Introduction
Laryngeal squamous cell carcinoma (LSCC) is a common
type of laryngeal cancer globally that frequently occurs in the
larynx and accounts for 95% of laryngeal cancer cases in the recent
decade (1–3). Surgical resection, chemotherapy,
radiotherapy and immunotherapy have been used in the treatment of
LSCC (4). However, the mortality
rate of patients with advanced LSCC remains high due to the
limitations of these therapeutic strategies, such as chemo- and
radio-resistance (5). The 5-year
overall survival rate of patients with LSCC is ~64% (5). In addition, some patients progress to
advanced stages of the disease due to disease recurrence and
metastasis (6). Thus, understanding
the pathogenesis and identification of novel therapeutic targets
are critical for the effective treatment of LSCC.
MicroRNAs (miRNAs/miRs) are a class of small
non-coding RNA molecules that are ~22 nucleotides in length
(7–9). miRNAs were primarily identified as key
regulators of gene expression by binding the 3′-untranslated region
(UTR) of target mRNAs (9).
Increasing evidence suggests that miRNAs serve important roles
across a variety of physiological and pathological conditions,
including cell proliferation, apoptosis and cell cycle progression
(10,11). Notably, the novel function of miRNAs
as potential biomarkers and therapeutic targets in the treatment of
different types of cancer is emerging (12–15).
miRNAs negatively modulate the expression of cancer-associated
genes, and thus regulate cancer progression (16,17).
Aberrant expression of miRNAs has been reported in LSCC, and is
associated with angiogenesis, tumor growth and metastasis (18–20). For
example, miR-143-3p suppresses the proliferation and invasion of
LSCC cells by targeting melanoma-associated antigen A9 (21). A recent study demonstrated that
miR-154 inhibits the progression of LSCC by regulating
N-acetylgalactosaminyltransferase 7 (GALNT7) (22). Furthermore, miR-4497 was identified
as a tumor suppressor in LSCC that negatively modulates
gastrulation brain homeobox 2 (GBX2) expression (23). Notably, a previous meta-analysis
predicted the potential function and clinical significance of
miR-375-3p in head and neck squamous cell carcinoma (HNSCC)
(24). In this study, a total of 21
studies involving 1,685 subjects were analyzed to evaluate the
relationship between miRNA and the prognosis of HNSCC (24). Significantly decreased expression of
miR-375-3p was found in HNSCC and was associated with poor
prognosis of patients (24).
However, the underlying molecular mechanism of miR-375-3p in LSCC
remains unknown.
Aerobic glycolysis, also known as the ‘Warburg
effect’, is considered the primary metabolic process for cancer
cells that facilitates cell proliferation under hypoxic conditions
(25–27). The glycolysis of cancer cells is
catalyzed by several key regulators, including glucose transporter
1 (GLUT1), lactate dehydrogenase A (LDHA) and hypoxia-inducible
factor 1α (HIF1α) (26). Previous
studies have reported that the hepatocyte nuclear factor 1β
(HNF-1β), also known as TCF2, is a key transcription factor that
promotes carcinogenesis by regulating glucose metabolism (28–30).
Overexpression of HNF1β has been observed in different types of
human cancer and is associated with the poor prognosis of patients
with cancer (31–34). A previous study demonstrated that
HNF1β is targeted by miRNAs and negatively modulated in cancer
cells (35).
The aim of this study was to explore the function
and mechanism of miR-375-3p in LSCC. The expression of miR-375-3p
in LSCC tissues and cells was detected using reverse-transcription
quantitative (RT-q)PCR. CCK-8 assay, cell apoptosis and cell cycle
analysis were performed to evaluate the effects of miR-375-3p on
the malignant behavior of LSCC cells. The results of the present
study indicated that miR-375-3p expression decreased in LSCC
tissues and cell lines. Furthermore, overexpression of miR-375-3p
inhibited LSCC cell proliferation. Functional analysis demonstrated
that miR-375-3p targeted HIF1β and inhibited its expression. Taken
together, these results provide a novel insight by which
miR-375/HNF1β signaling regulates the progression of LSCC.
Materials and methods
LSCC tissues, cell lines and
plasmids
A total of 50 paired LSCC tissues and matched
adjacent normal tissues (>5 cm from the margin of the LSCC
tissues) were collected from patients (20 female and 30 male; age
range, 39–74 years old; mean age, 58.5 years old) diagnosed with
LSCC at the First Affiliated Hospital of Zhengzhou University
between January 2012 to December 2014. Tissues were collected via
surgical resection and confirmed via histopathological analysis by
3 independent pathologists from the First Affiliated Hospital of
Zhengzhou University (Zhengzhou, China). Tissues were frozen in
liquid nitrogen and stored at −80°C before further experiments. The
present study was approved by the Ethical Institution of The First
Affiliated Hospital of Zhengzhou University (Zhengzhou, China;
approval no. 20120134-445) and performed in accordance with The
Declaration of Helsinki. All patients provided written informed
consent prior to the study start. Patients were divided into high
and low miR-375-3p expression groups with a median cut-off value of
3.68.
The LSCC cell lines, AMC-HN-8, Tu-177 and Tu-212
(the Tu-212 cell line used in the present study was authenticated
by STR profiling) were purchased from the American Type Culture
Collection, while the human bronchial epithelial 16 HBE cell line
(cat. no. scc150) was purchased from Sigma-Aldrich; Merck KGaA.
Cells were maintained in DMEM (Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS (Invitrogen; Thermo Fisher Scientific,
Inc.) at 37°C in 5% CO2.
The Flag-HIF1β plasmid was generated by amplifying
the full-length of HIF1β via PCR using the cDNA from AMC-HN-8 cells
with JumpStartTM Taq DNA polymerase (cat. no. D9307; Sigma-Aldrich;
Merck KGaA). The thermocycling conditions used were as follows:
initial denaturation at 95°C for 5 min followed by 33 cycles of
denaturation at 95°C for 30 sec, annealing at 58°C for 30 sec,
elongation at 72°C for 1 min and final extension at 72°C for 5 min.
The PCR products were inserted into the backbone of the Flag-vector
(Beijing Solarbio Science and Technology Co., Ltd) at the
restriction enzyme sites of NotI and ApaI. The
primers of HIF1β were designed as follows: forward,
5′-GCGGCGATGGCGGCGACTA-3′ and reverse,
5′-GGGCCCCTAGAGTTCCTGTTG-3′.
Transfection
Transfection of AMC-HN-8 and Tu-212 cells was
performed using Lipofectamine 2000 reagent (Invitrogen; Thermo
Fisher Scientific, Inc.). miR-375-3p mimic
(5′-UUUGUUCGUUCGGCUCGCGUGA-3′), miR-375-3p inhibitor
(5′-UCACGCGAGCCGAACGAACAAA-3′) and control miRNA
(5′-GGUUCGUACGUACACUGUUCA-3′) were obtained from the Guangzhou
RiboBio Co., Ltd. 50 nm of miRNA was diluted with 100 µl OPTI-MEM
(Invitrogen; Thermo Fisher Scientific, Inc.) and subsequently
incubated with 5 µl Lipofectamine 2000 for 15 min at room
temperature. Subsequent experimentation was performed 48 h
post-transfection.
RT-qPCR
Total RNA was extracted from LSCC tissues or cells
using TRIzol regent (Invitrogen; Thermo Fisher Scientific, Inc.),
and RNA concentration was measured using the NanoDrop 2000
spectrophotometer (Thermo Fisher Scientific, Inc.). RT was
performed using the Revert Aid First Strand cDNA synthesis kit
(Thermo Fisher Scientific, Inc.), with random primers at of 25°C
for 5 min, followed by 42°C for 60 min and 70°C for 5 min. The
level of miR-375-3p was assessed using the SYBR Green PCR Mixture
(Applied Biosystems; Thermo Fisher Scientific, Inc.) and ABI Prism
7900 detection system (Bio-Rad Laboratories, Inc.). The following
primer sequences were used for qPCR: miR-375-3p forward,
5′-CGGGTTTGTTCGTTCGGCT-3′ and reverse, 5′-GTGCAGGGTCCGAGGTATT-3′;
U6 forward, 5′-GCTTCGGCAGCACATATACT-3′ and reverse,
5′-GTGCAGGGTCCGAGGTATTC-3′; HNF1β forward,
5′-ACACACCTCCCATCCTCAAG-3′ and reverse, 5′-CATTTTAGCAGCCCTCCAAG-3′;
and GAPDH forward, 5′-AAATCCCATCACCATCTTCCAG-3′ and reverse,
5′-TGATGACCCTTTTGGCTCCC-3′. The following thermocycling conditions
were used for qPCR: initial denaturation at 95°C for 2 min followed
by 40 cycles of denaturation at 95°C for 15 sec, annealing and
elongation at 60°C for 30 sec. Relative expression levels were
calculated using the 2−∆∆Cq method (36) and normalized to the internal
reference genes U6 and GAPDH.
Cell proliferation assay
The proliferative rate of AMC-HN-8 and Tu-212 cells
transfected with miR-375-3p mimics or inhibitor was determined
using the cell counting Kit-8 (CCK-8) according to the
manufacturer's instructions. Cells were seeded into 96-well plates
at a density of 2,000 cells/well and cultured with Dulbecco's
Modified Eagle's Medium (DMEM) at 37°C overnight. Subsequently, 10
µl CCK-8 solution (Dojindo Molecular Technologies, Inc.) was added
into each well and further incubated for 4 h at 37°C. Cell
proliferation was analyzed at a wavelength of 450 nm, using a
microplate reader (Bio-Rad Laboratories, Inc.). All experiments
were performed in triplicate.
Target prediction
Targets of miR-375-3p were predicted using the miRDB
online database (http://mirdb.org/) by providing the
name of miRNA as ‘miR-375-3p’.
Dual-luciferase reporter assay
AMC-HN-8 and Tu-212 cells were seeded into 96-well
plates at a density of 1,000 cells/well and cultured with DEME
medium at 37°C overnight. Cells were subsequently co-transfected
with miR-375-3p mimic or inhibitor and the pmirGLO luciferase
vector (Promega Corporation) containing wild type (WT) or mutant
(MUT) 3′-UTR of HNF1β using Lipofectamine 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.). Following incubation for 48 h at 37°C,
cells were harvested and firefly and Renilla luciferase
activities were detected using a Dual-luciferase Reporter Assay
system (Promega Corporation), according to the manufacturer's
protocol. Firefly luciferase activity was normalized to
Renilla luciferase activity. All experiments were performed
in triplicate.
Western blotting
Total protein was extracted using RIPA lysis buffer
(Beyotime Institute of Biotechnology) containing protease
inhibitors and phosphatase inhibitors (Thermo Fisher Scientific,
Inc.). Protein concentration was determined using a bicinchoninic
acid (BCA) assay and an equal amount of protein (20 µg) was
separated by 15% SDS-PAGE. The separated proteins were subsequently
transferred onto PVDF western blotting membranes (Roche
Diagnostics) and blocked with 5% skim milk for 1 h at room
temperature. Membranes were washed twice with TBST (0.1% Tween-20)
and incubated with primary antibodies against HNF1β (1:1,000
dilution; cat. no. ab236759; Abcam) and GAPDH (1:2,000 dilution;
cat. no. 5174; Cell Signaling Technology, Inc.) overnight at 4°C.
Membranes were re-washed twice with TBST (0.1% Tween-20) and
incubated with goat anti-mouse (cat. no. 170-6516) or goat
anti-rabbit (cat. no. 170-6515) IgG (H+L)-horseradish
peroxidase-conjugated secondary antibodies (1:5,000 dilution;
Bio-Rad Laboratories, Inc.) at room temperature for 1 h. Protein
bands were visualized using the ECL Western Blotting Substrate
(Thermo Fisher Scientific, Inc.), according to the manufacturer's
protocol.
Flow cytometric analysis of
apoptosis
AMC-HN-8 and Tu-212 cells were seeded into 6-well
plates (1×105 cells/well) and transfected with
miR-375-3p mimics or inhibitor. After 48 h, cells were collected,
washed twice with PBS and stained with Annexin V (Invitrogen;
Thermo Fisher Scientific, Inc.) for 15 min in the dark at room
temperature. Cells were subsequently stained with propidium iodide
(PI) solution (Invitrogen; Thermo Fisher Scientific, Inc.) at room
temperature for 1 min. The apoptotic cells were analyzed using a
flow cytometer (BD FACSCalibur; BD Biosciences) and FlowJo software
version 10.6 (FlowJo LLC).
Glucose uptake
AMC-HN-8 and Tu-212 cells transfected with
miR-375-3p mimics or inhibitor were cultured in serum-free DMEM
medium at 37°C overnight. Glucose uptake was determined using the
Glucose Uptake Assay kit (Colorimetric, cat. no. ab136955; Abcam)
according to the manufacturer's instructions. Briefly, cells
(1×105) were washed twice with PBS and maintained in
KRPH (Krebs-Ringer phosphate HEPES)/2% BSA for 40 min at RT, prior
to incubation with 10 µl 2-deoxyglucose (2-DG) for 15 min at room
temperature. Standard 2-DG6P solutions (30 µl) or equal volumes of
media for miR-375-3p mimic or miR-control transfected cells were
added into 96-well plates, respectively. Subsequently, Assay buffer
(8 µl) and the enzyme mix (2 µl) were added to the solution and
incubated for 1 h at room temperature. Extraction buffer (90 µl)
was added and heated at 90°C for 30 min, prior to incubation with
reaction buffer B (35 µl) at room temperature for 5 min. Glucose
uptake was analyzed at a wavelength of 412 nm, using a microplate
reader (Bio-Rad Laboratories, Inc.). Total protein was quantified
using the BCA kit (Beyotime Institute of Biotechnology) according
to the manufacturer's protocol.
Lactate production
AMC-HN-8 and Tu-212 cells (1×105)
transfected with miR-375-3p mimics or inhibitor were seeded into
96-well plates and cultured with DMEM medium at 37°C. After 48 h,
lactate production was measured using the Lactate Assay kit (cat.
no. MAK064; Sigma-Aldrich; Merck KGaA), according to the
manufacturer's protocol. Cells were lysed and centrifuged at 10,000
× g for 10 min at 4°C. The supernatant (50 µl) was transferred into
the wells following deproteinization with a 10 kDa MWCO spin
filter. The lactate standard was established by adding 0, 2, 4, 6,
8 and 10 µl of 1 nmol/μl lactate solution to the lactate assay
buffer to make a final volume of 50 µl. Following incubation with
50 µl master reaction mix at room temperature for 30 min, lactate
production was analyzed at a wavelength of 570 nm, using a
microplate reader (Bio-Rad Laboratories, Inc.). Total protein was
quantified for each group using the BCA kit (Beyotime Institute of
Biotechnology), according to the manufacturer's protocol.
Statistical analysis
Statistical analysis was performed using GraphPad
Prism software (version 7.0; GraphPad Software Inc.) and data are
presented as the mean ± standard deviation. A paired Student's
t-test was used to compare differences between two groups. One-way
ANOVA followed by Tukey's post-hoc test were used to compare
differences between multiple groups. The χ2 test was
used to determine the association between miR-375-3p expression and
clinicopathological characteristics of patients with LSCC.
P<0.05 was considered to indicate a statistically significant
difference.
Results
miR-375-3p is downregulated in LSCC
tissues and cell lines
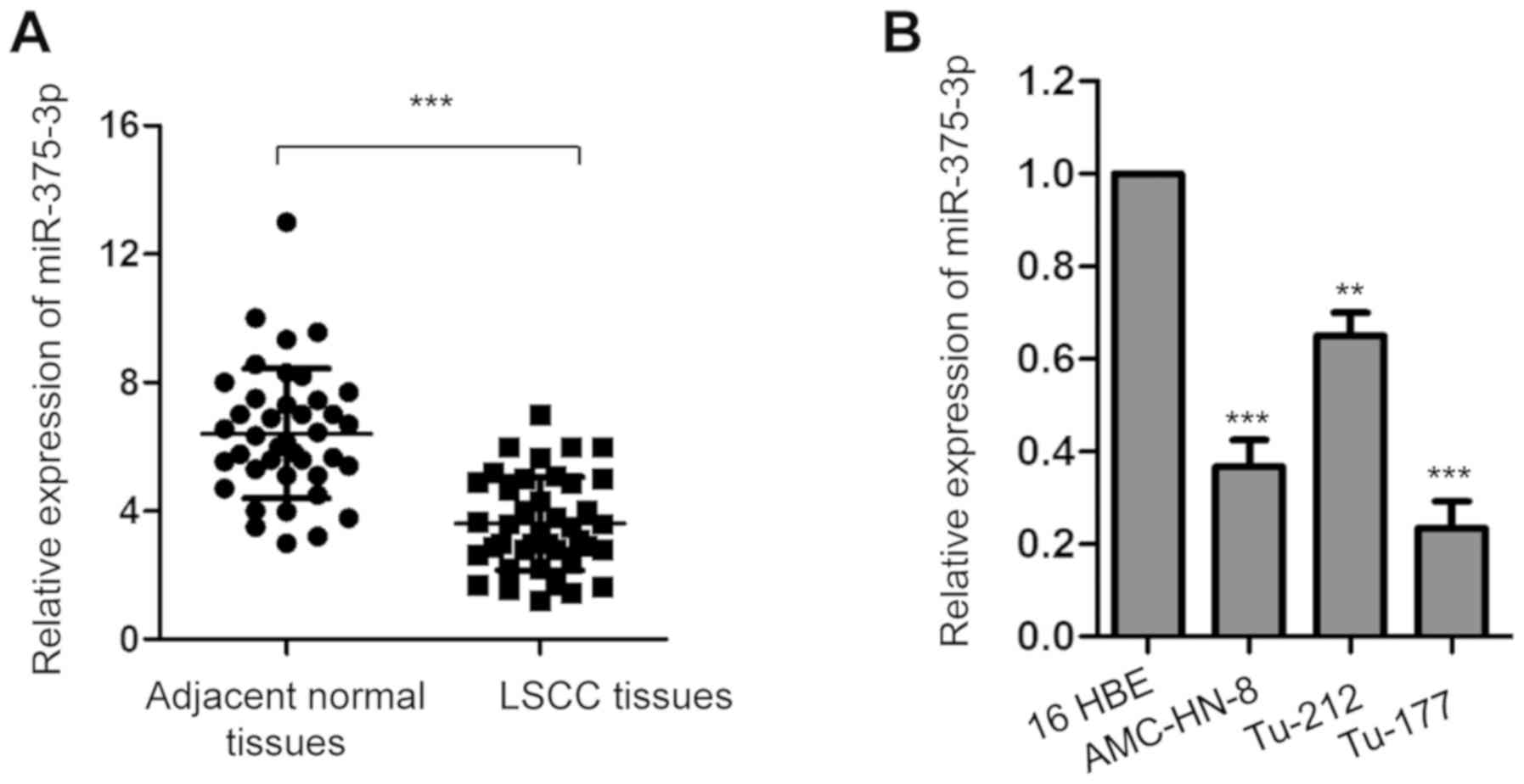
RT-qPCR analysis was performed to assess miR-375-3p
expression in 50 paired LSCC tissues and matched adjacent normal
tissues. miR-375-3p expression was significantly downregulated in
LSCC tissues compared with adjacent normal tissues (Fig. 1A; ***P<0.001). Consistently,
miR-375-3p expression was significantly downregulated in the LSCC
cell lines (AMC-HN-8, Tu-212 and Tu-177) compared with 16 HBE cells
(Fig. 1B; ***P<0.001).
In order to further determine the clinical
significance of miR-375-3p in LSCC, patients with LSCC were divided
into low and high miR-375-3p expression groups, respectively, with
a median miR-375-3p expression of 3.68 set as the cut-off value.
The results demonstrated that miR-375-3p expression was
significantly associated with tumor size, TNM stage (37), metastasis and histological grade
(38) of patients with LSCC
(Table I). Taken together, these
results indicate the potential involvement of miR-375-3p in
LSCC.
 | Table I.Association between miR-375-3p
expression and clinicopathological characteristics of patients with
laryngeal squamous cell carcinoma (n=50). |
Table I.
Association between miR-375-3p
expression and clinicopathological characteristics of patients with
laryngeal squamous cell carcinoma (n=50).
|
Characteristics | Number of patients,
n | High miR-375-3p
expression, n | Low miR-375-3p
expression, n | P-value |
|---|
| Age, years |
|
|
|
0.571 |
|
≤60 | 15 | 5 | 10 |
|
|
>60 | 35 | 10 | 25 |
|
| Sex |
|
|
|
0.632 |
|
Male | 26 | 8 | 18 |
|
|
Female | 24 | 7 | 17 |
|
| Tumor size, cm |
|
|
|
<0.001a |
| ≤4 | 25 | 13 | 12 |
|
|
>4 | 25 | 2 | 23 |
|
| Lymph node
metastasis |
|
|
|
<0.001a |
|
Negative | 22 | 10 | 12 |
|
|
Positive | 28 | 5 | 23 |
|
| Histological
grade |
|
|
|
<0.001a |
|
High | 30 | 10 | 20 |
|
|
Poor | 20 | 5 | 15 |
|
| TNM stage |
|
|
|
<0.001a |
|
I–II | 26 | 11 | 15 |
|
|
III–IV | 24 | 4 | 20 |
|
miR-375-3p regulates the proliferation
and apoptosis of AMC-HN-8 and Tu-212 cells
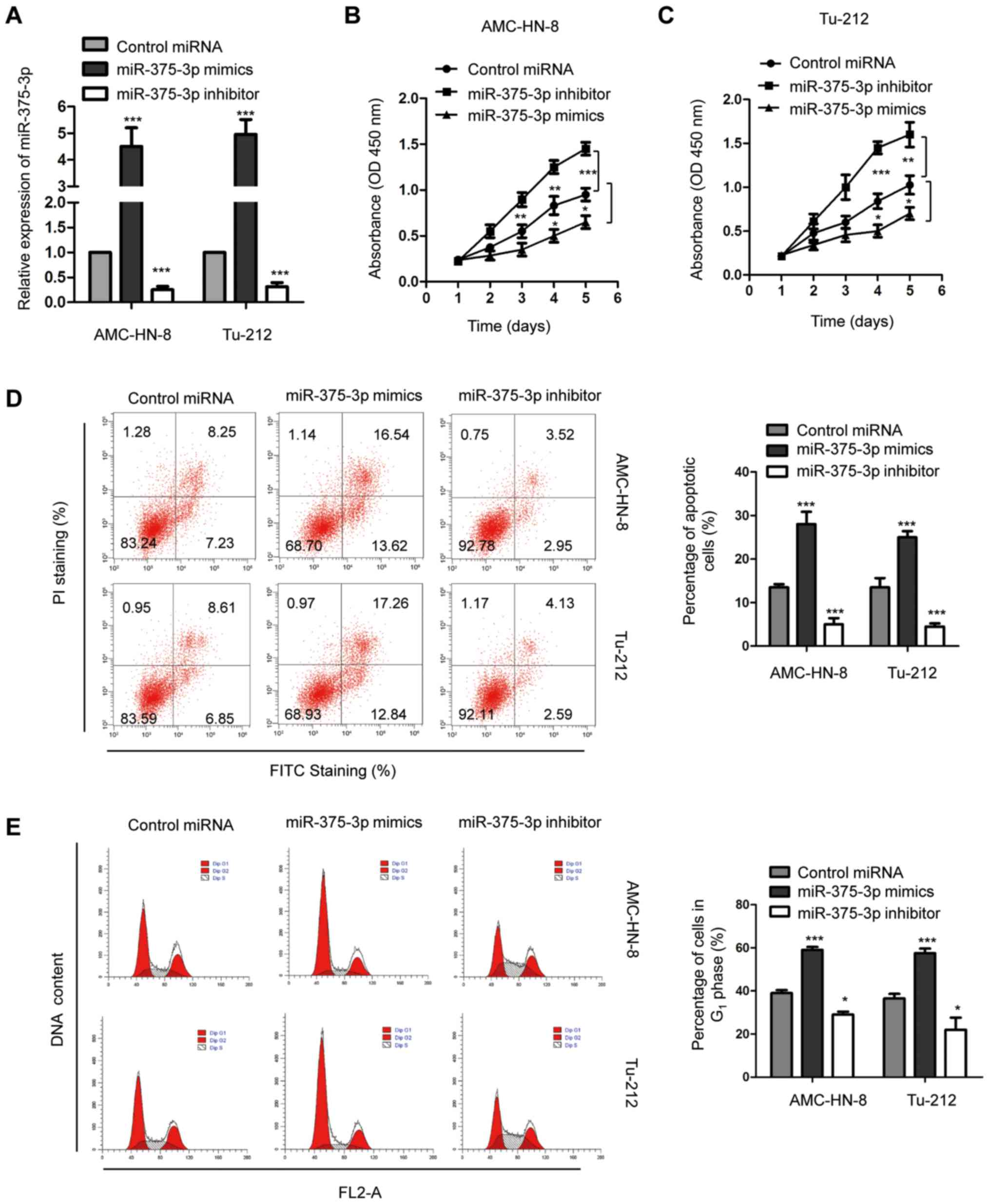
In order to further investigate the biological
function of miR-375-3p in LSCC, both AMC-HN-8 and Tu-212 cells,
which are widely used in the study of LSCC (39), were transfected with miR-375-3p
mimics or inhibitor, and transfection efficiency was determined via
RT-qPCR analysis (Fig. 2A;
***P<0.001). The effect of miR-375-3p on the proliferative rate
of AMC-HN-8 and Tu-212 cells was assessed via the CCK-8 assay.
Overexpression of miR-375-3p significantly inhibited the
proliferation of both AMC-HN-8 and Tu-212 cells, while miR-375-3p
knockdown promoted the proliferation of AMC-HN-8 and Tu-212 cells
(Fig. 2B and C; **P<0.01;
***P<0.001). Flow cytometric analysis was performed to detect
the effect of miR-375-3p on the apoptosis of AMC-HN-8 and Tu-212
cells. The result demonstrated that apoptosis of both AMC-HN-8 and
Tu-212 cells significantly increased following transfection with
miR-375-3p mimics compared with the control cells (Fig. 2D; ***P<0.001). Conversely,
miR-375-3p knockdown significantly decreased apoptosis of AMC-HN-8
and Tu-212 cells (Fig. 2D;
***P<0.001). Furthermore, overexpression of miR-375-3p
significantly increased the proportion of cells in the
G0/G1 phase (Fig.
2E; *P<0.05; ***P<0.001), suggesting that increased
miR-375-3p expression induces G1 cell cycle arrest.
miR-375-3p knockdown facilitated cell cycle progression from
G1 to S phase (Fig. 2E;
*P<0.05; ***P<0.001). Collectively, these results suggest the
potential tumor suppressive role of miR-375-3p in regulating
AMC-HN-8 and Tu-212 cell proliferation.
miR-375-3p targets HNF1β and decreases
its expression in AMC-HN-8 and Tu-212 cells
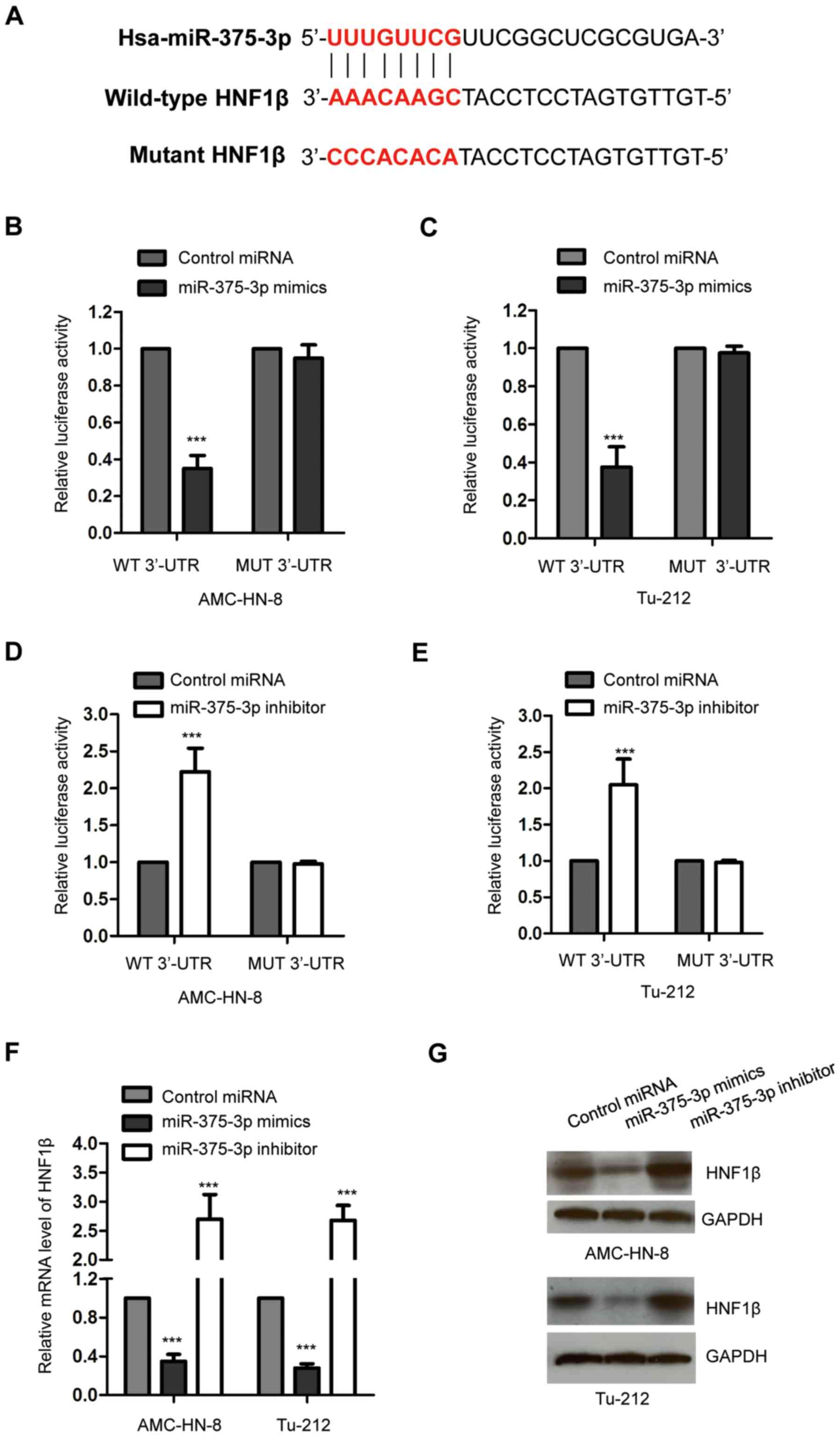
The potential targets of miR-375-3p were predicted
using the online miRDB database, in order to better understand the
molecular mechanism by which miR-375-3p negatively regulates the
progression of LSCC. The results indicated that the 3′-UTR of HNF1β
contains a putative complementary binding site of miR-375-3p
(Fig. 3A). In order to verify that
HNF1β is a target of miR-375-3p, both AMC-HN-8 and Tu-212 cells
were transfected with miR-375-3p mimics and the luciferase reporter
vector harboring WT or MUT 3′-UTR of HNF1β. The results
demonstrated that overexpression of miR-375-3p significantly
decreased the luciferase activity of cells expressing the WT but
not the MUT 3′-UTR of HNF1β (Fig. 3B and
C; ***P<0.001). This indicated the specific binding between
miR-375-3p and the 3′-UTR of HNF1β. In order to confirm these
results, both AMC-HN-8 and Tu-212 cells were transfected with
miR-375-3p inhibitor to decrease miR-375-3p expression. The
dual-luciferase reporter assay demonstrated that decreased
miR-375-3p expression significantly increased the luciferase
activity of cells harboring WT 3′-UTR of HNF1β (Fig. 3D and E; ***P<0.001). Taken
together, these results confirm the binding between miR-375-3p and
the 3′-UTR of HNF1β.
RT-qPCR analysis was performed to determine whether
the binding of miR-375-3p affected the stability of HNF1β mRNA in
AMC-HN-8 and Tu-212 cells transfected with miR-375-3p mimics or
inhibitor. The results demonstrated that overexpression of
miR-375-3p significantly decreased HNF1β mRNA expression, while
miR-375-3p knockdown significantly increased HNF1β mRNA expression
in AMC-HN-8 and Tu-212 cells compared with cells expressing
control-miRNA, respectively (Fig.
3F; ***P<0.001). Western blot analysis was performed to
assess HNF1β protein levels in AMC-HN-8 and Tu-212 cells
transfected with miR-375-3p mimics or inhibitor. The results
indicated that overexpression of miR-375-3p decreased HNF1β protein
expression, while miR-375-3p knockdown increased HNF1β protein
expression in both AMC-HN-8 and Tu-212 cells compared with cells
expressing control-miRNA, respectively (Fig. 3G). Collectively, these results
suggest that miR-375-3p targets HNF1β and decreases it expression
in LSCC cells.
miR-375-3p negatively regulates the
glucose metabolism of AMC-HN-8 and Tu-212 cells by targeting
HNF1β
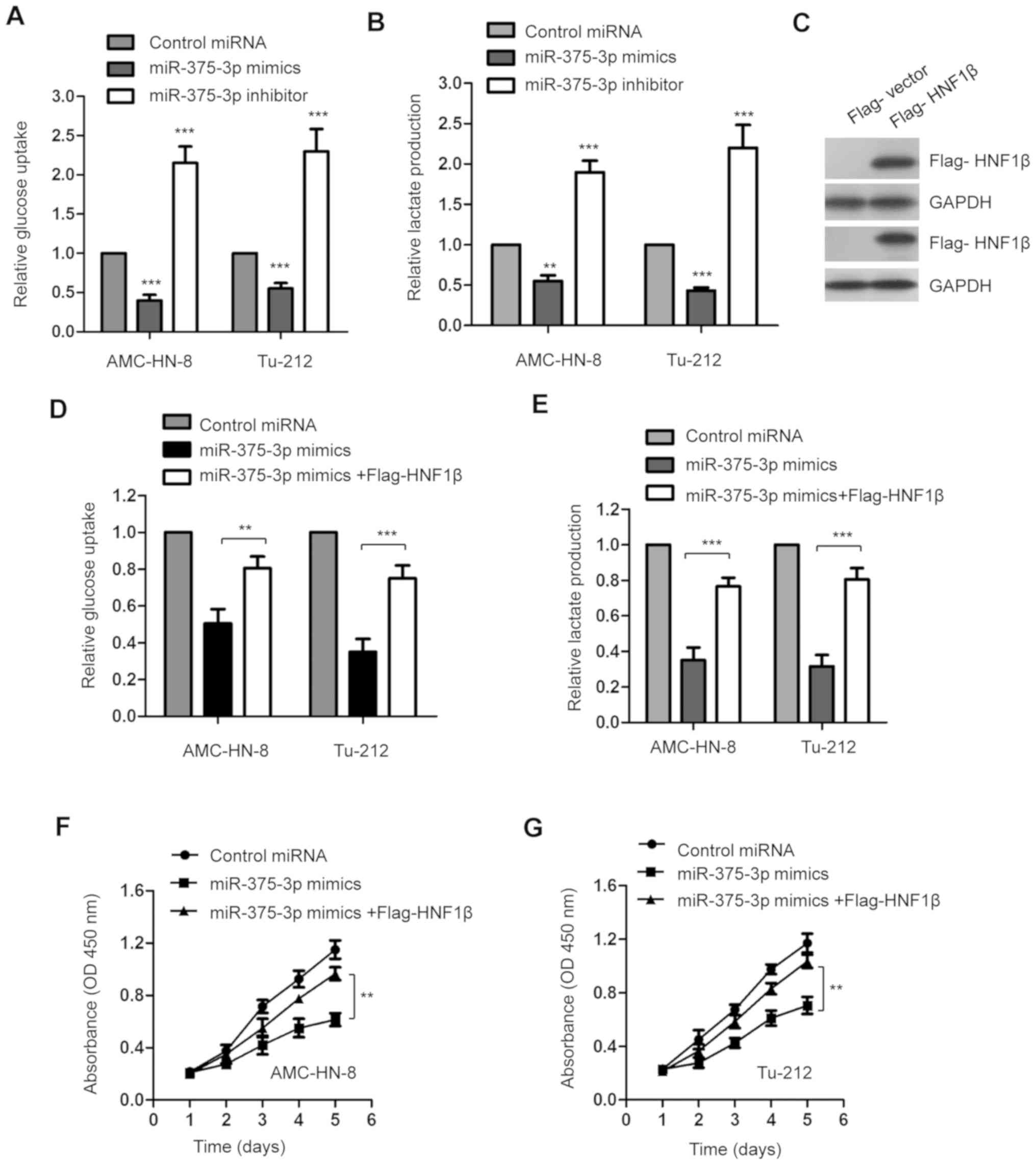
Given that miR-375-3p decreased HNF1β expression in
LSCC cells, the glucose uptake and lactate production of AMC-HN-8
and Tu-212 cells transfected with miR-375-3p mimics or inhibitor
were assessed to determine the influence of miR-375-3p on the
glucose metabolism of LSCC. The results indicated that
overexpression of miR-375-3p significantly suppressed glucose
consumption and lactate production in AMC-HN-8 and Tu-212 cells
(Fig. 4A and B; **P<0.01;
***P<0.001). Consistently, miR-375-3p knockdown significantly
promoted glucose metabolism of both AMC-HN-8 and Tu-212 cells
(Fig. 4A and B; **P<0.01;
***P<0.001).
In order to confirm the involvement of HNF1β in
suppressing glucose metabolism via miR-375-3p, HNF1β expression was
rescued by transfecting LSCC cells with Flag-HNF1β. A vector
encoding a FLAG tag was used to validate transfection efficiency of
HIFIβ (data not shown). Transfection with Flag-HNF1β was validated
via western blot analysis (Fig. 4C).
Restoration of HNF1β significantly attenuated the inhibitory role
of miR-375-3p in the glucose uptake and lactate generation of
AMC-HN-8 and Tu-212 cells (Fig. 4D and
E; **P<0.01; ***P<0.001).
Consistent with the important role of HNF1β in
miR-375-3p-mediated suppression of glucose metabolism,
overexpression of HNF1β significantly reversed the inhibitory
effect of miR-375-3p on the proliferation of AMC-HN-8 and Tu-212
cells (Fig. 4F and G; **P<0.01).
Taken together, these results suggest that miR-375-3p targets HNF1β
to regulate glucose metabolism of AMC-HN-8 and Tu-212 cells.
Discussion
The prognosis and treatment of LSCC have improved
with advancements in therapeutic strategies; the 5-year overall
survival rate of patients with LSCC is ~64% (5). Increasing evidence suggest that miRNAs
exert tumor suppressive or oncogenic functions in the progression
of LSCC (21,40–42). A
previous study reported that miR-375-3p is downregulated in head
and neck cancer (24). The results
of the present study demonstrated that miR-375-3p expression was
downregulated in LSCC tissues and cell lines, which was
significantly associated with larger tumor size, higher
pathological grades and distant metastasis of patients with LSCC.
Taken together, these results suggest the tumor suppressive role of
miR-375-3p in LSCC. However, further studies are required to
investigate the expression and function of miR-375-3p in different
types of cancer.
Due to the critical role miRNAs play in the
progression of malignancy, they are considered promising targets in
anticancer therapy (11,15,43).
Consistent with the results following miR-375-3p knockdown in LSCC,
overexpression of miR-375-3p inhibited the proliferation of LSCC
cells. Furthermore, overexpression of miR-375-3p increased the
apoptotic rate and induced cell cycle arrest at the G1
phase. Collectively, these results indicate that downregulation of
miR-375-3p may serve as a promising biomarker for the prognosis of
patients with LSCC, suggesting the role of miR-375-3p as a
potential therapeutic target for the intervention of LSCC. In order
to confirm the tumor suppressive role of miR-375-3p in LSCC,
prospective studies will focus on investigating the effect of
miR-375-3p on the growth of LSCC in vivo.
Given that miRNAs predominantly rely on modulating
the expression of target genes, bioinformatics analysis was
performed to identify the 3′-UTR of HNF1β containing the binding
site of miR-375-3p, in order to determine the molecular mechanism
underlying the suppressive function of miR-375-3p in LSCC. HNF1β is
a transcription factor that plays a critical role during several
processes, including cell proliferation, apoptosis and glucose
metabolism (30,34). In the present study, overexpression
of miR-375-3p significantly decreased the luciferase activity of
cells expressing 3′-UTR of HNF1β. Consistently, transfection with
miR-375-3p mimics inhibited mRNA and protein expression levels of
HNF1β in LSCC cells. Notably, overexpression of HNF1β significantly
reversed the inhibitory effect of miR-375-3p on the proliferation
of LSCC cells. Collectively, these results suggest that HNF1β is a
target of miR-375-3p, which mediates the suppressive role of
miR-375-3p in LSCC. Given the results of the present study, the
function of the miR-375-3p/HNF1β axis deserves further
investigation in different types of cancer. Notably, a single miRNA
has multiple targets (7,8), thus other targets of miR-375-3p may
play key roles in the progression of LSCC.
In conclusion, the results of the present study
demonstrated that miR-375-3p expression was downregulated in LSCC,
which was associated with a poor prognosis. Furthermore, miR-375-3p
targeted HNF1β and negatively regulated the proliferation of LSCC
cells. Studies with larger numbers of patients with LSCC may be
required to evaluate the clinical significance of miR-375-3p in the
diagnosis and prognosis of LSCC. Additionally, in vivo
studies are required to fully understand the tumor suppressive role
of miR-375-3p in the progression of LSCC. Taken together, these
results provide a novel insight into the pathogenesis of LSCC,
suggesting that the miR-375-3p/HNF1β axis may function as a
valuable therapeutic target for the treatment of LSCC.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author upon reasonable
request.
Author's contribution
KC designed the present study and performed the
experiments. ZW collected the tissue samples and performed the
RT-qPCR analysis. HC designed the study and drafted the initial
manuscript. All authors have revised and approved the
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethical
Institution of The First Affiliated Hospital of Zhengzhou
University (Zhengzhou, China; approval no. 20120134-445) and
performed in accordance with The Declaration of Helsinki. All
patients provided written informed consent prior to the study
start.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Lu ZM, Lin YF, Jiang L, Chen LS, Luo XN,
Song XH, Chen SH and Zhang SY: Micro-ribonucleic acid expression
profiling and bioinformatic target gene analyses in laryngeal
carcinoma. OncoTargets Ther. 7:525–533. 2014. View Article : Google Scholar
|
|
2
|
Tataru D, Mak V, Simo R, Davies EA and
Gallagher JE: Trends in the epidemiology of head and neck cancer in
London. Clin Otolaryngol. 42:104–114. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gao C and Hu S: miR-506 is a
YAP1-dependent tumor suppressor in laryngeal squamous cell
carcinoma. Cancer Biol Ther. 20:826–836. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Marur S and Forastiere AA: Head and Neck
Squamous Cell Carcinoma: Update on Epidemiology, Diagnosis, and
Treatment. Mayo Clin Proc. 91:386–396. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Palumbo A Jr, De Martino M, Esposito F,
Fraggetta F, Neto PN, Valverde Fernandes P, Santos IC, Dias FL,
Nasciutti LE, Meireles Da Costa N, et al: HMGA2, but not HMGA1, is
overexpressed in human larynx carcinomas. Histopathology.
72:1102–1114. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ozmen OA, Alpay M, Saraydaroglu O, Demir
UL, Kasapoglu F, Coskun HH and Basut OI: Prognostic significance of
soft tissue deposits in laryngeal carcinoma. Braz J
Otorhinolaryngol. 84:566–573. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fabian MR, Sonenberg N and Filipowicz W:
Regulation of mRNA translation and stability by microRNAs. Annu Rev
Biochem. 79:351–379. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mohr AM and Mott JL: Overview of microRNA
biology. Semin Liver Dis. 35:3–11. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gentilin E, Degli Uberti E and Zatelli MC:
Strategies to use microRNAs as therapeutic targets. Best Pract Res
Clin Endocrinol Metab. 30:629–639. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Momtazi AA, Shahabipour F, Khatibi S,
Johnston TP, Pirro M and Sahebkar A: Curcumin as a MicroRNA
Regulator in Cancer: A Review. Rev Physiol Biochem Pharmacol.
171:1–38. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Iorio MV and Croce CM: MicroRNA
dysregulation in cancer: Diagnostics, monitoring and therapeutics.
A comprehensive review. EMBO Mol Med. 9:8522017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Asadzadeh Z, Mansoori B, Mohammadi A,
Aghajani M, Haji-Asgarzadeh K, Safarzadeh E, Mokhtarzadeh A, Duijf
PHG and Baradaran B: microRNAs in cancer stem cells: Biology,
pathways, and therapeutic opportunities. J Cell Physiol.
234:10002–10017. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hosseinahli N, Aghapour M, Duijf PHG and
Baradaran B: Treating cancer with microRNA replacement therapy: A
literature review. J Cell Physiol. 233:5574–5588. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kwak PB, Iwasaki S and Tomari Y: The
microRNA pathway and cancer. Cancer Sci. 101:2309–2315. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Farazi TA, Spitzer JI, Morozov P and
Tuschl T: miRNAs in human cancer. J Pathol. 223:102–115. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li X, Wang HL, Peng X, Zhou HF and Wang X:
miR-1297 mediates PTEN expression and contributes to cell
progression in LSCC. Biochem Biophys Res Commun. 427:254–260. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yungang W, Xiaoyu L, Pang T, Wenming L and
Pan X: miR-370 targeted FoxM1 functions as a tumor suppressor in
laryngeal squamous cell carcinoma (LSCC). Biomed Pharmacother.
68:149–154. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Luo M, Sun G and Sun JW: miR-196b affects
the progression and prognosis of human LSCC through targeting
PCDH-17. Auris Nasus Larynx. 46:583–592. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Han L, Tang M, Xu X, Jiang B, Wei Y, Qian
H and Lu X: miR-143-3p suppresses cell proliferation, migration,
and invasion by targeting melanoma-associated antigen A9 in
laryngeal squamous cell carcinoma. J Cell Biochem. Oct 9–2018.(Epun
ahead of print).
|
|
22
|
Niu JT, Zhang LJ, Huang YW, Li C, Jiang N
and Niu YJ: miR-154 inhibits the growth of laryngeal squamous cell
carcinoma by targeting GALNT7. Biochem Cell Biol. 96:752–760. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen X, Zhang L and Tang S: MicroRNA-4497
functions as a tumor suppressor in laryngeal squamous cell
carcinoma via negatively modulation the GBX2. Auris Nasus Larynx.
46:106–113. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jamali Z, Asl Aminabadi N, Attaran R,
Pournagiazar F, Ghertasi Oskouei S and Ahmadpour F: MicroRNAs as
prognostic molecular signatures in human head and neck squamous
cell carcinoma: A systematic review and meta-analysis. Oral Oncol.
51:321–331. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zheng J: Energy metabolism of cancer:
Glycolysis versus oxidative phosphorylation (Review). Oncol Lett.
4:1151–1157. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Akram M: Mini-review on glycolysis and
cancer. J Cancer Educ. 28:454–457. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li XB, Gu JD and Zhou QH: Review of
aerobic glycolysis and its key enzymes - new targets for lung
cancer therapy. Thorac Cancer. 6:17–24. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Amano Y, Mandai M, Yamaguchi K, Matsumura
N, Kharma B, Baba T, Abiko K, Hamanishi J, Yoshioka Y and Konishi
I: Metabolic alterations caused by HNF1β expression in ovarian
clear cell carcinoma contribute to cell survival. Oncotarget.
6:26002–26017. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mandai M, Amano Y, Yamaguchi K, Matsumura
N, Baba T and Konishi I: Ovarian clear cell carcinoma meets
metabolism; HNF-1β confers survival benefits through the Warburg
effect and ROS reduction. Oncotarget. 6:30704–30714. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Okamoto T, Mandai M, Matsumura N,
Yamaguchi K, Kondoh H, Amano Y, Baba T, Hamanishi J, Abiko K,
Kosaka K, et al: Hepatocyte nuclear factor-1β (HNF-1β) promotes
glucose uptake and glycolytic activity in ovarian clear cell
carcinoma. Mol Carcinog. 54:35–49. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kato N and Motoyama T: Hepatocyte nuclear
factor-1beta (HNF-1beta) in human urogenital organs: Its expression
and role in embryogenesis and tumorigenesis. Histol Histopathol.
24:1479–1486. 2009.PubMed/NCBI
|
|
32
|
Kobayashi H, Yamada Y, Kanayama S,
Furukawa N, Noguchi T, Haruta S, Yoshida S, Sakata M, Sado T and Oi
H: The role of hepatocyte nuclear factor-1beta in the pathogenesis
of clear cell carcinoma of the ovary. Int J Gynecol Cancer.
19:471–479. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bockenhauer D and Jaureguiberry G:
HNF1B-associated clinical phenotypes: The kidney and beyond.
Pediatr Nephrol. 31:707–714. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bártů M, Dundr P, Němejcová K, Tichá I,
Hojný H and Hájková N: The Role of HNF1B in Tumorigenesis of Solid
Tumours: A Review of Current Knowledge. Folia Biol (Praha).
64:71–83. 2018.PubMed/NCBI
|
|
35
|
Zheng J, Liu X, Xue Y, Gong W, Ma J, Xi Z,
Que Z and Liu Y: TTBK2 circular RNA promotes glioma malignancy by
regulating miR-217/HNF1β/Derlin-1 pathway. J Hematol Oncol.
10:522017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Edge SB and Compton CC: The American Joint
Committee on Cancer: the 7th edition of the AJCC cancer staging
manual and the future of TNM. Ann Surgl Oncol. 17:1471–1474. 2010.
View Article : Google Scholar
|
|
38
|
Voigt JJ: Recommendations of the FNCLCC
Sarcoma Group for pathologic management of soft tissue sarcoma in
adults. Ann Pathol. 19:1531999.(In French). PubMed/NCBI
|
|
39
|
Shen Z, Yuan J, Tong Q, Hao W, Deng H, Li
Q, Zhou C, Hu Y and Xu J: Long non-coding RNA AC023794.4-201 exerts
a tumor-suppressive function in laryngeal squamous cell cancer and
may serve as a potential prognostic biomarker. Oncol Lett.
20:774–784. 2020.PubMed/NCBI
|
|
40
|
Fan Y, Xia X, Zhu Y, Diao W, Zhu X, Gao Z
and Chen X: Circular RNA expression profile in laryngeal squamous
cell carcinoma revealed by microarray. Cell Physiol Biochem.
50:342–352. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yu CH, Xing FY, Zhang JY, Xu JQ and Li YC:
A combination of mRNA expression profile and miRNA expression
profile identifies detection biomarkers in different tumor stages
of laryngeal squamous cell carcinoma. Eur Rev Med Pharmacol Sci.
22:7296–7304. 2018.PubMed/NCBI
|
|
42
|
Zhang F and Cao H: MicroRNA-143-3p
suppresses cell growth and invasion in laryngeal squamous cell
carcinoma via targeting the k-Ras/Raf/MEK/ERK signaling pathway.
Int J Oncol. 54:689–701. 2019.PubMed/NCBI
|
|
43
|
Kaboli PJ, Rahmat A, Ismail P and Ling KH:
MicroRNA-based therapy and breast cancer: A comprehensive review of
novel therapeutic strategies from diagnosis to treatment. Pharmacol
Res. 97:104–121. 2015. View Article : Google Scholar : PubMed/NCBI
|