 | Up | Proliferation,
apoptosis | (79) |
|
| J, Head and
neck: Tongue squamous carcinoma |
|
| First author,
year | miR-33a
expression | Target
gene | Target gene
expression |
Function | (Refs.) |
|
| Zuo et al,
2018 | Down | ZEB1 | Up | Proliferation,
migration, cell cycle | (81) |
|
| K, Skin:
Melanoma |
|
| First author,
year | miR-33a
expression | Target
gene | Target gene
expression |
Function | (Refs.) |
|
| Zhou et al,
2015 Cao et al, 2017 | Down | HIF1A | Up | Proliferation,
migration, invasion, apoptosis, radiosensitivity | (83,85) |
| Tian et al,
2016 | Down | CDK16 | Up | Proliferation | (86) |
| Lv et al,
2018 | Down | N/A | N/A | Proliferation,
migration, invasion | (87,88) |
| Fu et al,
2019 |
|
| L, Urinary
system: Renal cancer |
|
| First author,
year | miR-33a
expression | Target
gene | Target gene
expression |
Function | (Refs.) |
|
| Jiang et al,
2019 | Down | MDM4 | Up | Proliferation,
survival | (91) |
|
| M, Endocrine
system: Thyroid cancer |
|
| First author,
year | miR-33a
expression | Target
gene | Target gene
expression |
Function | (Refs.) |
|
| Takeshita et
al, 2013 | Down | MYC | Up | Proliferation | (93) |
|
| N, Nervous
system: Glioma |
|
| First author,
year | miR-33a
expression | Target
gene | Target gene
expression |
Function | (Refs.) |
|
| Chang et al,
2017 | Up | SIRT6 | Down | Apoptosis | (95) |
| Wang et al,
2014 | Up | PDE8A, UVRAG | Down | Self-renewal | (96) |
|
| O,
Musculoskeletal system: Osteosarcoma |
|
| First author,
year | miR-33a
expression | Target
gene | Target gene
expression |
Function | (Refs.) |
|
| Jiang et al,
2017 | Down | AXL | Up | Proliferation | (99) |
| Huang et al,
2018 | Down | CYR61 | Up | EMT, invasion | (100) |
| Zhang et al,
2015 | Down | N/A | N/A | Proliferation | (101) |
| Zhou et al,
2014 | Up | TWIST1 | Down | Cisplatin
resistance, apoptosis | (102) |
|
| P, Reproductive
system: Prostate cancer |
|
| First author,
year | miR-33a
expression | Target
gene | Target gene
expression |
Function | (Refs.) |
|
| Karatas et
al, 2017 | Down | PIM1 | Up | Proliferation,
invasion, migration | (106) |
| Li et al,
2017 | Down | EN-2 | Up | Proliferation,
migration, invasion | (107) |
| Dai et al,
2019 | Down | TGFBRI | Up | EMT, invasion,
migration, metastasis | (108) |
|
| Q, Reproductive
system: Breast cancer |
|
| First author,
year | miR-33a
expression | Target
gene | Target gene
expression |
Function | (Refs.) |
|
| Zhang et al,
2015 | Down | ADAM9 | Up | Proliferation,
migration | (112) |
|
|
| ROS1 |
|
|
|
| Wolfe et al,
2016 | Down | ABCA1 | Up |
Radiosensitivity | (113) |
| Weihua et
al, 2020 | Down | EZH2 | Up | Proliferation,
migration | (114) |
| Guan et al,
2019 | Down | eFI5A2 | Up | EMT, doxorubicin
sensitivity | (115) |
Signaling pathways affected by miR-33a
in carcinogenesis
The mechanisms underlying miR-33a-mediated
carcinogenesis are extremely complex and involve multiple signaling
pathways. There are nine signaling pathways mentioned in the
literature, namely the Wnt/β-catenin, Snail/Slug,
AKT/GSK-3β/β-catenin, mTOR, TGF-β, JAK2/STAT3, PKA, NOTCH and
PI3K/AKT signaling pathways.
Liu et al (29) have revealed that PNMA1, a target gene
of miR-33a, is upregulated in hepatocellular carcinoma. PNMA1
upregulation enables β-catenin to translocate to the nucleus of
hepatoma cells, stimulating Wnt/β-catenin signal transduction, and
consequently promoting the proliferation and EMT of hepatoma cells.
Fang et al (32) have
illustrated that miR-33a also directly targets β-catenin and thus
inhibits the proliferation of hepatoma cells. In general, GSK-3β
regulates the phosphorylation and degradation of β-catenin
(118), but a study by Fang et
al (32) has indicated that
GSK-3β is not associated with the miR-33a-mediated downregulation
of β-catenin in hepatoma cells. Liang et al (56) have revealed that PIM3, a target of
miR-33a, may regulate the AKT and Wnt/β-catenin signaling pathways.
PIM3 upregulation in pancreatic cancer increases the
phosphorylation of GSK-3β and AKT, and the expression levels of
β-catenin (56). Therefore, miR-33a
and PIM3 may act on the AKT/Wnt/β-catenin signaling pathway to
inhibit the proliferation and gemcitabine-resistance of pancreatic
tumors (56). In lung cancer, Pan
et al (73) have demonstrated
that miR-33a downregulation upregulates its target gene TWIST1 and
activates the Wnt/β-catenin signaling pathway, subsequently
promoting lung cancer cell proliferation and metastasis. TWIST1
upregulation, GSK-3β downregulation and activation of the
Wnt/β-catenin signaling pathway have also been observed in NSCLC by
Zheng et al (119).
According to the aforementioned studies, the association between
miR-33a and the Wnt/β-catenin signaling pathway in hepatocellular
carcinoma, pancreatic cancer and lung cancer remains unclear and
requires further clarification in the future.
Chen et al (42) have revealed that miR-33a inhibits the
EMT, invasion and migration of gastric tumor cells by targeting
SNAI2 to deactivate the Snail/Slug signaling pathway, which also
serves a similar role in tongue squamous cell carcinoma (120) and thyroid cancer (121). Cao et al (122) have reported that, in invasive
ductal carcinoma, SNAI2 promotes the EMT process by inhibiting
E-cadherin expression. Slug is encoded by the SNAI2 gene, and the
two constitute the Snail/Slug signaling pathway. Nevertheless, the
specific mechanism of miR-33a in this signaling pathway has not
been thoroughly clarified yet.
Liang et al (56) have demonstrated that miR-33a
upregulation in pancreatic cancer downregulates PIM3 and inhibits
the AKT/GSK-3β/β-catenin signaling pathway, thus suppressing the
proliferation and gemcitabine-resistance in pancreatic cells. A
study by Carreras-Torres et al (123) has demonstrated that type 2 diabetes
is one of the risk factors of pancreatic cancer. Furthermore, the
inhibition of the AKT/GSK-3β/β-catenin signaling pathway can
prevent the development of type 2 diabetes (124). In addition, Wijesekara et al
(125) have reported that miR-33a
overexpression in pancreatic tissues decreases insulin secretion,
and increases blood cholesterol levels. Although miR-33a is known
to suppress the AKT/GSK-3β/β-catenin signaling pathway in
pancreatic cancer cells, the mechanism requires further detailed
study.
mTOR can activate p70S6K and phosphorylation of
4EBP1, which are strongly associated with cancer cell proliferation
(126). Li et al (71) have illustrated that miR-33a targets
mTOR and downregulates p-p70S6K and p-4EBP1 in lung cancer,
indicating that miR-33a suppresses the proliferation of lung cancer
cells via the mTOR signaling pathway. In addition, Xia and Xu
(127) have illustrated that the
PI3K/AKT/mTOR signaling pathway can act altogether, and inhibiting
this axis can impede the self-renewal of tumor cells, as well as
the clone formation and differentiation of tumor stem cells. mTOR
is a downstream protein of AKT, and phosphorylation of AKT
increases mTOR expression (128,129).
Consequently, the expression levels of the downstream protein
p70S6K and phosphorylation of 4EBP1 are upregulated, promoting the
proliferation of cancer cells (126). Fu et al (88) have demonstrated that lncRNA PEG10
inhibits miR-33a and promotes melanoma cell proliferation,
migration and invasion by activating the PI3K/AKT and mTOR
signaling pathways. In osteosarcoma, Jiang et al (99) have demonstrated that lncRNA DANCR
upregulates the target protein AXL in osteosarcoma cells by
downregulating miR-33a-5p, thereby increasing the expression levels
of the downstream proteins PI3K and AKT. Therefore, this novel
regulatory network of DANCR-miR-33a-AXL-PI3K/AKT/mTOR is expected
to provide a novel therapeutic target for osteosarcoma (99). All aforementioned studies demonstrate
that miR-33a is closely associated with the proteins in the
PI3K/AKT/mTOR signaling pathway, which serves a vital role in tumor
development.
Dai et al (108) have demonstrated that increasing
miR-33a expression in PC cells downregulates the target gene
TGFBRI, leading to repression of the TGF-β signaling pathway and
suppression of bone metastasis. Additionally, Fournier et al
(130) have observed that blocking
the TGF-β signaling pathway suppresses bone metastasis in PC.
However, the inhibitory effect of miR-33a on bone metastasis in PC
via suppression of the TGF-β signaling pathway remains unclear, and
further research is required.
Chang et al (95) and Feng et al (131) have demonstrated that SIRT6, a
target protein of miR-33a, inhibits JAK2/STAT3 signaling pathway
activation in glioma cells by decreasing the phosphorylation of
JAK2 and STAT3. This signaling pathway is highly activated in
numerous malignant cells, including glioma cells, and its
repression can inhibit tumor growth, invasion and EMT (132,133).
Tang et al (134) have
indicated that the glioma tumor suppressor miR-876-3p downregulates
its target gene kinesin family member 20A, whereby the JAK2/STAT3
signaling pathway is repressed; consequently, the proliferation,
invasion and EMT of glioma cells are suppressed. High SIRT6
expression can block the growth of these cancer cells and
facilitate their apoptosis. In addition, in glioma cancer cells,
miR-33a targets PDE8A and UVRAG, whose overexpression activates the
cAMP/PKA and NOTCH signaling pathways, respectively (96). Notably, the two signaling pathways
can promote the self-renewal of glioma initiation cells only when
simultaneously activated (91). In
glioma, Tanabe et al (135)
have revealed that the adenylyl cyclase/cAMP/PKA signaling pathway
increases IL-1β-induced IL-6 synthesis by enhancing the JAK2/STAT3
signaling pathway. Liu et al (136) have illustrated that the oncogene
TRPM7 promotes the proliferation, migration and invasion of glioma
cells by activating the JAK2/STAT3 and Notch signaling pathways.
Overall, the JAK2/STAT3, cAMP/PKA and NOTCH signaling pathways are
critical in glioma. The mechanisms underlying the regulation of
these signaling pathways by miR-33a require further study.
miR-33a in tumor prognosis and
treatment
Numerous studies have reported the crucial role of
miR-33a in carcinogenesis. Accordingly, miR-33a may be used for the
prognosis or treatment of tumors. Xie et al (137) have illustrated that low miR-33a
expression is associated with tumorigenesis and poor prognosis in
hepatoma. Hou et al (138)
have demonstrated that adjuvant chemotherapy improves the overall
and disease-free survival rates of patients with NSCLC with high
miR-33a expression. Therefore, miR-33a may be used as a biomarker
for NSCLC prognosis. In addition to the experimental evidence,
bioinformatics studies have predicted that miR-33a expression
patterns may be used as a prognostic marker in cancer. For example,
Pan et al (139) have
demonstrated that a two-miRNA signature consisting of miR-33a-5p
and miR-128 may serve as a novel biomarker for early NSCLC
detection. Kandimalla et al (140) have identified an eight-miRNA
signature, which includes miR-33a, that achieved excellent
prediction for colorectal tumor recurrence in stages two and
three.
As aforementioned, miRNAs may be used as novel
therapeutic targets for cancer (9).
The involvement of miR-33a in carcinogenesis enables cancer
treatment by modulating the expression levels of miR-33a. Previous
studies have indicated that high miR-33a levels affect the
chemosensitivity of hepatocellular carcinoma (26,36,37),
glioma (141) and osteosarcoma
(102), as well as colorectal
(51), pancreatic (56,57),
lung (67,71) and breast cancer (115). The radiosensitivity of cancer cells
can also be affected by miR-33a expression; high levels of miR-33a
decrease HDL-induced radiosensitivity of breast cancer (113), whereas in melanoma, miR-33a
increases the radiosensitivity by repressing the glycolysis process
(85).
Overall, miR-33a is a potential cancer-associated
miRNA that deserves further investigation. At present, only a small
part of the literature mentions the signaling pathways affected by
miR-33a and its target genes in tumors. Most of the
miR-33a-modulated signaling pathways associated with cancer remain
unclear. Once the action mechanisms of miR-33a in cancer are
clarified, it may be possible to determine high-risk cancer cases
and predict prognosis according to the levels of miR-33a and its
targets in the cancer cells of the patients, and effectively design
targeted therapies. Finally, to the best of our knowledge, the
present paper is the first review of the involvement of miR-33a and
its target genes in diverse types of cancer, thereby providing
comprehensive information for future research on the involvement of
miR-33a in cancer.
Conclusions
Abnormal expression levels of miRNAs, such as
miR-33a, are associated with various diseases, including cancer. In
the present review, the roles of miR-33a and its 38 target genes
were summarized in more than a dozen types of cancer. Among the 38
target genes of this miRNA, 8 have been confirmed in various types
of cancer, while the rest have been validated in only one type of
cancer each.
miR-33a and its target genes affect different
biological behaviors of tumor cells, such as proliferation,
metastasis, migration, invasion, cell cycle, apoptosis,
self-renewal, chemoresistance and radioresistance, as well as
patient outcomes, such as prognosis and survival. miR-33a serves
diverse regulatory roles in cancer. It is a tumor suppressor in
most types of cancer, whereas it acts as an oncogene in glioma, and
it possibly serves controversial roles in osteosarcoma. These
different roles of miR-33a imply that it may affect different types
of cancer via different molecular mechanisms.
miR-33a participates in nine signaling pathways
that are associated with cancer. These pathways are the
Wnt/β-catenin, Snail/Slug, AKT/GSK-3β/β-catenin, mTOR, TGF-β,
JAK2/STAT3, PKA, NOTCH and PI3K/AKT pathways, although the
mechanistic details of their involvement in cancer remain elusive.
Therefore, further studies are required to explore the precise
mechanistic roles of miR-33a in these types of cancer, so that
novel targeted treatments can be designed and developed to improve
patient outcomes.
Acknowledgements
Not applicable.
Funding
The present review was supported by the National
Natural Science Foundation of China (grant no. 31770774) and the
Provincial Major Project of Basic or Applied Research in Natural
Science, Guangdong Provincial Education Department (grant no.
2016KZDXM038).
Availability of data and materials
Not applicable.
Authors' contributions
ZH and CG conceptualized the present review. CG, JW
and TT contributed to the formal analysis and investigation. CG
wrote the original draft. ZH revised and edited the original draft,
and provided funding acquisition. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
ABCA1
|
ATP-binding cassette transporter
A1
|
|
AXL
|
AXL receptor tyrosine kinase
|
|
CDK
|
cyclin-dependent kinase
|
|
eIF5A2
|
eukaryotic translation initiation
factor 5A2
|
|
HIF1A
|
hypoxia inducible factor 1 subunit
α
|
|
HK1/2
|
hexokinase 1/2
|
|
HMGA2
|
high mobility group AT-hook 2
|
|
IL
|
interleukin
|
|
JAK2
|
Janus kinase 2
|
|
miRNA/miR
|
microRNA
|
|
PDE8A
|
phosphodiesterase 8A
|
|
PIM1/3
|
Pim-1/3 proto oncogene
|
|
PNMA1
|
paraneoplastic antigen Ma1
|
|
RAFFS1C
|
Ras association domain family member
1
|
|
SIRT6
|
sirtuin 6
|
|
SNAI1/2
|
Snail family transcriptional
repressor 1/2
|
|
SOX9
|
SRY-box 9
|
|
ST8SIA1
|
ST8α-N-acetyl-neuraminide-α-2,8-sialyltransferase 1
|
|
S6K
|
ribosomal protein S6K
|
|
TGFBRI
|
transforming growth factor β receptor
1
|
|
TWIST1
|
Twist basic helix-loop-helix
transcription factor 1
|
|
UVRAG
|
UV radiation resistance
associated
|
|
ZEB1
|
zinc finger E-Box binding homeobox
1
|
|
4EBP1
|
eukaryotic translation initiation
factor 4E binding protein 1
|
References
|
1
|
Finnegan EF and Pasquinelli AE: MicroRNA
biogenesis: Regulating the regulators. Crit Rev Biochem Mol Biol.
48:51–68. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Acunzo M, Romano G, Wernicke D and Croce
CM: MicroRNA and cancer-a brief overview. Adv Biol Regul. 57:1–9.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lee YS and Dutta A: MicroRNAs in cancer.
Annu Rev Pathol. 4:199–227. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kappel A and Keller A: miRNA assays in the
clinical laboratory: Workflow, detection technologies and
automation aspects. Clin Chem Lab Med. 55:636–647. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
The Lancet, . GLOBOCAN 2018: Counting the
toll of cancer. Lancet. 392:9852018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Calin GA, Sevignani C, Dumitru CD, Hyslop
T, Noch E, Yendamuri S, Shimizu M, Rattan S, Bullrich F, Negrini M
and Croce CM: Human microRNA genes are frequently located at
fragile sites and genomic regions involved in cancers. Proc Natl
Acad Sci USA. 101:2999–3004. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Calin GA, Dumitru CD, Shimizu M, Bichi R,
Zupo S, Noch E, Aldler H, Rattan S, Keating M, Rai K, et al:
Frequent deletions and down-regulation of micro-RNA genes miR15 and
miR16 at 13q14 in chronic lymphocytic leukemia. Proc Natl Acad Sci
USA. 99:15524–15529. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
He L, Thomson JM, Hemann MT,
Hernando-Monge E, Mu D, Goodson S, Powers S, Cordon-Cardo C, Lowe
SW, Hannon GJ and Hammond SM: A microRNA polycistron as a potential
human oncogene. Nature. 435:828–833. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shell S, Park SM, Radjabi AR, Schickel R,
Kistner EO, Jewell DA, Feig C, Lengyel E and Peter ME: Let-7
expression defines two differentiation stages of cancer. Proc Natl
Acad Sci USA. 104:11400–11405. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pineau P, Volinia S, McJunkin K, Marchio
A, Battiston C, Terris B, Mazzaferro V, Lowe SW, Croce CM and
Dejean A: miR-221 overexpression contributes to liver
tumorigenesis. Proc Natl Acad Sci USA. 107:264–269. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Felli N, Fontana L, Pelosi E, Botta R,
Bonci D, Facchiano F, Liuzzi F, Lulli V, Morsilli O, Santoro S, et
al: MicroRNAs 221 and 222 inhibit normal erythropoiesis and
erythroleukemic cell growth via kit receptor down-modulation. Proc
Natl Acad Sci USA. 102:18081–18086. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liang Z, Li Y, Huang K, Wagar N and Shim
H: Regulation of miR-19 to breast cancer chemoresistance through
targeting PTEN. Pharm Res. 28:3091–3100. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shi GH, Ye DW, Yao XD, Zhang SL, Dai B,
Zhang HL, Shen YJ, Zhu Y, Zhu YP, Xiao WJ and Ma CG: Involvement of
microRNA-21 in mediating chemo-resistance to docetaxel in
androgen-independent prostate cancer PC3 cells. Acta Pharmacol Sin.
31:867–873. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Acunzo M, Visone R, Romano G, Veronese A,
Lovat F, Palmieri D, Bottoni A, Garofalo M, Gasparini P, Condorelli
G, et al: miR-130a targets MET and induces TRAIL-sensitivity in
NSCLC by downregulating miR-221 and 222. Oncogene. 31:634–642.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bao L, Hazari S, Mehra S, Kaushal D, Moroz
K and Dash S: Increased expression of P-glycoprotein and
doxorubicin chemoresistance of metastatic breast cancer is
regulated by miR-298. Am J Pathol. 180:2490–2503. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rottiers V and Näär AM: MicroRNAs in
metabolism and metabolic disorders. Nat Rev Mol Cell Biol.
13:239–250. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fernández-Hernando C, Suárez Y, Rayner KJ
and Moore KJ: MicroRNAs in lipid metabolism. Curr Opin Lipidol.
22:86–92. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Brown MS and Goldstein JL: The SREBP
pathway: Regulation of cholesterol metabolism by proteolysis of a
membrane-bound transcription factor. Cell. 89:331–340. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bengoechea-Alonso MT and Ericsson J: SREBP
in signal transduction: Cholesterol metabolism and beyond. Curr
Opin Cell Biol. 19:215–222. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Boren J and Brindle KM: Apoptosis-induced
mitochondrial dysfunction causes cytoplasmic lipid droplet
formation. Cell Death Differ. 19:1561–1570. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gharipour M and Sadeghi M: Pivotal role of
microRNA-33 in metabolic syndrome: A systematic review. ARYA
Atheroscler. 9:372–376. 2013.PubMed/NCBI
|
|
25
|
Nishida N and Goel A: Genetic and
epigenetic signatures in human hepatocellular carcinoma: A
systematic review. Curr Genomics. 12:130–137. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Karkampouna S, van der Helm D, Gray PC,
Chen L, Klima I, Grosjean J, Burgmans MC, Farina-Sarasqueta A,
Snaar-Jagalska EB, Stroka DM, et al: CRIPTO promotes an aggressive
tumour phenotype and resistance to treatment in hepatocellular
carcinoma. J Pathol. 245:297–310. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xu W, Liu H, Liu ZG, Wang HS, Zhang F,
Wang H, Zhang J, Chen JJ, Huang HJ, Tan Y, et al: Histone
deacetylase inhibitors upregulate Snail via Smad2/3 phosphorylation
and stabilization of Snail to promote metastasis of hepatoma cells.
Cancer Lett. 420:1–13. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gingold JA, Zhu D, Lee DF, Kaseb A and
Chen J: Genomic profiling and metabolic homeostasis in primary
liver cancers. Trends Mol Med. 24:395–411. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu P, Chen B, Gu Y and Liu Q: PNMA1,
regulated by miR-33a-5p, promotes proliferation and EMT in
hepatocellular carcinoma by activating the Wnt/β-catenin pathway.
Biomed Pharmacother. 108:492–499. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Guo XF, Wang AY and Liu J:
HIFs-MiR-33a-Twsit1 axis can regulate invasiveness of
hepatocellular cancer cells. Eur Rev Med Pharmacol Sci.
20:3011–3016. 2016.PubMed/NCBI
|
|
31
|
Li Y, Chen G, Yan Y and Fan Q: CASC15
promotes epithelial to mesenchymal transition and facilitates
malignancy of hepatocellular carcinoma cells by increasing TWIST1
gene expression via miR-33a-5p sponging. Eur J Pharmacol.
860:1725892019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Fang Y, Feng Y, Wu T, Srinivas S, Yang W,
Fan J, Yang C and Wang S: Aflatoxin B1 negatively regulates
Wnt/β-catenin signaling pathway through activating miR-33a. PLoS
One. 8:e730042013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Han SY, Han HB, Tian XY, Sun H, Xue D,
Zhao C, Jiang ST, He XR, Zheng WX, Wang J, et al: MicroRNA-33a-3p
suppresses cell migration and invasion by directly targeting PBX3
in human hepatocellular carcinoma. Oncotarget. 7:42461–42473. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wu S, Ai H, Zhang K, Yun H and Xie F: Long
non-coding RNA EGOT promotes the malignant phenotypes of
hepatocellular carcinoma cells and increases the expression of
HMGA2 via down-regulating miR-33a-5p. Onco Targets Ther.
12:11623–11635. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Han S, Han B, Li Z and Sun D:
Downregulation of long noncoding RNA CRNDE suppresses drug
resistance of liver cancer cells by increasing microRNA-33a
expression and decreasing HMGA2 expression. Cell Cycle.
18:2524–2537. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Meng W, Tai Y, Zhao H, Fu B, Zhang T, Liu
W, Li H, Yang Y, Zhang Q, Feng Y and Chen G: Downregulation of
miR-33a-5p in hepatocellular carcinoma: A possible mechanism for
chemotherapy resistance. Med Sci Monit. 23:1295–1304. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hou H, Kang Y, Li Y, Zeng Y, Ding G and
Shang J: miR-33a expression sensitizes Lgr5+ HCC-CSCs to
doxorubicin via ABCA1. Neoplasma. 64:81–91. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chang W, Zhang L, Xian Y and Yu Z:
MicroRNA-33a promotes cell proliferation and inhibits apoptosis by
targeting PPARα in human hepatocellular carcinoma. Exp Ther Med.
13:2507–2514. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Pasechnikov V, Chukov S, Fedorov E,
Kikuste I and Leja M: Gastric cancer: Prevention, screening and
early diagnosis. World J Gastroenterol. 20:13842–13862. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hartgrink HH, Jansen EP, van Grieken NC
and van de Velde CJ: Gastric cancer. Lancet. 374:477–490. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wang Y, Zhou X, Shan B, Han J, Wang F, Fan
X, Lv Y, Chang L and Liu W: Downregulation of microRNA-33a promotes
cyclin- dependent kinase 6, cyclin D1 and PIM1 expression and
gastric cancer cell proliferation. Mol Med Rep. 12:6491–6500. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chen DD, Cheng JT, Chandoo A, Sun XW,
Zhang L, Lu MD, Sun WJ and Huang YP: microRNA-33a prevents
epithelial-mesenchymal transition, invasion, and metastasis of
gastric cancer cells through the Snail/Slug pathway. Am J Physiol
Gastrointest Liver Physiol. 317:G147–G160. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wu Q, Xiang S, Ma J, Hui P, Wang T, Meng
W, Shi M and Wang Y: Long non-coding RNA CASC15 regulates gastric
cancer cell proliferation, migration and epithelial mesenchymal
transition by targeting CDKN1A and ZEB1. Mol Oncol. 12:799–813.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA Cancer J Clin. 69:7–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Herbey II, Ivankova NV, Katkoori VR and
Mamaeva OA: Colorectal cancer and hypercholesterolemia: Review of
current research. Exp Oncol. 27:166–178. 2005.PubMed/NCBI
|
|
47
|
Gray RT, Loughrey MB, Bankhead P, Cardwell
CR, McQuaid S, O'Neill RF, Arthur K, Bingham V, McGready C, Gavin
AT, et al: Statin use, candidate mevalonate pathway biomarkers, and
colon cancer survival in a population-based cohort study. Br J
Cancer. 116:1652–1659. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Murai T: Cholesterol lowering: Role in
cancer prevention and treatment. Biol Chem. 396:1–11. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Jacobs RJ, Voorneveld PW, Kodach LL and
Hardwick JC: Cholesterol metabolism and colorectal cancers. Curr
Opin Pharmacol. 12:690–695. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wang Y, Liu C and Hu L: Cholesterol
regulates cell proliferation and apoptosis of colorectal cancer by
modulating miR-33a-PIM3 pathway. Biochem Biophys Res Commun.
511:685–692. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Shan Y, Liu Y, Zhao L, Liu B, Li Y and Jia
L: MicroRNA-33a and let-7e inhibit human colorectal cancer
progression by targeting ST8SIA1. Int J Biochem Cell Biol.
90:48–58. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Yan Y, Zhang D, Lei T, Zhao C, Han J, Cui
J and Wang Y: MicroRNA-33a-5p suppresses colorectal cancer cell
growth by inhibiting MTHFD2. Clin Exp Pharmacol Physiol.
46:928–936. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Ibrahim AF, Weirauch U, Thomas M,
Grünweller A, Hartmann RK and Aigner A: MicroRNA replacement
therapy for miR-145 and miR-33a is efficacious in a model of colon
carcinoma. Cancer Res. 71:5214–5224. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Sener SF, Fremgen A, Menck HR and
Winchester DP: Pancreatic cancer: A report of treatment and
survival trends for 100,313 patients diagnosed from 1985–1995,
using the national cancer database. J Am Coll Surg. 189:1–7. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Moffat GT, Epstein AS and O'Reilly EM:
Pancreatic cancer-A disease in need: Optimizing and integrating
supportive care. Cancer. 125:3927–3935. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Liang C, Yu XJ, Guo XZ, Sun MH, Wang Z,
Song Y, Ni QX, Li HY, Mukaida N and Li YY: MicroRNA-33a-mediated
downregulation of Pim-3 kinase expression renders human pancreatic
cancer cells sensitivity to gemcitabine. Oncotarget. 6:14440–14455.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Liang C, Wang Z, Li YY, Yu BH, Zhang F and
Li HY: miR-33a suppresses the nuclear translocation of β-catenin to
enhance gemcitabine sensitivity in human pancreatic cancer cells.
Tumour Biol. 36:9395–9403. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Sharma A, Sharma KL, Gupta A, Yadav A and
Kumar A: Gallbladder cancer epidemiology, pathogenesis and
molecular genetics: Recent update. World J Gastroenterol.
23:3978–3998. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Zhang M, Gong W, Zuo B, Chu B, Tang Z,
Zhang Y, Yang Y, Zhou D, Weng M, Qin Y, et al: The microRNA miR-33a
suppresses IL-6-induced tumor progression by binding Twist in
gallbladder cancer. Oncotarget. 7:78640–78652. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Arnold M, Soerjomataram I, Ferlay J and
Forman D: Global incidence of oesophageal cancer by histological
subtype in 2012. Gut. 64:381–387. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Abnet CC, Arnold M and Wei WQ:
Epidemiology of esophageal squamous cell carcinoma.
Gastroenterology. 154:360–373. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Zhang C, Wang L, Yang J, Fu Y, Li H, Xie L
and Cui Y: MicroRNA-33a-5p suppresses esophageal squamous cell
carcinoma progression via regulation of lncRNA DANCR and ZEB1. Eur
J Pharmacol. 861:1725902019. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Wang L, Yu X, Zhang Z, Pang L, Xu J, Jiang
J, Liang W, Chai Y, Hou J and Li F: Linc-ROR promotes esophageal
squamous cell carcinoma progression through the derepression of
SOX9. J Exp Clin Cancer Res. 36:1822017. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Molina JR, Yang P, Cassivi SD, Schild SE
and Adjei AA: Non-small cell lung cancer: #pidemiology, risk
factors, treatment, and survivorship. Mayo Clin Proc. 83:584–594.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Du M, Zhang Y, Mao Y, Mou J, Zhao J, Xue
Q, Wang D, Huang J, Gao S and Gao Y: MiR-33a suppresses
proliferation of NSCLC cells via targeting METTL3 mRNA. Biochem
Biophys Res Commun. 482:582–589. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Kang M, Li Y, Zhao Y, He S and Shi J:
miR-33a inhibits cell proliferation and invasion by targeting CAND1
in lung cancer. Clin Transl Oncol. 20:457–466. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Amaar YG and Reeves ME: RASSF1C regulates
miR-33a and EMT marker gene expression in lung cancer cells.
Oncotarget. 10:123–132. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Yang L, Yang J, Li J, Shen X, Le Y, Zhou
C, Wang S, Zhang S, Xu D and Gong Z: MircoRNA-33a inhibits
epithelial-to- mesenchymal transition and metastasis and could be a
prognostic marker in non-small cell lung cancer. Sci Rep.
5:136772015. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Kang J, Kim W, Lee S, Kwon D, Chun J, Son
B, Kim E, Lee JM, Youn H and Youn B: TFAP2C promotes lung
tumorigenesis and aggressiveness through miR-183- and
miR-33a-mediated cell cycle regulation. Oncogene. 36:1585–1596.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Wang Y, Zhao W and Zhang S: STAT3-induced
upregulation of circCCDC66 facilitates the progression of non-small
cell lung cancer by targeting miR-33a-5p/KPNA4 axis. Biomed
Pharmacother. 126:1100192020. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Li YJ, Sun YX, Hao RM, Wu P, Zhang LJ, Ma
X, Ma Y, Wang PY, Xie N, Xie SY and Chen W: miR-33a-5p enhances the
sensitivity of lung adenocarcinoma cells to celastrol by regulating
mTOR signaling. Int J Oncol. 52:1328–1338. 2018.PubMed/NCBI
|
|
72
|
Kuo PL, Liao SH, Hung JY, Huang MS and Hsu
YL: MicroRNA-33a functions as a bone metastasis suppressor in lung
cancer by targeting parathyroid hormone related protein. Biochim
Biophys Acta. 1830:3756–3766. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Pan J, Fang S, Tian H, Zhou C, Zhao X,
Tian H, He J, Shen W, Meng X, Jin X and Gong Z: lncRNA
JPX/miR-33a-5p/Twist1 axis regulates tumorigenesis and metastasis
of lung cancer by activating Wnt/β-catenin signaling. Mol Cancer.
19:92020. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Rice SJ, Lai SC, Wood LW, Helsley KR,
Runkle EA, Winslow MM and Mu D: MicroRNA-33a mediates the
regulation of high mobility group AT-hook 2 gene (HMGA2) by thyroid
transcription factor 1 (TTF-1/NKX2-1). J Biol Chem.
288:16348–16360. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Zhu C, Zhao Y, Zhang Z, Ni Y, Li X and
Yong H: MicroRNA-33a inhibits lung cancer cell proliferation and
invasion by regulating the expression of β-catenin. Mol Med Rep.
11:3647–3651. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Boldrini L, Giordano M, Niccoli C, Melfi
F, Lucchi M, Mussi A and Fontanini G: Role of microRNA-33a in
regulating the expression of PD-1 in lung adenocarcinoma. Cancer
Cell Int. 17:1052017. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Li M, Zhao X, Liu Y, An J, Xiao H and Wang
C: Aberrant expression of CDK8 regulates the malignant phenotype
and associated with poor prognosis in human laryngeal squamous cell
carcinoma. Eur Arch Otorhinolaryngol. 274:2205–2213. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Marioni G, Marchese-Ragona R, Cartei G,
Marchese F and Staffieri A: Current opinion in diagnosis and
treatment of laryngeal carcinoma. Cancer Treat Rev. 32:504–515.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Karatas OF: Antiproliferative potential of
miR-33a in laryngeal cancer Hep-2 cells via targeting PIM1. Head
Neck. 40:2455–2461. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Karatas OF, Oner M, Abay A and Diyapoglu
A: MicroRNAs in human tongue squamous cell carcinoma: From
pathogenesis to therapeutic implications. Oral Oncol. 67:124–130.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Zuo Z, Ma L, Gong Z, Xue L and Wang Q:
Long non-coding RNA CASC15 promotes tongue squamous carcinoma
progression through targeting miR-33a-5p. Environ Sci Pollut Res
Int. 25:22205–22212. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Trotter SC, Sroa N, Winkelmann RR, Olencki
T and Bechtel M: A global review of melanoma follow-up guidelines.
J Clin Aesthet Dermatol. 6:18–26. 2013.PubMed/NCBI
|
|
83
|
Zhou J, Xu D, Xie H, Tang J, Liu R, Li J,
Wang S, Chen X, Su J, Zhou X, et al: miR-33a functions as a tumor
suppressor in melanoma by targeting HIF-1α. Cancer Biol Ther.
16:846–855. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Dimitriou F, Krattinger R, Ramelyte E,
Barysch MJ, Micaletto S, Dummer R and Goldinger SM: The world of
melanoma: Epidemiologic, genetic, and anatomic differences of
melanoma across the globe. Curr Oncol Rep. 20:872018. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Cao K, Li J, Chen J, Qian L, Wang A, Chen
X, Xiong W, Tang J, Tang S, Chen Y, et al: microRNA-33a-5p
increases radiosensitivity by inhibiting glycolysis in melanoma.
Oncotarget. 8:83660–83672. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Tian F, Wei H, Tian H, Qiu Y and Xu J:
miR-33a is downregulated in melanoma cells and modulates cell
proliferation by targeting PCTAIRE1. Oncol Lett. 11:2741–2746.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Lv L, Jia JQ and Chen J: The lncRNA CCAT1
upregulates proliferation and invasion in melanoma cells via
suppressing miR-33a. Oncol Res. 26:201–208. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Fu Y, Bi Y, Wang F, Chen X and Liu H:
Declination of long noncoding RNA paternally expressed gene 10
inhibits A375 cells proliferation, migration, and invasion via
mediating microRNA-33a. J Cell Biochem. 120:19868–19877. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Oltean MA, Matuz R, Sitar-Taut A, Mihailov
A, Rednic N, Tantau A, Toganel R, Minciuna IA, Orasan O, Muresan F
and Cozma A: Renal cell carcinoma with extensive tumor thrombus
into the inferior vena cava and right atrium in a 70-year-old man.
Am J Mens Health. 13:15579883198464042019. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Capitanio U, Bensalah K, Bex A, Boorjian
SA, Bray F, Coleman J, Gore JL, Sun M, Wood C and Russo P:
Epidemiology of renal cell carcinoma. Eur Urol. 75:74–84. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Jiang K, Sun F, Zhu J, Luo G, Ban Y and
Zhang P: miR-33a inhibits cell growth in renal cancer by
downregulation of MDM4 expression. Mol Genet Genomic Med.
7:e8332019. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Kim J, Gosnell JE and Roman SA: Geographic
influences in the global rise of thyroid cancer. Nat Rev
Endocrinol. 16:17–29. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Takeshita H, Shiozaki A, Bai XH, Iitaka D,
Kim H, Yang BB, Keshavjee S and Liu M: XB130, a new adaptor
protein, regulates expression of tumor suppressive microRNAs in
cancer cells. PLoS One. 8:e590572013. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Awad AJ, Burns TC, Zhang Y and Abounader
R: Targeting MET for glioma therapy. Neurosurg Focus. 37:E102014.
View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Chang M, Qiao L, Li B, Wang J, Zhang G,
Shi W, Liu Z, Gu N, Di Z, Wang X and Tian Y: Suppression of SIRT6
by miR-33a facilitates tumor growth of glioma through apoptosis and
oxidative stress resistance. Oncol Rep. 38:1251–1258. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Wang H, Sun T, Hu J, Zhang R, Rao Y, Wang
S, Chen R, McLendon RE, Friedman AH, Keir ST, et al: miR-33a
promotes glioma-initiating cell self-renewal via PKA and NOTCH
pathways. J Clin Invest. 124:4489–4502. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Ottaviani G and Jaffe N: The epidemiology
of osteosarcoma. Cancer Treat Res. 152:3–13. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Mirabello L, Troisi RJ and Savage SA:
Osteosarcoma incidence and survival rates from 1973 to 2004: Data
from the surveillance, epidemiology, and end results program.
Cancer. 115:1531–1543. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Jiang N, Wang X, Xie X, Liao Y, Liu N, Liu
J, Miao N, Shen J and Peng T: lncRNA DANCR promotes tumor
progression and cancer stemness features in osteosarcoma by
upregulating AXL via miR-33a-5p inhibition. Cancer Lett. 405:46–55.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Huang Y, Zhang J, Shao H, Liu J, Jin M,
Chen J and Zhao H: miR-33a mediates the anti-tumor effect of
lovastatin in osteosarcoma by targeting CYR61. Cell Physiol
Biochem. 51:938–948. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Zhang J, Wang D, Xiong J, Chen L and Huang
J: MicroRNA-33a-5p suppresses growth of osteosarcoma cells and is
downregulated in human osteosarcoma. Oncol Lett. 10:2135–2141.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Zhou Y, Huang Z, Wu S, Zang X, Liu M and
Shi J: miR-33a is up-regulated in chemoresistant osteosarcoma and
promotes osteosarcoma cell resistance to cisplatin by
down-regulating TWIST. J Exp Clin Cancer Res. 33:122014. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Gaudreau PO, Stagg J, Soulières D and Saad
F: The present and future of biomarkers in prostate cancer:
Proteomics, genomics, and immunology advancements. Biomark Cancer.
8 (Suppl 2):S15–S33. 2016.
|
|
104
|
Nelson WG, DeWeese TL and DeMarzo AM: The
diet, prostate inflammation, and the development of prostate
cancer. Cancer Metastasis Rev. 21:3–16. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Marshall JR: Diet and prostate cancer
prevention. World J Urol. 30:157–165. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Karatas OF, Wang J, Shao L, Ozen M, Zhang
Y, Creighton CJ and Ittmann M: miR-33a is a tumor suppressor
microRNA that is decreased in prostate cancer. Oncotarget.
8:60243–60256. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Li Q, Lu S, Li X, Hou G, Yan L, Zhang W
and Qiao B: Biological function and mechanism of miR-33a in
prostate cancer survival and metastasis: Via downregulating
Engrailed-2. Clin Transl Oncol. 19:562–570. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Dai Y, Wu Z, Lang C, Zhang X, He S, Yang
Q, Guo W, Lai Y, Du H, Peng X and Ren D: Copy number gain of ZEB1
mediates a double-negative feedback loop with miR-33a-5p that
regulates EMT and bone metastasis of prostate cancer dependent on
TGF-β signaling. Theranostics. 9:6063–6079. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Hong W and Dong E: The past, present and
future of breast cancer research in China. Cancer Lett. 351:1–5.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Shield KD, Soerjomataram I and Rehm J:
Alcohol use and breast cancer: A critical review. Alcohol Clin Exp
Res. 40:1166–1181. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Liang J and Shang Y: Estrogen and cancer.
Annu Rev Physiol. 75:225–240. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Zhang C, Zhang Y, Ding W, Lin Y, Huang Z
and Luo Q: MiR-33a suppresses breast cancer cell proliferation and
metastasis by targeting ADAM9 and ROS1. Protein Cell. 6:881–889.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Wolfe AR, Bambhroliya A, Reddy JP, Debeb
BG, Huo L, Larson R, Li L, Ueno NT and Woodward WA: MiR-33a
decreases high-density lipoprotein-induced radiation sensitivity in
breast cancer. Int J Radiat Oncol Biol Phys. 95:791–799. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Zeng W, Zuo G, Cao X and Li W: MiR-33a
functions as a tumor suppressor in triple-negative breast cancer by
targeting EZH2. Cancer Cell Int. 20:852020. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Guan X, Gu S, Yuan M, Zheng X and Wu J:
MicroRNA-33a-5p overexpression sensitizes triple-negative breast
cancer to doxorubicin by inhibiting eIF5A2 and
epithelial-mesenchymal transition. Oncol Lett. 18:5986–5994.
2019.PubMed/NCBI
|
|
116
|
Jang JS, Jeon HS, Sun Z, Aubry MC, Tang H,
Park CH, Rakhshan F, Schultz DA, Kolbert CP, Lupu R, et al:
Increased miR-708 expression in NSCLC and its association with poor
survival in lung adenocarcinoma from never smokers. Clin Cancer
Res. 18:3658–3667. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Wu X, Liu T, Fang O, Dong W, Zhang F,
Leach L, Hu X and Luo Z: MicroRNA-708-5p acts as a therapeutic
agent against metastatic lung cancer. Oncotarget. 7:2417–2432.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Morin PJ, Sparks AB, Korinek V, Barker N,
Clevers H, Vogelstein B and Kinzler KW: Activation of
beta-catenin-Tcf signaling in colon cancer by mutations in
beta-catenin or APC. Science. 275:1787–1790. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Zheng S, Zhang X, Wang X and Li J: MIR31HG
promotes cell proliferation and invasion by activating the
Wnt/β-catenin signaling pathway in non-small cell lung cancer.
Oncol Lett. 17:221–229. 2019.PubMed/NCBI
|
|
120
|
Zheng M, Jiang YP, Chen W, Li KD, Liu X,
Gao SY, Feng H, Wang SS, Jiang J, Ma XR, et al: Snail and Slug
collaborate on EMT and tumor metastasis through miR-101-mediated
EZH2 axis in oral tongue squamous cell carcinoma. Oncotarget.
6:6797–6810. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Buehler D, Hardin H, Shan W,
Montemayor-Garcia C, Rush PS, Asioli S, Chen H and Lloyd RV:
Expression of epithelial-mesenchymal transition regulators SNAI2
and TWIST1 in thyroid carcinomas. Mod Pathol. 26:54–61. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Cao YW, Wan GX, Sun JP, Cui XB, Hu JM,
Liang WH, Zheng YQ, Li WQ and Li F: Implications of the
Notch1-Snail/Slug-epithelial to mesenchymal transition axis for
lymph node metastasis in infiltrating ductal carcinoma. Kaohsiung J
Med Sci. 31:70–76. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Carreras-Torres R, Johansson M, Gaborieau
V, Haycock PC, Wade KH, Relton CL, Martin RM, Davey Smith G and
Brennan P: The role of obesity, type 2 diabetes, and metabolic
factors in pancreatic cancer: A mendelian randomization study. J
Natl Cancer Inst. 109:djx0122017. View Article : Google Scholar
|
|
124
|
Ercin M, Sancar-Bas S, Bolkent S and
Gezginci-Oktayoglu S: Tub and β-catenin play a key role in insulin
and leptin resistance-induced pancreatic beta-cell differentiation.
Biochim Biophys Acta Mol Cell Res. 1865:1934–1944. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Wijesekara N, Zhang LH, Kang MH, Abraham
T, Bhattacharjee A, Warnock GL, Verchere CB and Hayden MR: miR-33a
modulates ABCA1 expression, cholesterol accumulation, and insulin
secretion in pancreatic islets. Diabetes. 61:653–658. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Lekmine F, Uddin S, Sassano A, Parmar S,
Brachmann SM, Majchrzak B, Sonenberg N, Hay N, Fish EN and
Platanias LC: Activation of the p70 S6 kinase and phosphorylation
of the 4E-BP1 repressor of mRNA translation by type I interferons.
J Biol Chem. 278:27772–27780. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Xia P and Xu XY: PI3K/Akt/mTOR signaling
pathway in cancer stem cells: From basic research to clinical
application. Am J Cancer Res. 5:1602–1609. 2015.PubMed/NCBI
|
|
128
|
Yu JS and Cui W: Proliferation, survival
and metabolism: The role of PI3K/AKT/mTOR signalling in
pluripotency and cell fate determination. Development.
143:3050–3060. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Yip PY: Phosphatidylinositol
3-kinase-AKT-mammalian target of rapamycin (PI3K-Akt-mTOR)
signaling pathway in non-small cell lung cancer. Transl Lung Cancer
Res. 4:165–176. 2015.PubMed/NCBI
|
|
130
|
Fournier PG, Juárez P, Jiang G, Clines GA,
Niewolna M, Kim HS, Walton HW, Peng XH, Liu Y, Mohammad KS, et al:
The TGF-β signaling regulator PMEPA1 suppresses prostate cancer
metastases to bone. Cancer Cell. 27:809–821. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Feng J, Yan PF, Zhao HY, Zhang FC, Zhao WH
and Feng M: SIRT6 suppresses glioma cell growth via induction of
apoptosis, inhibition of oxidative stress and suppression of
JAK2/STAT3 signaling pathway activation. Oncol Rep. 35:1395–1402.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Liu SC, Huang CM, Bamodu OA, Lin CS, Liu
BL, Tzeng YM, Tsai JT, Lee WH and Chen TM: Ovatodiolide suppresses
nasopharyngeal cancer by targeting stem cell-like population,
inducing apoptosis, inhibiting EMT and dysregulating JAK/STAT
signaling pathway. Phytomedicine. 56:269–278. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Liu K, Tian T, Zheng Y, Zhou L, Dai C,
Wang M, Lin S, Deng Y, Hao Q, Zhai Z and Dai Z: Scutellarin
inhibits proliferation and invasion of hepatocellular carcinoma
cells via down-regulation of JAK2/STAT3 pathway. J Cell Mol Med.
23:3040–3044. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Tang J, Xu J, Zhi Z, Wang X, Wang Y, Zhou
Y and Chen R: MiR-876-3p targets KIF20A to block JAK2/STAT3 pathway
in glioma. Am J Transl Res. 11:4957–4966. 2019.PubMed/NCBI
|
|
135
|
Tanabe K, Kozawa O and Iida H: cAMP/PKA
enhances interleukin-1β-induced interleukin-6 synthesis through
STAT3 in glial cells. Cell Signal. 28:19–24. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Liu M, Inoue K, Leng T, Guo S and Xiong
ZG: TRPM7 channels regulate glioma stem cell through STAT3 and
Notch signaling pathways. Cell Signal. 26:2773–2781. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Xie RT, Cong XL, Zhong XM, Luo P, Yang HQ,
Lu GX, Luo P, Chang ZY, Sun R, Wu TM, et al: MicroRNA-33a
downregulation is associated with tumorigenesis and poor prognosis
in patients with hepatocellular carcinoma. Oncol Lett.
15:4571–4577. 2018.PubMed/NCBI
|
|
138
|
Hou LK, Ma YS, Han Y, Lu GX, Luo P, Chang
ZY, Xie RT, Yang HQ, Chai L, Cai MX, et al: Association of
microRNA-33a molecular signature with non-small cell lung cancer
diagnosis and prognosis after chemotherapy. PLoS One.
12:e01704312017. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Pan J, Zhou C, Zhao X, He J, Tian H, Shen
W, Han Y, Chen J, Fang S, Meng X, et al: A two-miRNA signature
(miR-33a-5p and miR-128-3p) in whole blood as potential biomarker
for early diagnosis of lung cancer. Sci Rep. 8:166992018.
View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Kandimalla R, Gao F, Matsuyama T, Ishikawa
T, Uetake H, Takahashi N, Yamada Y, Becerra C, Kopetz S, Wang X and
Goel A: Genome-wide discovery and identification of a novel miRNA
signature for recurrence prediction in stage II and III colorectal
cancer. Clin Cancer Res. 24:3867–3877. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Ma Y, Zhou G, Li M, Hu D, Zhang L, Liu P
and Lin K: Long noncoding RNA DANCR mediates cisplatin resistance
in glioma cells via activating AXL/PI3K/Akt/NF-κB signaling
pathway. Neurochem Int. 118:233–241. 2018. View Article : Google Scholar : PubMed/NCBI
|














