Introduction
Glioma is one of the most common and aggressive
primary malignancies of the brain, and the global incidence was 5%
in 2013; it was reported to account for ~80% of malignant neoplasia
and ~30-40% of primary tumors in the central nervous system,
worldwide (1). Despite recent
advances in adjuvant radiotherapy, hormone therapy, chemotherapy
and targeted biologic therapy, the overall survival outcomes of
most patients with glioma remain poor (2,3).
Furthermore, the aforementioned treatments exhibit side effects,
such as anemia and constipation (3);
thus, the development of novel therapeutic strategies against
glioma remains essential.
Long non-coding RNAs (lncRNAs) are a novel class of
non-protein coding RNAs, >200 nt long (4–6). lncRNAs
are involved in a number of biological processes, including cell
differentiation and gene expression (7). Increasing evidence suggests that
lncRNAs are associated with the initiation and progression of
different types of tumor, including glioma (8–15).
Furthermore, aberrant expression levels of lncRNAs have been
reported in tumor cells, indicating the potential role of lncRNAs
in tumorigenesis (16–18). Thus, investigation of the regulatory
functions of lncRNAs in cancer may help facilitate the development
of effective therapies.
lncRNA NBAT1 is involved in carcinogenesis, and
previous studies have reported the dysregulation of NBAT1 in
different types of cancer (19,20).
NBAT1 promotes the initiation and development of tumors via several
pathways, including PRC2 signaling (21,22), and
thus is considered a promising biomarker and prognostic indicator
of glioma (20,23). However, the potential molecular
targets of NBAT1 in glioma have not yet been fully
investigated.
MicroRNAs (miRNAs/miRs) are endogenously expressed
non-coding RNAs that are ~22 nt in length and are considered
potential targets of lncRNAs (24,25).
Increasing evidence suggests that the levels of miRNAs are
dysregulated in cancer, which may contribute to tumor development
(26,27). In addition, miRNAs such as miR-140
and miR-152 are able to interact with lncRNAs in glioma (28,29).
However, the specific functions of miRNAs in glioma remain
unclear.
The present study aimed to investigate the functions
of the NBAT1-mediated miR-21/SOX7 signaling pathway in glioma cell
proliferation and metastasis, which may provide novel insight for
the effective treatment of patients with glioma.
Materials and methods
Patient samples
A total of 40 pairs of primary tumor and
paracarcinoma tissue samples (>5 cm from tumor margin) were
collected from patients with glioma (16 men and 24 women; age
range, 45–65 years; mean age, 51 years) undergoing surgical
resection at The First Affiliated Hospital of Jinzhou Medical
University (Jinzhou, China) between June 2015 and August 2017.
Prior to surgery, none of the patients have received chemotherapy
or radiotherapy. Metastasis was observed in 14 and absent in 26
patients, and 22 cases were diagnosed with grade I or II glioma,
while 18 were diagnosed with grade III or IV glioma, according to
the World Health Organization grading system (https://www.cancer.net/cancer-types/brain-tumor/grades-and-prognostic-factors).
The biopsies were examined by two independent pathologists from the
Department of Histopathology, The First Affiliated Hospital of
Jinzhou Medical University (Jinzhou, China). All samples were
immediately snap-frozen in liquid nitrogen and stored at −80°C
until further experimentation. The present study was approved by
the Institutional Review Board of The First Affiliated Hospital of
Jinzhou Medical University (approval no. JYD159302; Jinzhou,
China). Written informed consent was provided by all patients prior
to the study start.
Cell culture
The human glioma cell lines (AM38, Gli-6, GSC11 and
A172) and normal human astrocyte cell line (A735) were purchased
from the American Type Culture Collection. Cells were maintained in
Dulbecco's Modified Eagle's Medium (DMEM) supplemented with 10%
fetal bovine serum (FBS), 100 U/ml penicillin and 100 µg/ml
streptomycin (all purchased from HyClone; Cytiva) at 37°C in a
humidified incubator containing 5% CO2.
Cell transfection
To establish the NBAT1 knockdown model, short
hairpin (sh)RNA sequences targeting NBAT1 (sh-NBAT1), as well as
the negative control (sh-NC) were synthesized by Shanghai GenePharm
Co., Ltd. Following annealing, the shRNAs were integrated into the
lentiviral pU6-Luc-Puro vector (Shanghai GenePharm Co., Ltd.). To
establish the NBAT1 overexpression model, wild-type (WT; o/e-NBAT1)
or mutant (o/e-NC) NBAT1 fragments were amplified by PCR using Taq
polymerase (Thermo Fisher Scientific, Inc.) and subcloned into the
pcDNA3.1 vector (Invitrogen; Thermo Fisher Scientific, Inc.). The
primer sequences used were as follows: Forward,
5′-TCAGCAGAAACGGCACGAT-3′ and reverse 5′-AGATGACCCAGGCACCTCC-3′.
The following thermocycling conditions were used: 95°C for 30 sec,
followed by 30 cycles at 95°C for 15 sec, 60°C for 20 sec, 68°C for
10 sec and a final extension at 68°C for 5 min.
A172 and AM38 cells were seeded into 6-well plates
at a density of 2×105 and subsequently transfected with
the recombinant lentiviral vectors (50 nM) using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) for 8 h at 37°C. The culture medium was replaced
with fresh DMEM supplemented with 10% FBS 8 h post-transfection.
Knockdown and overexpression of NBAT1 were confirmed by reverse
transcription-quantitative (RT-q)PCR analysis.
The miR-21 mimics or inhibitors and the
corresponding NCs (all 40 pg/µl) were purchased from Shanghai
GenePharm Co., Ltd. and transfected into glioma cells
(2×105) using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) for 8 h at 37°C. The
culture medium was replaced with fresh DMEM supplemented with 10%
FBS at 8 h post-transfection. Subsequent experiments were performed
24 h post-transfection.
RT-qPCR
miRNA was extracted from tissues or cell lines using
the miRNeasy Mini kit (Qiagen China Co., Ltd.), and miR-21
expression was determined using the TaqMan MicroRNA Assay kit (cat.
no. 4427975; Applied Biosystems; Thermo Fisher Scientific, Inc.).
qPCR was subsequently performed using an Applied Biosystem 7500 PCR
instrument (Applied Biosystems; Thermo Fisher Scientific, Inc.).
The following thermocycling conditions were used for qPCR: 95°C for
10 min, followed by 40 cycles at 95°C for 15 sec and 60°C for 1
min. U6 small nuclear RNA was used as the internal control.
Total RNA was extracted from clinical samples or
cell lines using TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocol.
RNA concentration was determined using a NanoDrop 1000
spectrophotometer (Thermo Fisher Scientific, Inc.) and subsequently
reverse-transcribed into cDNA using the PrimeScript™ RT
kit (Takara Biotechnology Co., Ltd.). The temperature protocol was
as follows: 42°C for 45 min, 99°C for 5 min and 5°C for 5 min. qPCR
was performed using the SYBR® Green PCR Master Mix
(Takara Biotechnology Co., Ltd.), according to the manufacturer's
protocol. The following primer sequences were used for qPCR: NBAT1
forward, 5′-ATTTCTGCTCCTGGGTCTTAC-3′ and reverse,
5′-GCAAGAGCACAAGAGGAAGA-3′; SOX7 forward,
5′-CAAGGACGAGAGGAAACGGC-3′ and reverse, 5′-TACGGCCTCTTCTGGGACAG-3′;
and GAPDH forward, 5′-GCAAGAGCACAAGAGGAAGA-3′ and reverse,
5′-ACTGTGAGGAGGGGAGATTC-3′. The following thermocycling conditions
were used for qPCR: 95°C for 5 min, followed by 45 cycles at 95°C
for 15 sec, 60°C for 20 sec and 72°C for 10 sec. Relative mRNA
expression levels were calculated using the 2−∆∆Cq
method (30) and normalized to the
internal reference gene GAPDH.
Western blotting
Total protein was extracted from tissues or cells
using radioimmunoprecipitation assay buffer (Beyotime Institute of
Biotechnology). Protein concentration was determined using a
bicinchoninic acid assay kit (Beyotime Institute of Biotechnology),
and 40 µg protein/lane was separated via 10% SDS-PAGE. The
separated proteins were subsequently transferred onto
nitrocellulose membranes (EMD Millipore) and blocked with
tris-buffered saline (TBS) containing 5% skimmed milk for 2 h at
room temperature. The membranes were incubated with primary
antibodies against SOX7 (1:1,000; cat. no. MAB2766; Novus
Biologicals, Ltd.) and GAPDH (1:1,000; cat. no. sc-47724; Santa
Cruz Biotechnology Inc.) overnight at 4°C. Membranes were washed
three with TBS + Tween-20 (0.05%) and subsequently incubated with
horseradish peroxidase-conjugated anti-mouse IgG secondary antibody
(1:5,000; cat. no. sc-2371; Santa Cruz Biotechnology Inc.) for 1 h
at 37°C. Protein bands were visualized using an enhanced
chemiluminescence protein detection kit (Pierce; Thermo Fisher
Scientific, Inc.) and quantified via densitometric analysis using
ImageJ software (version 1.48; National Institutes of Health).
Cell counting kit-8 (CCK-8) assay
Transfected A172/AM38 cells were harvested 24 h
post-transfection and seeded into 96-well plates at a density of
1×104 cells/well. Briefly, 10 µl of CCK-8 solution
(Dojindo Molecular Technologies, Inc.) was added to the wells on
days 1, 2, 3 and 4 post-inoculation, according to the
manufacturer's protocol. Following incubation at 37°C for an
additional 2 h, cell proliferation was subsequently analyzed at a
wavelength of 450 nm using a microplate reader (Bio-Rad
Laboratories, Inc.).
Transwell assay
For the migration assay, a total of 1×105
A172/AM38 cells were plated in the upper chamber of Transwell
plates with a pore size of 8 µm (BD Biosciences) in FBS-free DMEM
medium. For the invasion assay, Transwell membranes were precoated
with Matrigel® (Sigma-Aldrich; Merck KGaA) at room
temperature for 48 h. DMEM medium (500 µl) supplemented with 10%
FBS was added to the lower chambers. Following overnight incubation
at 37°C, non-migratory/invasive cells were removed using a cotton
swab, whereas the migratory/invasive cells in the lower chambers
were fixed with 4% paraformaldehyde for 15 min and stained with
0.5% crystal violet for 20 min at room temperature. Stained cells
were counted in five randomly selected fields using an inverted
light microscope (magnification, ×100; Olympus Corporation).
Northern blot analysis
Total RNA was extracted from tissues or cell lines
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). An equal amount of RNA (25 µg) was loaded onto
15% TBE-urea gels and separated using a 15% urea-PAGE gel. The
separated proteins were subsequently transferred onto positively
charged nylon membranes (Cytiva) and cross-linked by UV
irradiation. The blots were hybridized by DIG-labelled probe for
miR-21 (cat. no. 339546; Qiagen AB) overnight at 42°C. The
membranes were washed three times with a low-stringency buffer
(2×SSC containing 0.1% SDS; Thermo Fisher Scientific, Inc.), and
the RNA levels were determined using the DIG Luminescent Detection
kit [Roche Diagnostics (Shanghai) Co., Ltd] and semi-quantified
using Multi Gauge software (version 3.0; Fujifilm Wako Pure
Chemical Corporation). U6 (cat. no. 339508; Qiagen) RNA was used as
the loading control.
Bioinformatics prediction and
dual-luciferase reporter assay
TargetScan (www.targetscan.org), miRanda (www.microrna.org/microrna) and LncBase Predicted v2
(http://carolina.imis.athenainnovation.gr/diana_tools/web/index.php?r=lncbasev2%2Findex-predicted)
databases were used to predict the potential binding sites of NBAT1
and SOX7 transcripts on miR-21.
WT fragments of the 3′-untranslated region (3′UTR)
of NBAT1 and SOX7 containing the potential binding sites of miR-21
were purchased from Shanghai GenePharma Co., Ltd. and cloned into
the pmirGLO Dual-Luciferase miRNA Target Expression vector (Promega
Corporation) according to the manufacturer's protocol. The
QuikChange Multi Site-Directed Mutagenesis kit (Agilent
Technologies, Inc.) was used to generate the NBAT1- and
SOX7-3′UTR-MUT reporter containing mutant miR-21 binding sites. The
constructed luciferase vectors were co-transfected with miR-21
mimics or miR-NC into DH5α competent cells (Thermo Fisher
Scientific, Inc.). Following incubation at 37°C for 48 h, firefly
and Renilla luciferase activities were detected using a Dual
Luciferase Reporter assay system (Promega Corporation) according to
the manufacturer's protocol. Firefly luciferase activity was
normalized to Renilla luciferase activity.
Statistical analysis
Statistical analysis was performed using SPSS
software 17.0 (SPSS, Inc.), and data are presented as the mean ±
standard deviation. Student's t-test (paired and unpaired) was used
to compare differences between two groups. One-way analysis of
variance followed by Student-Newman-Keuls or Tukey's post-hoc test
were performed to compare differences among multiple groups.
Pearson's correlation analysis was performed to determine the
correlations between mRNA expression levels. All experiments were
performed in triplicate. P<0.05 was considered to indicate a
statistically significant difference.
Results
NBAT1 is downregulated in glioma
tissues and cells and associated with poor prognosis
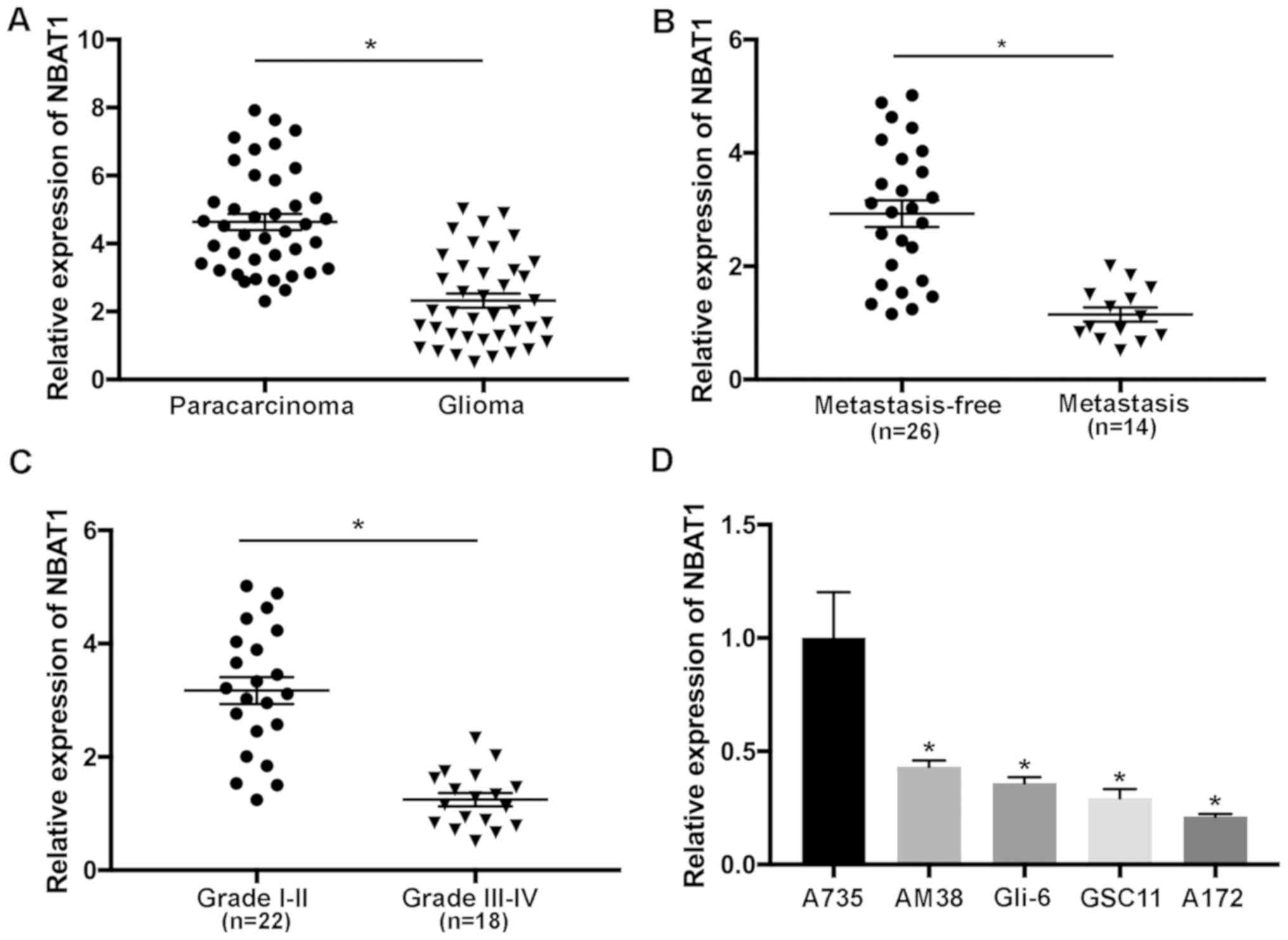
The expression levels of NBAT1 in 40 paired glioma
and paracarcinoma samples were determined using RT-qPCR analysis.
The results demonstrated that NBAT1 expression was significantly
downregulated in glioma tissues compared with that in the
paracarcinoma tissues (P<0.05; Fig.
1A). The effects of NBAT1 on metastasis of patients with glioma
were also investigated. The results revealed that NBAT1 expression
was significantly decreased in patients with metastatic glioma
compared with the controls (patients with non-metastatic glioma;
P<0.05; Fig. 1B). In addition,
NBAT1 expression was significantly decreased in aggressive (grade
III or IV) compared with low-grade (I or II) glioma, suggesting
that downregulation of NBAT1 was associated with the development of
glioma (P<0.05; Fig. 1C). NBAT1
expression was significantly downregulated in glioma cells compared
with A735 cells (P<0.05; Fig.
1D). Taken together, these results suggested that NBAT1
expression was downregulated in glioma, which may also be
associated with metastasis.
Overexpression of NBAT1 suppresses
glioma cell proliferation, migration and invasion
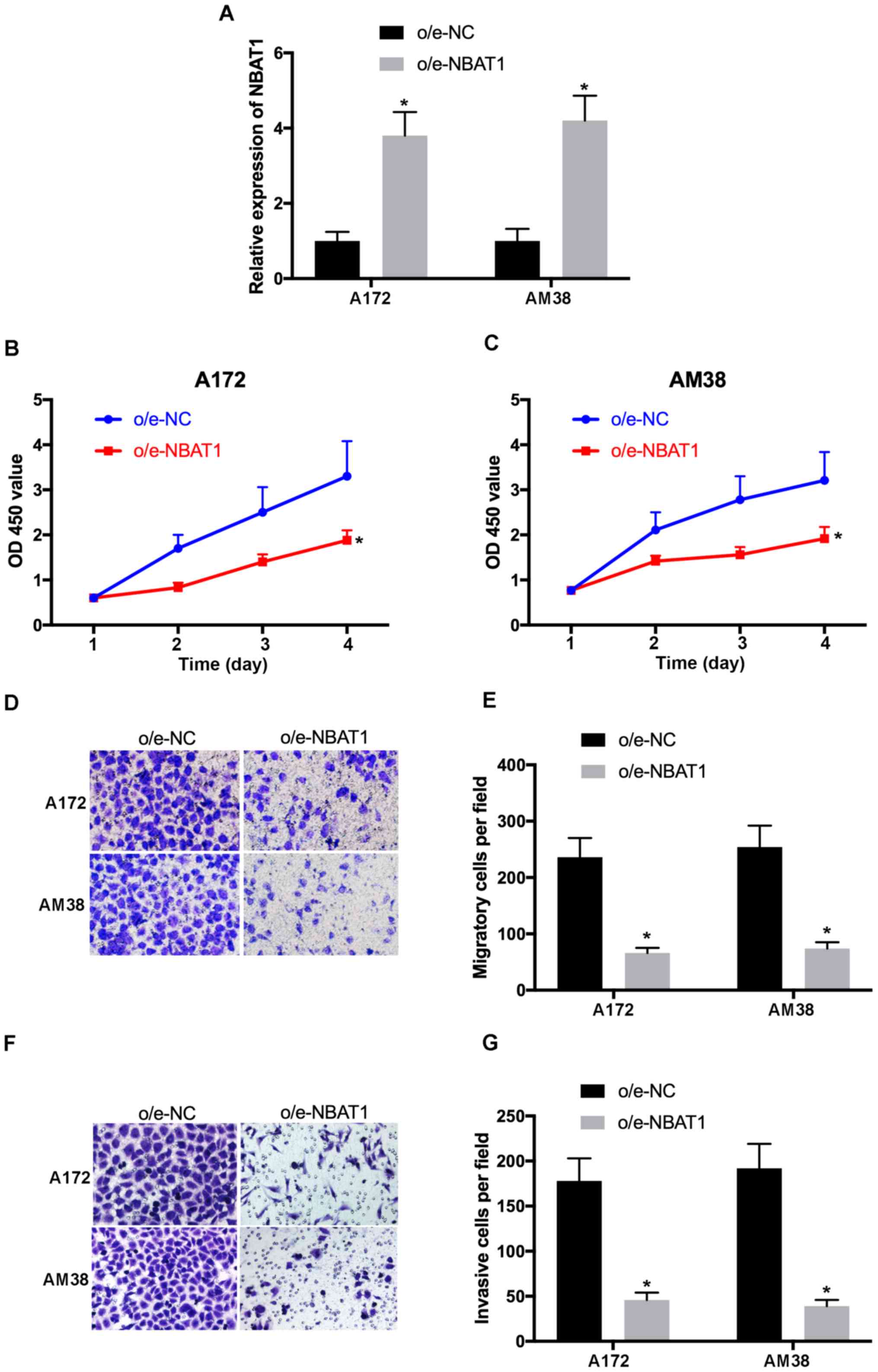
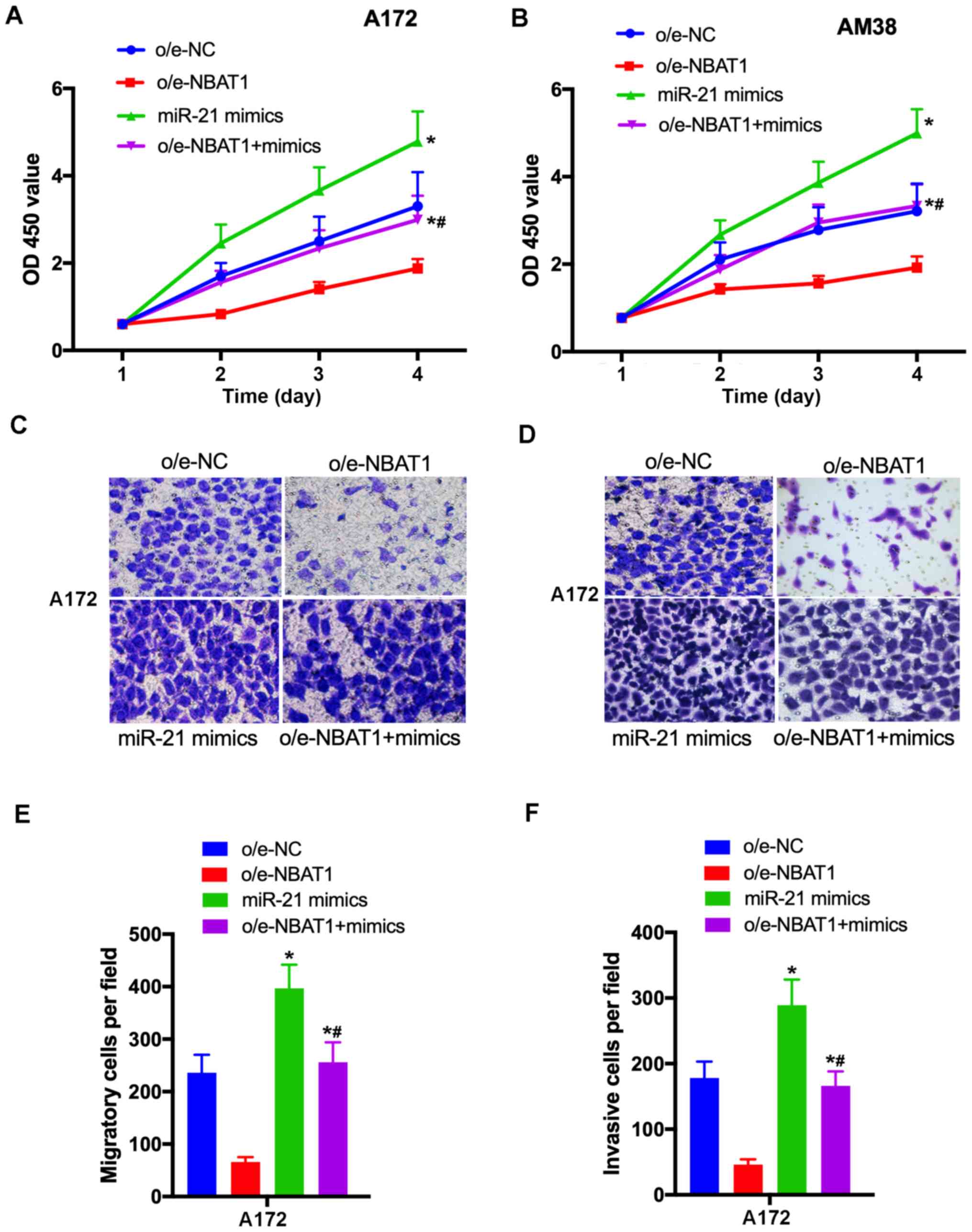
To determine the effects of NBAT1 on glioma cell
proliferation, migration and invasion, NBAT1 was overexpressed in
A172 and AM38 cells, and transfection efficiency was determined by
RT-qPCR (Fig. 2A). The CCK-8 assay
demonstrated that the proliferative ability of
o/e-NBAT1-transfected A172 and AM38 cells significantly decreased
compared with that of the control (P<0.05; Fig. 2B and C). In addition, the Transwell
assay results revealed that the migratory and invasive abilities of
o/e-NBAT1-transfected A172 and AM38 cells were significantly
inhibited compared with those of the control groups (P<0.05;
Fig. 2D-G). Collectively, these
results suggested that overexpression of NBAT1 may suppress the
proliferation, migration and invasion of glioma cells.
miR-21 is the potential target of
NBAT1 in glioma
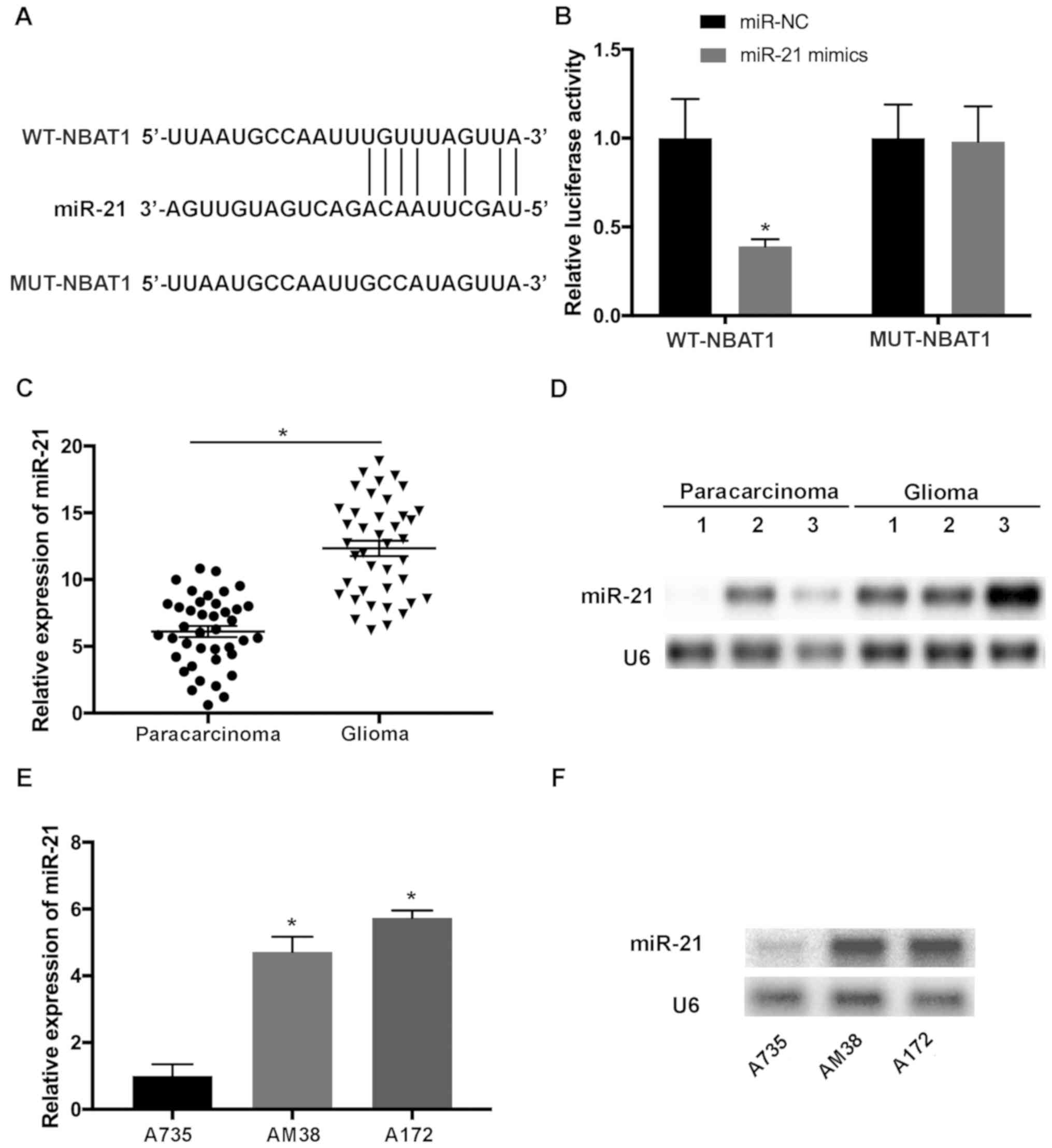
To determine whether NBAT1 functions by suppressing
its target miRNAs in glioma, the potential binding sites of miR-21
in NBAT1 transcripts were predicted using LncBase Predicted v2
(http://carolina.imis.athena-innovation.gr/diana_tools/web/index.php?r=lncbasev2%2Findex-predicted)
(Fig. 3A). Luciferase reporter
vectors containing WT-NBAT1 and MUT-NBAT1 sequences of predicted
miR-21 binding sites were constructed. The results demonstrated
that miR-21 mimics significantly attenuated the activity of the
luciferase plasmid containing the WT binding sites compared with
that of the MUT control (P<0.05; Fig.
3B). RT-qPCR and northern blot analyses indicated that miR-21
expression was upregulated in glioma tissues compared with that in
paracarcinoma tissues (P<0.05; Fig.
3C and D). In addition, upregulated miR-21 expression was
observed in A172 and AM38 cells compared with that in A735 cells
(P<0.05; Fig. 3E and F).
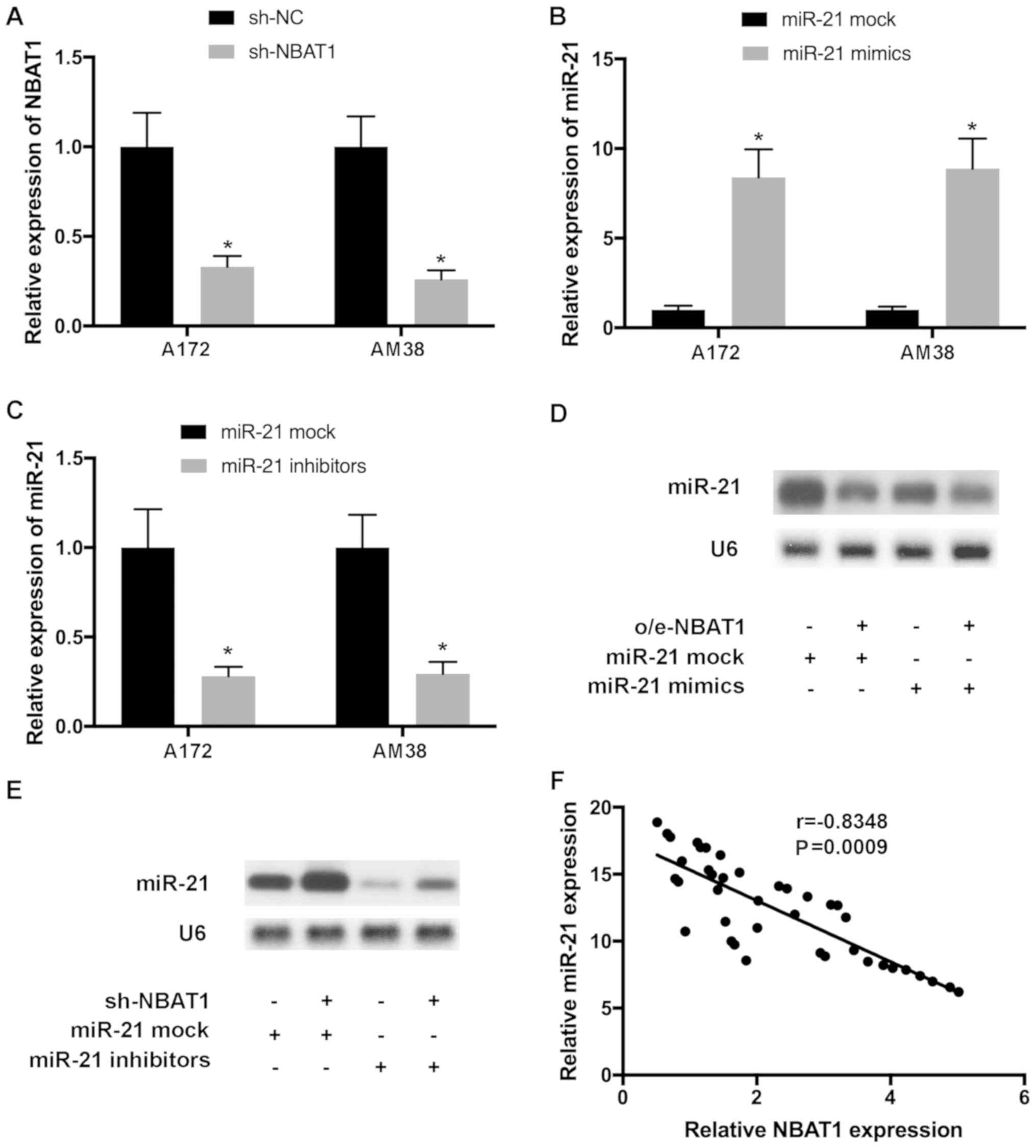
To further investigate the effects of NBAT1 on
miR-21 expression, glioma cells were transfected with o/e-NBAT1 and
miR-21 mimics, or with sh-NBAT1 and miR-21 inhibitors, and
transfection efficiency was determined by RT-qPCR (Fig. 4A-C). Northern blot analysis
demonstrated that upregulation of miR-21 expression by transfection
with miR-21 mimics was reversed following the addition of o/e-NBAT1
(Fig. 4D). Similarly, downregulation
of miR-21 expression by transfection with miR-21 inhibitors was
counteracted following co-transfection with sh-NBAT1 (Fig. 4E). Pearson's correlation analysis
demonstrated that the levels of miR-21 and NBAT1 were negatively
correlated in glioma tissues (Fig.
4F).
miR-21 serves important roles in the
proliferation and metastasis of glioma cells
The effects of miR-21 on the progression of glioma
were further assessed. The CCK-8 assay demonstrated that the
proliferative ability of glioma cells significantly increased
following transfection with the miR-21 mimics compared with the
vehicle only control (P<0.05; Fig. 5A
and B). In addition, the migratory and invasive abilities of
glioma cells were enhanced following transfection with the miR-21
mimics compared with the vehicle only control (P<0.05; Fig. 5C-F). Taken together, these results
suggest that miR-21 promoted glioma cell proliferation, migration
and invasion.
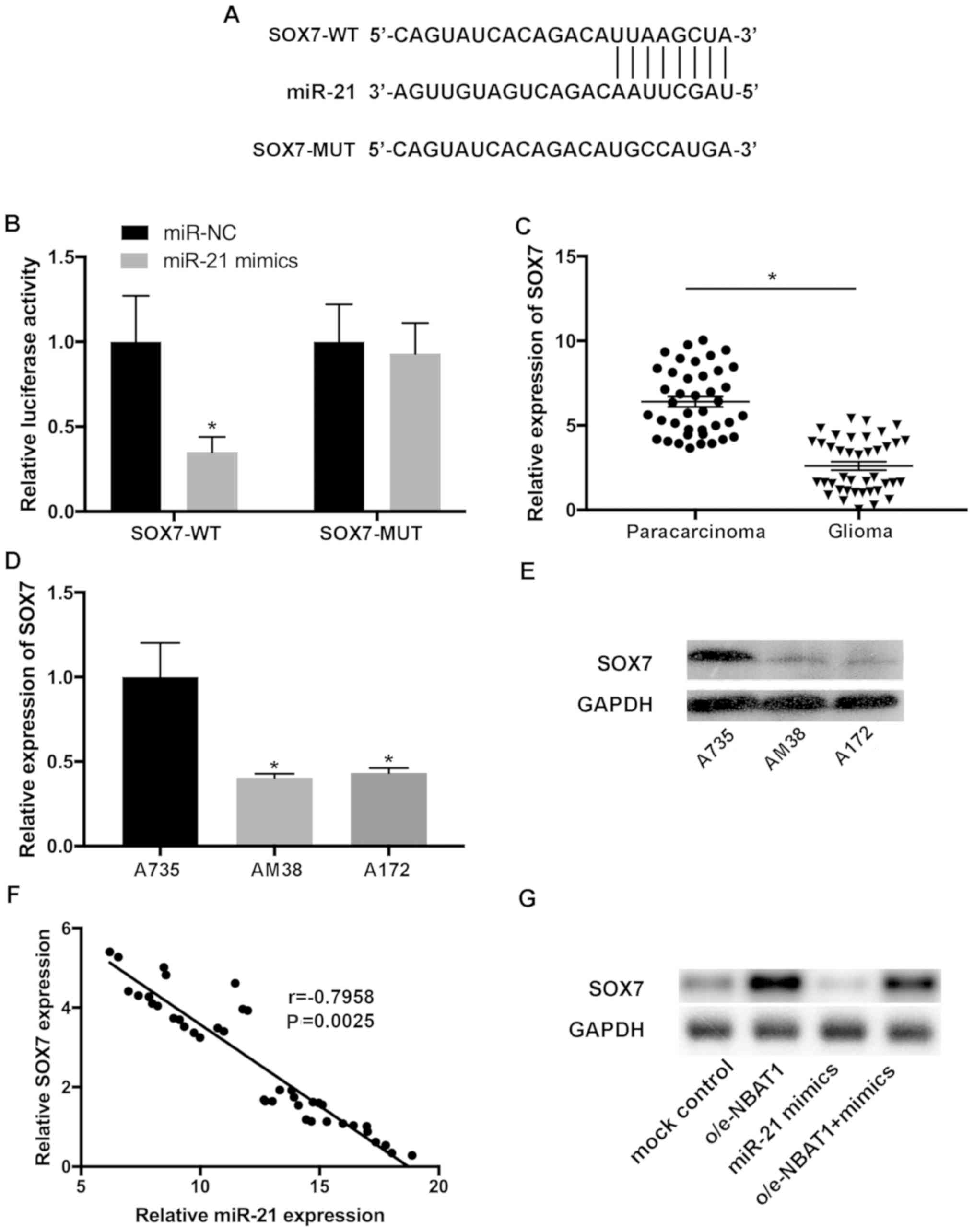
SOX7 is a downstream target of
miR-21
To identify the putative targets of miR-21, the
complementary sequence of miR-21 in SOX7 transcripts was predicted
(Fig. 6A). The interaction between
SOX7 and miR-21 was confirmed using a dual-luciferase assay.
Luciferase reporter plasmids containing SOX7-WT) and SOX7-MUT
fragments of the predicted miR-21 binding sites were constructed.
The results demonstrated that the miR-21 mimics significantly
decreased the activity of the luciferase vector containing the
SOX7-WT sequence, but not the mutant control (Fig. 6B). RT-qPCR analysis indicated that
SOX7 expression was downregulated in glioma tissues compared with
that in the paracarcinoma tissues (Fig.
6C). In addition, downregulated SOX7 expression was observed in
A172 and AM38 cells compared with that in A735 cells (P<0.05;
Fig. 6D and E). Pearson's
correlation analysis demonstrated that the levels of SOX7 and
miR-21 were inversely correlated in glioma samples (Fig. 6F).
To further investigate the effects of SOX7 on the
progression of glioma, the expression levels of SOX7 were assessed
in glioma cells transfected with control, o/e-NBAT1 or the miR-21
mimics, or co-transfected with o/e-NBAT1 and the miR-21 mimics.
SOX7 expression significantly decreased following transfection with
the miR-21 mimics compared with that in the control group, which
was reversed following overexpression of NBAT1 (Fig. 6G). Collectively, these results
suggested that SOX7 may be a novel target of miR-21, and that the
NBAT1/miR-21/SOX7 signaling pathway may be involved in the
progression of glioma.
Discussion
lncRNAs are a novel class of non-protein coding RNAs
that are >200 nt in length (4).
Previous studies on lncRNAs have reported their importance on the
pathogenesis of cancer, and increasing evidence indicates that
lncRNAs act as key factors in the regulation of cancer cells
(25–29). lncRNAs function as oncogenes or tumor
suppressors in cancer, and aberrant expression levels of lncRNAs
are associated with tumor development (31). For example, lncRNA PVT1 modulates the
cell proliferation, invasion and tumor growth in orthotopic
xenografts of breast cancer (32).
In addition, upregulated LINC01116 expression is associated with
unfavorable overall survival and metastasis of patients with breast
cancer (33). The expression levels
of lncRNA PlncRNA-1 are downregulated in cancer tissues; however,
restored PlncRNA-1 expression inhibits proliferation and promotes
apoptosis of breast cancer cells via the TGF-β1/phosphoglycerate
dehydrogenase axis (34).
Furthermore, downregulation of lncRNA SNHG5 suppresses
proliferation and migration of gastric cancer cells via the
miR-32/Kruppel-like factor 4 axis (35). These findings suggest that lncRNAs
are key regulators during the initiation and progression of cancer.
Consistent with previous findings, the results of the present study
demonstrated the key roles of NBAT1 on the proliferation and
metastasis of glioma cells.
A number of lncRNAs can regulate gene expression by
interacting with their target miRNAs directly. For example, lncRNA
BC032469 binds to miR-1207-5p and human telomerase reverse
transcriptase, and induces cancer cell proliferation (36). Furthermore, lncRNA H19 regulates the
proliferation, migration and invasion of gastric cancer cells
through its target miRNAs, such as miR-675 (37,38).
However, the functions and underlying molecular mechanisms of
lncRNAs remain largely unknown and require further investigation.
Similarly, the present study demonstrated the regulatory function
of miR-21 during the proliferation, migration and invasion of
glioma cells, and its underlying molecular mechanisms were
elucidated.
Increasing evidence suggests that aberrant NBAT1
expression is associated with various types of cancer, such as
ovarian cancer, breast cancer and osteosarcoma (19,21,22).
NBAT1 inhibits the proliferation and promotes apoptosis of ovarian
cancer cells (19). NBAT1 regulates
the migration and invasion of breast cancer cells by targeting
Dickkopf Wnt signaling pathway inhibitor 1/polycomb repressive
complex 2 signaling (21). A
previous study has reported that miR-21 is a potential target of
NBAT1 in osteosarcoma cells (22).
NBAT1 has also been demonstrated to be associated with the
proliferation and metastasis of glioma cells (20). Consistent with previous findings, the
results of the present study demonstrated that NBAT1 was
downregulated in glioma tissues and cells. Furthermore,
overexpression of NBAT1 suppressed glioma cell proliferation,
migration and invasion. However, the effects of sh-NBAT1 on the
biological behavior of glioma cells were not investigated in the
present study.
miRNAs may function as oncogenes or tumor
suppressors by modulating gene expression, and are considered to be
targets of lncRNAs (24,25). Previous studies have reported that
the expression levels of miRNAs are impaired in several types of
cancer, including glioma, and miR-140 and miR-152 interact with
lncRNAs in glioma cells (26–29). The
present study aimed to identify novel targets of NBAT1 in glioma.
The results of the present study suggested that NBAT1 was able to
bind with miR-21, the expression levels of which were notably
upregulated in glioma tissues and cells. Additionally, the
expression of miR-21 was reduced by NBAT1 in glioma cells, and
their expression levels were inversely correlated in glioma
tissues. Inhibition of miR-21 suppressed the proliferation,
migration and invasion of glioma cells. The results of the present
study were consistent with a previous study, which suggested that
NBAT1 inhibited the progression of osteosarcoma via miR-21
signaling (22).
SOX7 is a member of the SOX transcription factors
(39) and is considered to be an
essential regulator in cardiovascular development (40). SOX7 functions as a tumor suppressor
in endometrial cancer and as an oncogene in gastric cancer
(41–44). The positive association between SOX7
expression and distant metastasis-free survival has been reported
in prostate cancer and breast cancer (45,46). In
the present study, SOX7 was identified as a downstream target of
miR-21, and the results demonstrated that SOX7 expression levels
were decreased in glioma tissues and cells compared with those in
paracarcinoma tissues and normal astrocytes, respectively. Of note,
downregulated SOX7 expression induced by transfection with miR-21
mimics was reversed following overexpression of NBAT1.
Collectively, these results highlighted the key functions of
NBAT1/miR-21/SOX7 signaling on the progression of glioma. A major
limitation of the present study is that in vivo analysis was
not performed, thus prospective studies will focus on performing
in vivo experiments to validate these findings. Furthermore,
the expression levels of proliferation- and invasion-associated
molecules should be investigated to confirm the existing
findings.
In conclusion, the results of the present study
suggested that NBAT1 may act as a potential tumor suppressor by
upregulating SOX7 via suppressing miR-21, thus inhibiting glioma
cell proliferation, migration and invasion. These results indicate
the essential roles of NBAT1 and its underlying molecular
mechanisms in the proliferation, migration and invasion of glioma
cells, providing evidence on the potential functions of NBAT1 in
tumorigenesis, and suggest that the NBAT1/miR-21/SOX7 axis may be a
promising therapeutic target for the treatment of patients with
glioma.
Acknowledgements
Not applicable.
Funding
The present study was funded by the Outstanding
Talent Cultivation Foundation of Liaoning Province (grant no.
2015020349), the President Foundation of Jinzhou Medical University
(grant no. XZJJ20130206) and the Natural Science Foundation of
Liaoning Province (grant no. 2019-ZD-0824).
Availability of data and materials
The datasets generated or analyzed during the
present study are included in this published article.
Authors' contributions
WG conceived and designed the present study. NG, RW,
XF and CL performed the experiments and interpreted the data. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Institutional
Review Board of The First Affiliated Hospital of Jinzhou Medical
University (approval no. JYD159302; Jinzhou, China). Written
informed consent was provided by all patients for the use of
clinical tissue prior to the study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Lai NS, Wu DG, Fang XG, Lin YC, Chen SS,
Li ZB and Xu SS: Serum microRNA-210 as a potential noninvasive
biomarker for the diagnosis and prognosis of glioma. Br J Cancer.
112:1241–1246. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mansouri A, Mansouri S, Hachem LD,
Klironomos G, Vogelbaum MA, Bernstein M and Zadeh G: The role of
5-aminolevulinic acid in enhancing surgery for high-grade glioma,
its current boundaries, and future perspectives: A systematic
review. Cancer. 122:2469–2478. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Delgado-López PD and Corrales-García EM:
Survival in glioblastoma: A review on the treatment modalities.
Clin Transl Oncol. 18:1062–1071. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gibb EA, Brown CJ and Lam WL: The
functional role of long non-coding RNA in human carcinomas. Mol
Cancer. 10:382011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mills JD, Chen J, Kim WS, Waters PD,
Prabowo AS, Aronica E, Halliday GM and Janitz M: Long intervening
non-coding RNA 00320 is human brain-specific and highly expressed
in the cortical white matter. Neurogenetics. 16:201–213. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hsiao J, Yuan TY, Tsai MS, Lu CY, Lin YC,
Lee ML, Lin SW, Chang FC, Liu Pimentel H, Olive C, et al:
Upregulation of haploinsufficient gene expression in the brain by
targeting a long non-coding RNA improves seizure phenotype in a
model of dravet syndrome. EBioMedicine. 9:257–277. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li Z, Dong M, Fan D, Hou P, Li H, Liu L,
Lin C, Liu J, Su L, Wu L, et al: lncRNA ANCR down-regulation
promotes TGF-β-induced EMT and metastasis in breast cancer.
Oncotarget. 8:67329–67343. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang X, Sun S, Pu JK, Tsang AC, Lee D,
Man VO, Lui WM, Wong ST and Leung GK: Long non-coding RNA
expression profiles predict clinical phenotypes in glioma.
Neurobiol Dis. 48:1–8. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Grzmil M, Morin P Jr, Lino MM, Merlo A,
Frank S, Wang Y, Moncayo G and Hemmings BA: MAP kinase-interacting
kinase 1 regulates SMAD2-dependent TGF-β signaling pathway in human
glioblastoma. Cancer Res. 71:2392–2402. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang P, Ren Z and Sun P: Overexpression of
the long non-coding RNA MEG3 impairs in vitro glioma cell
proliferation. J Cell Biochem. 113:1868–1874. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Han L, Zhang K, Shi Z, Zhang J, Zhu J, Zhu
S, Zhang A, Jia Z, Wang G, Yu S, et al: lncRNA profile of
glioblastoma reveals the potential role of lncRNAs in contributing
to glioblastoma pathogenesis. Int J Oncol. 40:2004–2012.
2012.PubMed/NCBI
|
|
12
|
Zhang XQ, Sun S, Lam KF, Kiang KM, Pu JK,
Ho AS, Lui WM, Fung CF, Wong TS and Leung GK: A long non-coding RNA
signature in glioblastoma multiforme predicts survival. Neurobiol
Dis. 58:123–131. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhi F, Wang Q, Xue L, Shao N, Wang R, Deng
D, Wang S, Xia X and Yang Y: The use of three long non-coding RNAs
as potential prognostic indicators of astrocytoma. PLoS One.
10:e01352422015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Vital AL, Tabernero MD, Castrillo A,
Rebelo O, Tão H, Gomes F, Nieto AB, Resende Oliveira C, Lopes MC
and Orfao A: Gene expression profiles of human glioblastomas are
associated with both tumor cytogenetics and histopathology. Neuro
Oncol. 12:991–1003. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhou J, Xu T, Yan Y, Qin R, Wang H, Zhang
X, Huang Y, Wang Y, Lu Y, Fu D and Chen J: MicroRNA-326 functions
as a tumor suppressor in glioma by targeting the Nin one binding
protein (NOB1). PLoS One. 8:e684692013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang K, Sun X, Zhou X, Han L, Chen L, Shi
Z, Zhang A, Ye M, Wang Q, Liu C, et al: Long non-coding RNA HOTAIR
promotes glioblastoma cell cycle progression in an EZH2 dependent
manner. Oncotarget. 6:537–546. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Amit D, Matouk IJ, Lavon I, Birman T,
Galula J, Abu-Lail R, Schneider T, Siegal T, Hochberg A and Fellig
Y: Transcriptional targeting of glioblastoma by diphtheria toxin-A
driven by both H19 and IGF2-P4 promoters. Int J Clin Exp Med.
5:124–135. 2012.PubMed/NCBI
|
|
18
|
Matouk IJ, Mezan S, Mizrahi A, Ohana P,
Abu-Lail R, Fellig Y, Degroot N, Galun E and Hochberg A: The
oncofetal H19 RNA connection: Hypoxia, p53 and cancer. Biochim
Biophys Acta. 1803:443–451. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yan C, Jiang Y, Wan Y, Zhang L, Liu J,
Zhou S and Cheng W: Long noncoding RNA NBAT-1 suppresses
tumorigenesis and predicts favorable prognosis in ovarian cancer.
Onco Targets Ther. 10:1993–2002. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Pandey GK, Mitra S, Subhash S, Hertwig F,
Kanduri M, Mishra K, Fransson S, Ganeshram A, Mondal T, Bandaru S,
et al: The risk-associated long noncoding RNA NBAT-1 controls
neuroblastoma progression by regulating cell proliferation and
neuronal differentiation. Cancer Cell. 26:722–737. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hu P, Chu J, Wu Y, Sun L, Lv X, Zhu Y, Li
J, Guo Q, Gong C, Liu B and Su S: NBAT1 suppresses breast cancer
metastasis by regulating DKK1 via PRC2. Oncotarget. 6:32410–32425.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang C, Wang G, Yang J and Wang L: Long
noncoding RNA NBAT1 negatively modulates growth and metastasis of
osteosarcoma cells through suppression of miR-21. Am J Cancer Res.
7:2009–2019. 2017.PubMed/NCBI
|
|
23
|
Pandey GK and Kanduri C: Fighting
neuroblastomas with NBAT1. Oncoscience. 2:79–80. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xie H, Fu JL and Xie C: miR-138-5p is
downregulated in patients with atrial fibrillation and reverses
cardiac fibrotic remodeling via repressing CYP11B2. Eur Rev Med
Pharmacol Sci. 22:4642–4647. 2018.PubMed/NCBI
|
|
25
|
Zhao X, Sun J, Chen Y, Su W, Shan H, Li Y,
Wang Y, Zheng N, Shan H and Liang H: lncRNA PFAR promotes lung
fibroblast activation and fibrosis by targeting miR-138 to regulate
the YAP1-Twist axis. Mol Ther. 26:2206–2217. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sullivan TB, Robert LC, Teebagy PA, Morgan
SE, Beatty EW, Cicuto BJ, Nowd PK, Rieger-Christ KM and Bryan DJ:
Spatiotemporal microRNA profile in peripheral nerve regeneration:
miR-138 targets vimentin and inhibits Schwann cell migration and
proliferation. Neural Regen Res. 13:1253–1262. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liang Z, Feng Q, Xu L, Li S and Zhou L:
CREPT regulated by miR-138 promotes breast cancer progression.
Biochem Biophys Res Commun. 493:263–269. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhao H, Peng R, Liu Q, Liu D, Du P, Yuan
J, Peng G and Liao Y: The lncRNA H19 interacts with miR-140 to
modulate glioma growth by targeting iASPP. Arch Biochem Biophys.
610:1–7. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chen L, Wang Y, He J, Zhang C, Chen J and
Shi D: Long non-coding RNA H19 promotes proliferation and invasion
in human glioma cells by downregulating miR-152. Oncol Res. Feb
8–2018.(Epub ahead of print). View Article : Google Scholar
|
|
30
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhou W, Ye XL, Xu J, Cao MG, Fang ZY, Li
LY, Guan GH, Liu Q, Qian YH and Xie D: The lncRNA H19 mediates
breast cancer cell plasticity during EMT and MET plasticity by
differentially sponging miR-200b/c and let-7b. Sci Signal.
10:eaak95572017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tang J, Li Y, Sang Y, Yu B, Lv D, Zhang W
and Feng H: LncRNA PVT1 regulates triple-negative breast cancer
through KLF5/beta-catenin signaling. Oncogene. 37:4723–4734. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hu HB, Chen Q and Ding SQ: LncRNA
LINC01116 competes with miR-145 for the regulation of ESR1
expression in breast cancer. Eur Rev Med Pharmacol Sci.
22:1987–1993. 2018.PubMed/NCBI
|
|
34
|
Li Q, Gao H, Zhou S and Liao Y: lncRNA
PlncRNA-1 overexpression inhibits the growth of breast cancer by
upregulating TGF-β1 and downregulating PHGDH. Breast Cancer.
25:619–625. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhao L, Han T, Li Y, Sun J, Zhang S, Liu
Y, Shan B, Zheng D and Shi J: The lncRNA SNHG5/miR-32 axis
regulates gastric cancer cell proliferation and migration by
targeting KLF4. FASEB J. 31:893–903. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lü MH, Tang B, Zeng S, Hu CJ, Xie R, Wu
YY, Wang SM, He FT and Yang SM: Long noncoding RNA BC032469, a
novel competing endogenous RNA, upregulates hTERT expression by
sponging miR-1207-5p and promotes proliferation in gastric cancer.
Oncogene. 35:3524–3534. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yan J, Zhang Y, She Q, Li X, Peng L, Wang
X, Liu S, Shen X, Zhang W, Dong Y, et al: Long noncoding RNA
H19/miR-675 axis promotes gastric cancer via FADD/Caspase 8/Caspase
3 signaling pathway. Cell Physiol Biochem. 42:2364–2376. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Li P, Tong L, Song Y, Sun J, Shi J, Wu Z,
Diao Y, Li Y and Wang Z: Long noncoding RNA H19 participates in
metformin-mediated inhibition of gastric cancer cell invasion. J
Cell Physiol. 234:4515–4527. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Bowles J, Schepers G and Koopman P:
Phylogeny of the SOX family of developmental transcription factors
based on sequence and structural indicators. Dev Biol. 227:239–255.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Francois M, Koopman P and Beltrame M: Soxf
genes: Key players in the development of the cardio-vascular
system. Int J Biochem Cell Biol. 42:445–448. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Katoh M: Expression of human SOX7 in
normal tissues and tumors. Int J Mol Med. 9:363–368.
2002.PubMed/NCBI
|
|
42
|
Stovall DB, Cao P and Sui G: SOX7: From a
developmental regulator to an emerging tumor suppressor. Histol
Histopathol. 29:439–445. 2014.PubMed/NCBI
|
|
43
|
Chan DW, Mak CS, Leung TH, Chan KK and
Ngan HY: Down-regulation of Sox7 is associated with aberrant
activation of Wnt/b-catenin signaling in endometrial cancer.
Oncotarget. 3:1546–1556. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Cui J, Xi H, Cai A, Bian S, Wei B and Chen
L: Decreased expression of Sox7 correlates with the upregulation of
the Wnt/β-catenin signaling pathway and the poor survival of
gastric cancer patients. Int J Mol Med. 34:197–204. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhong WD, Qin GQ, Dai QS, Han ZD, Chen SM,
Ling XH, Fu X, Cai C, Chen JH, Chen XB, et al: SOXs in human
prostate cancer: Implication as progression and prognosis factors.
BMC Cancer. 12:2482012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Stovall DB, Wan M, Miller LD, Cao P,
Maglic D, Zhang Q, Stampfer MR, Liu W, Xu J and Sui G: The
regulation of SOX7 and its tumor suppressive role in breast cancer.
Am J Pathol. 183:1645–1653. 2013. View Article : Google Scholar : PubMed/NCBI
|