Introduction
According to 2018 GLOBOCAN statistics, pancreatic
cancer had 458,918 new cases and 432,242 mortalities, which
accounted for 4.5% of cancer-associated deaths worldwide (1,2).
Pancreatic ductal adenocarcinoma (PDAC), the most common type of
pancreatic cancer, has a poor prognosis with a five-year overall
survival rate of 5%, which has not significantly improved over the
last two decades despite improved chemotherapeutic and biological
agents (3). Hence, there is a
requirement to better understand the underlying molecular
mechanisms of the disease as a basis for identifying novel and more
effective therapeutic targets.
More than one growth factor potentiates
carcinogenesis and multiple growth factor signalling pathways are
consequently upregulated in cancerous cells (4–6). The
epidermal growth factor family of receptors (EGFR) are
overexpressed in >60% of PDAC tumours (7); however, EGFR-targeted therapies have
been unsuccessful in improving overall patient survival (5,8). Hence,
there must be a simultaneous upregulation of alternative pathways.
One such potential pathway is the insulin/insulin-like growth
factor (IGF) pathway. In PDAC, overexpression of both IGF-1 and
type 1 IGF receptor (IGF-1R) has been demonstrated (9). Furthermore, EGFR has been identified to
be co-expressed with IGF-1R, an interaction that confers resistance
to EGFR-specific therapies (10).
The IGF signalling pathway is important for cellular
growth, proliferation and differentiation, as well as for tissue
integrity (11). It has been
indicated to be aberrant in PDAC (12), enhancing tumourigenesis and
therapeutic resistance (13);
furthermore, IGF-1R is associated with increased proliferation and
angiogenesis and decreased apoptosis (14–16). A
recent study by our group reported on the upregulation of the IGF
signalling pathway in resected pancreatic tumours obtained from
South African patients (17).
Furthermore, IGF-1R has been demonstrated to have predictive and
prognostic value, as its overexpression has been associated with
metastases and decreased overall survival in PDAC (5), highlighting the requirement to further
investigate the regulation of this pathway and its mechanism of
action in PDAC.
MicroRNAs (miRNAs) are non-coding RNAs ~17–24
nucleotides in length, which are involved in biological and
cellular processes by regulation of mRNA translation (18,19).
Circulating miRNAs have been determined to be involved in
tumorigenesis serving as biomarkers and therapeutic targets
(20–24). In the present study, differentially
expressed miRNAs in patients with PDAC compared to patients with
chronic pancreatitis (CP) and a control group (CG) were identified.
Using bioinformatics analysis, the possible regulation of the
insulin receptor (INSR)/IGF signalling pathway activity by miRNAs
was further investigated.
Patients and methods
Patients
A total of 112 patients with cytologically or
histologically confirmed treatment-naive PDAC located in the head
of the pancreas were recruited at the time of diagnosis in Chris
Hani Baragwanath Hospital, Johannesburg, South Africa from January
2014 to December 2016. Participants were categorized into three
clinical groups according to their TNM staging (25). Group 1 consisted of participants with
resectable, non-metastatic disease (TNM stage 0-IIB), Group 2
comprised patients with non-resectable disease without evidence of
metastases (TNM stage III) and Group 3 included participants with
metastatic disease (TNM stage IV). Participants with tumours other
than PDAC were excluded. All clinical information was collected and
stored using REDCap v6.7 electronic data capture tools (26).
As the CP group, patients who had completed at least
one year of out-patient follow-up with the Hepatopancreatobiliary
unit in Chris Hani Baragwanath Hospital, (Johannesburg, South
Africa) and admitted between January 2014 to December 2016 and had
an abdominal CT scan demonstrating no mass or lesion suspicious of
a tumour in their pancreas within two weeks of their recruitment
date were included. CP was diagnosed radiologically or
endoscopically according to international guidelines (27,28). Any
patient with CP and a pancreas mass or cyst was excluded.
As the CG, consecutive patients admitted to the
vascular ward in Chris Hani Baragwanath Hospital (Johannesburg,
South Africa) and admitted between January 2014 to December 2016
with severe peripheral arterial disease who underwent an abdominal
CT scan as part of their clinical work-up were included if the scan
demonstrated a healthy pancreas. Participants were excluded if they
had a history of any previous malignancy or pancreatic disease,
obstructive jaundice, unexplained diarrhoea or weight loss.
Sample processing
From each participant, venous blood samples were
collected in 4.5-ml tubes containing EDTA and stored at room
temperature. Samples were centrifuged for 15 min at 1,500 × g at
room temperature within 4 h of collection. The plasma was aliquoted
into 1-ml Eppendorf tubes and stored at −80°C. Analysis was
performed within 12 months of sample collection and storage.
Reverse transcription-quantitative PCR
(RT-qPCR)
Plasma miRNA was extracted using the miRNeasy
serum/plasma miRNA isolation kit (Qiagen GmbH). The ID3EAL miRNA
knowledge panel 384 Target kit (MiRXES) was used and experiments
were performed according to the manufacturer's instructions. The
kit contained the ID3EAL Panel RT Primer Pool for the miRNAs with a
set of three proprietary spike-in RNAs [ID3EAL Panel RNA Spike-In
(MiRXES)] to normalize variations in RNA isolation efficiency.
Isolated serum RNA was subjected to RT using the ID3EAL Reverse
Transcriptase kit (MiRXES). A 6-log serial dilution of synthetic
templates for each miRNA was concurrently reversed-transcribed.
Using the miRNA-specific qPCR assays-ID3EAL miRNA qPCR Master Mix
(MiRXES), >300 candidate miRNAs were measured in each
complementary DNA sample. The following cycling conditions were
used: 95°C for 10 min, 40°C for 5 min, and then 45 cycles of 95°C
for 10 sec and 60°C for 30 sec. Absolute copy numbers of each miRNA
were determined by interpolation of the Cq values to that of the
synthetic miRNA standard curves and adjusted for RT-qPCR efficiency
variation. The 2−ΔΔCq method was used for quantification
(29).
Data normalization and statistical
analysis
A set of three endogenous reference miRNAs were
identified using geNorm and NormFinder (30,31). The
geometric mean of the three normalizers was used to normalize the
miRNA expression data across subjects. Student's t-test was applied
to compare miRNA expression between cancer and control groups, with
false discovery rate correction using Bonferroni-type comparison
procedures (32). The Sequential
Forward Floating Search algorithm (33) was used to optimize the biomarker
selection during the internal cross-validation process for the
discovery study and AUC values were set as the optimization target.
The linear support vector machine was used to construct
multi-variant biomarker panels with optimal performance in
classifying control and cancer groups. Demographic data and
clinical characteristics of the three patient groups were compared
using one-way analysis of variance. Bonferroni's test was used for
post hoc analysis. P<0.05 was considered to indicate statistical
significance. The Stata v14 statistical programme (StataCorp) was
used for analyses and values expressed as n (%) or median (range).
For values represented as n (%), a student's t-test was performed
for the normally distributed continuous variables, and P<0.05
was considered significant.
Bioinformatics analysis
Dysregulated miRNAs were considered if they had a
significance of P<0.01 and absolute fold change of >1.5. The
DIANA-mirPath v.3 tool was used for interaction and pathway
analyses (34). The tool uses
Tarbase v.8 to identify miRNA-mRNA interactions and probes the
Kyoto Encyclopedia of Genes and Genomes for pathway analysis
(35,36). GeneMania and HMDD v3.0 were used to
visualize interactions of target genes and observe
miRNA-interacting partners, respectively (37,38). The
highlighting of key genes of interest on pathways and network was
drawn using Microsoft PowerPoint v2013 (Microsoft).
Results
Clinicopathological characteristics of
study participants
The study included 208 subjects, comprising 112
patients with PDAC, 50 patients with CP and 46 patients with
critical limb ischaemia as a CG. The demographic features and
traditional risk factors of the subjects are provided in Table I. In the PDAC group, there were 15
patients in Group 1, 57 patients in Group 2 and 40 in Group 3. The
6-month overall survival rate for Groups 1, 2 and 3 was 80.0, 91.2,
and 32.5%, respectively (Table SI).
There were 35 patients with survival for <6 months, 27 of which
were from Group 3. The body mass index of all of the patients was
determined and was not significantly different between the three
study groups.
 | Table I.Demographic and clinical
characteristics of patients recruited for the study. |
Table I.
Demographic and clinical
characteristics of patients recruited for the study.
| Variable | PDAC (n=112) | CP (n=50) | Control (n=46) | P-value (PDAC vs.
CP) | P-value (PDAC vs.
control) |
|---|
| Male/female
ratio | 1.65:1 |
4.2:1 | 1.4:1 | 0.179 | 0.901 |
| Agea, years | 59 (33–85) | 52 (38–69) | 61 (35–89) | P<0.001 | 0.704 |
|
Chronicb
illness | 69 (62) | 16 (32) | 31 (67.3) | 0.095 | 0.692 |
| Smoking
statusb |
|
|
|
|
|
| Ever
smoked | 72 (64) | 44 (88) | 26 (57.1) | P<0.003 | 0.120 |
| Current
smoking | 29 (26) | 34 (68) | 7 (15) | P<0.001 | P<0.02 |
| Alcohol
useb | 78 (70) | 45 (90) | 23 (50) |
P<0.004 | 0.088 |
| Body mass
indexa,
kg/m2 | 21.4
(15.6–29.2) | 19.0
(14.8–24.5) | 22.1
(16.1–26.3) | 0.366 | 0.423 |
Differential expression of miRNA
miRNA expression was compared between the PDAC group
and the CG and 42 significantly upregulated and 42 downregulated
miRNAs were identified (P<0.01; Table SII). When comparing the PDAC group
to both the control and chronic pancreatitis groups combined
together (PDAC vs. CG+CP), 16 significantly upregulated and 16
downregulated miRNAs in PDAC were identified (P<0.01; Fig. 1; Table
SIII). Of note, the downregulated miRNAs included members of
the let-7 family, including Homo sapiens (hsa)-let-7f-5p and
hsa-let-7a-5p. Comparison of the PDAC group with solely the CP
group provided only three significantly upregulated and four
downregulated miRNAs (P<0.01; Table
SIV).
Let-7 miRNAs target the INSR and IGF
signalling pathways
Using a targeted bioinformatics analysis, it was
demonstrated that the downregulated and upregulated miRNAs targeted
several components of the insulin pathway. However, only the
downregulated miRNAs were observed to target INSR (Figs. 2 and 3). Amongst the downregulated miRNAs,
members of the let-7 family, hsa-let-7f-5p and hsa-let-7a-5p, were
implicated in targeting INSR.
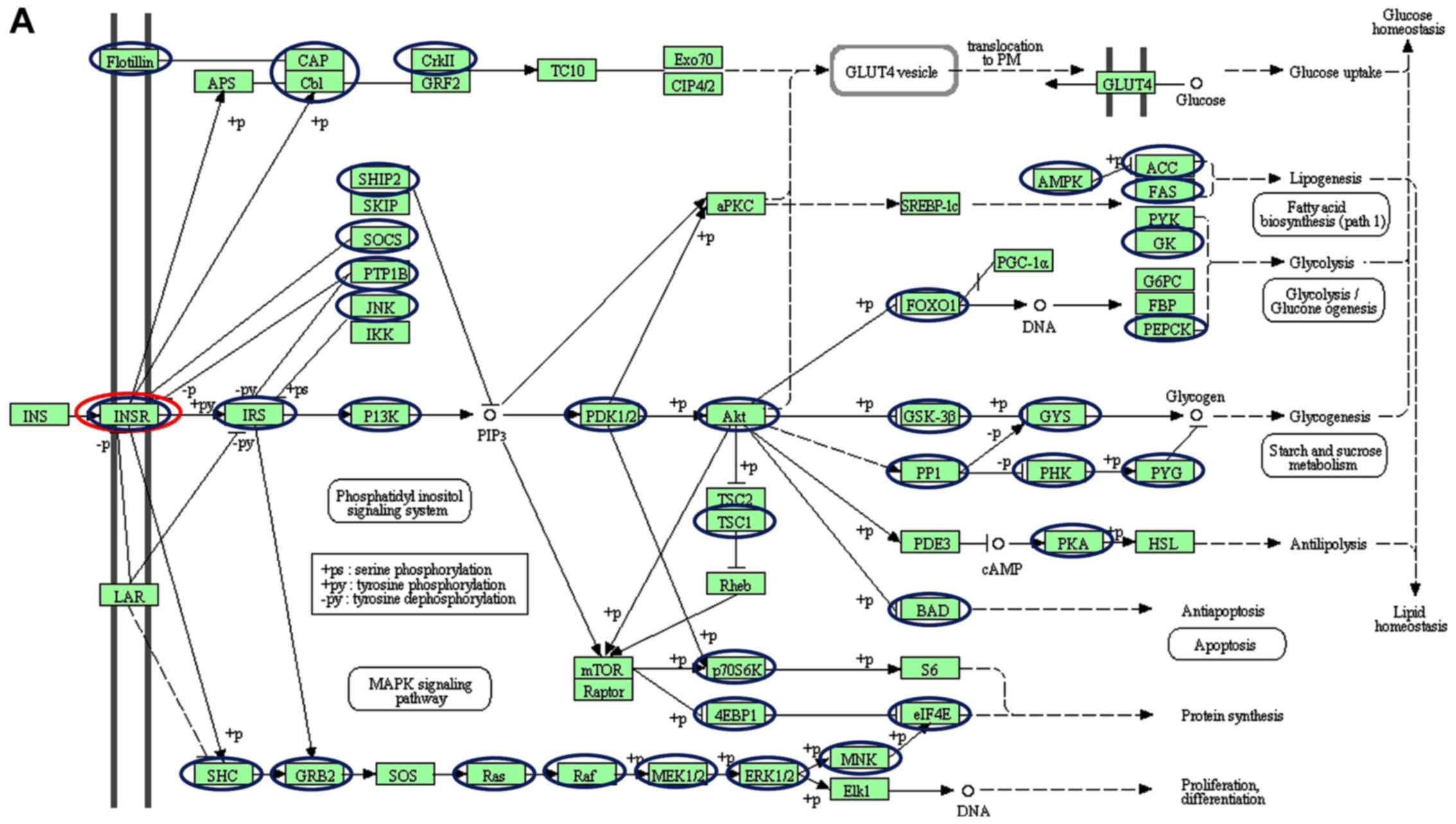 | Figure 2.Schematic of the insulin signalling
pathways, including the different genes targeted by (A)
downregulated and (B) upregulated miRNAs. Blue and red nodes
indicate genes in the pathways determined by the KEGG software to
be targeted by dysregulated miRNAs. Of interest, INSR is only
targeted by the downregulated let-7 family (red nodes). The figure
was generated from the Kyoto Encyclopedia of Genes and Genomes
pathways. The nodes (circles) were drawn onto the KEGG-generated
figure using Microsoft Powerpoint v2013 to highlight targeted
genes. INSR, insulin receptor; miRNA, microRNA; CAP, catabolite
gene activator protein; Cb1; cannabinoid receptor interacting
protein 1; CRKII, CRK proto-oncogene II; GRF2, general regulatory
factor 2; TC10, TC10 protein; EXO70; exocyst complex protein; CIP,
Cdc42-interacting protein; GLUT4, glucose transporter type 4; Lar,
low antibody response; SHIP2 (INPPL1), inositol polyphosphate
phosphatase like 1; SKIP, skeletal muscle and kidney-enriched
inositol phosphatase; SOCS (CISH), cytokine inducible SH2
containing protein; PTPIB, peptidylprolyl isomerase B; JNK, c-Jun
NH2-terminal kinase; IKK, I-κ B kinase; P13K, phosphoinositide
3-kinase; αPKC, protein kinase Cα; PDK, pyruvate dehydrogenase
kinase; SREBP-1C, SREBP-sterol regulatory element binding protein
1C; PRKAA1 (AMPK), protein kinase AMP-activated catalytic subunit
α1; ACC, acetyl-CoA carboxylase; FAS, Fas cell surface death
receptor; PyK, pyruvate kinase; GK, glycerol kinase; G6PC,
glucose-6-phosphatase catalytic subunit; FBP,
fructose-1,6-bisphosphatase 1; PEPCK, phosphoenolpyruvate
carboxykinase; PGC-1α, peroxisome proliferator-activated receptor γ
coactivator 1-α; FOXO1, Forkhead box protein O1; AKT (PKB), protein
kinase B; GSK3B, glycogen synthase kinase 3β; GYS, glycogen
synthase; PP1, serine/threonine protein phosphatase; PHKA2 (PHK),
phosphorylase kinase regulatory subunit α 2; PYG, glycogen
phosphorylase; TSC1, TSC complex subunit 1; PDE3, phosphodiesterase
3; PKA, protein kinase; HSL, hormone-sensitive lipase; BAD,
BCL2-associated agonist of cell death; RPS6KB1 (P70S6K), ribosomal
protein S6 kinase B1; MTOR, mechanistic target of rapamycin kinase;
EIF4EBP1 (4EBP1), eukaryotic translation initiation factor 4E
binding protein 1; S6, ribosomal protein S6; EIF4E, eukaryotic
translation Initiation factor 4E; MNK, mitogen-activated protein
kinase (MAPK) interacting protein kinase; ELK1, ETS transcription
factor; ERK, extracellular signal-regulated kinase; MEK (MAP2K),
mitogen-activated protein kinase kinase; RAF, RAF proto-oncogene
serine/threonine-protein kinase; RAS proto-oncogene; GRB2, growth
factor receptor bound protein 2; SHC, SHC adaptor protein 1; SOS,
SOS Ras/Rac guanine nucleotide exchange factor. |
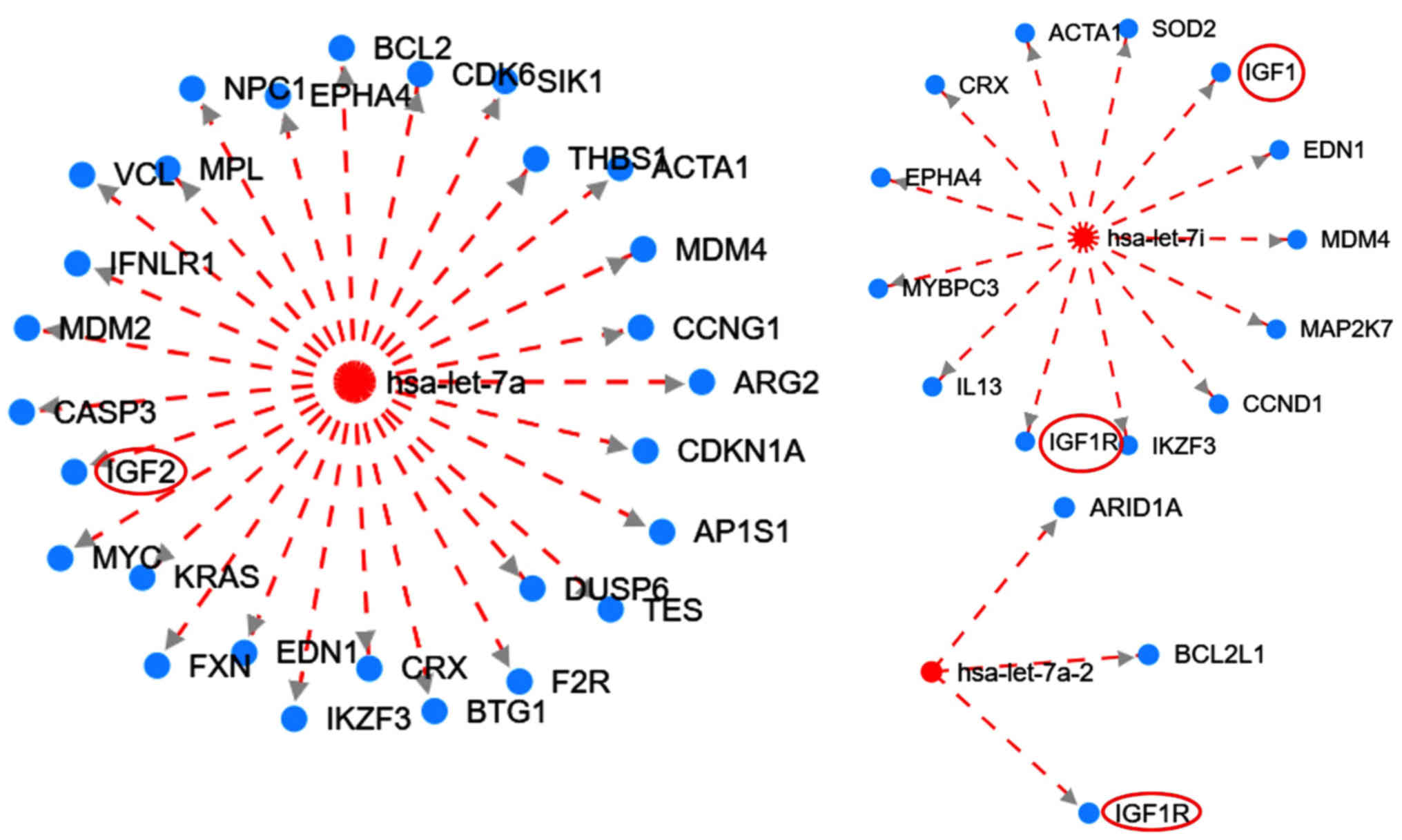 | Figure 3.Interaction network of the let-7
family targeting the IGF family members generated from HMDD version
3. Red circles (added with Microsoft PowerPoint v2013) indicate
members of the IGF signaling pathway. The broken red lines indicate
downregulation. Red and blue nodes are microRNAs and genes,
respectively. IGF1R, type 1 insulin-like growth factor receptor;
IGF, insulin-like growth factor; ARID1A, AT-rich interaction domain
1A; BCL2L1, BCL2 like 1; EDN1, endothelin 1; MDM4, MDM4 regulator
of p53; MAP2K7, mitogen-activated protein kinase kinase 7; CCND1,
cyclin D1; IKZF3, IKAROS family zinc finger 3; IL13, interleukin
13; MYBPC3, myosin binding protein C3; EPHA4, EPH receptor A4; CRX,
cone-rod homeobox; ACTA1, actin α1 skeletal muscle; SOD2,
superoxide dismutase 2; BCL2, BCL2 apoptosis regulator; CDK6,
cyclin dependent kinase 6; SIK1, salt inducible kinase 1; THBS1,
thrombospondin 1; CCNG1, cyclin G1; ARG2, arginase 2; CDKN1A,
cyclin dependent kinase inhibitor 1A; AP1S1, adaptor related
protein complex 1 subunit σ1; DUSP6, dual specificity phosphatase
6; TES, testin LIM domain protein; F2R, coagulation factor II
thrombin receptor; BTG1, BTG anti-proliferation factor 1; FXN,
frataxin; KRAS, KRAS proto-oncogene; MYC, MYC proto-oncogene;
CASP3, caspase 3; MDM2, MDM2 proto-oncogene; IFNLR1, interferon λ
receptor 1; VCL, vinculin; MPL, MPL proto-oncogene; EPHA4, EPH
receptor A4; hsa, Homo sapiens. |
Furthermore, the HDMM tool was used to illustrate
the interaction between the let-7 family of miRNAs with several
genes, including members of the IGF signalling pathway (Fig. 3). The analysis suggested that let-7
targets the IGF signalling pathway for downregulation. Let-7 has an
inhibitory effect on the IGF pathway and is downregulated,
suggesting the subsequent upregulation of this pathway.
Expression of the let-7 family in
different stages of PDAC
The levels of let-7 miRNAs were differentially
expressed in different groups of PDAC (Table SVa). Further analysis across the
PDAC groups indicated that 5 miRNAs were significantly upregulated
in Group 3 compared to Groups 1 and 2 combined (P<0.05; Table SVb). Although the differences in
expression levels were not significant between ‘Group 3 vs. 1+2’,
certain members of the let-7 miRNAs were differentially expressed
across the various groups. When comparing Group 3 to a combination
of Groups 1 and 2, hsa-let-7b-5p, hsa-let-7a-3p and hsa-let-7d-3p
were upregulated and hsa-let-7a-5p and hsa-let-7f-5p were
downregulated. Furthermore, hsa-let-7b-3p, hsa-let-7d-3p,
hsa-let-7a-5p and hsa-let-7e-3p were upregulated in Group 3 vs.
Group 2 but downregulated in Group 2 vs. 1. On the other hand,
hsa-let-7a-5p and hsa-let-7f-5p were downregulated in Group 3 vs. 2
but upregulated in Group 2 vs. 1. The potential prognostic value of
the let-7 miRNAs was also assessed by comparing levels observed in
patients with a ‘poor prognosis’ and those with a ‘good prognosis’
(Table SI). A poor or good
prognosis was defined by the survival of patients in months; and
since patients were followed up for only one year, those with a
survival time of <12 months were categorized as having a ‘poor
prognosis’. Comparing patients in the poor vs. good prognosis
group, a total of 20 miRNAs were associated with poor prognosis, as
they were significantly upregulated in the poor prognosis group
(P<0.05). However, none of the let-7 family members was
significantly dysregulated (Table
SVI).
Discussion
In the present study, the number of dysregulated
miRNAs in patients with PDAC compared with the CG was higher than
that in the PDAC compared with the CP group. The decreased number
of significantly dysregulated miRNAs between the CP and the PDAC
group may be due to shared pathological molecular mechanisms, as CP
is a risk factor for the development of PDAC (39). The present study indicated that
members of the let-7 miRNA family are dysregulated in PDAC. Of
note, the let-7 family members were significantly downregulated in
patients with PDAC. Let-7a-5p and let-7f-5p were also downregulated
in the late stages of PDAC. This finding corroborates the
observation that downregulation of let-7 in gemcitabine-resistant
pancreatic cancer cells was associated with activation of the
epithelial-to-mesenchymal pathway characteristic of metastasis
(40).
The let-7 family inhibits the expression of
components of the INSR/IGF signalling pathway (Figs. 2 and 3) and their downregulation may result in
increased activity of this pathway. This suggests that
hypothetically, an increase in let-7 may inhibit the INSR/IGF
signalling pathway and subsequently inhibit cellular proliferation
and promote chemotherapeutic sensitivity (Fig. 4). The let-7 family was initially
identified in the nematode Caenorhabditis elegans. In
humans, the let-7 family has 13 members, which have been
demonstrated to have a significant role in the process of
carcinogenesis (41). In 2016,
Encarnación et al (42)
indicated that overexpression of let-7b in patients with breast
cancer resulted in increased DNA repair capacity. Aberrant DNA
repair capacity is characteristic of PDAC (43,44).
Previous studies have suggested that expression of the let-7 family
was significantly reduced in PDAC tumor cells as compared with
normal acinar cells and that induction of let-7 expression
inhibited cellular proliferation (45). One study indicated that treatment
with diflourinated curcumin, a curcumin analogue with anti-oxidant
properties, inhibited tumor growth and this was associated with
increased levels of let-7 (46).
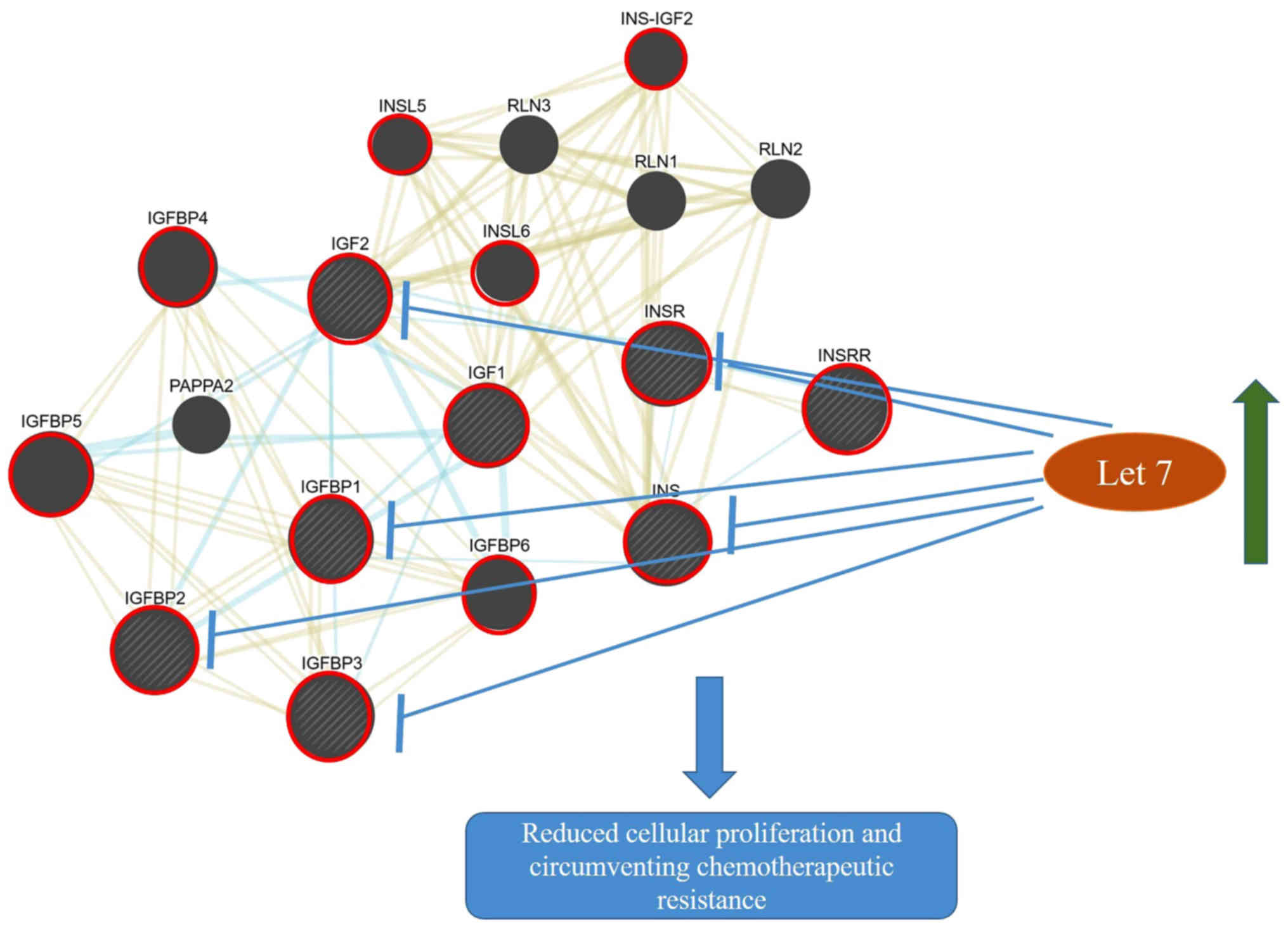 | Figure 4.Crosstalk between the insulin/IGF
signalling pathways and the potential targeting of let-7. The
upregulation of let-7 microRNAs may block the overexpression of
components of the INSR/IGF pathways resulting in reduced cell
proliferation and increase sensitivity to therapy. Blue and yellow
networks indicate shared pathways (INSR/IGF pathway) and protein
domains (N- and C-domains), respectively. The node sizes are
inversely proportional to the gene score rank determined by
GeneMANIA. Shaded nodes indicate genes inputted into the software.
Red circles are used to highlight key genes involved in the
INSR/IGF pathway. The interacting network was generated from
GeneMania. Microsoft PowerPoint v2013 was also used to make
additional drawings including let-7 node, arrows indicating
upregulation, inhibition and the subsequent possible effect of this
inhibition INSR, insulin receptor; IGF1, insulin-like growth factor
1; IGF2, insulin-like growth factor 2; IGFBP, Insulin-like growth
factor-binding protein; INS-IGF2, Insulin, isoform 2; INSL,
Insulin-like; INSRR, insulin receptor-related receptor; INS,
insulin; PAPPA2, pappalysin-2; RLN, relaxin. |
Early-onset diabetes and obesity are risk factors
for PDAC (47). It is known that
obesity-linked upregulation of the insulin/IGF signalling pathway
occurs in carcinogenesis via an increase in the NF-κβ pathway,
which induces inflammation, cellular proliferation, migration and
metastasis (48). A study on
transgenic let-7 mice indicated decreased body weight and increased
glucose levels linked to inhibition of components of INSR/IGF and
the PI3K/mTOR pathway (49).
Several receptors of the insulin and IGF family such
as INSR and IGF-1R were also indicated to be targeted for
downregulation by let-7 members (Figs.
2 and 3). The receptors of
insulin and IGF are related, belonging to a family of receptor
tyrosine kinases. Each has two receptors, INSR-A and INSR-B for
insulin and IGF-1R and IGF-2R for IGF (50,51).
These receptors are overexpressed in PDAC and exhibit complex
cross-talk with each other (52).
The INSR potentiates tumor progression, as it was indicated to be
significantly overexpressed along with progression and increased
tumor stage (53). The IGF-1R is
primarily responsible for mediating cellular responses to the IGFs
(54). Both INSR and IGF-1R are
highly homologous and are able to hybridize, enhancing tumor growth
(53). In a recent study,
overexpression of IGF-1R was observed in urothelial carcinoma,
indicating its utility as a therapeutic target (55). Hence, IGF-1R overexpression
correlates with the tumor stage and its co-expression with EGFR has
been associated with reduced overall survival (5). Similarly, IGF-2R is overexpressed in
PDAC compared to normal ductal cells (56). Knockdown of the IGFR pathway in a
mouse model inhibited cellular proliferation and promoted apoptosis
of PDAC cells (57). Regrettably,
several drugs developed to target both INSRs and IGF have been
unsuccessful in inhibiting PDAC development, possibly due to
heterodimerization of the IR and its interaction with IGF-1R,
resulting in resistance to these drugs (13).
The present study further suggested that let-7
targets IGF-binding proteins (IGFBPs) which tightly regulate the
expression of IGFs. The IGFBPs were determined to be elevated in
patients with PDAC compared to those with chronic pancreatitis
(58) and may contribute to
exacerbating pancreatic tumourigenesis. IGF2BP1 was indicated to
promote cellular proliferation in pancreatic cancer cells via the
AKT signalling pathway (59). A
recent study performed by our group confirmed the upregulation of
the Akt pathway in resected pancreatic tumours (17). Similarly, IGF2BP2 expression was
determined to promote tumour progression and to be correlated with
poor survival of patients with pancreatic cancer (60). It may therefore be concluded that
IGFBPs may serve as a biomarker for the disease. Since let-7
targets IGFBPs, it may be hypothesized that strategies targeting
let-7 may provide an avenue for evading therapeutic resistance
observed in pancreatic cancer treatment.
It has also been suggested that IGF signalling is
important in the formation of the dense avascular stroma
characteristic of PDAC (12),
another crucial factor involved in chemotherapy resistance
(61). Therefore, upregulation of
let-7 may inhibit stromal formation, increasing tumor
chemosensitivity.
In conclusion, the present study demonstrated
downregulation of certain members of the let-7 family in plasma
samples of patients with PDAC compared to patients with CP, a
pre-malignant condition, as well as to a CG with significant risk
factors common to patients with PDAC. Since the in silico
analysis indicated that let-7 targets the INSR/IGF pathway, its
downregulation may increase the expression of the INSR/IGF pathway
and may therefore be an effective target for the development of
INSR/IGF pathway-specific treatment strategies. Future studies
investigating the effects of let-7 in modulating the IGF pathway in
pancreatic cancer by using in vitro and in vivo
systems should be performed to validate this hypothesis.
Additionally, it would be pertinent to conduct a longer follow-up
Kaplan Meier analyses for overall survival between patients that
have high or low let-7 expression.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank Dr Lihan Zhou and Dr
Cheng He of MiRXES PTY Ltd, National University Singapore, for
assistance with the miRNA analyses.
Funding
This study was funded by the South African National
Research Foundation (grant no. 91508, ref. no. CSUR13091741850).
EEN was funded by a South African Medical Research Council Grant
awarded to the Wits Common Epithelial Cancer Research Centre.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
MB was responsible for the conceptualization of the
study, drafted and revised the manuscript. EEN performed data
analyses, drafted and revised the manuscript. Both authors read and
approved the final manuscript.
Ethics approval and informed consent to
participate
A prospective study including black South African
participants was undertaken. Ethics approval was obtained from the
University of the Witwatersrand Human Research Ethics Committee
(Medical), certificate no. M130551. All patients recruited for the
study provided written informed consent prior to sample
collection.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Rawla P, Sunkara T and Gaduputi V:
Epidemiology of pancreatic cancer: Global trends, etiology and risk
factors. World J Oncol. 10:10–27. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ferrone CR, Brennan MF, Gonen M, Coit DG,
Fong Y, Chung S, Tang L, Klimstra D and Allen PJ: Pancreatic
adenocarcinoma: The actual 5-year survivors. J Gastrointest Surg.
12:701–706. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Trajkovic-Arsic M, Kalideris E and Siveke
JT: The role of insulin and IGF system in pancreatic cancer. J Mol
Endocrinol. 50:R67–R74. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Valsecchi ME, McDonald M, Brody JR, Hyslop
T, Freydin B, Yeo CJ, Solomides C, Peiper SC and Witkiewicz AK:
Epidermal growth factor receptor and insulinlike growth factor 1
receptor expression predict poor survival in pancreatic ductal
adenocarcinoma. Cancer. 118:3484–3493. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nandy D and Mukhopadhyay D: Growth factor
mediated signaling in pancreatic pathogenesis. Cancers (Basel).
3:841–871. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Faller BA and Burtness B: Treatment of
pancreatic cancer with epidermal growth factor receptor-targeted
therapy. Biologics. 3:419–428. 2009.PubMed/NCBI
|
|
8
|
Zielinski R, Przytycki PF, Zheng J, Zhang
D, Przytycka TM and Capala J: The crosstalk between EGF, IGF, and
Insulin cell signaling pathways-computational and experimental
analysis. BMC Syst Biol. 3:882009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bergmann U: Insulin-like growth factor I
overexpression in human pancreatic cancer: Evidence for autocrine
and paracrine roles. Cancer Res. 55:2007–2011. 1995.PubMed/NCBI
|
|
10
|
Chakravarti A, Loeffler JS and Dyson NJ:
Insulin-like growth factor receptor I mediates resistance to
anti-epidermal growth factor receptor therapy in primary human
glioblastoma cells through continued activation of phosphoinositide
3-kinase signaling. Cancer Res. 62:200–207. 2002.PubMed/NCBI
|
|
11
|
LeRoith D and Roberts Jr CT: The
insulin-like growth factor system and cancer. Cancer Lett.
195:127–137. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mutgan AC, Besikcioglu HE, Wang S, Friess
H, Ceyhan GO and Demir IE: Insulin/IGF-driven cancer cell-stroma
crosstalk as a novel therapeutic target in pancreatic cancer. Mol
Cancer. 17:662018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Singh P, Alex JM and Bast F: Insulin
receptor (IR) and insulin-like growth factor receptor 1 (IGF-1R)
signaling systems: Novel treatment strategies for cancer. Med
Oncol. 31:8052014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu W, Bloom DA, Cance WG, Kurenova EV,
Golubovskaya VM and Hochwald SN: FAK and IGF-IR interact to provide
survival signals in human pancreatic adenocarcinoma cells.
Carcinogenesis. 29:1096–1107. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ma J, Sawai H, Matsuo Y, Ochi N, Yasuda A,
Takahashi H, Wakasugi T, Funahashi H, Sato M and Takeyama H: IGF-1
mediates PTEN suppression and enhances cell invasion and
proliferation via activation of the IGF-1/PI3K/Akt signaling
pathway in pancreatic cancer cells. J Surg Res. 160:90–101. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Neid M, Datta K, Stephan S, Khanna I, Pal
S, Shaw L, White M and Mukhopadhyay D: Role of insulin receptor
substrates and protein kinase C-zeta in vascular permeability
factor/vascular endothelial growth factor expression in pancreatic
cancer cells. J Biol Chem. 279:3941–3948. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nweke E, Ntwasa M, Brand M, Devar J, Smith
M and Candy G: Increased expression of plakoglobin is associated
with upregulated MAPK and PI3K/AKT signalling pathways in early
resectable pancreatic ductal adenocarcinoma. Oncol Lett.
19:4133–4141. 2020.PubMed/NCBI
|
|
18
|
Khan MA, Zubair H, Srivastava SK, Singh S
and Singh AP: Insights into the role of microRNAs in pancreatic
cancer pathogenesis: Potential for diagnosis, prognosis, and
therapy. Adv Exp Med Biol. 889:71–87. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mattick JS and Gagen MJ: The evolution of
controlled multitasked gene networks: The role of introns and other
noncoding RNAs in the development of complex organisms. Mol Biol
Evol. 18:1611–1630. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gong L, Wang C, Gao Y and Wang J:
Decreased expression of microRNA-148a predicts poor prognosis in
ovarian cancer and associates with tumor growth and metastasis.
Biomed Pharmacother. 83:58–63. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ono S, Lam S, Nagahara M and Hoon DS:
Circulating microRNA biomarkers as liquid biopsy for cancer
patients: Pros and cons of current assays. J Clin Med. 4:1890–1907.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Vafaee F, Diakos C, Kirschner MB, Reid G,
Michael MZ, Horvath LG, Alinejad-Rokny H, Cheng ZJ, Kuncic Z and
Clarke S: A data-driven, knowledge-based approach to biomarker
discovery: Application to circulating microRNA markers of
colorectal cancer prognosis. NPJ Syst Biol Appl. 4:202018.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bratulic S, Gatto F and Nielsen J: The
translational status of cancer liquid biopsies. Regen Eng Transl
Med. Nov 25–2019. View Article : Google Scholar
|
|
24
|
Lawrie CH, Gal S, Dunlop HM, Pushkaran B,
Liggins AP, Pulford K, Banham AH, Pezzella F, Boultwood J,
Wainscoat JS, et al: Detection of elevated levels of
tumour-associated microRNAs in serum of patients with diffuse large
B-cell lymphoma. Br J Haematol. 141:672–675. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chun YS, Pawlik TM and Vauthey JN: 8th
Edition of the AJCC Cancer Staging Manual: Pancreas and
Hepatobiliary cancers. Ann Surg Oncol. 25:845–847. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Harris PA, Taylor R, Thielke R, Payne J,
Gonzalez N and Conde JG: Research electronic data capture
(REDCap)-A metadata-driven methodology and workflow process for
providing translational research informatics support. J Biomed
Inform. 42:377–381. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Conwell DL, Lee LS, Yadav D, Longnecker
DS, Miller FH, Mortele KJ, Levy MJ, Kwon R, Lieb JG, Stevens T, et
al: American pancreatic association practice guidelines in chronic
pancreatitis: Evidence-based report on diagnostic guidelines.
Pancreas. 43:1143–1162. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Löhr JM, Dominguez-Munoz E, Rosendahl J,
Besselink M, Mayerle J, Lerch MM, Haas S, Akisik F, Kartalis N,
Iglesias-Garcia J, et al: United European Gastroenterology
evidence-based guidelines for the diagnosis and therapy of chronic
pancreatitis (HaPanEU). United European Gastroenterol J. 5:153–199.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lanoix D, Lacasse AA, St-Pierre J, Taylor
SC, Ethier-Chiasson M, Lafond J and Vaillancourt C: Quantitative
PCR Pitfalls: The case of the human placenta. Mol Biotechnol.
52:234–243. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
St-Pierre J, Grégoire JC and Vaillancourt
C: A simple method to assess group difference in RT-qPCR reference
gene selection using GeNorm: The case of the placental sex. Sci
Rep. 7:169232017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen SY, Feng Z and Yi X: A general
introduction to adjustment for multiple comparisons. J Thorac Dis.
9:1725–1729. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Reneker J and Shyu CR: Applying sequential
forward floating selection to protein structure prediction with a
study of HIV-1 RP. AMIA Annu Symp Proc. 2006:10722006.
|
|
34
|
Vlachos IS, Zagganas K, Paraskevopoulou
MD, Georgakilas G, Karagkouni D, Vergoulis T, Dalamagas T and
Hatzigeorgiou AG: DIANA-miRPath v3.0: Deciphering microRNA function
with experimental support. Nucleic Acids Res. 43:W460–W466. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Karagkouni D, Paraskevopoulou MD,
Chatzopoulos S, Vlachos IS, Tastsoglou S, Kanellos I, Papadimitriou
D, Kavakiotis I, Maniou S, Skoufos G, et al: DIANA-TarBase v8: A
decade-long collection of experimentally supported miRNA-gene
interactions. Nucleic Acids Res. 46:D239–D245. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kanehisa M and Goto S: KEGG: Kyoto
Encyclopedia of genes and genomes. Nucleic Acids Res. 28:27–30.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Warde-Farley D, Donaldson SL, Comes O,
Zuberi K, Badrawi R, Chao P, Franz M, Grouios C, Kazi F, Lopes CT,
et al: The GeneMANIA prediction server: Biological network
integration for gene prioritization and predicting gene function.
Nucleic Acids Res. 38:W214–W220. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Huang Z, Shi J, Gao Y, Cui C, Zhang S, Li
J, Zhou Y and Cui Q: HMDD v3.0: A database for experimentally
supported human microRNA-disease associations. Nucleic Acids Res.
47:D1013–D1017. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kirkegård J, Mortensen FV and
Cronin-Fenton D: Chronic pancreatitis and pancreatic cancer Risk: A
systematic review and meta-analysis. Am J Gastroenterol.
112:1366–1372. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Li Y, VandenBoom TG, Kong D, Wang Z, Ali
S, Philip PA and Sarkar FH: Up-regulation of miR-200 and let-7 by
natural agents leads to the reversal of epithelial-to-mesenchymal
transition in gemcitabine-resistant pancreatic cancer cells. Cancer
Res. 69:6704–6712. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Boyerinas B, Park SM, Hau A, Murmann AE
and Peter ME: The role of let-7 in cell differentiation and cancer.
Endocr Relat Cancer. 17:F19–F36. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Encarnación J, Ortiz C, Vergne R, Vargas
W, Coppola D and Matta JL: High DRC levels are associated with
Let-7b overexpression in women with breast cancer. Int J Mol Sci.
17:8652016. View Article : Google Scholar
|
|
43
|
Perkhofer L, Illing A, Gout J, Frappart PO
and Kleger A: Precision medicine meets the DNA damage response in
pancreatic cancer. Oncoscience. 5:6–8. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
McWilliams RR, Bamlet WR, Cunningham JM,
Goode EL, de Andrade M, Boardman LA and Petersen GM: Polymorphisms
in DNA repair genes, smoking, and pancreatic adenocarcinoma risk.
Cancer Res. 68:4928–4935. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Torrisani J, Bournet B, du Rieu MC,
Bouisson M, Souque A, Escourrou J, Buscail L and Cordelier P: let-7
MicroRNA transfer in pancreatic cancer-derived cells inhibits in
vitro cell proliferation but fails to alter tumor progression. Hum
Gene Ther. 20:831–844. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Bao B, Ali S, Banerjee S, Wang Z, Logna F,
Azmi AS, Kong D, Ahmad A, Li Y, Padhye S and Sarkar FH: Curcumin
analogue CDF inhibits pancreatic tumor growth by switching on
suppressor microRNAs and attenuating EZH2 expression. Cancer Res.
72:335–345. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Bracci PM: Obesity and pancreatic cancer:
Overview of epidemiologic evidence and biologic mechanisms. Mol
Carcinog. 51:53–63. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Bowers LW, Rossi EL, O'Flanagan CH,
deGraffenried LA and Hursting SD: The role of the Insulin/IGF
system in cancer: Lessons learned from clinical trials and the
energy balance-cancer link. Front Endocrinol (Lausanne). 6:772015.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zhu H, Shyh-Chang N, Segrè AV, Shinoda G,
Shah SP, Einhorn WS, Takeuchi A, Engreitz JM, Hagan JP, Kharas MG,
et al: The Lin28/let-7 axis regulates glucose metabolism. Cell.
147:81–94. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Bailyes EM, Navé BT, Soos MA, Orr SR,
Hayward AC and Siddle K: Insulin receptor/IGF-I receptor hybrids
are widely distributed in mammalian tissues: Quantification of
individual receptor species by selective immunoprecipitation and
immunoblotting. Biochem J. 327:209–215. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Pollak M: Insulin and insulin-like growth
factor signalling in neoplasia. Nat Rev Cancer. 8:915–928. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Ohmura E, Okada M, Onoda N, Kamiya Y,
Murakami H, Tsushima T and Shizume K: Insulin-like growth factor I
and transforming growth factor alpha as autocrine growth factors in
human pancreatic cancer cell growth. Cancer Res. 50:103–107.
1990.PubMed/NCBI
|
|
53
|
Ulanet DB, Ludwig DL, Kahn CR and Hanahan
D: Insulin receptor functionally enhances multistage tumor
progression and conveys intrinsic resistance to IGF-1R targeted
therapy. Proc Natl Acad Sci USA. 107:10791–10798. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Brahmkhatri VP, Prasanna C and Atreya HS:
Insulin-Like growth factor system in cancer: Novel targeted
therapies. Biomed Res Int. 2015:5380192015. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Eich ML, Tregnago AC, Faraj SF, Palsgrove
DN, Fujita K, Bezerra SM, Munari E, Sharma R, Chaux A and Netto GJ:
Insulin-like growth factor-1 receptor expression in upper tract
urothelial carcinoma. Virchows Arch. 474:21–27. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Ishiwata T, Bergmann U, Kornmann M, Lopez
M, Beger HG and Korc M: Altered expression of insulin-like growth
factor II receptor in human pancreatic cancer. Pancreas.
15:367–373. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Tian X, Hao K, Qin C, Xie K, Xie X and
Yang Y: Insulin-like growth factor 1 receptor promotes the growth
and chemoresistance of pancreatic cancer. Dig Dis Sci.
58:2705–2712. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Karna E, Surazynski A, Orłowski K,
Łaszkiewicz J, Puchalski Z, Nawrat P and Pałka J: Serum and tissue
level of insulin-like growth factor-I (IGF-I) and IGF-I binding
proteins as an index of pancreatitis and pancreatic cancer. Int J
Exp Pathol. 83:239–245. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Wan BS, Cheng M and Zhang L: Insulin-like
growth factor 2 mRNA-binding protein 1 promotes cell proliferation
via activation of AKT and is directly targeted by microRNA-494 in
pancreatic cancer. World J Gastroenterol. 25:6063–6076. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Dahlem C, Barghash A, Puchas P, Haybaeck J
and Kessler SM: The insulin-like growth factor 2 mRNA binding
protein IMP2/IGF2BP2 is overexpressed and correlates with poor
survival in pancreatic cancer. Int J Mol Sci. 20:32042019.
View Article : Google Scholar
|
|
61
|
Rhim AD, Oberstein PE, Thomas DH, Mirek
ET, Palermo CF, Sastra SA, Dekleva EN, Saunders T, Becerra CP,
Tattersall IW, et al: Stromal elements act to restrain, rather than
support, pancreatic ductal adenocarcinoma. Cancer Cell. 25:735–747.
2014. View Article : Google Scholar : PubMed/NCBI
|















