Introduction
Giant cell tumor of bone (GCTB) is a common
intermediate bone tumor that is locally aggressive and rarely
metastasizes. Recurrence ranges from 16.7–39% after surgery alone,
although the recurrence rate is very high (>80%) following drug
withdrawal, and lung metastasis may occur in 1–3.2% of cases
(1,2). In pathology, GCTB tissue is composed
primarily of neoplastic stromal (NS) cells and multinucleated giant
(MNG) cells. NS cells secrete receptor activator of nuclear factor
κB ligand (RANKL), which binds to its receptor, RANK, leading to
MNG cell fusion and excessive activation of osteoclasts (3). NS cells have tumor proliferation
characteristics, which is the ultimate cause of GCTB recurrence
(4). However, the mechanism by which
NS cell proliferation is regulated has yet to be elucidated.
p62 (also known as sequestosome-1, SQSTM-1 or A170)
is a crucial molecule in the regulation of cell growth, survival
and proliferation (5). The p62
protein contains different types of protein-protein interaction
domains, a characteristic that is consistent with its role as a
versatile adaptor, especially for bone tumors and metabolic bone
diseases (6) p62 is overexpressed in
bone tumors, such as osteosarcoma (7) and myeloma (8). It is also associated with tumor
invasion and metastasis (9).
Moreover, p62 gene mutations are considered to be the main cause of
Paget's disease of bone, which is a skeletal disorder characterized
by excessive activation of osteoclasts, similar to GCTB (10). However, to date, the role of p62 in
the mechanisms underlying the biology of GCTB has not been
elucidated.
In the present study, mRNA and protein expression
levels of p62 were detected by reverse transcription-quantitative
PCR (RT-qPCR) and western blotting, respectively, in 8 paired fresh
GCTB tumor tissues and adjacent normal cancellous bone tissues. The
association between p62 expression levels and patient prognosis was
subsequently analyzed in 54 paraffin-embedded GCTB specimens via
immunohistochemistry assay. NS cells were isolated from GCTB
primary cell cultures, and the role of p62 was evaluated with
respect to the regulation of cell proliferation, invasion,
migration and apoptosis in vitro. The results revealed that
p62 mRNA and protein were overexpressed in tumor tissues, and that
high p62 expression levels were significantly associated with the
recurrence of GCTB. Furthermore, p62 downregulation led to the
inhibition of NS cell proliferation, invasion and migration, and an
increase in the rate of apoptosis. Therefore, it has been
demonstrated that p62 overexpression is associated with GCTB
recurrence via the promotion of NS cell proliferation, and p62 may
be both a novel prognostic indicator and a potential therapeutic
target for GCTB.
Materials and methods
Tissue specimens and patients
For RT-qPCR and western blotting assays, 8 paired
GCTB tumor tissues and the adjacent normal cancellous bone tissues
(at a distance >2 cm from the tumor) were collected during
surgery between May 2017 and July 2018. In addition, 54
paraffin-embedded specimens of GCTB tumor tissues were collected
between January 2008 and December 2014 for immunohistochemical
testing. All cases were histologically and clinically diagnosed at
The Third Affiliated Hospital of Kunming Medical University, Tumor
Hospital of Yunnan Province (China). The median follow-up time of
the patients was 77 months (ranging from 7–128 months). The
protocol for the present study was approved by the Medical
Institutional and Clinical Research Ethics Committee of Tumor
Hospital of Yunnan Province. All patients included in the present
study provided informed verbal consent.
RT-qPCR detection of p62 expression
and point mutation
Total RNA was extracted using TRIzol (Thermo Fisher
Scientific, Inc.) from fresh GCTB tissues. RT-qPCR was performed
using the All-in-One™ First-Strand cDNA Synthesis kit (GeneCopoeia,
Inc.). The temperature protocol was as follows: 42°C for 60 min,
70°C for 5 min. The quantitative primer sequences for p62 and GAPDH
primer sequences were as follows: p62 forward,
5′-TCAGGCTGTGGATGAAGTGGAA-3′ and reverse,
5′-CAGAAGATGTTTGTGGCGAGGA-3′; and GAPDH forward,
5′-TGACTTCAACAGCGACACCCA-3′ and reverse,
5′-CACCCTGTTGCTGTAGCCAAA-3′. The 7th and 8th exon sequencing were
as follows: Exon 7 primer forward, 5′-AGACCCCTGCAGCCTTAACT-3′ and
reverse, 5′-CCAACTCCTAACCTCCCACA-3′; and exon 8 primer forward,
5′-AGTTGAGCAGTGTGAAAAAGA-3′ and reverse,
5′-GCAGGGAGGGGTCAGGAGCGC-3′. RT-qPCR sequencing was performed using
the SYBR-Green Master with Rox kit (GeneCopoeia, Inc.). The
reaction conditions for qPCR were 95°C for 10 min, followed by 40
cycles of 95°C for 15 sec, 60°C for 30 sec and 72°C for 30 sec. The
mRNA expression levels in each group were quantified using the
2−ΔΔCq method (11).
Sequencing of the 7 and 8th exon amplification products was
performed.
Western blot assay
Fresh GCTB tissues were harvested using RIPA lysis
buffer (Beyotime Institute of Biotechnology). Protein
concentrations were measured using the bicinchoninic acid protein
assay kit (Beyotime Institute of Biotechnology). The total protein
of each specimen (30 mg/lane) were separated by SDS-PAGE (10%
gels), and then transferred onto a polyvinylidene difluoride
membrane (EMD Millipore). Β-actin was used as a loading control.
The membrane was blocked with 5% bovine serum albumin (BSA, Beijing
Solarbio Science & Technology Co., Ltd.) at room temperature
for 40 min, and subsequently incubated with mouse anti-p62
(1:1,500; cat. no. ab56416; Abcam), mouse anti-RANKL (1:1,000; cat.
no. ab45039; Abcam), mouse anti-RANK (1:1,000; cat. no. ab13918;
Abcam) and mouse anti-β-actin (1:5,000; cat. no. ab6276; Abcam)
primary antibodies overnight at 4°C. The membranes were then
incubated with peroxidase-conjugated goat anti-mouse IgG (1:20,000;
cat. no. A4416; Sigma-Aldrich; Merck KGaA) at room temperature for
1 h. Protein bands were visualized using SuperSignal™
West Femto Maximum Sensitivity Substrate reagents (Thermo Fisher
Scientific, Inc.). The relative gray value of the immune reactive
bands was compared using ImageJ software (version 1.46, National
Institutes of Health).
Immunohistochemistry assay
In brief, GCTB tissues were fixed in 10%
formaldehyde for 12 h at room temperature. Then paraffin-embedded
specimens of GCTB tumor tissues were cut into 4-µm sections and
baked at 65°C for 30 min. The sections were washed with xylene and
rehydrated with 70, 80, 90 and 100% graded ethanol solutions.
Tissue sections were submerged for 2 min into an EDTA buffer a 95°C
and 90 kPa for antigen retrieval. Subsequently, the sections were
treated with 3% hydrogen peroxide in methanol, followed by
incubation with 1% rabbit serum albumin (Cell Signaling Technology,
Inc.) at room temperature for 10 min. The specimens were incubated
overnight at 4°C with an anti-p62 antibody (1:800; cat. no. 16177S;
Cell Signaling Technology, Inc.). The specimens were then incubated
with SignalStain® Boost IHC Detection Reagent (1:1,000;
cat. no. 8114P; Cell Signaling Technology, Inc.) at 37°C for 30
min. The degree of immunostaining of sections was reviewed by light
microscope and scored by two independent pathologists. p62
cytoplasmic staining was scored from 0–3 as follows: i) 0, no or
faint staining; ii) 1, weak staining; iii) 2, moderate staining;
and iv) 3, strong staining. p62 nuclear staining was scored as
follows: i) 0, nuclear staining visible in <10% of nuclei; ii)
1, nuclear staining visible in >10% of nuclei. The total score
was calculated, and the samples were divided into low expression
(0–2) or high expression (3–4) groups (12).
GCTB NS cells isolation and
culture
GCTB fresh tissue was placed in 40 ml HANK's
balanced salt solution and stored at 0–4°C. The tissue was washed
three times with saline containing 3% penicillin-streptomycin
double antibody (cat. no. 516106, diluted in 10,000 IU
penicillin/ml and 10,000 µg streptomycin/ml; EMD Millipore). Twice
the tissue weight of pancreatin (1 g/ml; Thermo Fisher Scientific,
Inc.) was added to digest tissue at 37°C for 3 min, aspirate the
solution, neutralize with an equal volume of 15% fetal bovine serum
(FBS, Gibco; Thermo Fisher Scientific, Inc.), mix well, and
incubated at 4°C for 2 h. 0.25%. In total, 20 ml Trypsin (EMD
Millipore) was continuously added to the tissue fragments and the
aforementioned steps were repeated six times. The remaining tissue
was then resuspended with collagenase digestion solution (DMEM, 10%
FBS, 3% double antibody, 5 mg/ml collagenase II, 0.22 µm
filtration, all Biosharp Life Sciences). The tissues were then
shaken at 200 rpm on Rocking Device for 2 h at 37°C (CellNest
Shaker; http://www.shbiotech.com/). The digested
cells were filtered with a 70-µm filter and washed twice with PBS.
NS cells were cultured in DMEM/F12 supplemented with 15% FBS
(Gibco; Thermo Fisher Scientific) and 2% streptomycin/penicillin
(EMD Millipore).
Cell senescence assay
First, the NS cells were seeded in 6-well plates
(EMD Millipore). Then culture fluid was aspirated, washed once with
PBS and 1 ml β-galactosidase staining fixative was added (Beijing
Solarbio Science & Technology Co., Ltd.), and cells were fixed
at room temperature for 15 min. Then the cell fixative was
aspirated and the cells were washed three times with PBS, each for
3 min. Then PBS was aspirated and 1 ml staining working solution
(Beijing Solarbio Science & Technology Co., Ltd.) was added to
each well. Then 300 µl dyeing working fluid was added in each well
and incubated at 37°C overnight. The 6-well plate was sealed with
parafilm or plastic wrap to prevent evaporation. Lastly, cells were
observed under a light microscope at 200× magnification.
Lentivirus infection of NS cells
p62 (NM_003900) overexpression plasmid
(pcDNA3.1-p62) was purchased from GeneCopoeia Co., Ltd. (cat. no.
EX-M0245-Lv201). Lv201 empty lentiviral vector was purchased from
GeneCopoeia (cat. no. EX-NEG-Lv201) for negative control. For p62
knockdown, short hairpin RNA (p62-shRNA, cat. no.
HSH021660-LVRU6GP) and scrambled shRNA (cat. no. SHCTR001-LVRU6GP)
were both purchased from GeneCopoeia. The shRNA sequences were as
follows: p62-shRNA, 5′-CCATCCAGTATTCAAAGCATC-3′ and scrambled
shRNA, 5′-GCTTCGCGCCGTAGTCTTA-3′. Transfections of the NS cells
with plasmids (1 µg/well) and shRNA (50 nm) were performed using a
Lipofectamine® 3000 kit (Gibco; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocols. Cell
density was 106 cells/25 cm dish. Puromycin (1 µg/ml)
was used to kill any cells that were not successfully transfected.
NS cells either stably expressing, or silenced for p62 were
constructed. Subsequent experimentation was conducted after
transfection for 48 h. RT-qPCR was used to verify the expression of
p62 mRNA, and western blotting was performed to evaluate the
expression of p62 protein. NS cells were split into five groups,
including: GCTB control, normal NS cells; shp62, silenced p62 gene
lentivirus-transfected NS cells; NCshp62, shp62 scrambled control;
mp62, exogenous p62 gene expression lentivirus-transfected NS cells
and NCmp62, mp62 negative control (Lv201 empty vector).
Cell proliferation assay
NS cells stably expressing p62 were plated in
96-well plates in DMEM with 15% FBS (both Gibco; Thermo Fisher
Scientific, Inc.) at a density of 5,000 cells/well. To quantify
cell viability, cultures were stained after 1–4 and 5 days; 10 µl
Cell Counting Kit-8 (CCK-8, Dojindo Molecular Technologies, Inc.)
working solution was then added into the wells for 2 h at 37°C
according to the manufacturer's instructions, after which the
absorbance was measured at 450 nm using an Epoch Multi-Volume
Spectrophotometer system (BioTek Instruments, Inc.).
Cell apoptosis assay
Cell early and late apoptosis assays were performed
using an Annexin V-Fluorescein Isothiocyanate Apoptosis Detection
kit (BD Biosciences) and NS cells stably expressing p62. All cells
were collected, resuspended in 1X binding buffer, and stained with
Annexin V-FITC and propidium iodide at room temperature for 5 min,
followed by flow cytometry detection (Attune NxT; Thermo Fisher
Scientific, Inc.). The rate of early and late apoptosis was
analyzed using CellQuest™ software (version 3.3; BD
Biosciences).
Cell invasion and migration
assays
Cells (1×105), suspended in DMEM
containing 0.1% BSA were added to the top of the Boyden chamber
(EMD Millipore) at 37°C for 2 h, which was coated with 50 µl
Matrigel (1 mg/ml final concentration). The lower chamber contained
10% serum-containing medium. After incubation for 24 h at 37°C, a
Transwell chamber (EMD Millipore) was used to determine cell
invasion and migration ability. A total of 1×105
lentivirus-infected NS cells were suspended in DMEM containing 10%
FBS at a density of 5,000 cells/well. Cells were subsequently
placed onto the top of each chamber. Medium containing 10% FBS was
added to the bottom of the chamber. The cells were incubated for 24
h, and then cells on the upside of the membrane were wiped off to
remove the non-migrated cells. Cells that had migrated to the
underside of the membrane were stained with crystal violet at room
temperature for 20 min and visualized under Nikon Eclipse TE2000-U
microscope. A total of 4 random fields (magnification, ×100) were
scanned and analyzed using the aforementioned ImageJ software.
Statistics analysis
Statistical analyses of immunohistochemistry assay
were performed using the SPSS 17.0 software package (SPSS, Inc.).
The significance of the differences between groups was estimated
using the χ2 test. Recurrence-free survival curves were
plotted according to the Kaplan-Meier method, and compared using
the log-rank test. The significance of survival variables was
evaluated using a multivariate Cox proportional hazards regression
analysis. Statistical graphs were drawn using GraphPad Prism v.6.0
(GraphPad Software, Inc.). The differences of p62 mRNA and protein
expression levels between two groups of GCTB tumor tissues and the
adjacent normal cancellous bone tissues were tested with a
Student's t-test. The differences of p62 protein expression levels,
NS cell proliferation rates and invasion cell numbers between
multiple groups were tested by one-way ANOVA with post-hoc Tukey's
test. All data are presented as the mean ± standard deviation of
three independent experiments. P<0.05 was considered to indicate
a statistically significant difference.
Results
p62 mRNA and protein are overexpressed
in fresh GCTB tumor tissues
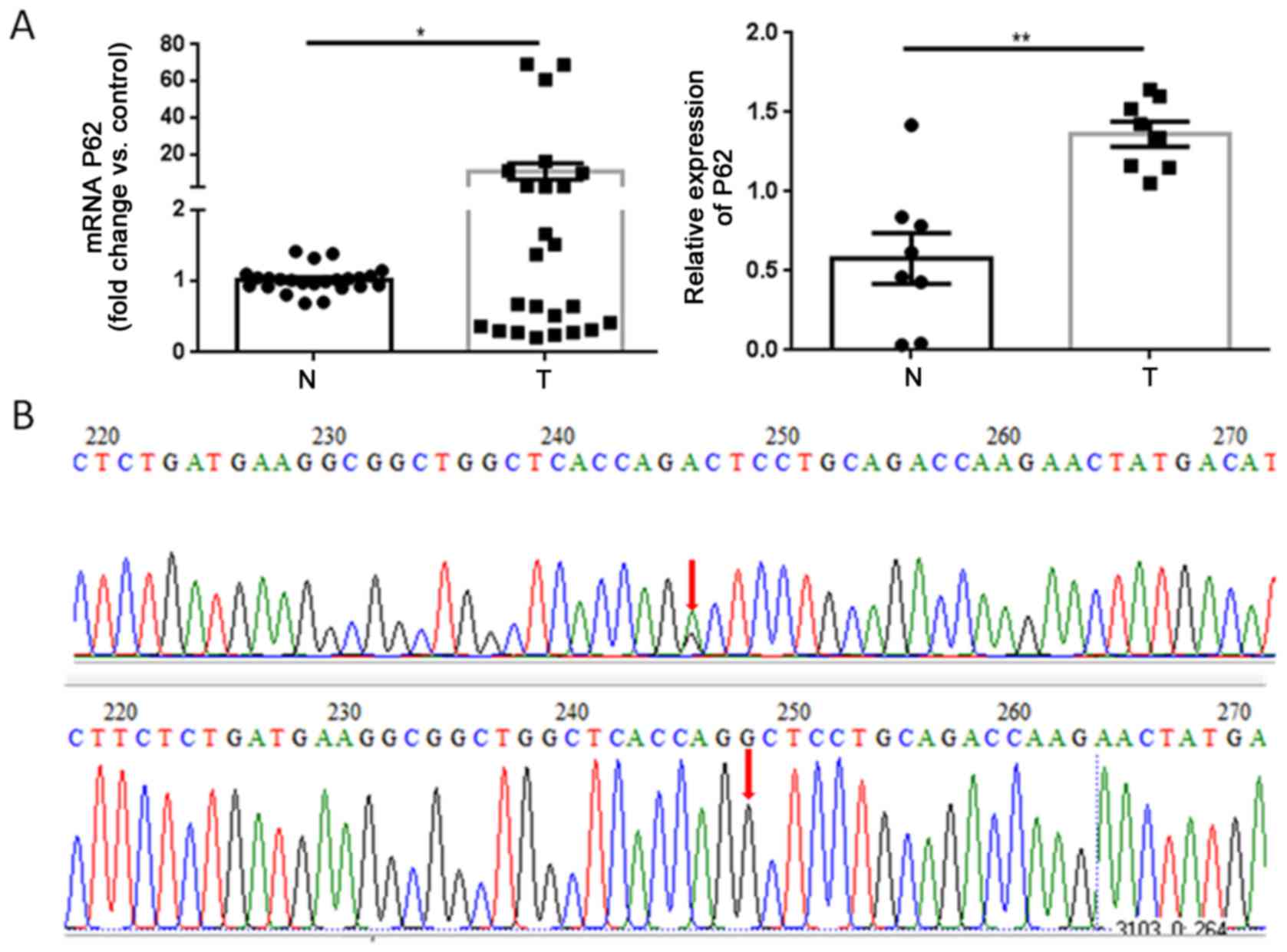
In 8 paired fresh GCTB tumor tissues and adjacent
normal cancellous bone tissues, p62 mRNA and protein were
overexpressed in tumors compared with normal tissues, as determined
by RT qPCR and western blot assays, respectively (Fig. 1A). Gene sequencing revealed that the
G1340A mutation occurred at the 1,340 bp position in two tumor
specimens between exons 7 and 8 of the p62 gene (Fig. 1B). However, the amino acid R
(arginine) at position 415 was not changed.
p62 overexpression is associated with
poor prognosis of patients with GCTB
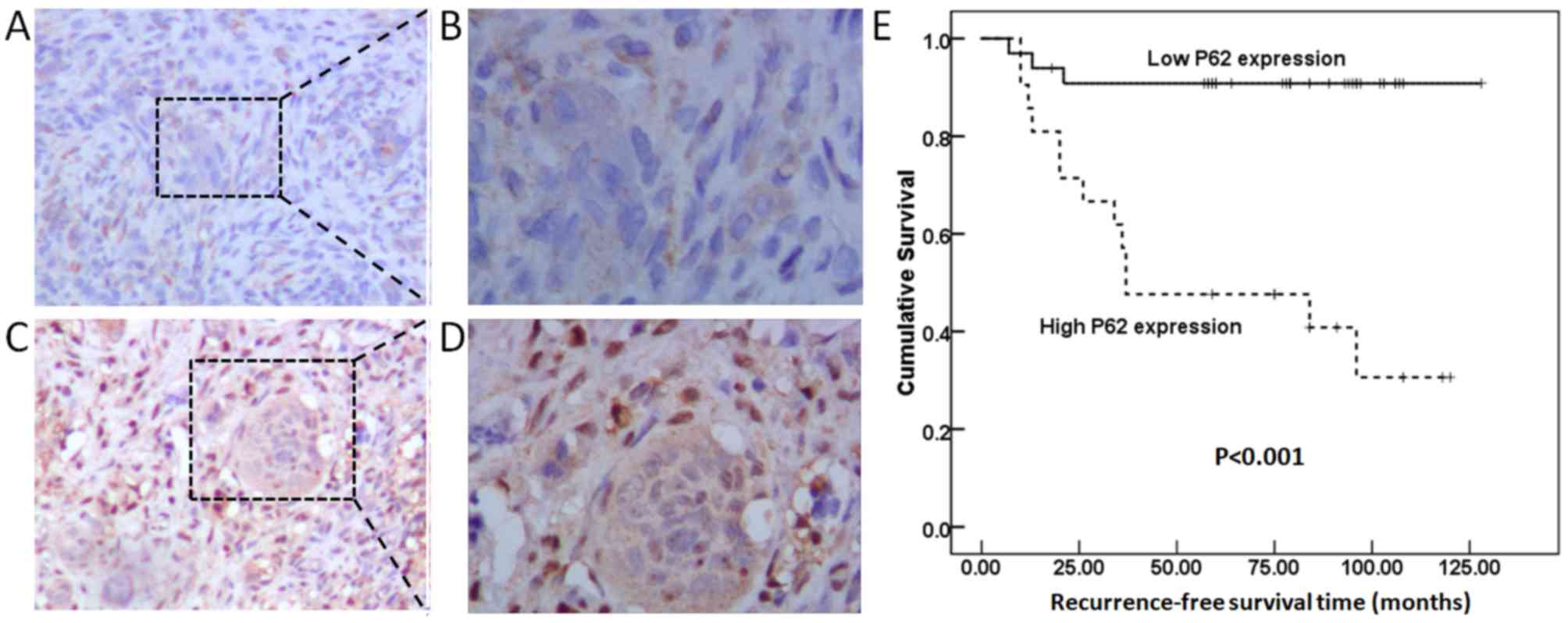
As shown in Fig.
2A-D, and as demonstrated by strong cytoplasmic and nuclear
staining, the p62 protein was widely expressed in NS and MNG cells.
To assess the association between p62 expression levels and
clinicopathological features, the GCTB specimens were classified
into a high p62 expression level group (n=20) and a low p62
expression level group (n=34). High p62 expression levels were
significantly associated with GCTB recurrence (P=0.001; Table I). The patients in the high p62
expression level group had shorter 5-year recurrence-free survival
rates compared with patients in the low p62 expression level group
(P<0.001; Fig. 2E). Cox
regression analysis identified p62 expression as an independent
prognostic indicator for recurrence-free survival of patients with
GCTB (P<0.001; Table II).
 | Table I.Association of p62 expression with
clinicopathological features of patients with giant cell tumor of
bone. |
Table I.
Association of p62 expression with
clinicopathological features of patients with giant cell tumor of
bone.
|
|
| p62 expression |
|
|---|
|
|
|
|
|
|---|
| Clinicopathological
features | Number of cases | Low, n (%) | High, n (%) | P-value |
|---|
| Age, years |
| ≤40 | 43 | 29 (67.4) | 14 (32.6) | 0.178 |
|
>40 | 11 | 5 (45.5) | 6 (54.5) |
|
| Sex |
| Male | 28 | 20 (71.4) | 8 (28.6) | 0.181 |
|
Female | 26 | 14 (53.8) | 12 (46.2) |
|
| Recurrence |
| Yes | 16 | 3 (18.8) | 13 (81.2) | 0.001 |
| No | 38 | 31 (81.6) | 7 (18.4) |
|
| Pathological
fracture |
| Yes | 40 | 26 (65) | 14 (35) | 0.600 |
| No | 14 | 8 (57.1) | 6 (42.9) |
|
| Campanacci grade |
| I | 2 | 1 (50) | 1 (50) | 0.906 |
| II | 9 | 6 (66.7) | 3 (33.3) |
|
| III | 43 | 27 (62.8) | 16 (37.25) |
|
| Table II.Cox regression analysis of
recurrence-free survival in 54 patients with giant cell tumor of
bone. |
Table II.
Cox regression analysis of
recurrence-free survival in 54 patients with giant cell tumor of
bone.
|
|
|
|
|
|
|
| 95% CI for Exp
(B) |
|---|
|
|
|
|
|
|
|
|
|
|---|
| Clinicopathological
features | B | SE | Wald | df | Sig | Exp(B) | Lower | Upper |
|---|
| Age | −0.073 | 0.625 | 0.014 | 1 | 0.907 | 0.930 | 0.273 | 3.165 |
| Sex | −0.048 | 0.534 | 0.008 | 1 | 0.929 | 0.954 | 0.335 | 2.716 |
| Pathological
fracture | 0.136 | 0.817 | 0.028 | 1 | 0.868 | 1.146 | 0.231 | 5.678 |
| Campanacci
grade | 0.342 | 0.769 | 0.198 | 1 | 0.657 | 1.407 | 0.312 | 6.349 |
| p62 expression | −2.227 | 0.650 | 11.753 | 1 | 0.001 | 0.108 | 0.030 | 0.385 |
p62 downregulation inhibits the
proliferation, invasion and migration of NS cells
GCTB primary cells were isolated, and it was
observed that the primary cells were mainly composed of
spindle-shaped NS cells and osteoclast-like MNG cells. After 2–3
passage generations, it was found that the population of MNG cells
gradually decreased. In the 4th passage generation, only NS cells
remained (Fig. 3A-B). To further
ensure that only NS cells remained in the 4th passage generation,
western blotting was used to measure the expression of two markers
(RANKL and RANK; Fig. S1). A cell
senescence assay indicated more and more aging NS cells emerged
from 4th generation (Fig. 3C and D).
Therefore, 4th generation cells were used for subsequent
experiments.
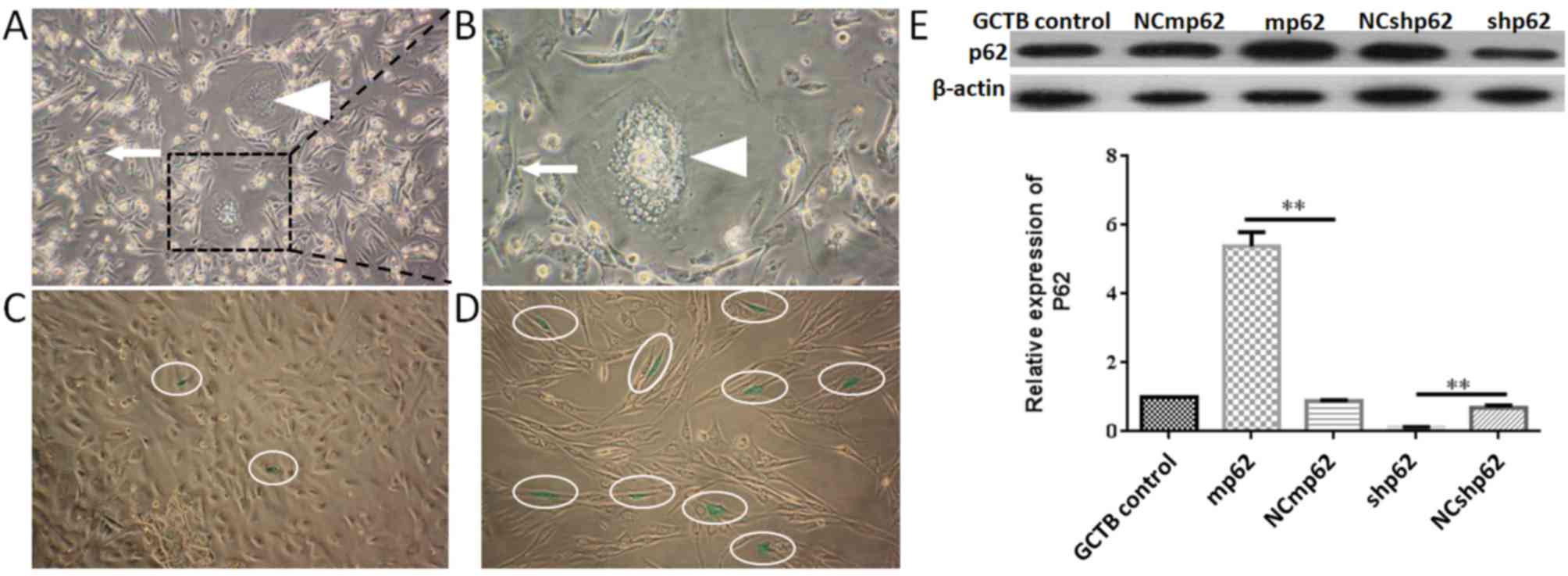 | Figure 3.GCTB primary cell culture and passage.
(A) GCTB primary cells 48 h after passage (magnification, ×100).
Cells are mainly composed of spindle shaped NS cells (white arrow)
and osteoclast like multinucleated giant cells (white triangle).
(B) GCTB primary cells 48 h after passage (magnification, ×400). NS
cells (white arrow) and multinucleated giant cells (white triangle)
were marked. (C) β galactosidase staining assay (green staining was
indicted by white oval) of NS cells at the 4th passage
(magnification, ×200). Cells grew well, and the senescence rate was
2.02±0.21%. (D) NS cells at the 7th passage became tenuous
(magnification, ×200), and the senescence rate was 8.40±1.14%. (E)
Lentivirus infection of NS cells, **P<0.01. GCTB control, normal
NS cells; shp62, silenced p62 gene lentivirus transfected NS cells;
NCshp62, shp62 scrambled control; mp62, exogenous p62 gene
expression lentivirus transfected NS cells; NCmp62, mp62 negative
control; GCTB, giant cell tumor of bone; NS, neoplastic
stromal. |
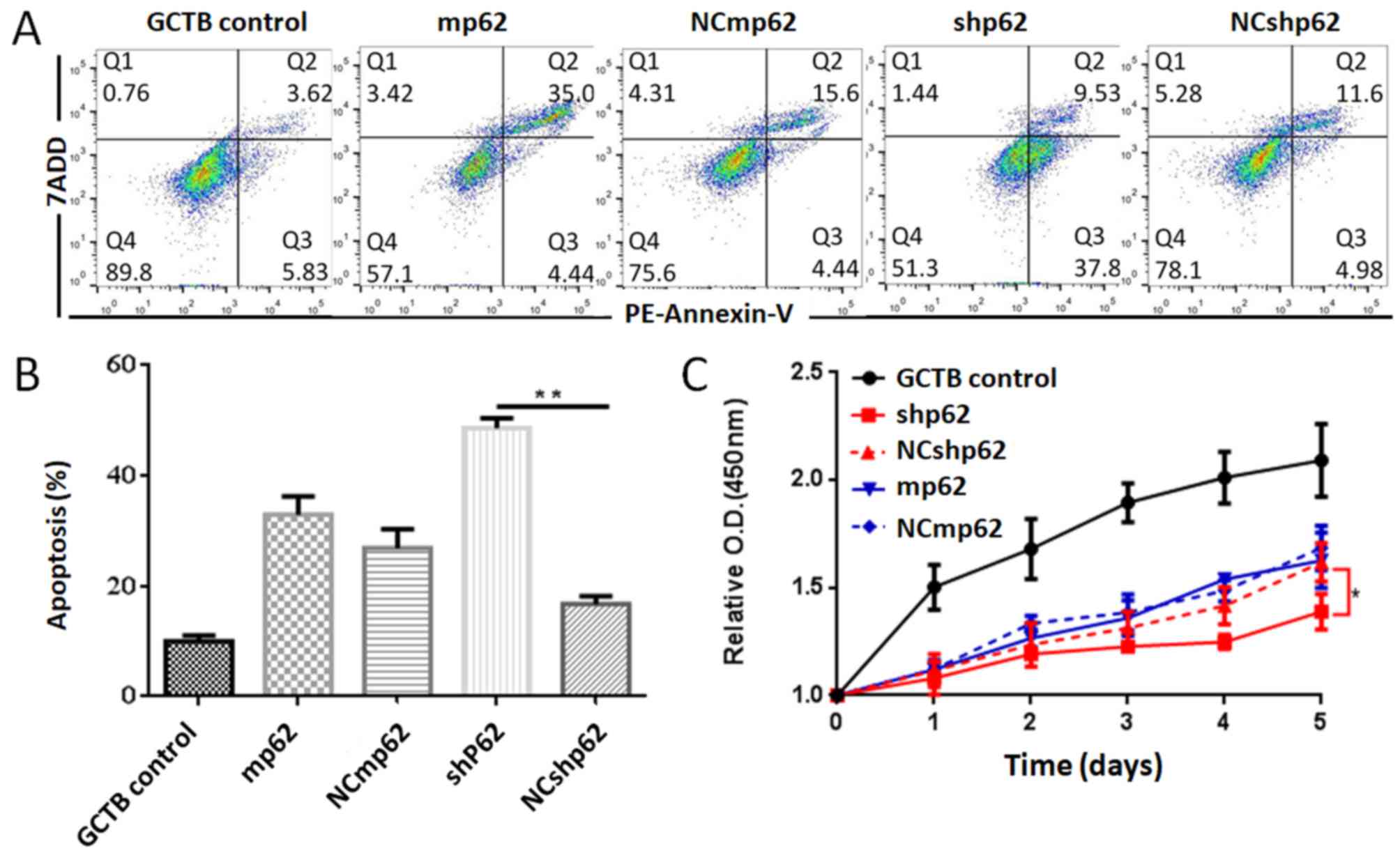
Subsequently, the expression of p62 was either
downregulated or upregulated using lentiviral plasmid transfection
(Fig. 3E). The differences between
the silenced p62 gene lentivirus-transfected NS cells (the shp62
downregulation group) and the scrambled control (NCshp62) group
were analyzed. The percentage of early and late apoptotic NS cells
in the shp62 group was 47.33%, whereas the percentage of early and
late apoptotic cells in the NCshp62 group was 16.58% (Fig. 4A and B). The difference in
proliferation measured by relative OD values between the two groups
was found to be statistically significant on day 4 (P=0.0343) and
on day 5 (P=0.0305) (Fig. 4C).
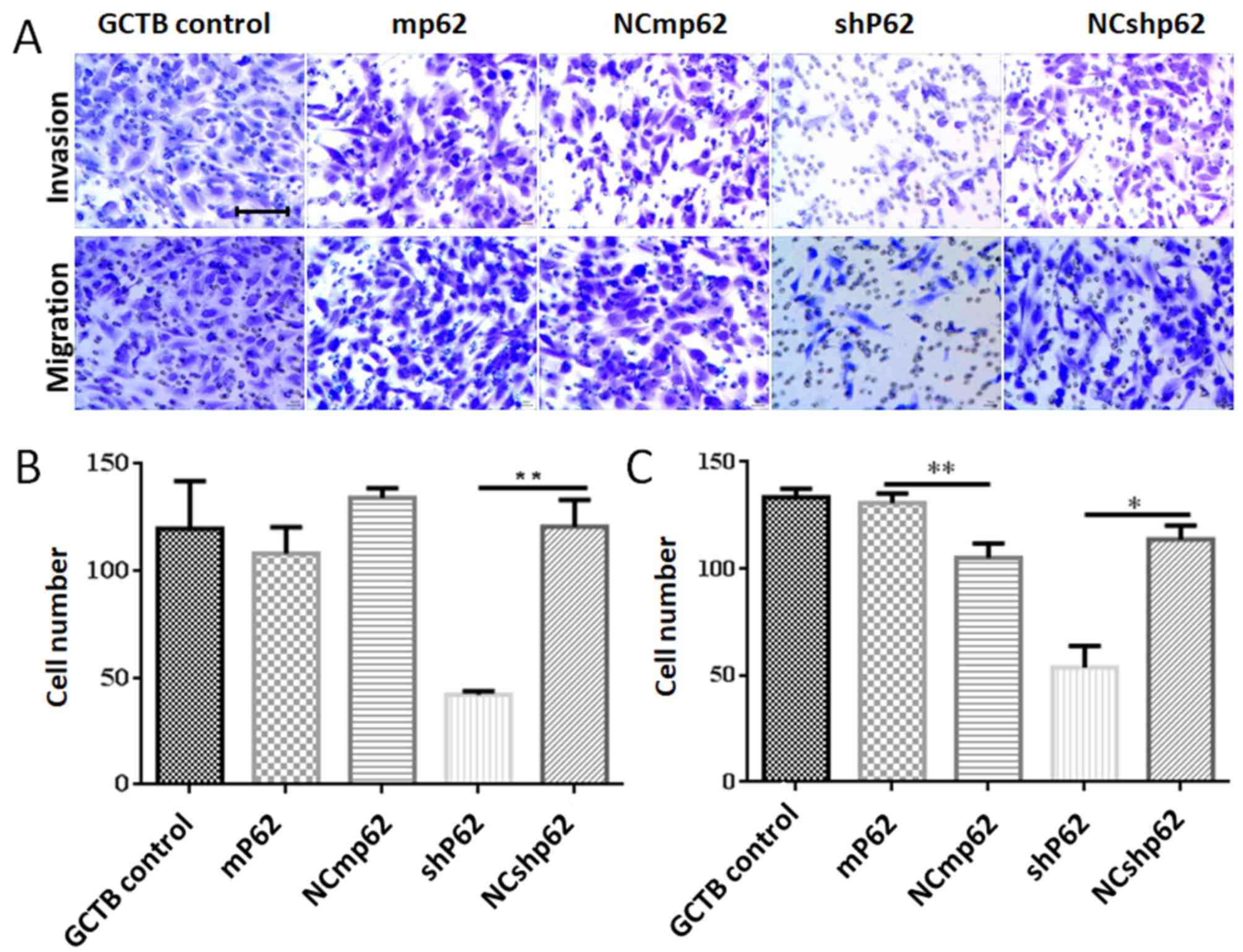
Downregulation of p62 also led to the inhibition of NS cell
invasion (P=0.042; Fig. 5B) and
migration (P=0.002; Fig. 5C).
Discussion
The overexpression of p62 has been detected among a
melanoma and hepatocellular carcinoma (13–15),
playing positive roles in both tumorigenesis and metastasis
(16). Recently, p62 was
demonstrated to be an effective oncotarget for solid tumors. In a
multicenter phase I/IIa clinical trial, a p62 vaccine (Elenagen)
alone or in combination with chemotherapy resulted in stable
disease for 8–32 weeks in 44% of patients with advanced solid
tumors (17). It also brought new
hope for identifying potential therapeutic targets of GCTB. In the
present study, overexpression of p62 mRNA and protein was observed,
indicating that p62 may contribute to GCTB initiation. It was also
possible to exclude the possibility of a p62 gene mutation between
exon 7 and exon 8 (P392L) in GCTB, which is the most common
mutation area of p62 in Paget's disease of bone (18). These results revealed that, in the
GCTB tumor microenvironment, p62 overexpression may be associated
with tumorigenesis that is unrelated to bone metabolism. High p62
expression levels were associated with GCTB recurrence, a finding
which agreed with previous findings in other bone tumors (6,7).
Finally, p62 overexpression was identified as an independent
indicator of poor prognosis of GCTB. To the best of our knowledge,
this is the first study that has analyzed the role of p62 in GCTB
prognosis.
GCTB is not a malignant tumor, and NS cells do not
have the ability to proliferate infinitely. This also explains the
lack of standard GCTB cell lines that have been available to us for
research purposes to date (19). To
clarify the cellular and molecular mechanism of GCTB recurrence, NS
cells were isolated from GCTB primary cell cultures. Fourth
generation cells were used for experiments in which the cells
maintained a rapid proliferation rate and stable growth
characteristics during this period. Functional assays revealed that
downregulation of p62 could inhibit the proliferation, invasion and
migration of NS cells in GCTB. These results were consistent with
most previous findings in other bone tumors (6). The present results also demonstrated
that downregulation of p62 could promote apoptosis of NS cells in
GCTB, a finding which provides a novel clue for the proliferative
regulatory mechanism of p62.
For advanced GCTB, recurrence is often observed
after denosumab withdrawal, in addition to surgery (20). The first fully humanized monoclonal
antibody against RANKL, denosumab has achieved a good response in
GCTB (21,22). However, the reason for the rapid
recurrence after denosumab withdrawal has yet to be identified.
Previous studies demonstrated that the expression of RANKL/RANK was
downregulated following denosumab treatment, although the
proliferation of NS cells was not inhibited; indeed, the rate even
increased (23,24). In the present study, it has been
demonstrated that p62 is able to promote the proliferation of NS
cells. This finding provides a further explanation towards the
understanding of GCTB recurrence after denosumab withdrawal.
The present study does, however, have some
limitations. First, it was a single-center study, and therefore the
number of clinical specimens to analyze was small. Secondly,
although the positive role of p62 in the proliferation of NS cells
was identified, the underlying signaling pathway associated with
p62 adaptor function is not clear. Consequently, our future studies
will focus on the interaction between p62 and other key regulators
of GCTB, such as RANKL and RANK.
In conclusion, to the best of our knowledge, the
present study is the first to have shown that p62 is overexpressed
in GCTB tumor tissues. High p62 expression levels were associated
with GCTB recurrence. Additionally, p62 overexpression was shown to
promote NS cell proliferation by regulating apoptosis. We propose
that p62 could be a novel prognostic indicator and a potential
therapeutic target for GCTB.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This work was supported by the National Nature
Science Foundation of China (grant no. 81760486), and Science and
Technology Plan of Yunnan province (grant no. 2018FB133).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JZ designed this research and revised the
manuscript. SL and FY performed the western blot analysis, RT-qPCR
assays and cell experiments, and wrote the manuscript. DL and CH
performed the immunohistochemistry assays. HH analyzed the data.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The protocol for the present study was approved by
the Medical Institutional and Clinical Research Ethics Committee of
Yunnan Tumor Hospital (approval no. KY202030; Kunming, China). All
patients included in the present study previously provided informed
verbal consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Mavrogenis AF, Igoumenou VG,
Megaloikonomos PD, Panagopoulos GN, Papagelopoulos PJ and Soucacos
PN: Giant cell tumor of bone revisited. SICOT J. 3:542017.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Niu X, Zhang Q, Hao L, Ding Y, Li Y, Xu H
and Liu W: Giant cell tumor of the extremity: Retrospective
analysis of 621 Chinese patients from one institution. J Bone Joint
Surg Am. 94:461–467. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lüke J, Hasenfratz M, Möller P and Barth
TFE: New aspects on giant cell tumor of bone. Pathologe.
39:125–131. 2018.(In German). View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhou Z, Li Y, Xu L, Wang X, Chen S, Yang C
and Xiao J: Biological characteristics of a novel giant cell tumor
cell line derived from spine. Tumour Biol. 37:9681–9689. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fan L, Yin S, Zhang E and Hu H: Role of
p62 in the regulation of cell death induction. Apoptosis.
23:187–193. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sánchez-Martín P, Saito T and Komatsu M:
p62/SQSTM1: ‘Jack of all trades’ in health and cancer. FEBS J.
286:8–23. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lu Y, Wang Q, Zhou Y, Sun L, Hu B, Xue H,
Li M, Zhang K, Ren C, Duan N, et al: Overexpression of p62 is
associated with poor prognosis and aggressive phenotypes in
osteosarcoma. Oncol Lett. 15:9889–9895. 2018.PubMed/NCBI
|
|
8
|
Sha Z, Schnell HM, Ruoff K and Goldberg A:
Rapid induction of p62 and GABARAPL1 upon proteasome inhibition
promotes survival before autophagy activation. J Cell Biol.
217:1757–1776. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang J, Yang Z and Dong J: P62: An
emerging oncotarget for osteolytic metastasis. J Bone Oncol.
5:30–37. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shaw B, Burrel CL, Green D,
Navarro-Martinez A, Scott D, Daroszewska A, van't Hof R, Smith L,
Hargrave F, Mistry S, et al: Molecular insights into an ancient
form of Paget's disease of bone. Proc Natl Acad Sci USA.
116:10463–10472. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Langer R, Neppl C, Keller MD, Schmid R,
Tschan MP and Berezowska S: Expression analysis of autophagy
related indicators LC3B, p62 and HMGB1 indicate an
autophagy-independent negative prognostic impact of high p62
expression in pulmonary squamous cell carcinomas. Cancers (Basel).
10:2812018. View Article : Google Scholar
|
|
13
|
Karras P, Riveiro-Falkenbach E, Cañón E,
Tejedo C, Calvo TG, Martínez-Herranz R, Alonso-Curbelo D, Cifdaloz
M, Perez-Guijarro E, Gómez-López G, et al: p62/SQSTM1 fuels
melanoma progression by opposing mRNA decay of a selective set of
pro-metastatic factors. Cancer Cell. 35:46–63 e10. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xing M, Li P, Wang X, Li J, Shi J, Qin J,
Zhang X, Ma Y, Francia G and Zhang JY: Overexpression of p62/IMP2
can promote cell migration in hepatocellular carcinoma via
activation of the Wnt/β-catenin pathway. Cancers (Basel). 12:72019.
View Article : Google Scholar
|
|
15
|
Umemura A, He F, Taniguchi K, Nakagawa H,
Yamachika S, Font-Burgada J, Zhong Z, Subramaniam S, Raghunandan S,
Duran A, et al: p62, upregulated during preneoplasia, induces
hepatocellular carcinogenesis by maintaining survival of stressed
HCC-Initiating cells. Cancer Cell. 29:935–948. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Moscat J, Karin M and Diaz-Meco MT: p62 in
Cancer: Signaling adaptor beyond autophagy. Cell. 167:606–609.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ponomarenko DM, Klimova ID, Chapygina YA,
Dvornichenko VV, Zhukova NV, Orlova RV, Manikhas GM, Zyryanov AV,
Burhanova LA, Badrtdinova II, et al: Safety and efficacy of P62 DNA
vaccine ELENAGEN in a first-in-human trial in patients with
advanced solid tumors. Oncotarget. 8:53730–53739. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Singer FR: Paget's disease of bone-genetic
and environmental factors. Nat Rev Endocrinol. 11:662–671. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cowan RW and Singh G: Giant cell tumor of
bone: A basic science perspective. Bone. 52:238–246. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lipplaa A, Dijkstra S and Gelderblom H:
Challenges of denosumab in giant cell tumor of bone, and other
giant cell-rich tumors of bone. Curr Opin Oncol. 31:329–335. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Branstetter DG, Nelson SD, Manive JC, Blay
JY, Chawla S, Thomas DM, Jun S and Jacobs I: Denosumab induces
tumor reduction and bone formation in patients with giant-cell
tumor of bone. Clin Cancer Res. 18:4415–4424. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chawla S, Henshaw R, Seeger L, Choy E,
Blay JY, Ferrari S, Kroep J, Grimer R, Reichardt P, Rutkowski P, et
al: Safety and efficacy of denosumab for adults and skeletally
mature adolescents with giant cell tumor of bone: Interim analysis
of an open-label, parallel-group, phase 2 study. Lancet Oncol.
14:901–908. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mukaihara K, Suehara K, Kohsaka S, Akaike
K, Tanabe T, Kubota D, Ishii M, Fujimura T, Kazuno S, Okubo T, et
al: Protein expression profiling of giant cell tumors of bone
treated with denosumab. PLoS One. 11:e01484012016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lau CP, Huang L, Wong KC and Kumta SM:
Comparison of the anti-tumor effects of denosumab and zoledronic
acid on the neoplastic stromal cells of giant cell tumor of bone.
Connect Tissue Res. 54:439–449. 2013. View Article : Google Scholar : PubMed/NCBI
|