Introduction
Lung cancer is the most common human malignancy,
accounting for 21.7% of all cancer-associated deaths worldwide
during 2015 (1). In addition, its
morbidity and mortality rank the highest among all malignant tumor
types worldwide (2). According to
the differentiation degree and morphological characteristics of
cancer cells, lung cancer can be roughly classified into
non-small-cell lung cancer (NSCLC) and small-cell lung cancer
(3). Among patients with lung
cancer, nearly 80% are diagnosed as NSCLC, which manifests with
earlier diffusion and metastasis (4). Currently, resection, chemotherapy,
radiotherapy and targeted therapy are the primary treatments for
lung cancer (5). For patients with
advanced NSCLC or those who are clinically incapacitated for
surgery, chemotherapy is a remarkably important treatment (6). Cisplatin (DDP) is widely applied in the
treatment of several malignancies, and it exhibits a broad spectrum
of antitumor effects by inducing DNA damage and hindering DNA
damage repair (7). Paclitaxel (PTX),
another commonly used chemotherapeutic agent in the clinic, targets
the microtubule cytoskeleton and impedes cell division (8,9). The
majority of patients have a good initial response to chemotherapy
agents; however, subsequent relapse is common and largely due to
the emergence of drug resistance (10). Thus, chemoresistance is considered
one of the main factors of poor prognosis in patients with advanced
NSCLC (6). Therefore, there is an
urgent need to investigate the target and mechanism of
chemoresistance in NSCLC.
Close homolog of L1 (CHL1) is a member of the L1
family of nerve cell adhesion molecules and is located on the 3q26
locus (11). As a nerve cell
adhesion molecule, CHL1 serves an important role in the
development, regeneration and plasticity of the nervous system
(12). The absence or mutation of
CHL1 can trigger 3p syndrome and schizophrenia (13). The abnormal expression of CHL1 may
lead to reduced working memory and social behavior, mental damage,
and abnormal behavior (14). CHL1
has been reported to be involved in carcinogenesis and progression
in a variety of human cancers. In esophageal squamous cell
carcinoma (ESCC), CHL1 downregulation is associated with invasion,
lymph node metastasis and poor overall survival (11). Functional studies revealed that CHL1
has anti-proliferation and anti-metastasis abilities (11). The expression of CHL1 is
downregulated by hypermethylation in human breast cancer, and its
negative expression contributes to breast tumorigenesis and
progression (15,16). In thyroid cancer (17) and colonic adenocarcinoma (18), CHL1 impedes cell proliferation and
invasion, and acts as a tumor suppressor. In lung cancer, Hӧtzel
et al (19) evaluated CHL1
expression in 2,161 NSCLC cases based on a tissue microarray, and
it was reported that CHL1 expression is associated with T stage in
adenocarcinomas, as well as with metastatic lymph node status and
improved survival. Additionally, by analyzing the Gene Expression
Omnibus dataset GSE21656 submitted by Sun et al (20), microarray results demonstrated that
CHL1 expression in DDP-resistant H460 cells is significantly lower
compared with that in parental cells, suggesting that CHL1 may be
involved in NSCLC chemoresistance; however, to the best of our
knowledge, the underlying mechanism remains unknown.
In the present study, the expression of CHL1 in DDP-
and PTX-resistant A549 cells and the parental cells was assessed.
Functional studies of CHL1 were performed to investigate its
potential role in chemoresistance.
Materials and methods
Data processing
The human GSE21656 microarray dataset (20) was downloaded from the NCBI Gene
Expression Omnibus (GEO) database (www.ncbi.nlm.nih.gov/geo). The available dataset,
GSE21656 was based on the GPL6244 platform (Affymetrix Human Gene
1.0 ST Array, Affymetrix; Thermo Fisher Scientific, Inc.). This
data includes H460 cells and DDP-resistant H460 cells sample, and
each cell has three repeats samples. The online tool, GEO2R
(http://www.ncbi.nlm.nih.gov/geo/geo2r) (21) was used to determine the
differentially expressed genes in H460 and DDP-resistant H460
cells. P<0.05 and |log2fold-change|≥1 were set as
cut-off standards.
Cell culture
The human NSCLC cell line A549, the PTX-resistant
cell line A549/PTX and the DDP-resistant cells A549/DDP were
purchased from Procell Life Science & Technology Co., Ltd. The
cells were cultured in Ham's F-12K medium supplemented with 10%
fetal bovine serum (both purchased from Thermo Fisher Scientific,
Inc.), 100 U/ml penicillin and 100 U/ml streptomycin (cat. no.
15140122; Thermo Fisher Scientific, Inc.), in a 37°C humidified
incubator with 5% CO2.
Cell transfection
The resistant cells A549/PTX and A549/DDP cells were
transfected with 4.0 µg CHL1 recombinant expression plasmid (cat.
no. HG10143-NY; Sino Biological, Inc.). Empty vector
(pCMV3-SP-N-HA) was used as the control. A549 cells were
transfected with 100 pmol small interfering (si)RNAs. The siRNA
sequence for CHL1 (Guangzhou RiboBio Co., Ltd.) were siRNA-1,
5′-GGAGCUAAUUUGACCAUAUtt-3′, siRNA-2, 5′-CAGCAAUAUUAGCGAGUAUtt-3′
and scrambled control, 5′-UUCUCCGAACGUGUCACGUtt-3′. Plasmids and
siRNAs were transfected into cells using Lipofectamine®
2000 (Thermo Fisher Scientific, Inc.) following the manufacturer's
instructions. The time interval between transfection and subsequent
experimentation was 48 h. For the rescue experiments, the CHL1
silenced A549 cells were treated with the Akt inhibitor SC66 (cat.
no. S5313; Selleck Chemicals), along with DDP (1.5 µg/ml) or PTX
(35 ng/ml; both purchased from Selleck Chemicals) for 24 h at
37°C.
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR) assay
Total RNAs were isolated using TRIzol reagent
(Thermo Fisher Scientific, Inc.) according to the manufacturer's
instructions, and the mixed DNAs were eliminated by DNase I (New
England Biolabs, Inc.). First-strand cDNA synthesis was conducted
using the GoScriptTM kit (Promega Corporation) according to the
manufacturer's instructions. The reaction conditions for reverse
transcription were 25°C for 5 min, 42°C for 60 min and 70°C for 5
min. The SYBR Green Real-Time PCR Master mix (Thermo Fisher
Scientific, Inc.) was used to perform RT-qPCR, using a
LightCycler480 system (Roche Diagnostics GmbH). The CHL1 primer
sequences were as follows: Forward, 5′-GGCTTGGTCTCTTGCTTTCC-3′ and
reverse, 5′-ATCTTCCCTCCCTTTGCACG-3′; and β-actin forward,
5′-TTCCTTCCTGGGCATGGAGTC−3′ and reverse,
5′-TCTTCATTGTGCTGGGTGCC-3′. The following thermocycling conditions
were used for qPCR: 1 min at 95°C, followed by 40 cycles at 95°C
for 20 sec, 30 sec at 60°C and a final extension at 72°C for 30
sec. Each reaction was conducted in triplicate. Relative expression
levels were calculated using the 2−ΔΔCq method (22).
Cell viability
Cell viability was detected by MTT assay. A cell
suspension (100 µl) was seeded into 96-well plates at a density of
1×104 cells/well and incubated overnight at 37°C. The
concentrations of DDP used to treat A549 cells were 0.5, 1, 1.5, 2
and 2.5 µg/ml. While the concentrations of PTX used to treat A549
cells were 10,20,30,40 and 50 ng/ml. The concentrations of DDP used
to treat A549/DDP cells were 2, 4, 6, 8 and 10 µg/ml. While the
concentrations of PTX used to treat A549/PTX cells were 50, 100,
150, 200 and 250 ng/ml. After treating with different
concentrations of DDP or PTX for 48 h at 37°C, 100 µl MTT (5 mg/ml)
solution was added to each well and incubated for 4 h at 37°C.
Subsequently, 150 µl DMSO was added to each well to dissolve the
blue formazan crystals and the absorbance was measured using a
microplate reader (BioTek Instruments, Inc.) at 570 nm.
Clone formation assay
A total of 1×103 cells were seeded into a
35-mm dish (in triplicate) and maintained in F-12K medium with or
without DDP or PTX at 37°C for 48 h. A total of 2 weeks later,
cells were fixed in 4% paraformaldehyde for 15 min at room
temperature and stained with 0.01% crystal violet dye at room
temperature for 15 min. The rate of colony formation was calculated
using the following equation: (Number of colonies/number of seeded
cells) ×100.
Flow cytometry
Apoptosis was detected using a FITC Annexin V
Apoptosis kit (BD Pharmingen; BD Biosciences) according to the
manufacturer's protocol. Cells (1×105) were collected
and washed twice with PBS prior to being suspended in 500 µl
binding buffer. Subsequently, cells were incubated with 5 µl
Annexin V-FITC and 5 µl propidium iodide in the dark for 10 min at
room temperature and apoptosis was analyzed using a CytoFlex flow
cytometer (Beckman Coulter, Inc.). Data were analyzed using
CytEXpert 2.0 software (Beckman Coulter, Inc.). The ratio between
early and late apoptosis was calculated.
Western blotting
Cells were collected, washed twice with PBS and
lysed with RIPA lysis buffer (Thermo Fisher Scientific, Inc.).
Proteins were isolated from the cell lysis buffer and quantified
using the Piercetm™ BCA Protein Assay kit (cat. no. 23227; Thermo
Fisher Scientific, Inc.) with bovine serum album as a standard.
Equal amount of protein (30 µg) proteins were separated by
10% SDS-PAGE gel. Next, the proteins were transferred onto a
polyvinylidene membrane (Thermo Fisher Scientific, Inc.), blocked
with 5% BSA (Thermo Fisher Scientific, Inc.) for 2 h at 4°C, and
incubated overnight at 4°C with primary antibodies against CHL1
(1:500; cat. no. 25250-1-AP; ProteinTech, Inc.), multi-drug
resistance gene 1 (MDR1; 1:500; cat. no. 22336-1-AP; ProteinTech,
Inc.), multidrug resistance-associated protein (MRP; 1:500; cat.
no. 67228-1-Ig; ProteinTech, Inc.), low-density lipoprotein
receptor-related protein (LRP; 1:500; cat. no. 22336-1-AP;
ProteinTech, Inc.), phosphorylated (p)-Akt (1:1,000; cat. no.
ab38449; Abcam,) and Akt (1:2,000; cat. no. ab227385; Abcam). After
washing three times with PBS, the membrane was incubated with
horseradish peroxide-conjugated goat anti-rabbit (1:2,000; cat. no.
ab6271; Abcam)_or rabbit anti-mouse (1:2,000; cat. no. ab6728;
Abcam) secondary antibodies for 2 h at room temperature and the
blots were detected with enhanced chemiluminescence reagent (Thermo
Fisher Scientific, Inc.). Protein expression was quantified using
Image-pro plus 6.0 software (Media Cybernetics, Inc.).
Animal experiments
The animal experiments were approved by the Medical
Ethics Committee of Xiangya Changde Hospital (approval no.
20190325) and were performed in compliance with all regulatory
institutional guidelines for animal welfare (the National
Institutes of Health Publications no. 80-23) (23). A total of 12 male BALB/c-nu mice
(4-week-old, 20±5 g; Hunan SJA Laboratory Animals Center of the
Chinese Academy of Sciences) were used in this study. All animals
were kept at the SPF level laboratory at 20–25°C, a relative
humidity of 30–70%, a 12/12 h light/dark cycle and 10 times/h of
fresh air exchange. All mice were given free access to food and
water. The bedding materials, drinking water, feeding cages and
other items in contact with the animals were all autoclaved prior
to use. A549/DDP cells (1×107) transfected with empty
vector and CHL1 overexpression vector, using
Lipofectamine® 2000 reagent (Thermo Fisher Scientific,
Inc.), were subcutaneously injected into the nude mice to establish
xenograft models, following anaesthesia with 4% chloral hydrate
(400 mg/kg). Xenografts were allowed to grow to ~100 mm3
over 2 weeks and the mice were randomly divided into four groups
(n=3/group) as follows: i) vector group (A549/DDP cells transfected
with empty vector and treated with 100 µl saline solution); ii)
vector-DDP group (A549/DDP cells transfected with empty vector and
treated with 10 mg/kg DDP); iii) CHL1 group (A549/DDP cells
transfected with CHL1 overexpression vector and treated with 100 µl
saline solution) and iv) CHL1-DDP group (A549/DDP cells transfected
with CHL1 overexpression vector and treated with 10 mg/kg DDP). DDP
was administered by intraperitoneal injection every 3 days for 2
weeks. The mice were observed daily, and the tumors were measured
by a vernier caliper every 7 days. The tumor volumes were
calculated as length × width2/2. A total of 5 weeks
post-injection, mice were euthanized with CO2 at 30%
volume displacement rate (VDR) per min using a programmable logic
controller (Barry-Wehmiller Design Group, Inc.). Mice were
monitored continuously and once the mice were immobile (except for
breathing) for 1 min, the VDR was provided at 100% for 2 min. The
animals remained in the euthanasia chamber for 5 min and were then
observed for an additional 5 min. Breathing and heart rate were
monitored to determine death.
Statistical analysis
All experiments were performed in triplicate and
data are presented as the mean ± standard deviation. All
experiments were performed at least three times. Paired Student's
t-test was performed for comparisons between two groups and one-way
analysis of variance followed by Tukey's multiple comparison
post-hoc analysis was performed for comparisons between multiple
groups. SPSS 20.0 (IBM Corp.) was used to perform the analysis.
P<0.05 was considered to indicate a statistically significant
difference.
Results
CHL1 is downregulated in A549/DDP and
A549/PTX-resistant cells
In order to investigate the mechanism of
chemoresistance in lung cancer, the lung adenocarcinoma cell line
A549, the DDP-resistant cells (A549/DDP) and PTX-resistant cells
(A549/PTX) were used in the present study. Cells were exposed to
different concentrations of DDP (0–8 µg/ml) and PTX (0–160 ng/ml),
and MTT assay was used to detect the cell survival rate. A549/DDP
and A549/PTX cells demonstrated higher resistance to DDP and PTX
compared with A549 cells (Fig. 1A).
The half maximal inhibitory concentration (IC50) of DDP
was significantly higher in A549/DDP cells (8.30±0.92 µg/ml)
compared with A549 cells (1.68±0.18 µg/ml), and the IC50
of PTX was significantly higher in A549/PTX cells (174.80±8.64
ng/ml) compared with A549 cells (36.97±2.56 ng/ml; Fig. 1B). In addition, the expression levels
of the drug-resistant markers MDR1, MRP and LRP were significantly
higher in A549/DDP and A549/PTX cells compared with A549 cells
(Fig. 1C). Additionally, the mRNA
and protein expression levels of CHL1 were significantly lower in
A549/DDP and A549/PTX cells compared with those in A549 cells
(Fig. 1D and E), and this was also
observed in H460 DDP-resistant cells obtained from the GEO dataset
(GSE21656; Fig. 1F). These results
suggested that CHL1 may be involved in regulating DDP and PTX
resistance in NSCLC.
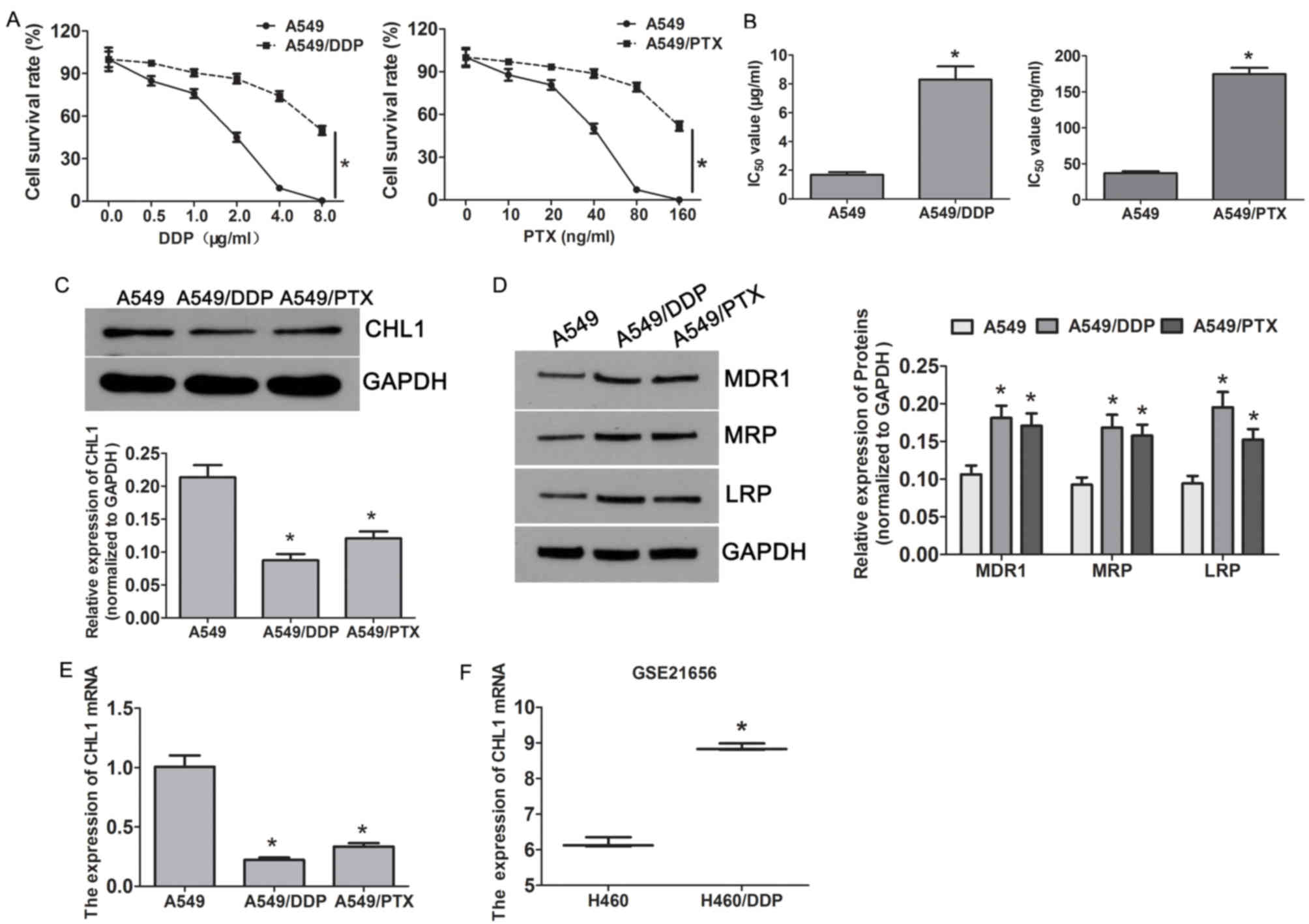 | Figure 1.CHL1 is downregulated in DDP and
PTX-resistant A549 cells. (A) Cell survival of A549 and
A549-resistant cells (A549/DDP and A549/PTX) treated with
increasing concentrations of DDP and PTX, as assessed by MTT assay.
(B) The IC50 values of DDP in A549/DDP and A549 cells,
and the IC50 values of PTX in A549/PTX and A549 cells.
*P<0.05 vs. A549 cells. (C) Western blotting demonstrated the
expression of drug resistance-related proteins MDR1, MRP and LRP in
A549 cells and A549-resistant cells (A549/DDP and A549/PTX).
*P<0.05 vs. A549 cells. The protein and mRNA expression levels
of CHL1 in A549 cells and A549-resistant cells (A549/DDP and
A549/PTX) were analysed by (D) western blotting and (E) reverse
transcription-quantitative PCR, respectively. *P<0.05 vs. A549
cells. (F) The mRNA expression of CHL1 in H460 and H460/DDP cells
in the GSE21656 dataset. *P<0.05 vs. H460 cells. CHL1, close
homolog of L1; DDP, cisplatin; PTX, paclitaxel; MDR1, multi-drug
resistance gene 1; MRP, multidrug resistance-associated protein;
LRP, low-density lipoprotein receptor-related protein; IC50, half
maximal inhibitory concentration. |
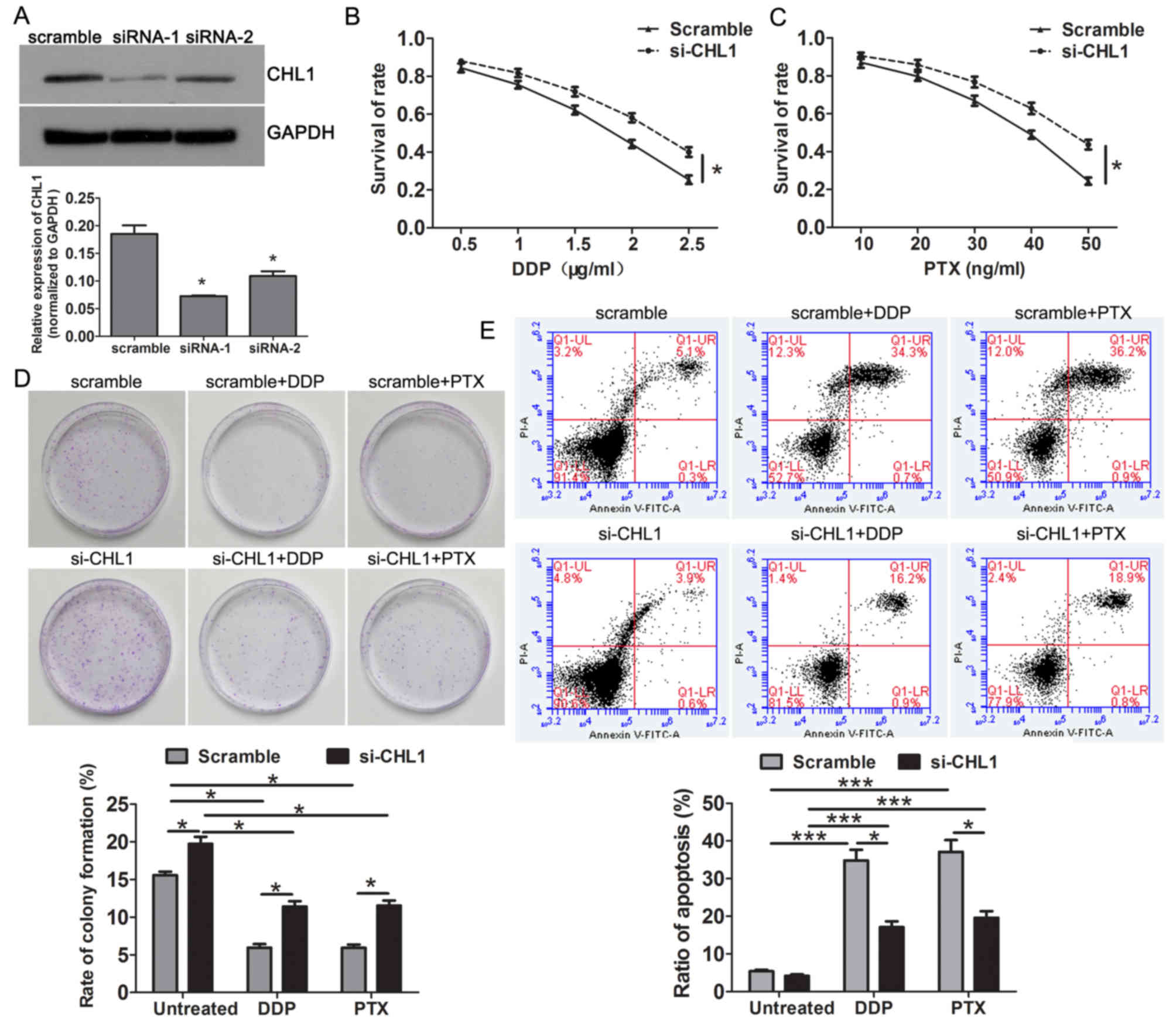
Knockdown of CHL1 enhances resistance
to DDP and PTX in A549 cells
As CHL1 was upregulated in A549 cells, CHL1 was
silenced in A549 cells using siRNAs. CHL1 expression was
significantly reduced in the CHL1 siRNA groups compared with that
of the scrambled control group (Fig.
2A). As siRNA-1 demonstrated the greatest interference
efficiency, it was selected for use in the following experiments.
Notably, CHL1-knockdown enhanced the resistance to DDP and PTX in
A549 cells (Fig. 2B and C). Colony
formation assay revealed that compared with the control group,
CHL1-knockdown significantly increased the rate of colony formation
in the absence of chemotherapeutics and enhanced the resistance to
DDP and PTX (Fig. 2D). Flow
cytometry results demonstrated significantly reduced apoptosis in
CHL1-knockdown cells after DDP and PTX treatment compared with that
of the control group (Fig. 2E).
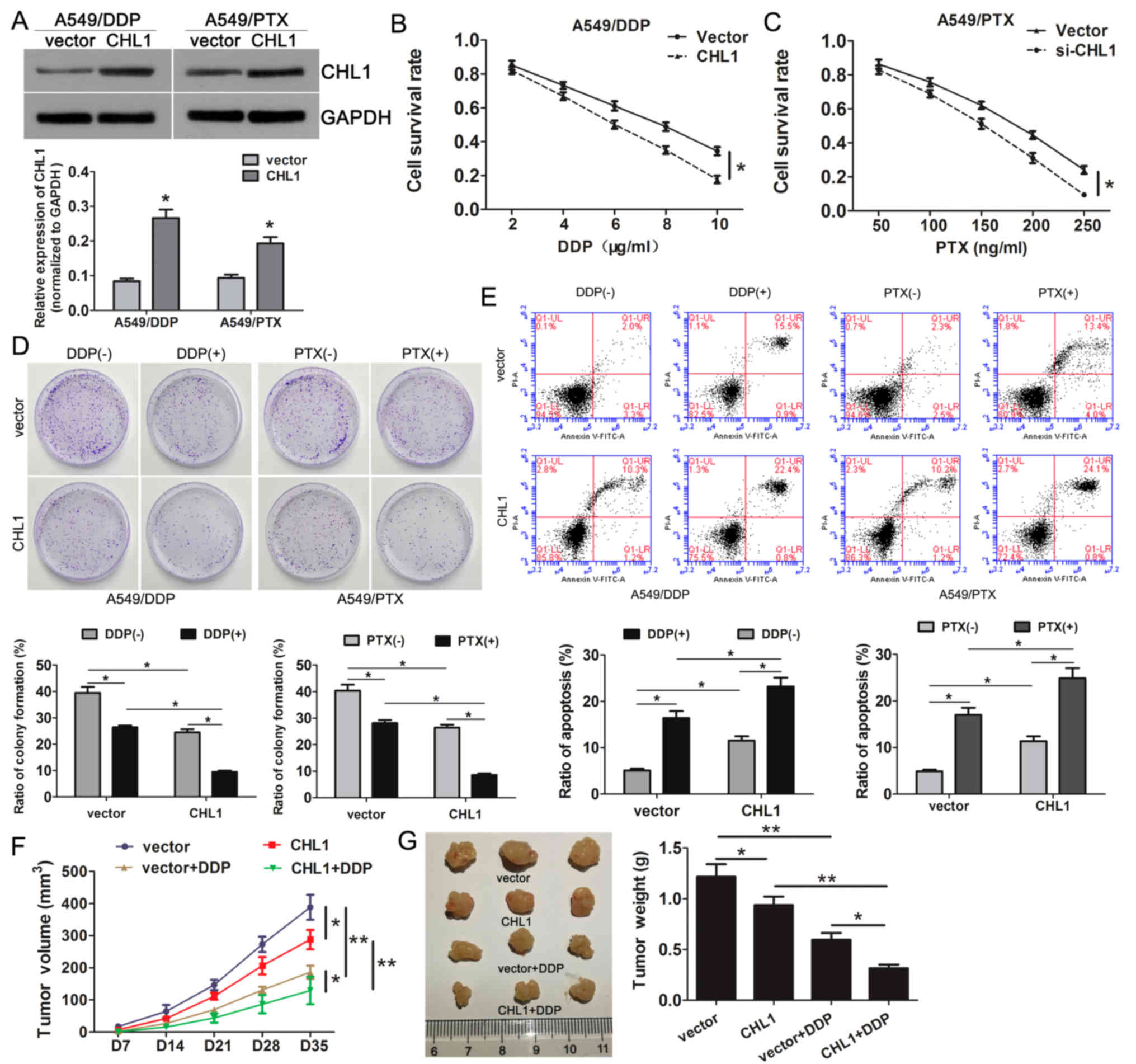
CHL1 overexpression enhances the
sensitivity of A549 resistant cells to DDP and PTX
As CHL1 is downregulated in A549/DDP and A549/PTX
cells, the present study successfully overexpressed CHL1 in these
cells using CHL1 recombinant expression plasmids (Fig. 3A). The results demonstrated that CHL1
overexpression alleviated the resistance to DDP and PTX compared
with that of the control group (Fig. 3B
and C). In addition, CHL1 overexpression inhibited colony
formation in the absence or presence of DDP and PTX (Fig. 3D). Additionally, flow cytometry
results demonstrated that restoration of CHL1 expression promoted
apoptosis in resistant cells following DDP and PTX treatment
(Fig. 3E).
To further validate the effects of CHL1
overexpression on DDP or PTX sensitivity, xenograft mice model
experiments were performed. The results demonstrated that CHL1
overexpression or DDP treatment significantly impeded the tumor
growth (Fig. 3F) and decreased the
tumor weight (Fig. 3G). In addition,
CHL1 overexpression further aggravated DDP-mediated repression on
tumor growth (Fig. 3F and G). These
data suggested that CHL1 overexpression suppressed tumor growth and
enhanced the chemosensitivity in NSCLC.
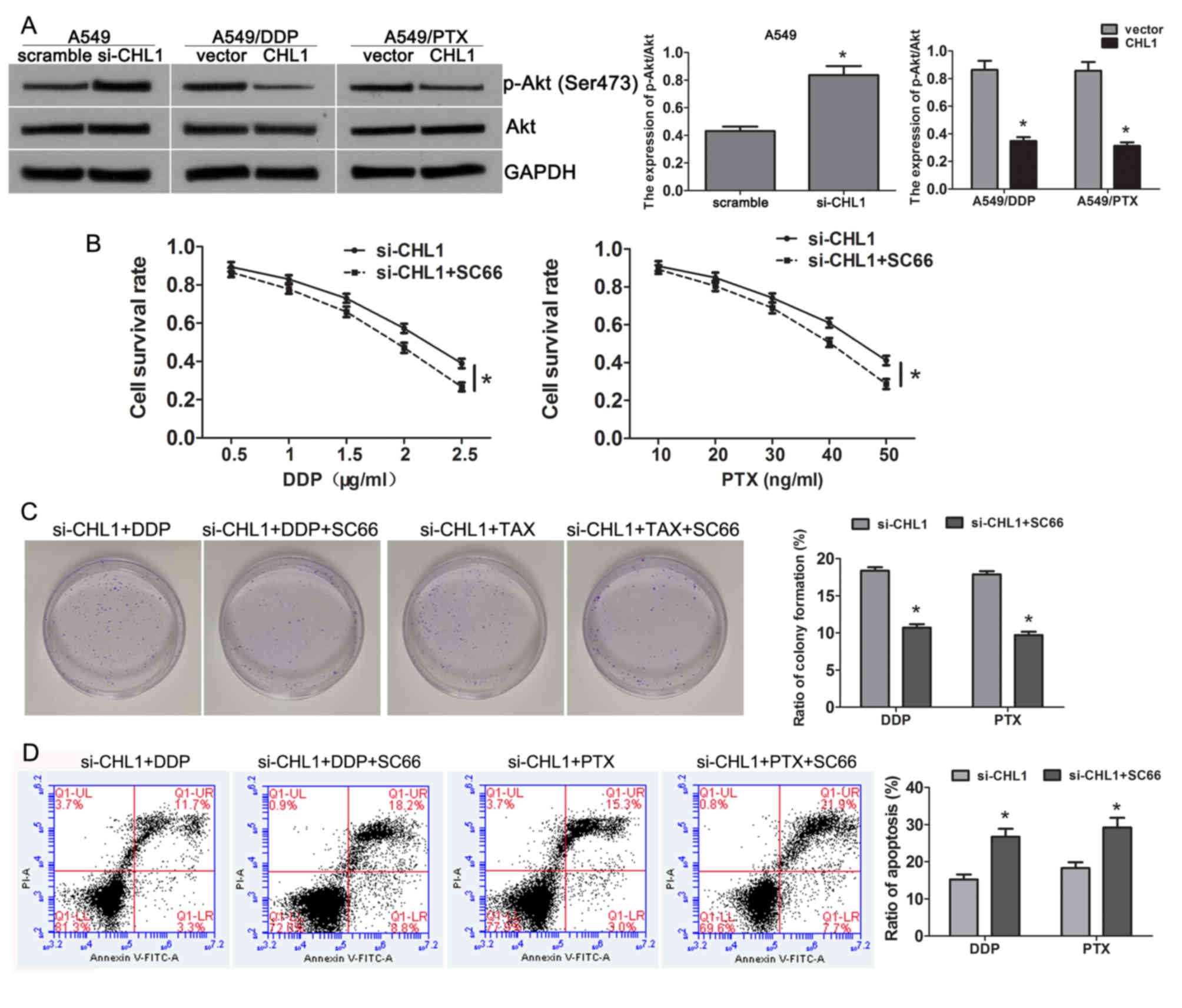
CHL1 mediates chemosensitivity by
inhibiting Akt activity
Recently, studies have confirmed that CHL1 inhibits
Akt activity in ESCC and neuroblastoma cell lines (11,24).
Thus, the present study investigated whether CHL1 mediates
chemoresistance via the Akt pathway in NSCLC. In A549 cells,
compared with the scrambled group, CHL1-knockdown elevated the
expression of p-Aktser473 (Fig. 4A). By contrast, restoring CHL1
expression in A549/DDP and A549/PTX cells inhibited the Akt
phosphorylation compared with the control group (Fig. 4A), suggesting CHL1 mediates
chemosensitivity via the Akt pathway. Subsequently, CHL1-silenced
A549 cells were treated with the Akt inhibitor SC66, and it was
demonstrated that inhibiting Akt activity significantly reduced the
promotive effects on cell survival (Fig.
4B) and clone formation (Fig.
4C), and the inhibitory effects on apoptosis (Fig. 4D) induced by CHL1-depletion. These
results confirmed that CHL1 mediates chemosensitivity in NSCLC by
inhibiting the Akt pathway.
Discussion
The results of the present study demonstrated that
CHL1 was significantly downregulated in A549/DDP and A549/PTX cells
compared with A549 cells. The knockdown of CHL1 in A549 cells
facilitated the cell survival and clone formation, and decreased
apoptosis when treated with or without DDP and PTX; whereas CHL1
overexpression in A549/DDP and A549/PTX cells inhibited cell
survival and clone formation, and increased apoptosis. The results
of the present study also demonstrated that CHL1 enhances NSCLC
chemosensitivity through inhibition of the Akt pathway. These data
suggested that CHL1 may be a promising target to improve the
efficacy of chemosensitivity in NSCLC.
CHL1 belongs to the L1 family of nerve cell adhesion
molecules, it was initially cloned in mice, and its expression in
mouse development was analyzed by Senchenko et al (25). Through cell-cell interactions and
mediating cell-cell and cell-matrix interactions, CHL1 has an
important effect on the development, regeneration and plasticity of
the nervous system (12). Previous
reports have demonstrated that CHL1 also participates in
carcinogenesis (11,15–18).
CHL1 was observed to be significantly downregulated in up to 11
types of tumor tissues compared with their adjacent normal tissues
(25). In most tumors, CHL1 is a
potential tumor suppressor gene whose silencing is associated with
tumor growth, invasion and metastasis (11,15–18). For
example, knockdown of CHL1 expression results in enhanced cervical
cancer cell invasion and migration (26,27). A
low expression of CHL1 in patients with neuroblastoma predicts a
poor prognosis, and enhancing CHL1 expression suppresses tumor
progression (24). In contrast, CHL1
has been reported to promote cell proliferation, metastasis and
migration in human gliomas (28).
However, to the best of our knowledge research on CHL1 and tumor
chemoresistance has rarely been reported.
The present study examined the differentially
expressed genes in NSCLC DDP-resistant cells in a GEO dataset. CHL1
was demonstrated to be upregulated in DDP-resistant cells compared
with parental cells, suggesting that CHL1 may be involved in NSCLC
chemotherapy resistance. Similarly, a study that compared and
analyzed the differentially expressed genes in chemosensitive
tumors and chemoresistant ovarian adenocarcinomas tissues reported
that the expression of CHL1 in chemotherapy-sensitive tumor tissues
is higher compared with that in drug-resistant tissues, suggesting
that CHL1 may help to predict the efficacy of chemotherapy for
ovarian cancer (29). In addition,
aberrant methylation of CHL1 may be associated with the recurrence
of colorectal cancer (CRC) following chemotherapy. 5-azadC
treatment restores 5-flurouracil sensitivity in vitro, which
also suggests that CHL1 may be involved in CRC chemotherapy
resistance (30). The results of the
present study demonstrated that CHL1 was downregulated in A549/DDP
cells. Additionally, as multiple drug resistance is a common
characteristic, another type of resistant cells (A549/TAX cells)
were also used in the current study. The results also demonstrated
that CHL1 was downregulated in A549/PTX cells. Compared with
control cells, overexpression of CHL1 significantly increased the
sensitivity of cells resistant to DDP and PTX, whereas knockdown of
CHL1 expression in parent A549 cells displayed the opposite
results. To the best of our knowledge, this study is the first
study to suggest that CHL1 may be involved in chemosensitivity in
lung cancer. The concentration of DDP used in vivo is 10
mg/kg (8,31), however, this may not be in line with
the concentrations that would be used in a clinical setting. In a
clinical trial, the human initial dose was calculated from the no
observed adverse effect levels (NOAELs) verified in animal
experiments. NOAEL is the maximum dose level without significant
adverse reactions. The NOAEL verified in animal experiments can be
converted to a human equivalent dose according to the body surface
area conversion, which is based on the area standardization
(mg/m2) proportional among different species (32). In the present study, the
concentration of DDP used in vivo was not the NOAEL, thus it
was not consistent with the concentrations used in clinical
settings.
Akt is a serine/threonine protein kinase that is
activated by phosphorylation (33).
As a key molecule of the PI3K/Akt signaling pathway, p-Akt
regulates cell survival, cell growth, cell motility and
angiogenesis, and prevents apoptosis (24). Additionally, Akt activation is
associated with tumor chemoresistance (33,34). The
results of the present study demonstrated that compared with the
control groups the expression of p-Akt was increased in
CHL1-knockdown A549 cells, and its expression was reduced in CHL1
overexpressed A549/DDP and A549/PTX cells. When Akt activity was
inhibited by the Akt inhibitor, the sensitivity to DDP and PTX in
CHL1-knockdown A549 cells was restored. This finding suggested that
CHL1 enhanced the chemosensitivity of NSCLC by inhibiting the Akt
pathway. Considering numerous studies have confirmed that the Akt
pathway mediates chemoresistance via regulation of ATP binding
cassette (ABC) members (35–37), the present study didn't further
investigate the specific ABC members and mechanisms, which was a of
the limitation to the present study; thus, this research should be
further investigated in vivo.
In summary, the present study demonstrated that CHL1
was downregulated in resistant cells A549/DDP and A549/PTX, and
upregulation of CHL1 enhanced the chemosensitivity of NSCLC via
inhibiting the Akt pathway. To the best of our knowledge, this was
the first study to confirm the function and mechanism of CHL1 in
mediating chemosensitivity in cancer. Thus, the development of
CHL1-based therapeutic strategies may improve the efficacy of
chemosensitivity in NSCLC.
Acknowledgements
The authors of the present study would like to thank
Mr. Dingliang Li (Xiangya Hospital, Changsha, China) for his
guidance and assistance in flow cytometric analysis.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author upon reasonable
request.
Authors' contributions
RH conceived and designed the present study. XC, BH,
YH and PL performed experiments and collected the data. SL, ZZ and
ZH analyzed and interpreted the data. ML and LZ analyzed the data
and prepared the figure. XC, ML and LZ drafted the initial
manuscript and revised it for intellectual content. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
The animal experiments were approved by the Medical
Ethics Committee of Xiangya Changde Hospital (Changde, China;
approval no. 20190325).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Parascandola M and Xiao L: Tobacco and the
lung cancer epidemic in China. Transl Lung Cancer Res. 8
(Suppl):S21–S30. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen W: Cancer statistics: Updated cancer
burden in China. Chin J Cancer Res. 27:12015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Oser MG, Niederst MJ, Sequist LV and
Engelman JA: Transformation from non-small-cell lung cancer to
small-cell lung cancer: Molecular drivers and cells of origin.
Lancet Oncol. 16:e165–e172. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Thatcher N, Faivre-Finn C, Blackhall F,
Anderson H and Lorigan P: Sequential platinum-based
chemotherapy-thoracic radiotherapy in early stage non-small cell
lung cancer. Clin Cancer Res. 11 (Suppl):S5051–S5056. 2005.
View Article : Google Scholar
|
|
5
|
Yano T, Okamoto T, Fukuyama S and Maehara
Y: Therapeutic strategy for postoperative recurrence in patients
with non-small cell lung cancer. World J Clin Oncol. 5:1048–1054.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fang Z, Chen W, Yuan Z, Liu X and Jiang H:
LncRNA-MALAT1 contributes to the cisplatin-resistance of lung
cancer by upregulating MRP1 and MDR1 via STAT3 activation. Biomed
Pharmacother. 101:536–542. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cai Y, Dong ZY and Wang JY: LncRNA NNT-AS1
is a major mediator of cisplatin chemoresistance in non-small cell
lung cancer through MAPK/Slug pathway. Eur Rev Med Pharmacol Sci.
22:4879–4887. 2018.PubMed/NCBI
|
|
8
|
Han ML, Zhao YF, Tan CH, Xiong YJ, Wang
WJ, Wu F, Fei Y, Wang L and Liang ZQ: Cathepsin L
upregulation-induced EMT phenotype is associated with the
acquisition of cisplatin or paclitaxel resistance in A549 cells.
Acta Pharmacol Sin. 37:1606–1622. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liu J, Meisner D, Kwong E, Wu XY and
Johnston MR: Translymphatic chemotherapy by intrapleural placement
of gelatin sponge containing biodegradable Paclitaxel colloids
controls lymphatic metastasis in lung cancer. Cancer Res.
69:1174–1181. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hassan WA, Yoshida R, Kudoh S, Kameyama H,
Hasegawa K, Niimori-Kita K and Ito T: Notch1 controls cell
chemoresistance in small cell lung carcinoma cells. Thorac Cancer.
7:123–128. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tang H, Jiang L, Zhu C, Liu R, Wu Y, Yan
Q, Liu M, Jia Y, Chen J, Qin Y, et al: Loss of cell adhesion
molecule L1 like promotes tumor growth and metastasis in esophageal
squamous cell carcinoma. Oncogene. 38:3119–3133. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu H, Focia PJ and He X: Homophilic
adhesion mechanism of neurofascin, a member of the L1 family of
neural cell adhesion molecules. J Biol Chem. 286:797–805. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tassano E, Biancheri R, Denegri L, Porta
S, Novara F, Zuffardi O, Gimelli G and Cuoco C: Heterozygous
deletion of CHL1 gene: Detailed array-CGH and clinical
characterization of a new case and review of the literature. Eur J
Med Genet. 57:626–629. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Morellini F, Lepsveridze E, Kahler B,
Dityatev A and Schachner M: Reduced reactivity to novelty, impaired
social behavior, and enhanced basal synaptic excitatory activity in
perforant path projections to the dentate gyrus in young adult mice
deficient in the neural cell adhesion molecule CHL1. Mol Cell
Neurosci. 34:121–136. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
He LH, Ma Q, Shi YH, Ge J, Zhao HM, Li SF
and Tong ZS: CHL1 is involved in human breast tumorigenesis and
progression. Biochem Biophys Res Commun. 438:433–438. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Martín-Sánchez E, Mendaza S,
Ulazia-Garmendia A, Monreal-Santesteban I, Blanco-Luquin I, Córdoba
A, Vicente-García F, Pérez-Janices N, Escors D, Megías D, et al:
CHL1 hypermethylation as a potential biomarker of poor prognosis in
breast cancer. Oncotarget. 8:15789–15801. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhu H, Fang J, Zhang J, Zhao Z, Liu L,
Wang J, Xi Q and Gu M: miR-182 targets CHL1 and controls tumor
growth and invasion in papillary thyroid carcinoma. Biochem Biophys
Res Commun. 450:857–862. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yu W, Zhu K, Wang Y, Yu H and Guo J:
Overexpression of miR-21-5p promotes proliferation and invasion of
colon adenocarcinoma cells through targeting CHL1. Mol Med.
24:362018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hötzel J, Melling N, Müller J, Polonski A,
Wolters-Eisfeld G, Izbicki JR, Karstens KF and Tachezy M: Protein
expression of close homologue of L1 (CHL1) is a marker for overall
survival in non-small cell lung cancer (NSCLC). J Cancer Res Clin
Oncol. 145:2285–2292. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sun Y, Zheng S, Torossian A, Speirs CK,
Schleicher S, Giacalone NJ, Carbone DP, Zhao Z and Lu B: Role of
insulin-like growth factor-1 signaling pathway in
cisplatin-resistant lung cancer cells. Int J Radiat Oncol Biol
Phys. 82:e563–e572. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Barrett T, Wilhite SE, Ledoux P,
Evangelista C, Kim IF, Tomashevsky M, Marshall KA, Phillippy KH,
Sherman PM, Holko M, et al: NCBI GEO: Archive for functional
genomics data sets-update. Nucleic Acids Res. 41((Database issue)):
D991–D995. 2013.PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen Y, Wang H, Xiong T, Zou R, Tang Z and
Wang J: The role of LASS2 in regulating bladder cancer cell
tumorigenicity in a nude mouse model. Oncol Lett. 14:5149–5156.
2017.PubMed/NCBI
|
|
24
|
Ognibene M, Pagnan G, Marimpietri D,
Cangelosi D, Cilli M, Benedetti MC, Boldrini R, Garaventa A,
Frassoni F, Eva A, et al: CHL1 gene acts as a tumor suppressor in
human neuroblastoma. Oncotarget. 9:25903–25921. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Senchenko VN, Krasnov GS, Dmitriev AA,
Kudryavtseva AV, Anedchenko EA, Braga EA, Pronina IV, Kondratieva
TT, Ivanov SV, Zabarovsky ER and Lerman MI: Differential expression
of CHL1 gene during development of major human cancers. PLoS One.
6:e156122011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chu Y, Ouyang Y, Wang F, Zheng A, Bai L,
Han L, Chen Y and Wang H: MicroRNA-590 promotes cervical cancer
cell growth and invasion by targeting CHL1. J Cell Biochem.
115:847–853. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Long MJ, Wu FX, Li P, Liu M, Li X and Tang
H: MicroRNA-10a targets CHL1 and promotes cell growth, migration
and invasion in human cervical cancer cells. Cancer Lett.
324:186–196. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang Z, Xie Q, Hu CL, Jiang Q, Shen HF,
Schachner M and Zhao WJ: CHL1 is expressed and functions as a
malignancy promoter in glioma cells. Front Mol Neurosci.
10:3242017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Choi CH, Choi JJ, Park YA, Lee YY, Song
SY, Sung CO, Song T, Kim MK, Kim TJ, Lee JW, et al: Identification
of differentially expressed genes according to chemosensitivity in
advanced ovarian serous adenocarcinomas: Expression of GRIA2
predicts better survival. Br J Cancer. 107:91–99. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Baharudin R, Ab Mutalib NS, Othman SN,
Sagap I, Rose IM, Mohd Mokhtar N and Jamal R: Identification of
predictive DNA methylation biomarkers for chemotherapy response in
colorectal cancer. Front Pharmacol. 8:472017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang CK, Zhang Y, Zhang ZJ, Qiu QW, Cao JG
and He ZM: Effects of VBMDMP on the reversal of cisplatin
resistance in human lung cancer A549/DDP cells. Oncol Rep.
33:372–382. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gao Y, Wang Y, Li Y, Han R, Li C, Xiao L,
Cho S, Ma Y, Fang C and Lee AW: Repeated sub-chronic oral toxicity
study of xylooligosaccharides (XOS) in dogs. Regul Toxicol
Pharmacol. 86:379–385. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wu YH, Huang YF, Chen CC and Chou CY: Akt
inhibitor SC66 promotes cell sensitivity to cisplatin in
chemoresistant ovarian cancer cells through inhibition of COL11A1
expression. Cell Death Dis. 10:3222019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Clark AS, West K, Streicher S and Dennis
PA: Constitutive and inducible Akt activity promotes resistance to
chemotherapy, trastuzumab, or tamoxifen in breast cancer cells. Mol
Cancer Ther. 1:707–717. 2002.PubMed/NCBI
|
|
35
|
Imai Y, Yamagishi H, Ono Y and Ueda Y:
Versatile inhibitory effects of the flavonoid-derived PI3K/Akt
inhibitor, LY294002, on ATP-binding cassette transporters that
characterize stem cells. Clin Transl Med. 1:242012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sims JT, Ganguly SS, Bennett H, Friend JW,
Tepe J and Plattner R: Imatinib reverses doxorubicin resistance by
affecting activation of STAT3-dependent NF-kappaB and HSP27/p38/AKT
pathways and by inhibiting ABCB1. PLoS One. 8:e555092013.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang X, He S, Gu Y, Wang Q, Chu X, Jin M,
Xu L, Wu Q, Zhou Q, Wang B, et al: Fatty acid receptor GPR120
promotes breast cancer chemoresistance by upregulating ABC
transporters expression and fatty acid synthesis. EBioMedicine.
40:251–262. 2019. View Article : Google Scholar : PubMed/NCBI
|