Introduction
Lung carcinoma, the most commonly diagnosed cancer
(11.6% of total cases) and the leading cause of cancer-associated
mortality (18.4% of total mortalities) worldwide, according to the
GLOBOCAN 2018 estimates of cancer incidence and mortality (1), is characterized by a high degree of
malignancy, metastasis and high mortality rate (1). Lung carcinoma is the most diagnosed
cancer and the leading cause of cancer-associated mortality in men
(1), and the third most common
cancer and second cause of cancer-associated mortality in women,
worldwide (1). Most lung cancer
tumors have no obvious symptoms in the early stage and are
therefore less likely to be detected; thus, the majority of
patients with lung carcinoma are diagnosed at an advanced stage,
which reduces the efficacy of treatment and the 5-year survival
rate is low (2,3). Therefore, novel potential prognostic
biomarkers and drug targets are required to improve the outcome and
individualized treatments of lung cancer.
SMADs are a family of genes that encode
signal transducers and transcriptional modulators, which mediate
various signaling pathways, such as TGF-β/SMAD, BMP/SMAD, ERK/MAPK,
Hippo, JAK/STAT and Wnt/β-catenin (4). In mammals, there are eight SMAD
proteins, which are sub-divided into three types:
Receptor-regulated SMADs (R-SMADs), common-mediator SMADs and
inhibitory SMADs (4–6). SMADs can recognize and bind several
sequence-specific and context-dependent transcriptional regulation
factors, such as FoxH1, Sp1, YY1 and p53, which have been found to
participate in various biological processes, including cell
proliferation, apoptosis, differentiation, as well as tumor
progression and immune regulation processes (7–9). The
majority of signaling pathways regulated by SMADs, such as
TGF-β/SMAD, BMP/SMAD, ERK/MAPK, JAK/STAT and Wnt/β-catenin, are
deregulated in various human malignant carcinomas, including lung
carcinoma, malignant melanoma, colorectal cancer, kidney cancer,
breast cancer, ovarian cancer and prostate cancer (10–17).
A total of seven mammalian SMAD proteins have been
reported to participate in the regulation of lung carcinoma
tumorigenesis or progression. The SMAD-1 gene participates
in negative regulation of the Akt/GSK3β pathway to maintain the
cancer stem cell-like characteristic of cancer stem cells in
non-small cell lung carcinoma (NSCLC) (18). Yang et al (19) found that SMAD1 knockdown inhibited
epithelial-mesenchymal transition (EMT) induced by fine particulate
matter (PM2.5) in human lung carcinoma cells. In addition, Tang
et al (20) reported that
transcriptional activation of SMAD-2 and −3 facilitates the growth
and metastasis of lung carcinoma through enhanced transforming
growth factor (TGF)-β1-induced EMT and from the generation of the
angiogenic factors, such as vascular endothelial growth factor and
connective tissue growth factor. SMAD4 is phosphorylated(p) at
Tyrosine 95 by an oncogenic tyrosine kinase ALK, preventing it from
binding to DNA and eliciting TGF-β-induced tumor suppressing
responses during lung cancer tumorigenesis (21). Phosphorylated (p)-SMAD-9 expression
levels were found to be markedly enhanced and associated with the
metastatic potential of non-small cell lung carcinoma A549
subclones from experimental brain metastases through four rounds of
intracardiac injection of A549 cells or its derivatives into
athymic nude mice (22). Inhibitory
SMADs, including SMAD-6 and −7, have been found to be involved in
lung carcinoma by regulating the stability or activity of TGF-β
receptors, including the modulation of TβRI ubiquitination and
degradation, by which TGF-β signaling is regulated and functions in
lung cancer growth and metastasis (6,23).
However, the underlying mechanisms by which SMADs are involved in
the regulation of lung carcinoma are not fully understood.
At present, the dysregulated mRNA expression levels
of SMAD proteins in human lung cancer and their associations with
lung carcinoma prognosis have not been investigated. In the present
study, bioinformatics analysis was used to investigate the roles of
SMAD proteins in human lung carcinoma. The expression patterns and
mutations of different SMAD proteins in patients with lung
carcinoma were analyzed, from the vast number of gene expression
data that has been previously published, to identify SMAD
expression patterns and potential prognostic values in human lung
carcinoma.
Materials and methods
Oncomine analysis
The Oncomine gene expression array dataset
(www.oncomine.org), was used for analyzing the
expression levels of SMADs in different carcinomas, including
bladder cancer, brain and central nervous system cancer, breast
cancer, cervical cancer, colorectal cancer, esophageal cancer,
gastric cancer, head and neck cancer, kidney cancer, leukemia,
liver cancer, lung cancer, lymphoma, melanoma, myeloma, ovarian
cancer, pancreatic cancer, prostate cancer and sarcoma (24). A comparison of SMAD mRNA
expression levels between clinical tumor specimens and adjacent
normal tissues was performed using unpaired two-tailed Student's
t-tests (number of samples: GSE19188: 10, GSE10072: 107, GSE32863:
116, GSE31210: 246 and GSE7670: 64). The cut-off values were set as
follows, P<0.01 and fold change >2, respectively.
Gene expression omnibus (GEO) dataset
analysis
GSE19188 (25),
GSE10072 (26), GSE32863 (25), GSE31210 (25), GSE7670 (27) were downloaded from the GEO database
(www.ncbi.nlm.nih.gov/geo/) to analyze
the mRNA expression levels of all SMAD subtypes in lung neoplasm
tissues and adjacent normal lung tissues (28). These datasets were obtained by
searching using the following key words: ‘lung cancer AND SMAD’ or
‘lung cancer AND gene’. To avoid generating less reliable results,
the 5 datasets were batch normalized in the R computing environment
in RStudio (version 1.2.5001) using the sva package and merged to
reduce the variability (29–31).
Gene expression profiling interactive
analysis (GEPIA) dataset
GEPIA is an online cancer database used for
analyzing RNA sequences and expression levels for 9,736 lung cancer
samples and 8,587 normal samples from The Cancer Genome Atlas
(TCGA) and GTEx projects, based on a standard processing pipeline
(32). GEPIA can be used to analyze
differential expression in tumor and adjacent normal tissues, tumor
type or clinical stage, patient survival analysis, correlation,
dimensionality reduction (reducing the dimensionality of high
dimensional expression datasets while maintaining most of the
variances based on principal component analysis) and similar gene
detection (32).
Kaplan-Meier plotter
The prognostic value of the SMADs transcriptional
levels was assessed using the online database Kaplan-Meier plotter
(www.kmplot.com) (33,34),
which contains gene expression data and survival information for
2,437 patients with NSCLC. Overall survival (OS), progression-free
survival (PFS) and post-progression survival (PPS) were analyzed
after the patients had been divided into 2 groups (high and low)
based on the median expression value, using Kaplan-Meier curves,
with the log rank test to determine any significant difference. The
hazard ratio (HR) and 95% confidence intervals (CI) were also
calculated. Only SMAD datasets selected using the JetSet best probe
package in R were used in Kaplan-Meier analysis.
TCGA data and cBioPortal
TGCA includes sequencing and pathology data for 30
different cancer types, including glioblastoma multiforme, head and
neck, kidney clear cell, lung adenocarcinoma, lung squamous cell
carcinoma and medulloblastoma (35).
Using cBioPortal (36), SMAD
analyses were conducted using the provisional lung adenocarcinoma
TCGA dataset. Using the Genomic Identification of Significant
Targets in Cancer package (version 1.12.0) which can identify
mutations, putative copy number alterations, this was included with
the z-scores (also known as standard score, it is obtained by
dividing the difference between a number and an average by the
standard deviation. In statistics, the standard score is the sign
number of the standard deviation of the value of an observation or
data point higher than the average of the observed value or
measured value. Its value is positively correlated with mRNA
expression level) (37) of mRNA
expression levels [RNA sequencing V2 RSEM (RNA-Seq by Expectation
Maximization)] (38) and the protein
expression level data (using reverse phase protein array data) to
create the genomic profile for 522 patients with lung
adenocarcinoma. The co-expression network was plotted using
cBioPortal. The functional roles of the target host gene of SMADs
were predicted using Gene Ontology (GO) enrichment analysis of
three elements: Biological process, cellular composition and
molecular function. Kyoto Encyclopedia of Genes and Genomes (KEGG)
function analysis of genes associated with SMAD alterations was
performed using the Database for Annotation, Visualization and
Integrated Discovery (DAVID; http://david.ncifcrf.gov/summary.jsp).
Statistical analysis
Student's t-tests was used to compare mRNA
expression levels in the Oncomine and GEO databases. Kaplan-Meier
plotter and Cramér-von Mises tests were used to analyze survival.
By defining disease state (tumor or normal) as a variable, GEPIA
analysis of variance was performed using one-way ANOVA. Two-tailed
Student's t-test was used to compare the expression levels of
SMAD mRNA between clinical tumor specimens and adjacent
normal tissues. Spearman's correlation analysis was used to
evaluate the correlation between gene expression levels in TCGA
database. P<0.05 was considered to indicate a statistically
significant difference.
Results
Variation of SMADs expression levels
among different types of lung carcinoma and adjacent normal
tissues
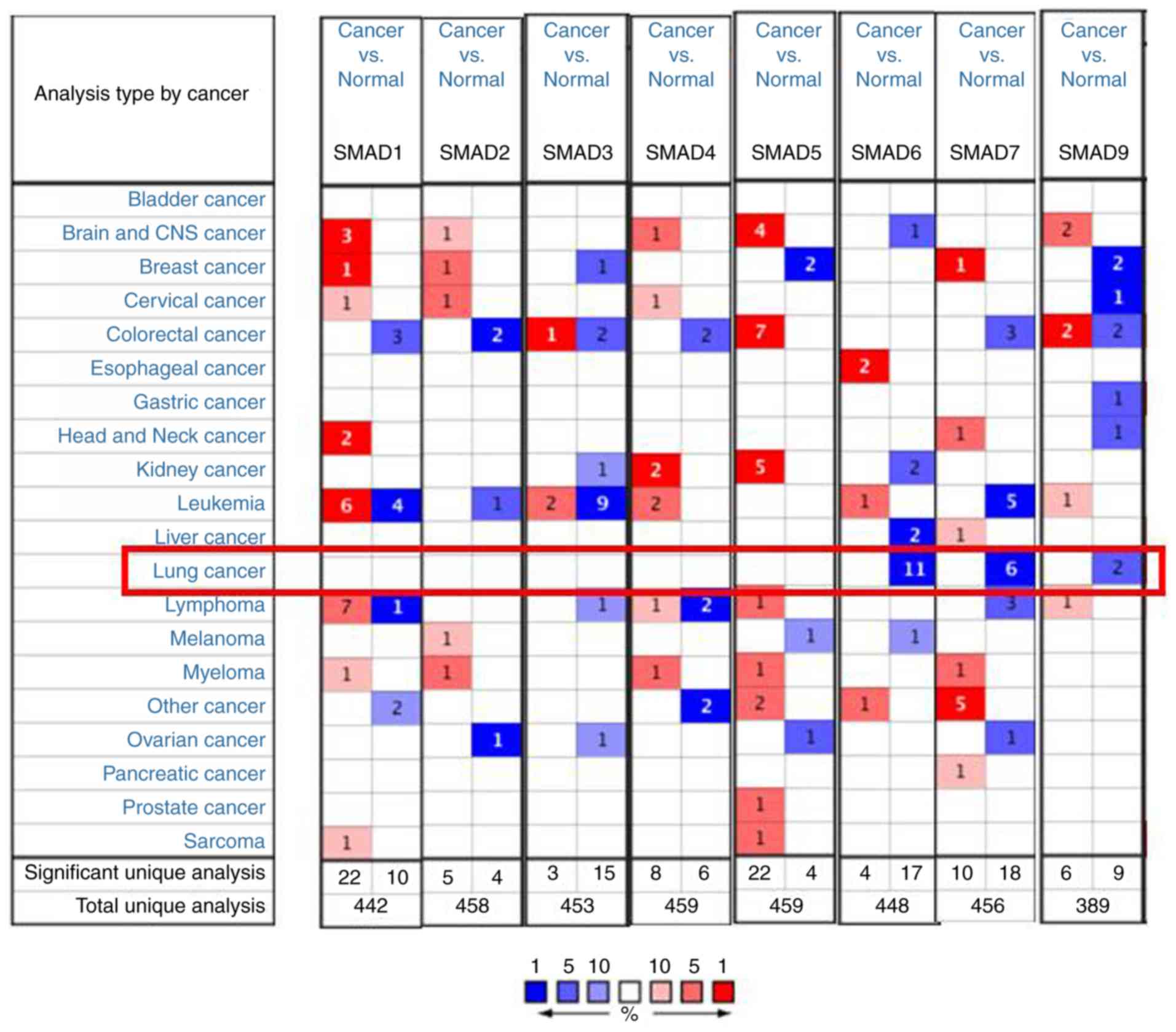
The expression levels of the eight SMADs of various
cancer types and in adjacent normal samples were compared using the
Oncomine database (Fig. 1). The
transcriptional levels of SMAD-6, −7 and −9 were significantly
decreased in patients with lung carcinoma (P<0.05). No
significant expression difference was observed for
SMAD-1/-2/-3/-4/-5 between lung cancer tissues and normal tissues.
With respect to lung adenocarcinoma, the expression levels of
SMAD-6, −7 and −9 were significantly decreased in cancer tissues
compared with that in adjacent normal tissues, with a fold change
of −4.622, −3.508 and −2.285, respectively (P<0.05; Table I) (39–41). In
particular, only the expression levels of SMAD-7 were decreased
compared with that in the adjacent normal samples in all types of
lung cancer; SMAD-7 was found to be decreased in squamous cell lung
carcinoma by −3.813-fold, in small cell lung carcinoma with a fold
change of −4.568 and in lung carcinoid tumor with a fold change of
−6.114 (Table I). There were no
significant differences in the transcription levels for the
remaining SMADs (P>0.05) (25–27).
 | Table I.Significant changes of SMAD
transcriptional levels between different types of lung cancer and
normal lung tissues using the Oncomine database. |
Table I.
Significant changes of SMAD
transcriptional levels between different types of lung cancer and
normal lung tissues using the Oncomine database.
| Genes | Types of lung
cancer vs. normal tissue | Fold change | P-value | t-test value | Author, year | (Refs.) |
|---|
| SMAD-1 | Lung adenocarcinoma
vs. normal | 1.185 | >0.05 | 1.555 | Beer et al,
2002 | (25) |
| SMAD-2 | Squamous cell lung
cancer vs. normal | 1.205 | >0.05 | 1.525 | Garber et
al, 2001 | (26) |
|
| Large cell lung
carcinoma vs. normal | 1.070 | >0.05 | 0.668 | Garber et
al, 2001 | (26) |
|
| Large cell lung
carcinoma vs. normal | 1.412 | >0.05 | 1.272 | Garber et
al, 2001 | (26) |
|
| Small cell lung
carcinoma vs. normal | −1.069 | >0.05 | −0.427 | Garber et
al, 2001 | (26) |
| SMAD-3 | Lung adenocarcinoma
vs. normal | 1.213 | >0.05 | 1.104 | Beer et al,
2002 | (25) |
| SMAD-4 | Lung adenocarcinoma
vs. normal | −1.110 | >0.05 | −1.405 | Beer et al,
2002 | (25) |
| SMAD-5 | Lung adenocarcinoma
vs. normal | 1.415 | >0.05 | 1.239 | Yamagata et
al, 2003 | (27) |
|
| Squamous cell
carcinoma vs. normal | 1.302 | >0.05 | 0.962 | Yamagata et
al, 2003 | (27) |
|
| Large cell
carcinoma vs. normal | 1.398 | >0.05 | 1.073 | Yamagata et
al, 2003 | (27) |
| SMAD-6 | Lung adenocarcinoma
vs. normal | −4.622 | <0.0001 | −12.757 | Selamat et
al, 2012 | (39) |
| SMAD-7 | Lung adenocarcinoma
vs. normal | −3.508 | <0.0001 | −5.632 | Bhattacharjee et
al, 2001 | (40) |
|
| Squamous cell
carcinoma vs. normal | −3.813 | <0.0001 | −4.561 | Bhattacharjee et
al, 2001 | (40) |
|
| Small cell lung
carcinoma vs. normal | −4.568 | <0.0001 | −6.228 | Bhattacharjee et
al, 2001 | (40) |
|
| Lung carcinoid
tumor vs. normal | −6.114 | <0.0001 | −7.804 | Bhattacharjee et
al, 2001 | (40) |
| SMAD-9 | Lung adenocarcinoma
vs. normal | −2.285 | <0.0001 | −5.387 | Okayama et
al, 2012 | (41) |
SMAD mRNA expression levels are
associated with the clinicopathological parameters of patients with
lung carcinoma
GEPIA and GEO were used to compare the mRNA
transcriptional levels of SMAD proteins between lung carcinoma
tissues and adjacent normal tissues. The results from GEPIA
analysis showed that there were significantly lower expression
levels of SMAD-6, −7 and −9 in lung adenocarcinoma and squamous
cell lung carcinoma tissues compared with normal tissues, while
there were no significant differences in the expression levels for
SMAD1-5 (Fig. 2A and C). The results
from GEO analysis showed similar results, and SMAD-7 showed
significantly increased expression in ADC, LCC and SCC samples as
compared with normal tissues (Fig.
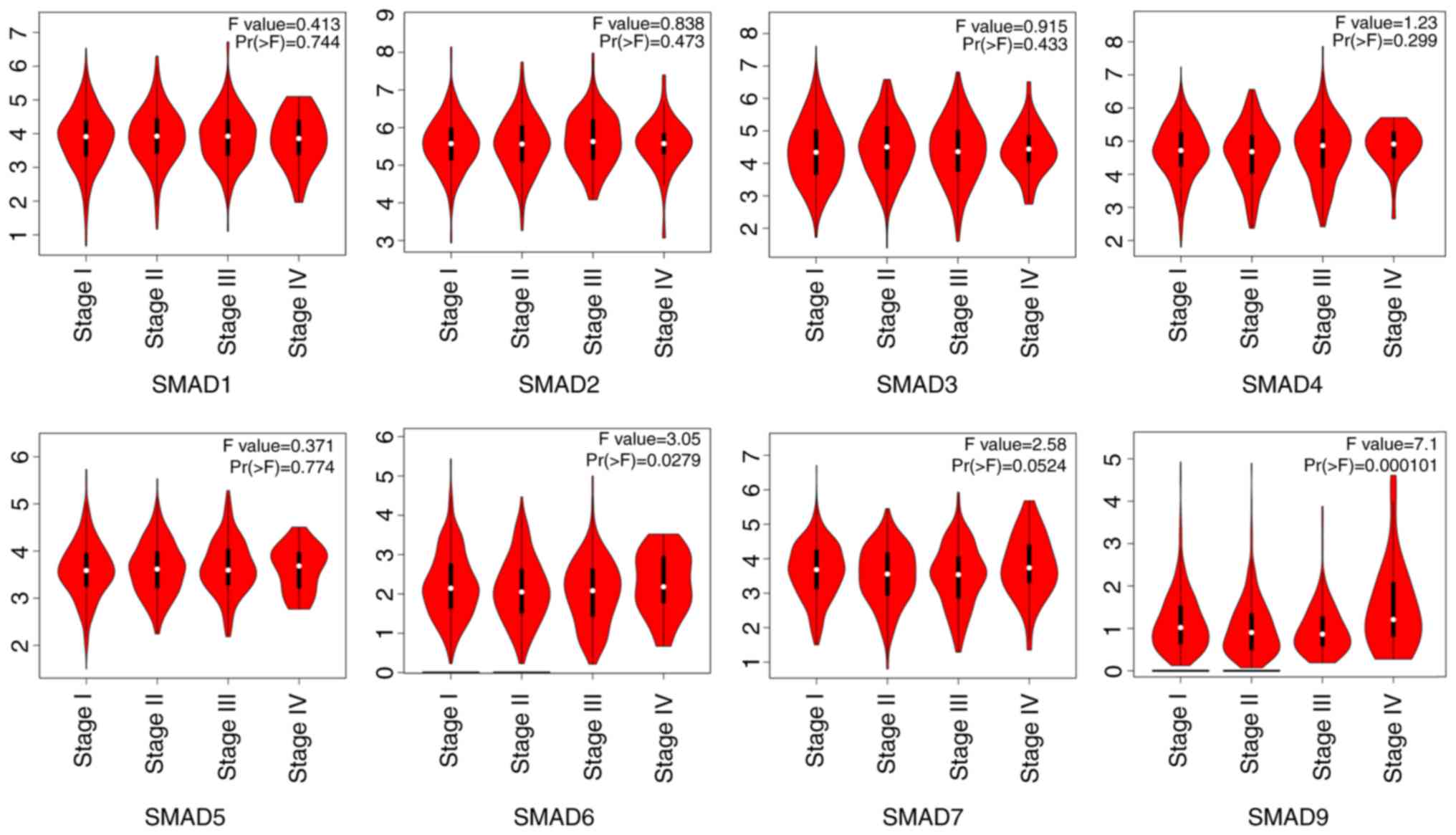
2B). The transcriptional levels of SMADs in different
pathological stages of lung adenocarcinoma and squamous cell
carcinoma tissues were also analyzed. The mRNA expression levels of
SMAD-6 and −9 were significantly different at stages I–IV,
according to the TNM staging system of American Joint Committee on
Cancer (AJCC) in 2010 (42), while
there were no significant differences in the expression levels of
SMAD-1, −2, −3, −4, −5 and −7 (Fig.
3).
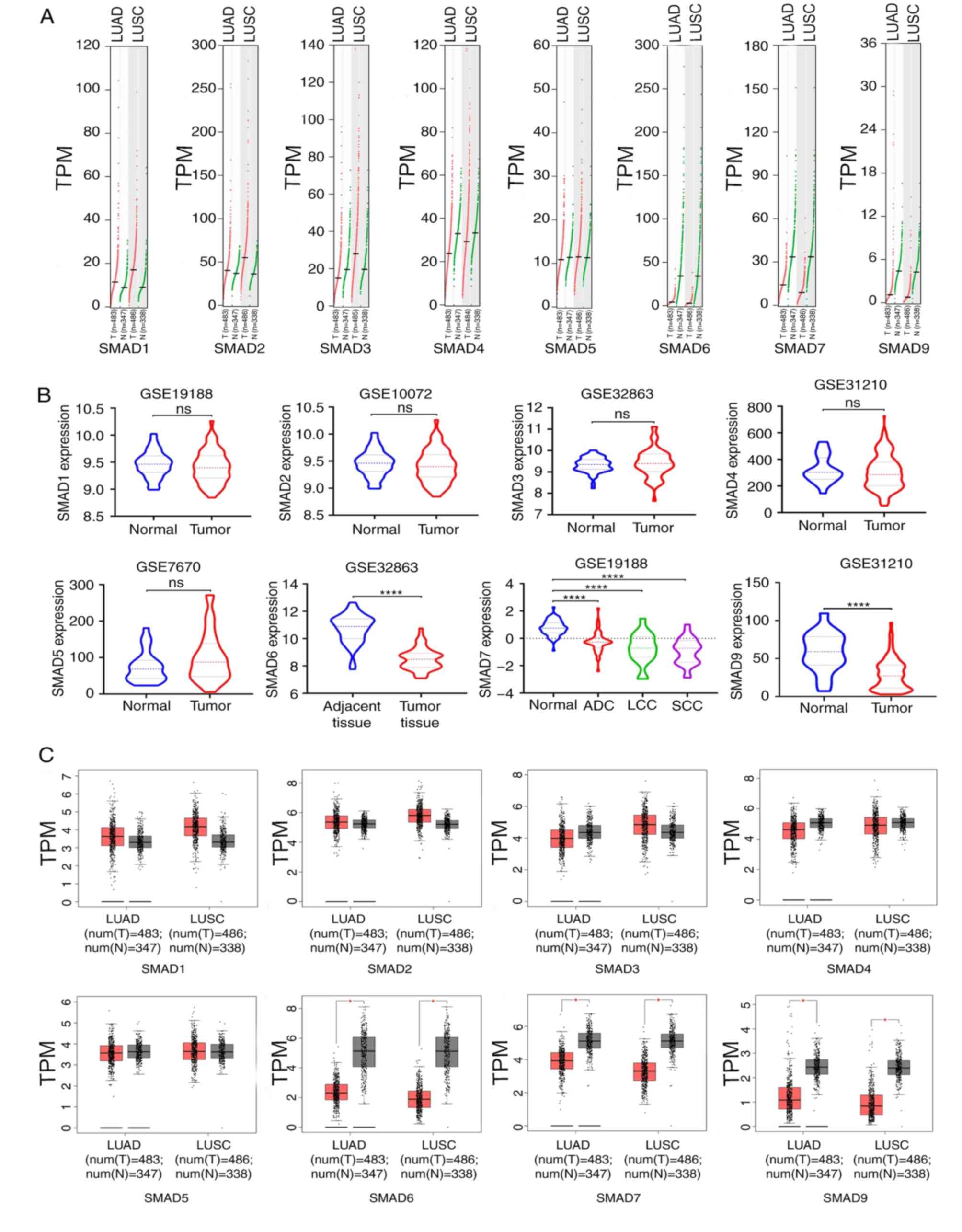 | Figure 2.Expression levels of SMADs in lung
cancer. (A) Analyses of SMAD expression levels were performed using
Gene Expression Profiling Interactive Analysis. (B) Analyses of
SMAD expression levels were performed using the Gene Expression
Omnibus and the results are depicted as a violin plot. (C) Analyses
of SMAD expression levels were performed using Gene Expression
Profiling Interactive Analysis and the results are depicted as a
box plot. ****P<0.0001 and *P<0.05. ns, not significant;
LUAD, lung adenocarcinoma; LUSC, lung squamous cell carcinoma; T,
tumor; N, normal; num, number; ADC, adenocarcinoma; LCC, large cell
carcinoma; SCC, squamous cell carcinoma; TPM, transcripts per
million. |
Association between decreased mRNA
expression levels of SMAD-6, −7 and −9 with the prognosis of
patients with lung carcinoma
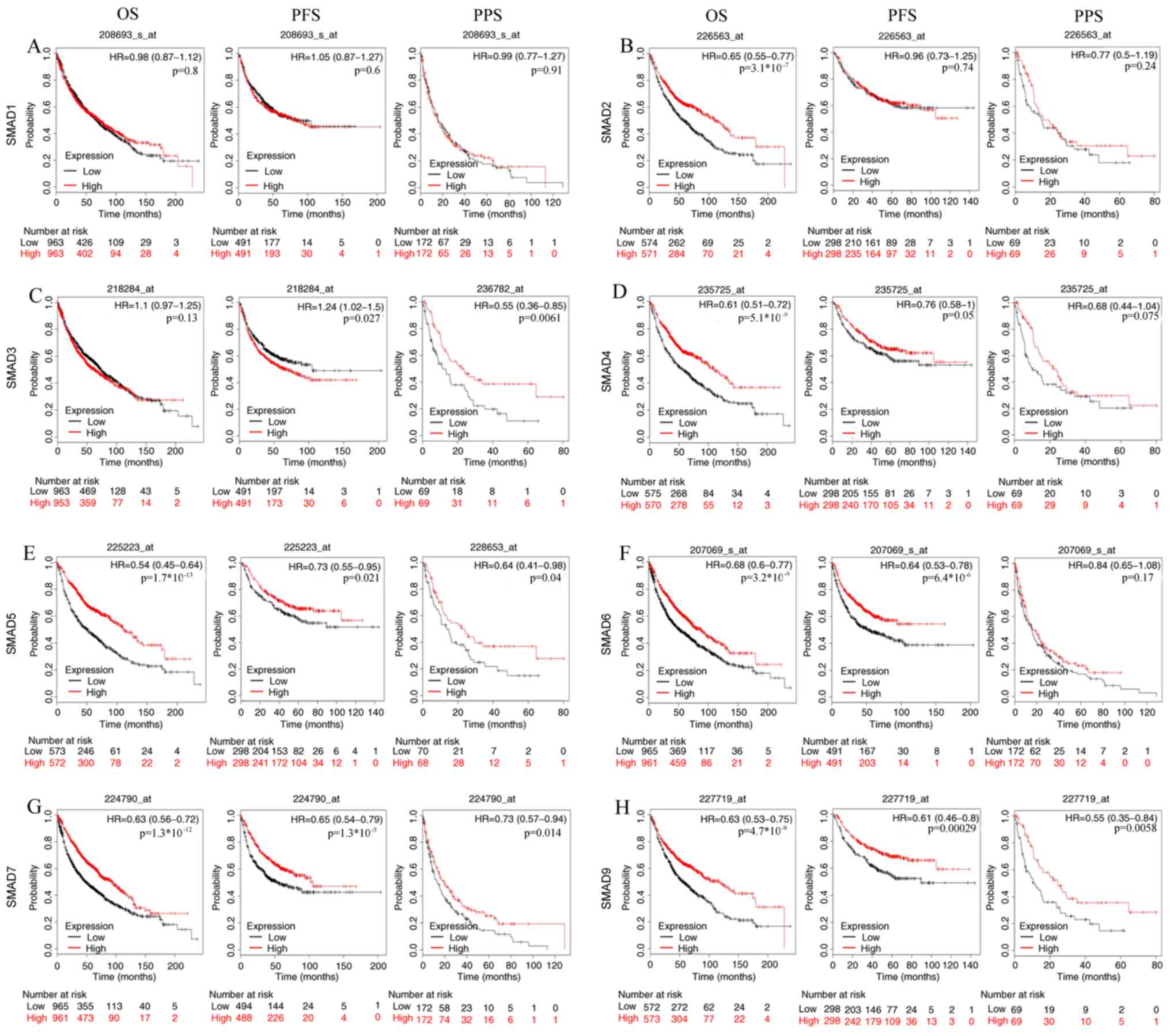
The association between the mRNA expression levels
of SMAD proteins and the survival of patients with NSCLC was
analyzed using Kaplan-Meier plotter, and the results revealed that
the patients with low mRNA expression levels of SMAD-5, −6, −7 and
−9 had poor OS, PFS and PPS rates (from left to right: OS, PFS and
PPS) (P<0.05, Fig. 4), suggesting
their potential roles as a tumor suppressor, except that SMAD6 is
not significantly associated with PPS. SMAD-2, −4 was identified as
a tumor suppressor by OS analysis; however, the results for SMAD-3
were ambiguous, which was demonstrated to be a tumor suppressor for
PPS but an oncogene for PFS.
Changes in SMAD protein expression
levels and their networks in patients with lung carcinoma
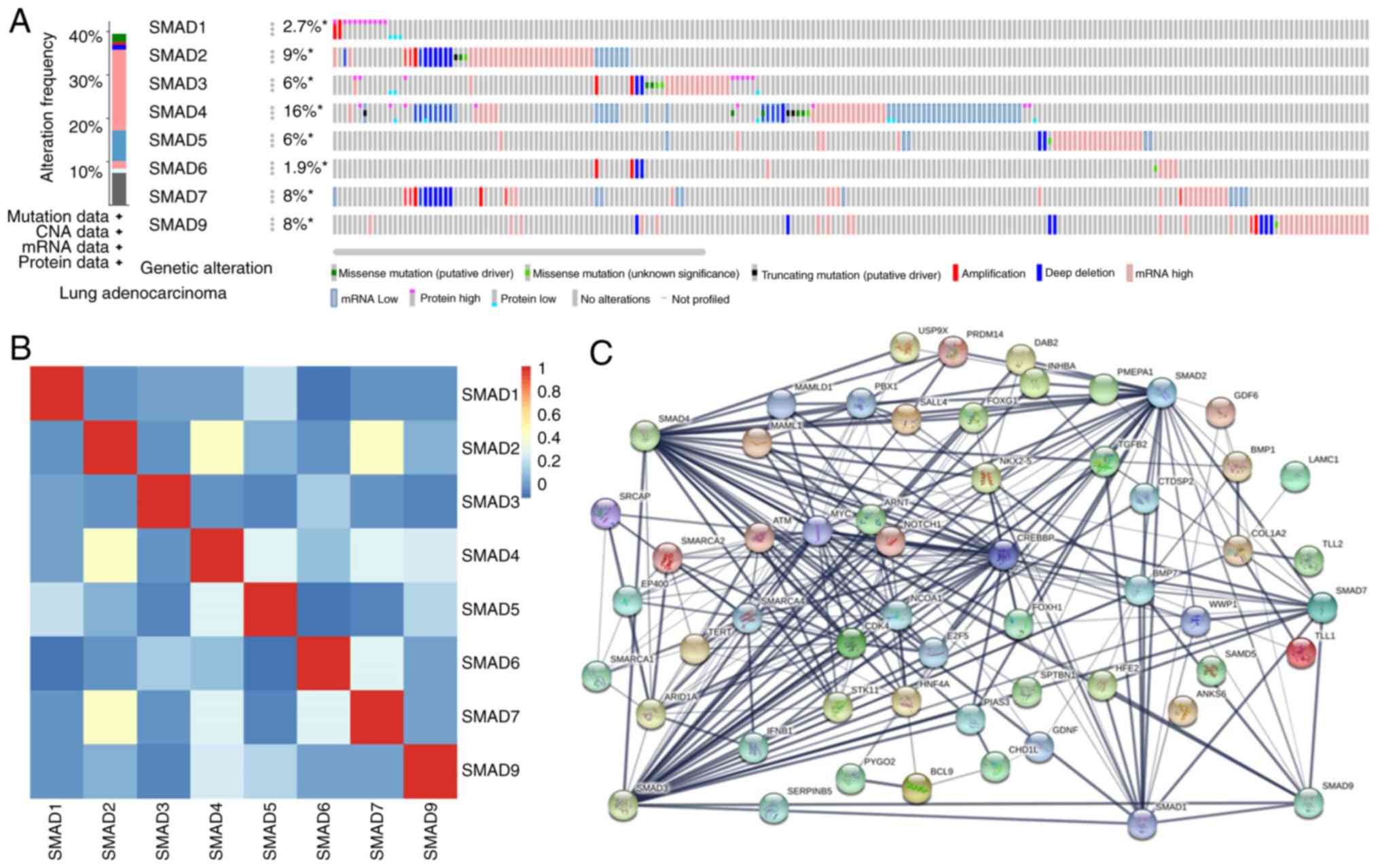
Alterations, associations and networks of SMAD
proteins were analyzed using the cBioPortal online tool for lung
adenocarcinoma. Of the 522 patients with lung adenocarcinoma, 208
(40%) had alterations in SMADs; while >2 changes were detected
in 83 samples (Fig. 5A). Among the
eight SMADs, SMAD-4 has the most alterations (with ‘mRNA low’ being
the most abundant type) while SMAD-6 has the least changes. Deep
deletion was identified in all SMADs except SMAD-1. Missense
mutations were identified in SMAD-2/3/4/5/6/9 but not SMAD-1/7.
Calculation of the correlations between each of the SMADs in lung
adenocarcinoma was performed using cBioPortal online tool based on
their mRNA expression levels. The results indicated that SMAD-2 was
significantly correlated with SMAD4 (R=0.55; P<0.001) and SMAD7
(R=0.4; P<0.001), while SMAD-1, −3, −5, −6 and −9 were not
significantly associated with any other SMADs (Fig. 5B). The interaction network between
SMADs and the 49 most-confirmed altered neighbors was then
constructed, demonstrating that cell cycle-associated genes, such
as CDK4, BCL-9, MYC, CREBBP and E2F5, were associated with changes
to SMAD expression levels (Fig.
5C).
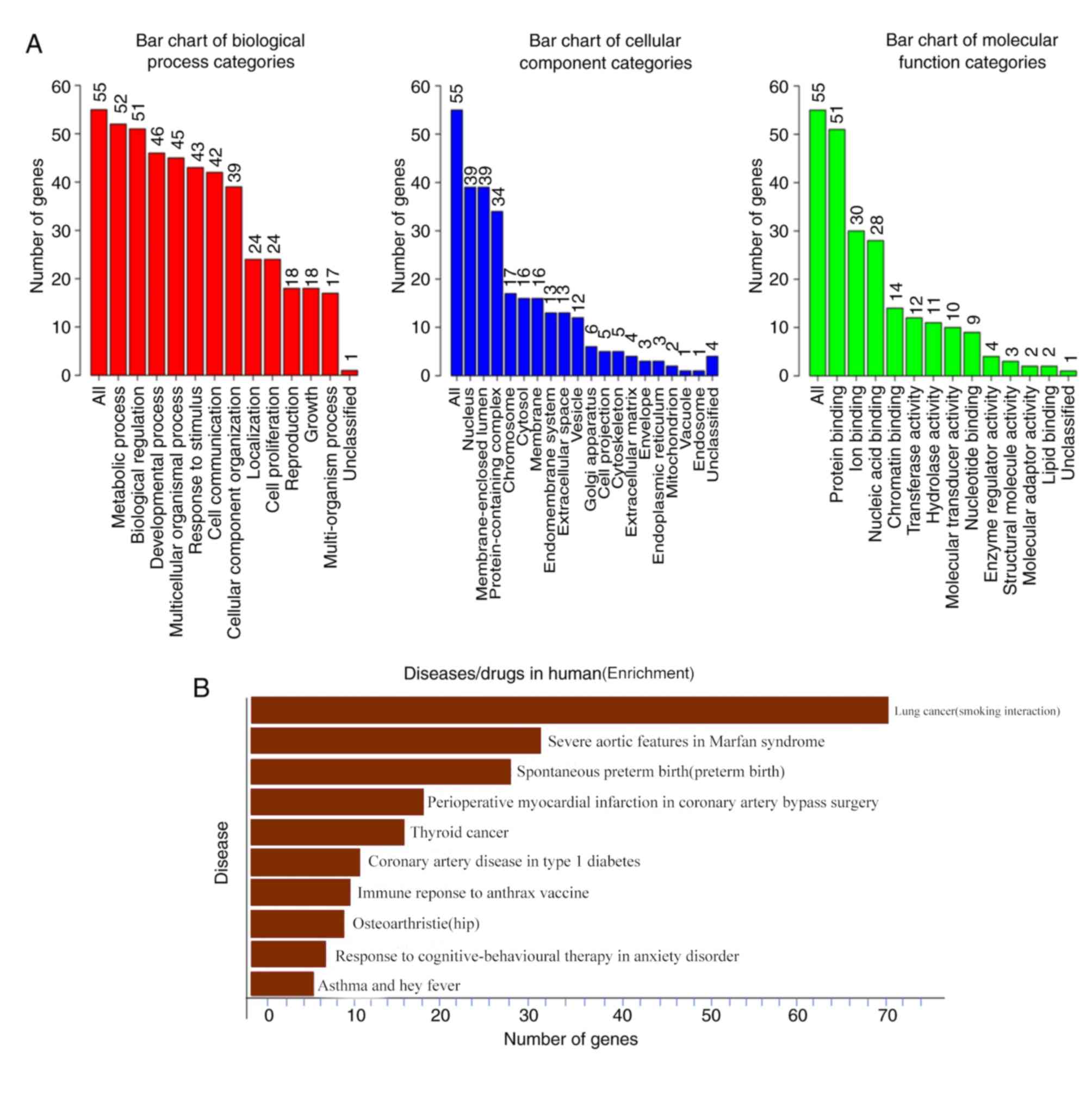
Using the Database for Annotation, Visualization and
Integrated Discovery (https://david.ncifcrf.gov/summary.jsp), the functions
of the SMADs and genes associated with SMAD expression level
changes were predicted. The functional roles of the target host
genes of SMADs were predicted using Gene Ontology (GO) enrichment
analysis of three elements: Biological process, cellular
composition and molecular function. It was found that all SMAD mRNA
expression changes in lung adenocarcinoma were found to be enriched
in cellular metabolic processes, biochemical processes,
multicellular progression and biochemical activity-related
proteins, such as stress (Fig. 6A).
Among the proteins associated with disease and drug sensitivity, it
was found that the SMAD proteins were involved in biological
activities such as ‘lung cancer (smoking interaction)’, ‘severe
aortic features in Marfan syndrome’ and ‘immune response to anthrax
vaccine’ (Fig. 6B).
Discussion
The majority of studies investigating SMAD proteins
are related to the TGF-β signaling pathway, which functions in
tumorigenesis and tumor progression as well as tumor
immunosuppression (11,43–47).
Dysregulation of several SMAD proteins has been proved in numerous
types of cancer. It has been reported that SMAD1 gene polymorphisms
influence colorectal cancer susceptibility (48), while SMAD2 signaling is enhanced in
chemoresistant colorectal cancer cells (49). Tone et al (50) identified R361G mutation of SMAD4 in
primary low-grade serous ovarian carcinoma samples, which is
situated within MH2 domain and is speculated to affect the
functional specificity and selectivity of SMAD4 protein. SMAD5 was
found to be overexpressed in human breast cancer cells and was
involved in the induction of cancer stem cell-like phenotype
(51). Further, SMAD7 expression is
decreased by the overexpression of SETDB1 in human breast cancer,
while upregulation of SMAD7 by SETDB1 knockdown inhibits breast
cancer metastasis (52). However,
bioinformatics analysis of SMAD proteins associated with human lung
cancer has not been performed, to the best of our knowledge. The
present study reported the mRNA expression levels and survival
prediction potential of SMAD proteins in human lung cancer for the
first time.
SMAD1 has been found to be involved in both the
positive modulation of EMT in human malignant lung neoplasm cells
and in the maintenance of stem-like cell traits (18,19).
Immunohistochemical analyses was performed to determine the protein
expression levels of SMAD1 in 60 cases of lung cancer tissues (lung
cancer group), 25 cases of normal alveolus tissues (alveolus
control group) and 29 cases of normal bronchial tissues (bronchial
control group), and the results revealed markedly lower expression
levels of SMAD1 in the lung cancer group compared with that in
normal tissue (53). Furthermore,
the expression levels of SMAD-1 protein were significantly
associated with lung carcinoma differentiation and lymphatic
metastasis (53). However, according
to the present analysis, there were no significant differences in
the expression levels of SMAD-1 between either lung adenocarcinoma
or lung squamous cell carcinoma and adjacent normal tissues. The
results from Liu et al (18)
and Yang et al (19) were
collected from lung cancer cell lines, which might account for
these differences. In addition, the difference between the findings
from the present study (based on 650 cases of lung cancer tissues)
and with that from Gao et al (53) (based on 60 cases of lung cancer
tissues) may be caused by the different sample size.
The regulatory functions of SMAD-2 and −3 have been
explored extensively in lung carcinoma. It was reported that SMAD-2
and −3 function as key R-SMAD proteins mediating the TGF-β/bone
morphogenetic protein signaling pathway to facilitate EMT of lung
carcinoma cells, as well as tumorigenesis, invasion and metastasis
(20,54–58).
Chen et al (59) also
reported that a higher transcriptional level of p-SMAD-2 in stromal
fibroblasts predicted less favorable survival in patients with
pathological stage I to IIIA NSCLC, according to TNM staging system
of American Joint Committee on Cancer (AJCC) in 2010 (42). SMAD3 was also reported to facilitate
the progression of NSCLC by upregulating PAX6 expression (60). However, in the present bioinformatics
study there were no significant differences in the expression
levels of SMAD-2 and −3 between lung carcinoma and adjacent normal
tissues. SMAD-2 was identified as a tumor suppressor using OS
analysis; however, the results for SMAD-3 were ambiguous, as SMAD3
was demonstrated to be a tumor suppressor for PPS but an oncogene
for PFS.
SMAD-4 binds to other active R-SMAD proteins
(SMAD-1/2/3/5/8) to form homo- and heterotypic complexes that
accumulate in the nucleus and regulate transcription of target
genes, including Nanog, CDKN1C, CDKN2B, Nodal, PAI-1, Lefty1 and
SMAD7 (6). SMAD-4 appears to
function as a tumor suppressor in lung carcinoma and is associated
with the differentiation status of lung carcinoma tissues (61,62). It
was found that the p.R361C mutation in SMAD-4 serves a role
in downregulating the TGF-β signaling pathway, causing a loss of
growth inhibition and transcriptional activation mediated by SMADs,
and it is hypothesized that the mutation p.R361C in SMAD-4
plays a crucial part in lung oncogenesis (63). It has been reported that the SMAD4
protein expression levels in NSCLC tissues are significantly lower
compared with that in a normal tracheal-bronchial epithelium
(P<0.05) using immunohistochemistry and that SMAD4 knockdown
initiates and promotes lung carcinoma progression (64). In addition, suppression of SMAD4
protein expression has been found to be associated with the
promotion of growth and invasion and metastasis of lung cancer
(65–67).
In the present study, SMAD-4 was shown to be
downregulated in both lung adenocarcinoma and lung squamous cell
carcinoma; however, this difference was not significant. OS
analysis also demonstrated that SMAD-4 functioned as a tumor
suppressor, which is consistent with the genetic alteration
analysis in lung adenocarcinoma (cBioPortal) shown in Fig. 5A (with both missense and truncating
mutations as putative drivers). A possible explanation for the
tumor suppressor function of SMAD-4 may be due to the missense
mutation sites, given that no difference in expression levels was
observed between cancer tissues and adjacent normal tissues,
similar with that for SMAD-2. This hypothesis is also in accordance
with a previous report indicating that some mutations in SMAD-2 and
−4 (D450H, del1434-6 in SMAD-2 and R420H, R441P in
SMAD-4) are associated with progression in lung cancer
(68), which further supports the
present finding that SMAD-2 is significantly correlated with SMAD-4
(Fig. 5B).
It was observed using a two-stage case-control
study, with 4,680 cases and controls, that there is a significant
association between SMAD-5 rs12719482 and increased risk of
lung carcinoma, suggesting that SMAD-5 participates in the
modulation of lung cancer tumorigenesis. It is speculated that
SMAD-5 rs12719482 is a candidate marker for lung cancer
susceptibility, which decreases the expression of SMAD-5 through
binding has-miR-1270, hsa-miR-571 or hsa-miR-920 (69). This may also explain why the OS
analysis of the present study suggested that SMAD-5 functions as a
tumor suppressor despite the fact that there was no significant
difference in SMAD-5 expression levels.
With regards to SMAD-9, which is another R-SMAD
protein, there have been no previously published studies
investigating its function in the regulation of lung cancer to the
best of our knowledge. The present findings indicated that no
significant mRNA expression level changes were observed for SMAD-5;
however, the expression levels of SMAD-9 was found to be
significantly decreased in lung adenocarcinoma and lung squamous
cell carcinoma tissues compared with that in adjacent normal
tissues, which suggests that SMAD-9 may also serve as a tumor
suppressor in lung cancer progression. In addition, the survival
analysis also suggested that low expression levels of SMAD-5 and −9
were significantly associated with poor OS, PFS and PPS in all
patients with lung carcinoma, which agrees with previous reports
demonstrating the tumor suppressor roles of SMAD-5 and −9 (69–71).
However, as there was no significant association between SMAD-5
expression levels and the cancer stage of lung carcinoma (Fig. 3), the value of SMAD5 as a potential
biomarker is limited.
The two inhibitory SMAD proteins, SMAD-6 and −7,
were found to be downregulated in both human lung adenocarcinoma
and lung squamous cell carcinoma tissues in the present study. Low
expression levels of SMAD6 were associated with poor OS and PFS,
but not PPS, in all patients with lung carcinoma, whilst low
expression levels of SMAD-7 were associated with poor OS, PFS and
PPS. These findings indicate the roles of SMAD-6 and −7 as tumor
suppressors in human lung cancer, which agrees with previous
published findings regarding these functions in lung cancer
(46,72). Moreover, it was found that SMAD7 is
markedly correlated with SMAD-2 as shown in Fig. 5B, which is also consistent with the
finding that the expression of SMAD-2 is regulated by inhibitory
SMAD-7 activity in a negative feedback in cancer cells (73).
As shown in Fig. 5,
different deep deletions were found for SMAD-6, −7 and −9 in
clinical lung adenocarcinoma samples, which is in accordance with
the present survival analysis. Missense mutations were also
identified for SMAD-6 and −9 but not SMAD-7, suggesting that both
deep deletions and missense mutations may serve an important role
in lung cancer progression.
In conclusion, the expression levels and prognostic
values of SMADs in lung cancer were analyzed in the present study,
which improved the understanding of the regulation of lung cancer.
The results demonstrated that decreased expression levels of
SMAD-6, −7 and −9 might function in the tumorigenesis and
development of lung carcinoma, and that these SMADs may also
function as biomarkers to identify patients with a high-risk of
lung carcinoma. Nevertheless, there are still some limitations for
the present study, for example, all results were obtained by
bioinformatics analyses and no experimental validation was
performed. It is suggested that both in vitro and in
vivo experimental validation is required for the confirmation
of the three molecules as bona fide biomarkers for lung
carcinoma, which should be conducted in future studies.
Furthermore, experiments need to be performed to validate the tumor
suppressive functions of the three SMADs in both lung cancer cells
and transplanted lung tumor models. The experimental validation and
the functional deciphering of the three SMADs are of great
importance to better understand the functions of TGF-β signaling in
human lung cancer.
Acknowledgements
Not applicable.
Funding
This study was supported by The National Key R and D
Program of China (grant nos. 2018YFC0115704 and 2018YFC0115705) and
The National Natural Science Foundation of China (grant nos.
11405235 and 81602794).
Availability of data and materials
The datasets generated and/or analyzed during the
present study are available in the [online] repository,
[http://ualcan.path.uab.edu/analysis.html, https://www.oncomine. org/resource/main.html,
https://www.cancer.gov/about-nci/organization/ccg/research/structural-genomics/tcga,
https://www.cbioportal.org/].
Authors' contributions
GZ, WH and HP conceived and designed the study. SP
conducted the bioinformatics analyses. WH and SP drafted the
initial manuscript. All authors read and approved the final
manuscript.
Ethic approval and consent to
participate
The present study was conducted according to The
Declaration of Helsinki. All the datasets were retrieved from open
databases and published literature.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Weller DP, Peake MD and Field JK:
Presentation of lung cancer in primary care. NPJ Prim Care Respir
Med. 29:212019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhao Y, Wang W, Liang H, Yang CJ, D'Amico
T, Ng CSH, Liu CC, Petersen RH, Rocco G, Brunelli A, et al: The
optimal treatment for stage IIIA-N2 non-small cell lung cancer: A
network meta-analysis. Ann Thorac Surg. 107:1866–1875. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Budi EH, Duan D and Derynck R:
Transforming growth factor-β receptors and smads: Regulatory
complexity and functional versatility. Trends Cell Biol.
27:658–672. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Luo K: signaling cross talk between
TGF-β/Smad and other signaling pathways. Cold Spring Harb Perspect
Biol. 9:a0221372017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yu Y and Feng XH: TGF-β signaling in cell
fate control and cancer. Curr Opin Cell Biol. 61:56–63. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kamato D, Do BH, Osman N, Ross BP, Mohamed
R, Xu S and Little PJ: Smad linker region phosphorylation is a
signalling pathway in its own right and not only a modulator of
canonical TGF-β signalling. Cell Mol Life Sci. 77:243–251. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Silvestri L, Nai A, Dulja A and Pagani A:
Hepcidin and the BMP-SMAD pathway: An unexpected liaison. Vitam
Horm. 110:71–99. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dai J, Xu M, Zhang X, Niu Q, Hu Y, Li Y
and Li S: Bi-directional regulation of TGF-β/Smad pathway by
arsenic: A systemic review and meta-analysis of in vivo and in
vitro studies. Life Sci. 220:92–105. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Eckhardt BL, Cao Y, Redfern AD, Chi LH,
Burrows AD, Roslan S, Sloan EK, Parker BS, Loi S, Ueno NT, et al:
Activation of canonical BMP4-SMAD7 signaling suppresses breast
cancer metastasis. Cancer Res. 80:1304–1315. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lang C, Dai Y, Wu Z, Yang Q, He S, Zhang
X, Guo W, Lai Y, Du H, Wang H, et al: SMAD3/SP1 complex-mediated
constitutive active loop between lncRNA PCAT7 and TGF-β signaling
promotes prostate cancer bone metastasis. Mol Oncol. 14:808–828.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang Y, Zeng Y, Liu T, Du W, Zhu J, Liu Z
and Huang JA: The canonical TGF-β/Smad signalling pathway is
involved in PD-L1-induced primary resistance to EGFR-TKIs in
EGFR-mutant non-small-cell lung cancer. Respir Res. 20:1642019.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tuncer E, Calcada RR, Zingg D, Varum S,
Cheng P, Freiberger SN, Deng CX, Kleiter I, Levesque MP, Dummer R
and Sommer L: SMAD signaling promotes melanoma metastasis
independently of phenotype switching. J Clin Invest. 129:2702–2716.
2019. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jung M, Lee JH, Lee C, Park JH, Park YR
and Moon KC: Prognostic implication of pAMPK immunohistochemical
staining by subcellular location and its association with SMAD
protein expression in clear cell renal cell carcinoma. Cancers
(Basel). 11:16022019. View Article : Google Scholar
|
|
15
|
Leng Z, Li Y, Zhou G, Lv X, Ai W, Li J and
Hou L: Krüppel-like factor 4 regulates stemness and mesenchymal
properties of colorectal cancer stem cells through the
TGF-β1/Smad/snail pathway. J Cell Mol Med. 24:1866–1877. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bai Y, Li LD, Li J, Chen RF, Yu HL, Sun
HF, Wang JY and Lu X: A FXYD5/TGF-β/SMAD positive feedback loop
drives epithelial-to-mesenchymal transition and promotes tumor
growth and metastasis in ovarian cancer. Int J Oncol. 56:301–314.
2020.PubMed/NCBI
|
|
17
|
Chen J, Deng Y, Ao L, Song Y, Xu Y, Wang
CC, Choy KW, Tony Chung KH, Du Q, Sui Y, et al: The high-risk HPV
oncogene E7 upregulates miR-182 expression through the TGF-β/Smad
pathway in cervical cancer. Cancer Lett. 460:75–85. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu CW, Li CH, Peng YJ, Cheng YW, Chen HW,
Liao PL, Kang JJ and Yeng MH: Snail regulates Nanog status during
the epithelial-mesenchymal transition via the Smad1/Akt/GSK3β
signaling pathway in non-small-cell lung cancer. Oncotarget.
5:3880–3894. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang D, Ma M, Zhou W, Yang B and Xiao C:
Inhibition of miR-32 activity promoted EMT induced by PM2.5
exposure through the modulation of the Smad1-mediated signaling
pathways in lung cancer cells. Chemosphere. 184:289–298. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tang YN, Ding WQ, Guo XJ, Yuan XW, Wang DM
and Song JG: Epigenetic regulation of Smad2 and Smad3 by profilin-2
promotes lung cancer growth and metastasis. Nat Commun. 6:82302015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang Q, Xiao M, Gu S, Xu Y, Liu T, Li H,
Yu Y, Qin L, Zhu Y, Chen F, et al: ALK phosphorylates SMAD4 on
tyrosine to disable TGF-β tumour suppressor functions. Nat Cell
Biol. 21:179–189. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rao G, Pierobon M, Kim IK, Hsu WH, Deng J,
Moon YW, Petricoin EF, Zhang YW, Wang Y and Giaccone G: Inhibition
of AKT1 signaling promotes invasion and metastasis of non-small
cell lung cancer cells with K-RAS or EGFR mutations. Sci Rep.
7:70662017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yu J, Lei R, Zhuang X, Li X, Li G, Lev S,
Segura MF, Zhang X and Hu G: MicroRNA-182 targets SMAD7 to
potentiate TGFβ-induced epithelial-mesenchymal transition and
metastasis of cancer cells. Nat Commun. 7:138842016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu W, Ouyang S, Zhou Z, Wang M, Wang T,
Qi Y, Zhao C, Chen K and Dai L: Identification of genes associated
with cancer progression and prognosis in lung adenocarcinoma:
Analyses based on microarray from oncomine and the cancer genome
atlas databases. Mol Genet Genomic Med. 7:e005282019.PubMed/NCBI
|
|
25
|
Beer DG, Kardia SL, Huang CC, Giordano TJ,
Levin AM, Misek DE, Lin L, Chen G, Gharib TG, Thomas DG, et al:
Gene-expression profiles predict survival of patients with lung
adenocarcinoma. Nat Med. 8:816–824. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Garber ME, Troyanskaya OG, Schluens K,
Petersen S, Thaesler Z, Pacyna-Gengelbach M, van de Rijn M, Rosen
GD, Perou CM, Whyte RI, et al: Diversity of gene expression in
adenocarcinoma of the lung. Proc Natl Acad Sci USA. 98:13784–13789.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yamagata N, Shyr Y, Yanagisawa K, Edgerton
M, Dang TP, Gonzalez A, Nadaf S, Larsen P, Roberts JR, Nesbitt JC,
et al: A training-testing approach to the molecular classification
of resected non-small cell lung cancer. Clin Cancer Res.
9:4695–4704. 2003.PubMed/NCBI
|
|
28
|
Pei YF, Xu XN, Wang ZF, Wang FW, Wu WD,
Geng JF and Liu XQ: Methyl-CpG binding domain protein 2 inhibits
the malignant characteristic of lung adenocarcinoma through the
epigenetic modulation of 10 to 11 translocation 1 and miR-200s. Am
J Pathol. 189:1065–1076. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Loraine AE, Blakley IC, Jagadeesan S,
Harper J, Miller G and Firon N: Analysis and visualization of
RNA-Seq expression data using RStudio, bioconductor, and integrated
genome browser. Methods Mol Biol. 1284:481–501. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Benso A, Di Carlo S, Politano G, Savino A
and Hafeezurrehman H: Building gene expression profile classifiers
with a simple and efficient rejection option in R. BMC
Bioinformatics. 12 (Suppl 13):S32011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Leek JT, Johnson WE, Parker HS, Jaffe AE
and Storey JD: The sva package for removing batch effects and other
unwanted variation in high-throughput experiments. Bioinformatics.
28:882–883. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tang Z, Li C, Kang B, Gao G, Li C and
Zhang Z: GEPIA: A web server for cancer and normal gene expression
profiling and interactive analyses. Nucleic Acids Res 45 (W1).
W98–W102. 2017. View Article : Google Scholar
|
|
33
|
Israel Y, Rachmiel A, Gourevich K and
Nagler R: Kaplan-Meier analysis of salivary gland tumors: Prognosis
and long-term survival. J Cancer Res Clin Oncol. 145:2123–2130.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kim J: Drawing guideline for JKMS
manuscript (01) Kaplan-Meier curve and survival analysis. J Korean
Med Sci. 34:e352019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sun CC, Zhou Q, Hu W, Li SJ, Zhang F, Chen
ZL, Li G, Bi ZY, Bi YY, Gong FY, et al: Transcriptional E2F1/2/5/8
as potential targets and transcriptional E2F3/6/7 as new biomarkers
for the prognosis of human lung carcinoma. Aging (Albany NY).
10:973–987. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Unberath P, Knell C, Prokosch HU and
Christoph J: Developing new analysis functions for a translational
research platform: Extending the cBioPortal for cancer genomics.
Stud Health Technol Inform. 258:46–50. 2019.PubMed/NCBI
|
|
37
|
Tarca AL, Romero R, Erez O, Gudicha DW,
Than NG, Benshalom-Tirosh N, Pacora P, Hsu CD, Chaiworapongsa T,
Hassan SS and Gomez-Lopez N: Maternal whole blood mRNA signatures
identify women at risk of early preeclampsia: A longitudinal study.
J Matern Fetal Neonatal Med. 1–12. 2020.(Online ahead of print).
View Article : Google Scholar
|
|
38
|
Li B and Dewey CN: RSEM: Accurate
transcript quantification from RNA-Seq data with or without a
reference genome. BMC Bioinformatics. 12:3232011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Selamat SA, Chung BS, Girard L, Zhang W,
Zhang Y, Campan M, Siegmund KD, Koss MN, Hagen JA, Lam WL, et al:
Genome-scale analysis of DNA methylation in lung adenocarcinoma and
integration with mRNA expression. Genome Res. 22:1197–1211. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Bhattacharjee A, Richards WG, Staunton J,
Li C, Monti S, Vasa P, Ladd C, Beheshti J, Bueno R, Gillette M, et
al: Classification of human lung carcinomas by mRNA expression
profiling reveals distinct adenocarcinoma subclasses. Proc Natl
Acad Sci USA. 98:13790–13795. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Okayama H, Kohno T, Ishii Y, Shimada Y,
Shiraishi K, Iwakawa R, Furuta K, Tsuta K, Shibata T, Yamamoto S,
et al: Identification of genes upregulated in ALK-positive and
EGFR/KRAS/ALK-negative lung adenocarcinomas. Cancer Res.
72:100–111. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Rami-Porta R, Asamura H, Travis WD and
Rusch VW: Lung cancer-major changes in the American joint committee
on cancer eighth edition cancer staging manual. CA Cancer J Clin.
67:138–155. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Gurrapu S, Franzolin G, Fard D, Accardo M,
Medico E, Sarotto I, Sapino A, Isella C and Tamagnone L: Reverse
signaling by semaphorin 4C elicits SMAD1/5- and ID1/3-dependent
invasive reprogramming in cancer cells. Sci Signal.
12:eaav20412019. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Girolami I, Veronese N, Smith L, Caruso
MG, Reddavide R, Leandro G, Demurtas J and Nottegar A: The
activation status of the TGF-β transducer Smad2 is associated with
a reduced survival in gastrointestinal cancers: A systematic review
and meta-analysis. Int J Mol Sci. 20:38312019. View Article : Google Scholar
|
|
45
|
Liang C, Xu J, Meng Q, Zhang B, Liu J, Hua
J, Zhang Y, Shi S and Yu X: TGFB1-induced autophagy affects the
pattern of pancreatic cancer progression in distinct ways depending
on SMAD4 status. Autophagy. 16:486–500. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Jeon HS, Dracheva T, Yang SH, Meerzaman D,
Fukuoka J, Shakoori A, Shilo K, Travis WD and Jen J: SMAD6
contributes to patient survival in non-small cell lung cancer and
its knockdown reestablishes TGF-beta homeostasis in lung cancer
cells. Cancer Res. 68:9686–9692. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Ma L, Jiang H, Xu X, Zhang C, Niu Y, Wang
Z, Tao Y, Li Y, Cai F, Zhang X, et al: Tanshinone IIA mediates
SMAD7-YAP interaction to inhibit liver cancer growth by
inactivating the transforming growth factor beta signaling pathway.
Aging (Albany NY). 11:9719–9737. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Karmokar PF, Asaduzzaman M, Islam MS,
Shahriar M and Shabnaz S: The influence of SMAD1 gene polymorphisms
on colorectal cancer susceptibility in Bangladeshi population: A
case-control study. FEBS Open Bio. 8:3402018.
|
|
49
|
Cheng M, Jiang Y, Yang H, Zhao D, Li L and
Liu X: FLNA promotes chemoresistance of colorectal cancer through
inducing epithelial-mesenchymal transition and smad2 signaling
pathway. Am J Cancer Res. 10:403–423. 2020.PubMed/NCBI
|
|
50
|
Tone AA, McConechy MK, Yang W, Ding J, Yip
S, Kong E, Wong KK, Gershenson DM, Mackay H, Shah S, et al:
Intratumoral heterogeneity in a minority of ovarian low-grade
serous carcinomas. BMC Cancer. 14:9822014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Opyrchal M, Gil M, Salisbury JL, Goetz MP,
Suman V, Degnim A, McCubrey J, Haddad T, Iankov I, Kurokawa CB, et
al: Molecular targeting of the Aurora-A/SMAD5 oncogenic axis
restores chemosensitivity in human breast cancer cells. Oncotarget.
8:91803–91816. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Ryu TY, Kim K, Kim SK, Oh JH, Min JK, Jung
CR, Son MY, Kim DS and Cho HS: SETDB1 regulates SMAD7 expression
for breast cancer metastasis. BMB Rep. 52:139–144. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Gao YQ, Liu M and Zhang H: Expression
profiles of Smad1 protein in lung cancer tissues and normal tissues
and its effect on lung cancer incidence. J Biol Regul Homeost
Agents. 30:165–171. 2016.PubMed/NCBI
|
|
54
|
Chae DK, Ban E, Yoo YS, Kim EE, Baik JH
and Song EJ: MIR-27a regulates the TGF-β signaling pathway by
targeting SMAD2 and SMAD4 in lung cancer. Mol Carcinog.
56:1992–1998. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Sun W, Ma Y, Chen P and Wang D:
MicroRNA-10a silencing reverses cisplatin resistance in the
A549/cisplatin human lung cancer cell line via the transforming
growth factor-β/Smad2/STAT3/STAT5 pathway. Mol Med Rep.
11:3854–3859. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Wang Z, Lu Y, Sheng B, Ding Y and Cheng X:
Catalpol inhibits TGF-β1-induced epithelial-mesenchymal transition
in human non-small-cell lung cancer cells through the inactivation
of Smad2/3 and NF-κB signaling pathways. J Cell Biochem. Sep
11–2018.(Online ahead of print).
|
|
57
|
Wei Y, Li D, Wang D, Qiu T and Liu K:
WITHDRAWN: Evaluation of microRNA-203 in bone metastasis of
patients with non-small cell lung cancer through TGF-β/SMAD2
expression. Oncol Rep. Sep 21–2017.(Online ahead of print).
View Article : Google Scholar
|
|
58
|
Zhang JX, Zhai JF, Yang XT and Wang J:
MicroRNA-132 inhibits migration, invasion and
epithelial-mesenchymal transition by regulating TGFβ1/Smad2 in
human non-small cell lung cancer. Eur Rev Med Pharmacol Sci.
20:3793–3801. 2016.PubMed/NCBI
|
|
59
|
Chen Y, Xing P, Chen Y, Zou L, Zhang Y, Li
F and Lu X: High p-Smad2 expression in stromal fibroblasts predicts
poor survival in patients with clinical stage I to IIIA non-small
cell lung cancer. World J Surg Oncol. 12:3282014. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Qian Z, Zhang QK, Hu Y, Zhang T, Li J, Liu
Z, Zheng H, Gao Y, Jia W, Hu A, et al: Investigating the mechanism
by which SMAD3 induces PAX6 transcription to promote the
development of non-small cell lung cancer. Respir Res. 19:2622018.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Ziemke M, Patil T, Nolan K, Tippimanchai D
and Malkoski SP: Reduced Smad4 expression and DNA topoisomerase
inhibitor chemosensitivity in non-small cell lung cancer. Lung
Cancer. 109:28–35. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Chen H, Wang JW, Liu LX, Yan JD, Ren SH,
Li Y and Lu Z: Expression and significance of transforming growth
factor-β receptor type II and DPC4/Smad4 in non-small cell lung
cancer. Exp Ther Med. 9:227–231. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
D'Haene N, Le Mercier M, Salmon I, Mekinda
Z, Remmelink M and Berghmans T: SMAD4 mutation in small cell
transformation of epidermal growth factor receptor mutated lung
adenocarcinoma. Oncologist. 24:9–13. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Haeger SM, Thompson JJ, Kalra S, Cleaver
TG, Merrick D, Wang XJ and Malkoski SP: Smad4 loss promotes lung
cancer formation but increases sensitivity to DNA topoisomerase
inhibitors. Oncogene. 35:577–586. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Zeng YY, Zhu JJ, Shen D, Qin H, Lei Z, Li
W, Liu Z and Huang JA: MicroRNA-205 targets SMAD4 in non-small cell
lung cancer and promotes lung cancer cell growth in vitro and in
vivo. Oncotarget. 8:30817–30829. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Takahashi K, Nishikawa S, Miyata R,
Noguchi M, Ishikawa H, Yutaka Y, Nakajima D, Hamaji M, Ohsumi A,
Menju T, et al: Tranilast inhibits TGF-beta-induced EMT and
invasion/metastasis via the suppression of smad4 in lung cancer
cell lines. Ann Oncol 29:. (Suppl 8):viii1–viii13. 2018.
|
|
67
|
Lee CC, Yang WH, Li CH, Cheng YW, Tsai CH
and Kang JJ: Ligand independent aryl hydrocarbon receptor inhibits
lung cancer cell invasion by degradation of Smad4. Cancer Lett.
376:211–217. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Yanagisawa K, Uchida K, Nagatake M, Masuda
A, Sugiyama M, Saito T, Yamaki K, Takahashi T and Osada H:
Heterogeneities in the biological and biochemical functions of
Smad2 and Smad4 mutants naturally occurring in human lung cancers.
Oncogene. 19:2305–2311. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Zhang Z, Wang J, Zeng X, Li D, Ding M,
Guan R, Yuan L, Zhou Q, Guo M, Xiong M, et al: Two-stage study of
lung cancer risk modification by a functional variant in the
3′-untranslated region of SMAD5 based on the bone morphogenetic
protein pathway. Mol Clin Oncol. 8:38–46. 2018.PubMed/NCBI
|
|
70
|
Ngeow J, Yu W, Yehia L, Niazi F, Chen J,
Tang X, Heald B, Lei J, Romigh T, Tucker-Kellogg L, et al: Exome
sequencing reveals germline SMAD9 mutation that reduces phosphatase
and tensin homolog expression and is associated with hamartomatous
polyposis and gastrointestinal ganglioneuromas. Gastroenterology.
149:886–889 e5. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Zhang Q, Gan H, Song W, Chai D and Wu S:
MicroRNA-145 promotes esophageal cancer cells proliferation and
metastasis by targeting SMAD5. Scand J Gastroenterol. 53:769–776.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Shi JQ, Wang B, Cao XQ, Wang YX, Cheng X,
Jia CL, Wen T, Luo BJ and Liu ZD: Circular RNA_LARP4 inhibits the
progression of non-small-cell lung cancer by regulating the
expression of SMAD7. Eur Rev Med Pharmacol Sci. 24:1863–1869.
2020.PubMed/NCBI
|
|
73
|
Jin L, Zhu C, Wang X, Li C, Cao C, Yuan J
and Li S: Urocortin attenuates TGFβ1-induced Snail1 and slug
expressions: Inhibitory role of Smad7 in Smad2/3 signaling in
breast cancer cells. J Cell Biochem. 116:2494–2503. 2015.
View Article : Google Scholar : PubMed/NCBI
|