Introduction
The latest report by the World Health Organization
revealed that there were 288,000 new cases of melanoma and 61,000
deaths from melanoma in 2018 worldwide (1). A study by the American Cancer Society
revealed that melanoma ranked as the fifth most prevalent cancer
among men and the sixth among women, with a gradually growing
incidence over the past years, whereas the incidence of most solid
cancers has tended to drop or stabilize (2,3).
Melanoma, which accounts for ~70% of skin cancer deaths, is the
most lethal type of skin cancer, characterized by rapid
deterioration, early metastasis and high mortality, and derives
from the pigment-producing melanocytes (2,4).
Melanin, a natural pigment that is produced by a specialized group
of cells known as melanocytes and by melanoma cells, can affect the
behavior of melanoma cells or their surrounding environment, and
abnormal melanocyte proliferation can develop into melanomas
(5). Melanogenesis is subject to
complex regulatory control by multiple agents, involving pathways
that are activated by receptor-dependent and receptor-independent
mechanisms in hormonal, auto-, para- or intracrine manners, and is
a metabolic pathway characteristic for melanocytes, in which
L-tyrosine is transformed to heterogenous melanin biopolymer
through a series of oxidoreduction reactions (6–9).
Lee et al (10) have demonstrated the inhibitory effect
of pectolinarigenin on melanogenesis. In addition, induction of
melanogenesis in melanoma cells is associated with increases of
nuclear hypoxia-inducible factor 1 α (HIF1-α) expression
accompanied by the upregulation of multiple HIF-1-dependent genes
involved in the regulation of glucose metabolism, angiogenesis and
stress responses, which suggests that melanogenesis serves a key
role in the regulation of cell metabolism (11). In addition, the presence of melanin
pigment or active melanogenesis attenuates the sensitivity and
efficacy of chemo-, radio- or immunotherapy in patients with
melanoma; thus, the inhibition of melanogenesis may sensitize
melanoma cells and improve the outcome of radiotherapy in patients
with melanoma (12,13). Since pectolinarigenin inhibits
melanogenesis, it may be an effective approach for
anti-melanogenesis treatment (10).
Melanin is produced by melanocytes and melanoma cells, and can
transform melanocytes to proliferate abnormally and develop into
melanomas. Therefore, it was hypothesized in the present study that
melanogenesis may attenuate the cytotoxic effect of
pectolinarigenin on melanoma cells, and inhibition of melanogenesis
may sensitize melanoma cells (11–13).
This suggests that treatment with pectolinarigenin may be
considered as an alternative, physiologically based approach for
the treatment of melanoma.
However, the exact biological action of
pectolinarigenin on melanoma cells and its underlying mechanisms
have not been reported in detail. The present study aimed to
evaluate the potential effects of pectolinarigenin on melanoma cell
proliferation and migration and invasion in vitro, as well
as its underlying molecular mechanisms. Previous studies have
reported that 30 µM pectolinarigenin treatment suppressed melanin
biosynthesis without cytotoxicity in melan-a cells (10), whereas when treated with 20 µM
pectolinarigenin for 48 h, the viability of bone marrow macrophages
was not influenced (14), and ≥100
µM pectolinarigenin did not affect the proliferation of skin
fibroblasts 142BR, the IC50 of which was >100 µM
pectolinarigenin (15). Thus, the
present study proposed that within the 40 µM range of
concentration, pectolinarigenin may be safe for normal cells. In
the present study, we investigated the role of pectolinarigenin in
cell proliferation, cell apoptosis, and cell migration and invasion
in melanoma cancer cells to provide a new agent for an ant-melanoma
strategy.
Materials and methods
Chemicals and reagents
Pectolinarigenin, with a purity >98% as measured
by HPLC, was purchased from Sichuan Weikeqi Biological Technology
Co., Ltd. 3-(4,5-Dimethylthiazol-
2-yl)-2,5-diphenyltetrazoliumbromide (MTT), dimethyl sulfoxide
(DMSO), rhodamine 123 (Rh123) and 2′,7′-dichlor-
odihydrofluorescein diacetate (DCFH-DA) were purchased from
Sigma-Aldrich; Merck KGaA. Hoechst 33258 and the Annexin V-FITC and
propidium iodide (PI) apoptosis detection kit were purchased from
Nanjing KeyGen Biotech Co., Ltd. For western blotting experiments,
the primary antibodies against matrix metalloproteinase-9 (MMP9,
cat. no. 13667T), matrix metalloproteinase-2 (MMP2, cat. no.
4022s), TIMP2 (cat. no. 5738S), Bax (cat. no. 2772S), Bcl2 (cat.
no. 4223S), Cleaved Caspase-3 (cat. no. 9661S), Stat3 (cat. no.
9139S), phosphorylated (p)-Stat3Tyr705 (cat. no. 7145T)
and β-actin (cat. no. 3700T) were purchased from Cell Signaling
Technology, Inc., and the secondary antibodies were obtained from
OriGene Technologies, Inc. For in vitro assays,
pectolinarigenin was prepared as a 40 mM stock solution in DMSO and
stored at −20°C. The stock solution was diluted with the relevant
medium for subsequent applications.
Cell lines and culture
Non-pigmented human melanoma cell lines A375 and
CHL-1, the former of which is recalcitrant and exhibits high
malignancy, and mouse pigmented melanoma cells with high metastatic
potential B16-F10 (B16) were selected for the present study based
on their origin, malignant degree and metastatic ability. The three
cell lines were obtained from the American Type Culture Collection.
The A375 and CHL-1 cells were cultured in DMEM (Hyclone; Cyvita),
and the B16-F10 cells were cultured in RPMI-1640 medium (Hyclone;
Cyvita), and both mediums were supplemented with 10%
heat-inactivated fetal bovine serum (FBS, Gibco; Thermo Fisher
Scientific, Inc.) and 1% penicillin and streptomycin. All cells
were incubated at 37°C in a humidified atmosphere of 5%
CO2.
Cell viability assay
The viability of pectolinarigenin-treated melanoma
cells was determined by an MTT colorimetric assay. A375, B16 and
CHL-1 cells in the logarithmic phase were seeded into 96-well
plates at 2–4×103 cells in 100 µl per well for
adherence, followed by the addition of pectolinarigenin diluted in
culture medium corresponding to each cell line in a final volume of
100 µl and a final concentration of 0, 2.5, 5, 10, 20 or 40 µM. The
cells were then cultured in a 37°C CO2 incubator for 24,
48 and 72 h. Subsequently, 20 µl 5 mg/ml MTT solution was added
into each well and incubated for an additional 2–4 h at 37°C. The
liquid phase was discarded, and the purple-colored formazan
precipitates were solubilized by the addition of 150 µl DMSO to
each well. The viability of melanoma cells was assessed by a
Spectra MAX M5 microplate spectrophotometer (Molecular Devices,
LLC) at 570 nm.
Colony formation assay
A colony formation assay was used to determine the
effects of pectolinarigenin on the colony formation rate of
melanoma cells. A375, B16 and CHL-1 cells were seeded in 6-well
plates at 500–800 cells per well. At 24 h, the supernatants were
replaced with fresh media containing the indicated doses (0, 2.5,
5, 10, 20 or 40 µM) of pectolinarigenin, and the plates were
cultured for 10–15 days in a 37°C CO2 incubator.
Subsequently, the cells were washed with PBS, followed by fixing
with 100% methanol and staining with a 0.5% crystal violet solution
in the room temperature for ~15 min, after which the colonies
(>50 cells) were counted using light microscopy (Olympus
Corp.).
Morphological analysis of cell nuclei
by Hoechst 33258 staining
Cell shrinkage, chromatin condensation and apoptotic
bodies, which are typical morphologic features of apoptotic cells,
were used to evaluate cell apoptosis as previously described
(16). Briefly, A375 and B16 cells
were seeded onto 18-mm coverslips in a 6-well plate at density of
3–8×104 per well and incubated in 37°C overnight. After
the cells were seeded on the coverslips, the groups treated with
various concentrations (0–40 µM) of pectolinarigenin for 48 h were
washed with ice-cold PBS twice and fixed in ice-cold 100% methanol
for ~15 min. The cells were stained with the Hoechst 33258
solution, and images were captured under a Leica DM4000B
fluorescence microscope (Leica Microsystems, Inc.) to observe the
nuclear morphology and identify the apoptotic bodies.
Apoptosis assay by flow cytometry
(FCM)
Flow cytometry was utilized to test apoptosis using
an Annexin V-FITC/PI apoptosis detection kit according to the
manufacturer's instructions. Briefly, A375 and B16 melanoma cells
were treated with different doses of pectolinarigenin (0–40 µM) for
48 h, harvested and washed twice with ice-cold PBS, followed by the
addition of Annexin V-FITC and PI in the dark for 5 min in the room
temperature. The apoptotic state in each group was analyzed by FCM
(FACSCalibur; BD Biosciences), and Flow Jo software version 7.6.1
(Treestar) was used to analyze the data and determine the average
apoptotic rates of three independent experiments.
Detection of mitochondrial membrane
potential (ΔΨm)
Mitochondrial membrane potential was detected by FCM
using Rh123 staining. A375 and B16 cells, exposed to a range of
concentrations of pectolinarigenin (0–40 µM) for 48 h, were
collected, washed and incubated with 10 µM Rh123 for 30 min at 37°C
in the dark. Following the incubation, the cells were washed twice
with cold PBS, and the fluorescence was subsequently detected by
dyeing of Rh123 and FCM (as aforementioned and detected using the
FITC channel). The mean values of triplicate experiment results
were calculated.
Detection of reactive oxygen species
(ROS)
ROS was detected by DCFH-DA, which is hydrolyzed by
intracellular esterases to produce a non-fluorescent DCFH product
that can be oxidized by ROS and other oxidizing species to produce
a highly fluorescent DCF product. A375 (5×104 per well)
and B16 (5×104 per well) cells were treated with 0–40 µM
pectolinarigenin for 48 h, collected, washed and incubated with 10
µM (DCFH-DA) for 30 min at 37°C in the dark. Following the
incubation, the cells were washed twice with ice-cold PBS, and the
fluorescence was subsequently assessed by FCM. This experiment was
repeated three times.
Migration assay
Cell migratory ability was measured using a modified
wound healing assay. The cells were cultured in 6-well plates until
they reached 80% confluence. A sterile 10 µl plastic pipette tip
was used to create a wound in each well, followed by washing with
PBS. Fresh medium containing 2% FBS and pectolinarigenin (0, 10, 20
or 40 µM) was added to the well. After 48-h incubation at 37°C with
5% CO2, the cells that migrated into the wounded area
were photographed under a light microscope and the cells within the
area were counted manually. The assay was performed three
times.
Transwell migration and invasion
assays
A Boyden chamber (8-µm pore size) migration assay
was conducted as previously described, with several modifications
(16). Briefly, 1.0×105
A375 or B16 cells suspended in 100 µl serum-free medium were added
into the upper chamber of a 24-well Transwell insert, and 600 µl
culture medium (DMEM for A375 and 1640 for B16) with 10% FBS was
put into the lower chamber. In addition, pectolinarigenin (0, 10,
20 or 40 µM) was added into both chambers. The cells were incubated
at 37° for 48 h, and the migrated cells that were attached to the
lower surface of the Transwell membrane were fixed in 100% methanol
and stained with 0.5% crystal violet at room temperature for 15
min. The non-migrated cells on the upper surface of the membrane
were removed by cotton swabs. Images of migrated cells in three
randomly selected fields were captured and evaluated under a light
microscope.
The invasion assay was performed according to
methods outlined in previous studies (17). Briefly, 60 µl diluted Matrigel (BD
Biosciences) was placed in the upper chamber of a 24-well Transwell
plate and incubated at 37°C for polymerization. The lower chamber
was filled with 600 µl medium (DMEM for A375 and 1640 for B16)
containing 10% FBS. Cells (A375 or B16) (1.0×105) in 100
µl serum-free medium were added into the upper chamber and treated
with pectolinarigenin (0, 10, 20 or 40 µM). Following 48-h
incubation at 37°C, the rest of the invasion assay was conducted
using the same protocol as the migration assay. Invading cells in
three areas per well were counted, and the inhibition rate of
invasion was calculated. The two assays were performed three
times.
Western blot analysis
A375 and B16 cells (2×105 cells) were
seeded in petri dishes and treated with pectolinarigenin (0, 10, 20
or 40 µM) for 48 h at 37°C. The cells were lysed in ice-cold RIPA
buffer (Beyotime Institute of Biotechnology) (containing protease
inhibitor cocktail and phosphalase inhibitor cocktail) for 30 min
on the ice after washing twice with PBS. Protein lysates were
centrifuged at 12,000 × g for 15 min at 4°C to collect the
supernatant of the lysate. Protein concentrations were determined
by the BCA Protein Assay Kit (Pierce; Thermo Fisher Scientific,
Inc.) using known amounts of bovine serum albumin (Beyotime
Institute of Biotechnology) to standardize and equalize the protein
concentration before loading onto gels. Protein lysates (50 µg per
lane) were resolved by electrophoresis on 10 or 12% SDS
polyacrylamide gels and transferred onto PVDF membranes (Amersham;
Cytiva). The membranes were blocked with 5% skim milk in
Tris-buffered saline containing 1% Tween 20 (TBST; pH 7.4) at room
temperature for 1 h, and incubated overnight at 4°C with primary
antibodies. After washing five times with TBST at room temperature,
the membranes were incubated with a 1:2,000 dilution of the
relevant HRP-conjugated secondary antibody for 2 h at room
temperature, followed by five washes with TBST. The blots were
developed using an enhanced chemiluminescence (ECL) kit (Amersham;
Cytiva). A monoclonal β-actin antibody was used as an internal
control. ImagePro Plus v6.0 (Media Cybernetics) was used for
densitometry analysis, and normalized against β-actin expression.
Original images of the blots may be viewed in Supplementary
Figure 1.
Statistical analysis
The data are expressed as the mean ± SD of at least
three independent experiments. The differences between the groups
were examined by one-way ANOVA followed by Dunnett's post hoc test
using SPSS v16.0 software (SPSS, Inc.). Excel 2016 software
(Microsoft Corporation) was used to process the figures. P<0.05
was considered to indicate a statistically significant
difference.
Results
Pectolinarigenin inhibits melanoma
cell viability
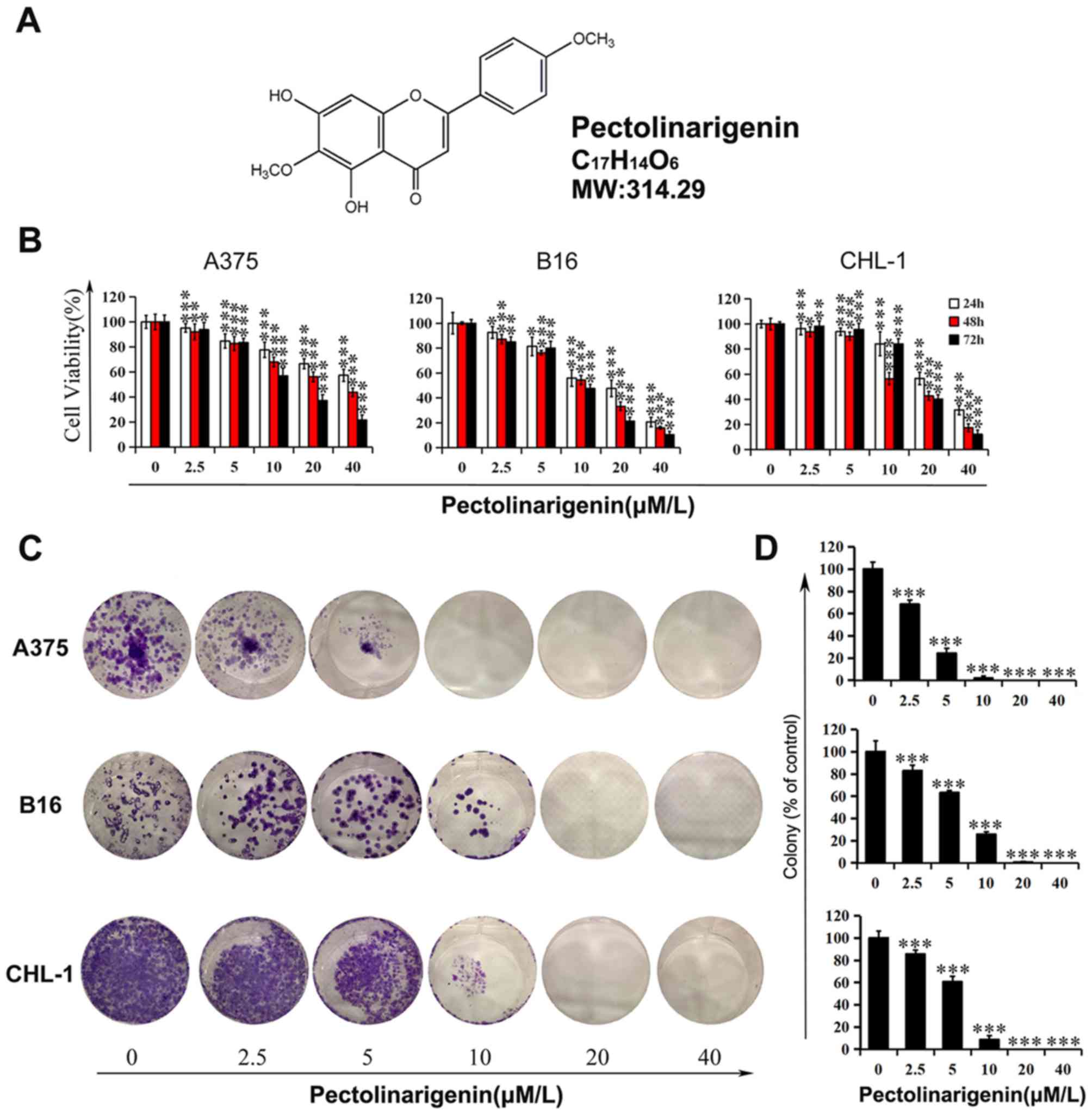
The chemical structure, molecular weight and
molecular formula of pectolinarigenin, a yellow powder composed of
flavonoids, are presented in Fig.
1A. First, cell viability was examined using a wide range of
pectolinarigenin concentrations in melanoma cells A375, B16 and
CHL-1 to analyze the toxic or protective function of
pectolinarigenin in the present study. The results demonstrated
that pectolinarigenin treatment reduced the viability of A375, B16
and CHL-1 cells compared with the untreated control groups, and the
effects appeared stronger with higher concentrations
(Fig. 1B). Following 48-h exposure
to 40 µM pectolinarigenin, the viability of the A375, B16 and CHL-1
cells was reduced to 43.79, 15.94 and 34.76% of that in the control
groups, respectively.
To validate the MTT assay results, a colony
formation assay was performed to detect the proliferation of
melanoma cells following continuous exposure to pectolinarigenin.
The results demonstrated that long-term treatment resulted in a
decrease in the colony formation of A375, B16 and CHL-1 cells
(Fig. 1C and D). More specifically,
pectolinarigenin exerted an inhibitory effect on colony number and,
with no visible colony formation observed in the high concentration
groups (20 and 40 µM). These results demonstrated that
pectolinarigenin inhibited the viability and proliferation in A375,
B16 and CHL-1 cells.
Pectolinarigenin induces apoptosis in
melanoma cells
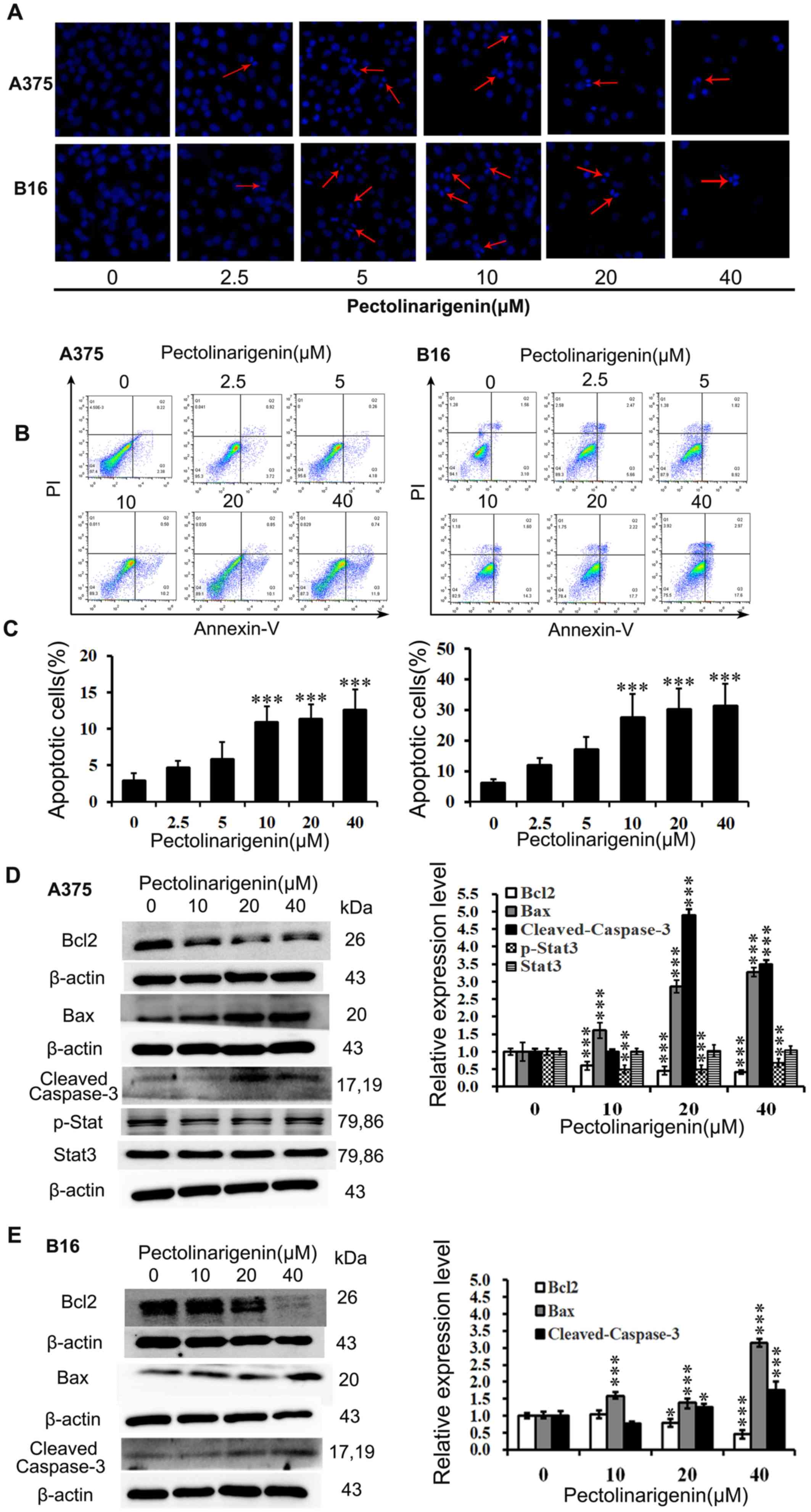
Hoechst 33258 staining assay was performed to
initially assess the effects of pectolinarigenin on the induction
of apoptosis in A375 and B16 cells. Upon exposure to
pectolinarigenin for 48 h, nuclear fragmentation and apoptotic
bodies in the treated groups were observed in the A375 and B16
cells (Fig. 2A). In addition, the
higher the pectolinarigenin concentration used, the more notable
the apoptotic hallmarks the lower number of living cells, thus
confirming that pectolinarigenin induced apoptosis in melanoma
cells.
To quantify the apoptotic rates induced by
pectolinarigenin, an Annexin V-FITC/PI assay was performed and
analyzed by FCM. The results demonstrated that pectolinarigenin
induced apoptosis in both A375 and B16 cells after treatment with
the indicated concentration of pectolinarigenin for 48 h (Fig. 2B and C). Treatment with 40 µM
pectolinarigenin led to an increase of the apoptotic rate by
>6-fold in A375 cells and 10-fold in B16 cells compared with the
respective untreated control groups. Thus, pectolinarigenin induced
apoptosis in A375 and B16 cells.
The expression levels of apoptosis-associated
proteins Bcl2, Bax and Cleaved Caspase-3 were determined by western
blotting. As presented in Fig. 2D,
in A375 cells, pectolinarigenin treatment reduced the levels of
Bcl2 expression, whereas those of pro-apoptotic proteins Bax and
Cleaved Caspase-3 were increased compared with the untreated
control group. In B16 cells, pectolinarigenin treatment
downregulated the expression of the anti-apoptotic Bcl2 and
upregulated the expression of pro-apoptotic Bax and Cleaved
Caspase-3; however, the loss of Bcl2 accumulation when B16 cells
were treated with the higher concentrations of pectolinarigenin may
have been affected by the different period of dose-response curve
(lag period and log period) (Fig.
2E). Thus, the higher concentrations of pectolinarigenin
induced more loss of Bcl2 in B16 cells.
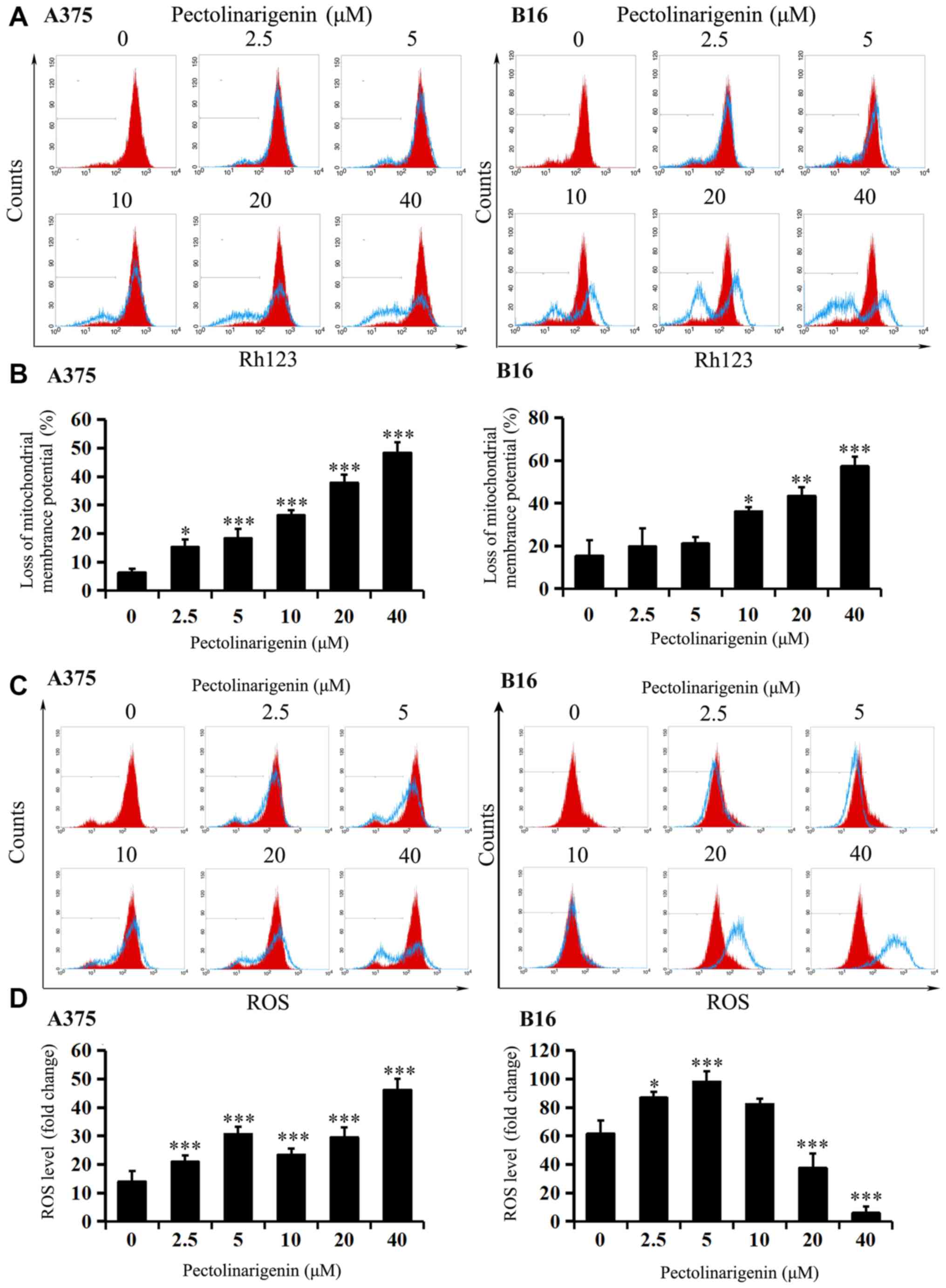
Effects of pectolinarigenin on ΔΨm and
ROS
To further test whether apoptosis induced by
pectolinarigenin was associated with the mitochondrial apoptotic
pathway, ΔΨm and intracellular ROS levels were measured.
After 48-h pectolinarigenin treatment, loss of ΔΨm (except
for the 2.5 and 5 µM in B16) compared with that in the untreated
controls was observed in both A375 and B16 cells (Fig. 3A and B), and the balance of cellular
ROS levels in the two cell types was visibly disturbed (Fig. 3C and D), that the changes of ROS
levels in the two cells varied significantly compared with the
control group (P<0.05). With the increasing of concentration,
ROS level may increase or decrease unsteadily. These results
suggested that the mitochondria-mediated apoptotic pathway may
serve an important role in pectolinarigenin-induced apoptosis in
B16 and A375 cells.
Pectolinarigenin decreases the
migratory and invasive abilities of A375 and B16 cells
Based on the antiproliferative and
apoptosis-inducing effects of pectolinarigenin, as well as on the
metastatic capability of tumor cells, which poses the predominant
threat in cancer-related mortality, the present study investigated
whether pectolinarigenin may affect the migration- and
invasion-associated abilities of melanoma cells. Wound healing and
Transwell assays were used to validate the antimetastatic effects
of pectolinarigenin on A375 and B16 cells.
In the wound healing assay with A375 and B16 cells,
significant reductions in cell migration were observed at 48 h in
the groups treated with pectolinarigenin compared with that in the
untreated control (Fig. 4A).
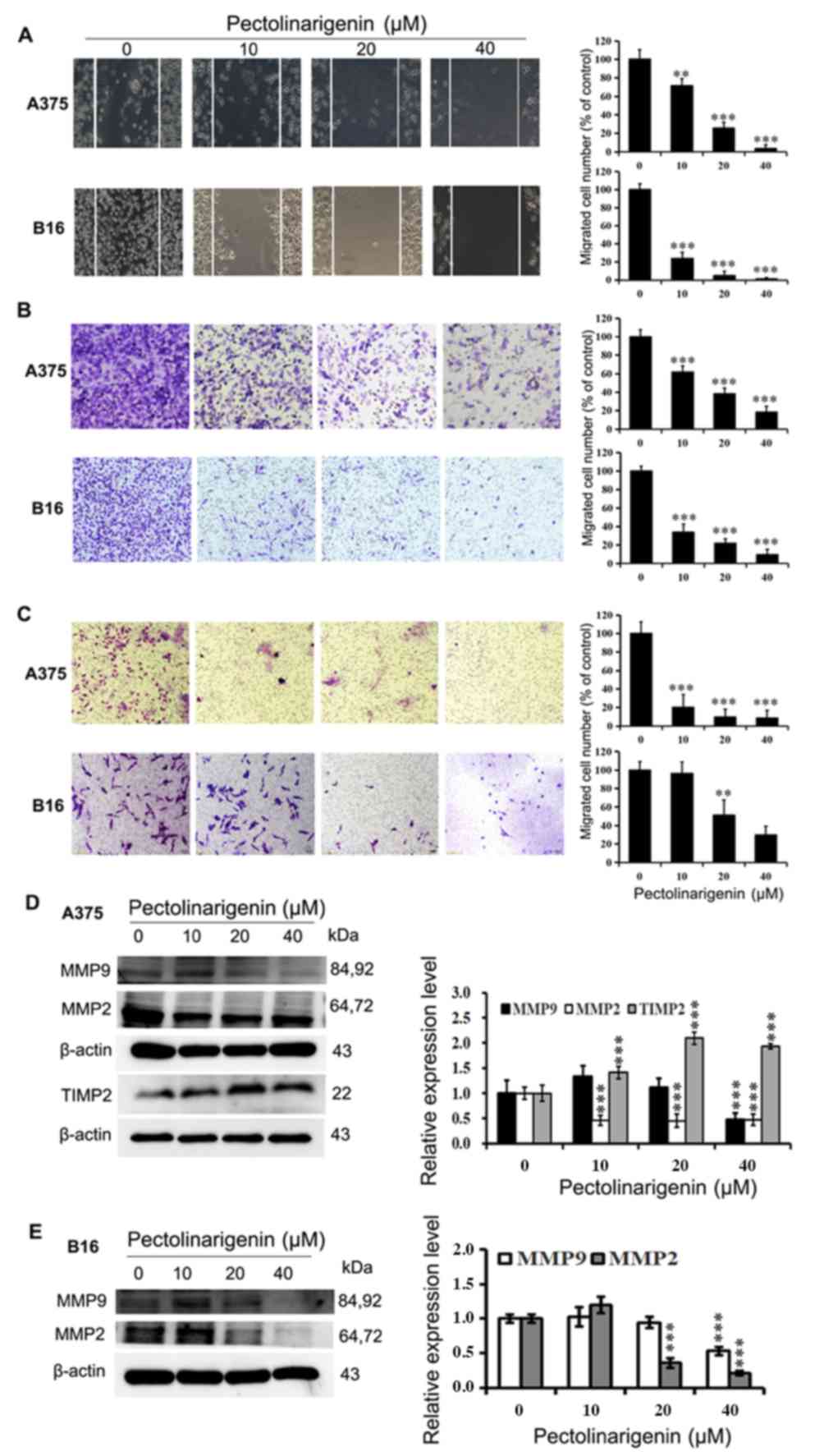 | Figure 4.Pectolinarigenin decreases A375 and
B16 cell migration and invasion. (A) Representative images from a
light microscope (×10 magnification) of the wound healing assay
using A375 and B16 cells treated with pectolinarigenin for 48 h;
The lines represent the area occupied by the initial scraping, and
migrated cells were counted. (B) Transwell migration assay of A375
and B16 cells upon treatment with the indicated concentration of
pectolinarigenin, after which the migrated cells were stained,
photographed and quantified (×20 magnification). (C) Transwell
invasion assay of A375 and B16 cells upon treatment with the
indicated concentration of pectolinarigenin, after which the
invading cells were stained, photographed and quantified (×20
magnification). (D and E) Expression of MMP2, MMP9, and TIMP2 were
determined by western blotting with β-actin as the internal
control. The protein band and the loading control that follows were
from the different parts of the same gel, and the loading controls
and protein bands were grouped together in the figure from
different gels. Data are presented as the means ± SD of at least
three independent experiments. **P<0.01 and ***P<0.001 vs. 0
µM pectolinarigenin. MMP, matrix metalloproteinase; TIMP2, tissue
inhibitor or metalloproteinases 2; p, phosphorylated. |
The results of the Transwell migration and invasion
assays demonstrated that the numbers of cells treated with
pectolinarigenin that migrated to the lower surfaces of the
Transwell membranes were reduced compared with the control group
cells. The migration and invasion of A375 and B16 cells were
notably diminished by pectolinarigenin (Fig. 4B and C). Taken together, these
results demonstrated that pectolinarigenin inhibited cell motility
and invasiveness in vitro.
The expression levels of proteins MMP2, MMP9, TIMP2,
Stat3, and p-Stat3(Tyr705), which are associated with
cell migration and invasion, were determined by western blotting.
The results demonstrated that the protein levels of MMP2 and MMP9
were decreased in the treated groups of B16 cells compared with
those in the untreated control group (Fig. 2D and 4D). A similar result was obtained in A375
cells (Fig. 4E), where
pectolinarigenin treatment also upregulated the protein expression
of TIMP2 and downregulated the expression of p-Stat3. These results
demonstrated that pectolinarigenin inhibited cell migration and
invasion in both A375 and B16 cells, which was associated with the
downregulation of MMP2 and MMP9 and/or the alteration of TIMP2 and
-Stat3.
Discussion
Melanoma is a highly malignant skin cancer with a
considerable metastatic potential, drug resistance and poor
treatment efficacy, as well as an increasing incidence in the last
decade (2,18). Thus, novel and efficient alternatives
to current anti-melanoma agents are urgently needed.
Extensive studies have demonstrated that components
derived from plants, which are generally considered safe, can exert
potent anticancer activities (19).
As a typical candidate, flavonoids have exhibited potential for
cancer suppression through apoptosis induction and other different
mechanisms, such as autophagy induction (20,21).
Pectolinarigenin is a natural extract from multiple herbal
medicinal plants, including Cirsium japonicum, Eupatorium
odoratum and Trollius chinensis (22). Previous studies have reported that
pectolinarigenin demonstrates potent inhibitory activity on
melanogenesis and effective antitumor activity in vivo and
in vitro (10,15).
To the best of our knowledge, there has been no
study on the biological activity of pectolinarigenin in melanoma
cells, and the precise role and mechanism of pectolinarigenin on
melanoma needs further research. To verify the anti-melanoma effect
of pectolinarigenin in this study, experiments were performed to
demonstrate the inhibitory effect of pectolinarigenin on the
proliferation and metastasis of melanoma cells. The results
demonstrated that pectolinarigenin suppressed the viability and
proliferation of melanoma cells. The next objective of the study
was to explore the effects of pectolinarigenin on apoptosis and on
the underlying mechanism. Apoptosis is an essential mechanism
targeted by numerous anticancer agents, characterized by shrinkage
of the cell and nucleus, chromatin condensation and finally the
formation of apoptotic bodies (23,24).
Thus, inducing apoptosis is regarded as a potential strategy for
cancer treatments (25). The results
of the Hoechst and the Annexin V-FITC/PI assays in the present
study confirmed that pectolinarigenin induced apoptosis in melanoma
cells. In addition, Bcl-2 family proteins are involved in the
induction of intrinsic apoptosis, with Bcl2 inhibiting apoptosis
and Bax promoting apoptosis in various cell types, such as prostate
and breast cancer cells (26). The
caspase family is considered to comprise critical regulators of
apoptosis initiation and execution, one of which, Caspase-3, is an
important effector protease that is cleaved and activated during
apoptosis (27). In the present
study, pectolinarigenin treatment significantly elevated the
expression of pro-apoptotic proteins Bax and Cleaved Caspase-3,
whereas the expression of the anti-apoptotic protein Bcl2 was
reduced, which suggested the occurrence of apoptosis induced by
pectolinarigenin and was consistent with previous reports that
demonstrated that pectolinarigenin exerted potent
anti-proliferative activity in breast cancer cells by inducing
apoptosis and the alteration of apoptosis-related proteins
(28). A375 and B16 cells are from
different origins, which may cause the differences in cell
sensitivity observed in the present study. Since pectolinarigenin
inhibits melanogenesis, it may be an effective approach for
anti-melanogenesis treatment (10).
In addition, inhibition of melanogenesis may sensitize melanoma
cells and improve the efficacy of therapy (13). Flavonoid extracts from flower of
Paeonia decomposita and Paeonia ostii inhibit
tyrosinase oxidase activity and downregulate the expression of
melanocortin 1 receptor (MC1-R), tyrosinase and
melanogenesis-related proteins (29). Pectolinarigenin, as a natural
flavonoid, may enhance the inhibitory effect of MC1-R expression in
B16, which is an MC1-R positive cell. Flavonoids target critical
oncogenic signaling pathways and receptors, such as PI3K/ATK, EGFR,
MAPK (30), which may affect the
sensitivity of different cell lines. Thus, subtle differences in
cell sensitivity were observed in the present study.
Excessive or low levels of ROS disrupt the
mitochondrial membrane potential and induce apoptosis, which is
associated with a decreasing mitochondrial transmembrane potential
(31,32). The results of the present study
demonstrated that the apoptosis induced by pectolinarigenin in
melanoma cells was associated with the loss of ΔΨm and
dysregulated ROS levels. Mitochondria and ROS-generating enzymes
can also be targeted by flavonoids; however, previous studies have
reported that flavonoids may serve either as ROS scavengers or
stimulators (30). These conflicting
results may be due to the potential of all antioxidants to be
converted into pro-oxidants (30).
It may be speculated that certain flavonoids may require a specific
dose range to act as antioxidants, whereas outside of this range,
they may stimulate ROS production. In the present study,
pectolinarigenin intervention affected the ROS levels in A375 and
B16 in different ways. In A375 cells, pectolinarigenin served as an
ROS stimulator to induce apoptosis. In B16 cells, low concentration
of pectolinarigenin (2.5 and 5 µM) served as a ROS stimulator,
whereas when the concentration was >5 µM, pectolinarigenin
exerted an antioxidant effect to decrease the ROS content. These
differences between A375 and B16 cells may have occurred due to the
varying sensitivities of the two cell types. The different
responses to pectolinarigenin in A375 and B16 melanoma cells from
different species are supported by a previous study Yang et
al (33) have demonstrated that
the ROS level in A375 cells treated with hinokiflavone was notably
increased, whereas that in B16 cells was first increased and then
decreased, and loss of ΔΨm and unbalanced the level of ROS
in both A375 and B16 cells was observed. Therefore, it is possible
that the ROS-mediated mitochondrial apoptotic pathway is involved
in melanoma cell death induced by pectolinarigenin.
Cancer is a complex process, and the hallmark
migratory and invasive ability of cancer cells is the primary cause
of cancer-associated mortality (34). Migration and invasion are
prerequisites for tumor metastasis, and suppressing the metastatic
capability of cancer cells is regarded as a principal strategy for
cancer therapy (35). Thus, the
present study conducted assays to evaluate the metastatic ability
of A375 and B16 cells following exposure to pectolinarigenin. The
results demonstrated that pectolinarigenin induced an inhibition of
cell migration and invasion compared with those in untreated cells.
In addition, the degradation of basement membranes is a key step in
cellular invasion and metastasis formation. Several lines of
evidence have implicated that MMPs serve a crucial role in these
complex multistep processes (36).
Two of these metalloproteinases, MMP2 and MMP9, have been reported
to be associated with metastasis, and cancer cell invasion and
migration into surrounding tissues are mediated by MMP2 and MMP9
(37,38). In addition, TIMP2, a specific
inhibitor of MMP2, is associated with the activation status of MMP2
(16). Stat3 is another important
transcription factor that is involved in cell proliferation,
survival, apoptosis and metastatic capability, which also has
important roles in cancer aggressiveness, making it an attractive
target for cancer therapy (39).
Thus, the present study assessed whether these
metastasis-associated proteins were associated with impaired
metastasis in melanoma cells by western blotting. The results
indicated that after treatment with pectolinarigenin, the
metastatic ability of B16 cells was inhibited via MMP2- and
MMP9-mediated pathways, and the metastatic ability of A375 was
inhibited via the Stat3/MMP2/MMP9 pathway. In addition, previous
studies have demonstrated that by interacting with the Stat3
pathway, pectolinarigenin can inhibit the proliferation, migration
and invasion of osteosarcoma (40),
breast cancer (39) and colorectal
carcinoma (41) cells, which is in
agreement with the results of the present study. In vitro
and in vivo experiments were carried out in the three cited
study, which demonstrated that pectolinarigenin suppressed
metastasis. Pectolinarigenin induced apoptosis in colorectal and
breast cancer cells by increasing the ratio of Bax/Bcl2 and the
expression of Cleaved Caspase-3 (39,41),
which was also demonstrated in the present study, and the
ROS-mediated mitochondrial apoptotic pathway was involved in the
pectolinarigenin induced apoptosis in melanoma cells. In addition,
Zhang et al (40) have
suggested that pectolinarigenin suppresses osteosarcoma growth and
metastasis by SHP-1-mediated Stat3 signaling inhibition in
vivo and in vitro, and pectolinarigenin significantly
suppresses osteosarcoma cell proliferation, induces apoptosis and
reduces the level of STAT3 downstream proteins cyclin D1, Survivin,
Bcl-2, B-cell lymphoma extra-large and myeloid cell leukemia 1. The
anti-melanoma effect of pectolinarigenin in vivo, and the
underlying mechanisms, need to be investigated in future
studies.
The results of the present study demonstrated that
pectolinarigenin exhibited antitumor activity on melanoma cells,
causing the inhibition of A375 and B16 cell viability,
proliferation, invasion and migration, and the induction of
apoptosis via the ROS-mitochondrial apoptotic pathway. As
pectolinarigenin inhibits melanogenesis, it may be an effective
approach for anti-melanogenesis therapies (10), and melanogenesis serves a key role in
the regulation of cellular metabolism (11). Of note, based on the present results
and on previous studies, pectolinarigenin may be offered as a new
potential agent for melanoma treatment based on its ability to
inhibit the cell proliferation, induce apoptosis and block
migration and invasion. However, there are certain limitations to
the present study. For example, in the wounding healing assay, the
same method was used to create identical wounds in each well at 0
h, and there were no differences between wells when observed under
a microscope; thus, images were not captured at 0 h, and the
relative wound closure could not be calculated. However, after
treatment for 48 h, the scratch was observed and images of every
well were captured under a microscope to count the cells that had
migrated into the wound area. Regarding the content of experiments,
the present study focused on the role of pectolinarigenin in
melanoma cell proliferation, apoptosis, migration and invasion;
this was a primary study on the anti-melanoma ability of
pectolinarigenin, providing a research basis for its anti-melanoma
activity and recognizing its anti-melanoma ability. The present
study had some limitations, including that on in vitro
studies were performed, without detecting changes in melanin
production in pigmented melanoma cells. In addition, the activity
of related enzymes, and related genes expression levels were not
analyzed. In our future studies, in vivo experiments will be
performed to validate the anti-melanoma effects of
pectolinarigenin. These studies will involve exploring the effect
of pectolinarigenin on tumorigenesis, malignancy and metastasis of
melanoma, and tissue inflammation and histopathological changes in
animal models to provide a scientific basis for its anti-melanoma
activity.
Overall, the results of the present study
demonstrated that pectolinarigenin significantly inhibited cell
proliferation, migration and invasion and induced apoptosis in
vitro via a ROS-mediated mitochondrial pathway and an MMP2 and
MMP9-mediated pathway. These results suggested that
pectolinarigenin may serve as a new agent for melanoma
treatment.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank the members of the
Tinghong Ye Lab (State Key of Laboratory of Biotherapy and Cancer
Center of West China Hospital, Sichuan University) for their
technical assistance.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YD and WY contributed to the conception and design
of the experiments. LW, QZ, YL, SY and CG performed the
experiments. XC made contributions to collection and analysis of
data. FH and TY made substantial contributions to the conception
and design of the study. All authors have read and approved the
final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 68:7–30. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zheng Y, Wang K, Wu Y, Chen Y, Chen X, Hu
CW and Hu F: Pinocembrin induces ER stress mediated apoptosis and
suppresses autophagy in melanoma cells. Cancer Lett. 431:31–42.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Xiong J, Wang Y, Gu Y, Xue Y, Dang L and
Li Y: CDK5RAP1 targeting NF-κB signaling pathway in human malignant
melanoma A375 cell apoptosis. Oncol Lett. 15:4767–4774.
2018.PubMed/NCBI
|
|
5
|
Slominski A, Zmijewski MA and Pawelek J:
L-tyrosine and L-dihydroxyphenylalanine as hormone-like regulators
of melanocyte functions. Pigment Cell Melanoma Res. 25:14–27. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Slominski A, Tobin DJ, Shibahara S and
Wortsman J: Melanin pigmentation in mammalian skin and its hormonal
regulation. Physiol Rev. 84:1155–1228. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Prota G: Melanins, melanogenesis and
melanocytes: Looking at their functional significance from the
chemist's viewpoint. Pigment Cell Res. 13:283–293. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Meredith P and Sarna T: The physical and
chemical properties of eumelanin. Pigment Cell Res. 19:572–594.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sarna T: Properties and function of the
ocular melanin-a photobiophysical view. J Photochem Photobiol B.
12:215–258. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lee S, Lee DH, Kim JC, Um BH, Sung SH,
Jeong LS, Kim YK, Kim SN, et al: Pectolinarigenin, an aglycone of
pectolinarin, has more potent inhibitory activities on
melanogenesis than pectolinarin. Biochem Biophys Res Commun.
493:765–772. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Slominski A, Kim TK, Brożyna AA,
Janjetovic Z, Brooks DLP, Schwab LP, Skobowiat C, Jóźwicki W and
Seagroves TN: The role of melanogenesis in regulation of melanoma
behavior: Melanogenesis leads to stimulation of HIF-1α expression
and HIF-dependent attendant pathways. Arch Biochem Biophys.
563:79–93. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Slominski A, Zbytek B and Slominski R:
Inhibitors of melanogenesis increase toxicity of cyclophosphamide
and lymphocytes against melanoma cells. Int J Cancer.
124:1470–1477. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Brożyna AA, Jozwicki W, Roszkowski K,
Filipiak J and Slominski AT: Melanin content in melanoma metastases
affects the outcome of radiotherapy. Oncotarget. 7:17844–17853.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang K, Lei J, He Y, Yang X, Zhang Z, Hao
D, Wang B and He B: A flavonoids compound inhibits osteoclast
differentiation by attenuating RANKL induced NFATc-1/c-Fos
induction. Int Immunopharmacol. 61:150–155. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bonesi M, Tundis R, Deguin B, Loizzo MR
and Menichini F, Tillequin F and Menichini F: In vitro biological
evaluation of novel 7-O-dialkylaminoalkyl cytotoxic
pectolinarigenin derivatives against a panel of human cancer cell
lines. Bioorg Med Chem Lett. 18:5431–5434. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Deng Y, Li Y, Yang F, Zeng A, Yang S, Luo
Y, Zhang Y, Xie Y, Ye T, Xia Y and Yin W: The extract from Punica
granatum (pomegranate) peel induces apoptosis and impairs
metastasis in prostate cancer cells. Biomed Pharmacother.
93:976–984. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li Y, Yang F, Zheng W, Hu M, Wang J, Ma S,
Deng Y, Luo Y, Ye T and Yin W: Punica granatum (pomegranate) leaves
extract induces apoptosis through mitochondrial intrinsic pathway
and inhibits migration and invasion in non-small cell lung cancer
in vitro. Biomed Pharmacother. 80:227–235. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tuong W, Cheng LS and Armstrong AW:
Melanoma: Epidemiology, diagnosis, treatment, and outcomes.
Dermatol Clin. 30:113–124. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Panth N, Manandhar B and Paudel KR:
Anticancer activity of Punica granatum (pomegranate): A review.
Phytother Res. 31:568–578. 2017. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang CZ, Calway TD, Wen XD, Smith J, Yu C,
Wang Y, Mehendale SR and Yuan CS: Hydrophobic flavonoids from
Scutellaria baicalensis induce colorectal cancer cell apoptosis
through a mitochondrial-mediated pathway. Int J Oncol.
42:1018–1026. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Elkady AI, Abu-Zinadah OA and Hussein RA:
Crude flavonoid extract of the medicinal herb nigella sativa
inhibits proliferation and induces apoptosis in breast cancer
cells. J Biomater Tiss Eng. 7:1235–1249. 2017. View Article : Google Scholar
|
|
22
|
Xu F, Gao X and Pan H: Pectolinarigenin
inhibits non-small cell lung cancer progression by regulating the
PTEN/PI3K/AKT signaling pathway. Oncol Rep. 40:3458–3468.
2018.PubMed/NCBI
|
|
23
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rahman N, Dhadi SR, Deshpande A and
Ramakrishna W: Rice callus suspension culture inhibits growth of
cell lines of multiple cancer types and induces apoptosis in lung
cancer cell line. BMC Complement Altern Med. 16:4272016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hunter AM, LaCasse EC and Korneluk RG: The
inhibitors of apoptosis (IAPs) as cancer targets. Apoptosis.
12:1543–1568. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Autret A and Martin SJ: Emerging role for
members of the Bcl-2 family in mitochondrial morphogenesis. Mol
Cell. 36:355–363. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Namura S, Zhu JM, Fink K, Endres M,
Srinivasan A, Tomaselli KJ, Yuan J and Moskowitz MA: Activation and
cleavage of caspase-3 in apoptosis induced by experimental cerebral
ischemia. J Neurosci. 18:3659–3668. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lu M, Kong Q, Xu X, Lu H, Lu Z, Yu W, Zuo
B, Su J and Guo R: Pectolinarigenin-a flavonoid compound from
cirsium japonicum with potential anti-proliferation activity
in MCF-7 breast cancer cell. Tropical J Pharma Res. 13:2252014.
View Article : Google Scholar
|
|
29
|
Guo L, Yin Z, Wen L, Xin J, Gao X and
Zheng X: Flower extracts from Paeonia decomposita and
Paeonia ostii inhibit melanin synthesis via
cAMP-CREB-associated melanogenesis signaling pathways in murine B16
melanoma cells. J Food Biochem. 43:e127772019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu-Smith F and Meyskens FL: Molecular
mechanisms of flavonoids in melanin synthesis and the potential for
the prevention and treatment of melanoma. Mol Nutr Food Res.
60:1264–1274. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Alfadda AA and Sallam RM: Reactive oxygen
species in health and disease. J Biomed Biotechnol.
2012:9364862012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang J and Yi J: Cancer cell killing via
ROS: To increase or decrease, that is the question. Cancer Biol
Ther. 7:1875–1884. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yang SP, Zhang YG, Luo Y, Xu B, Yao Y,
Deng Y, Yang F, Ye T, Wang G, Cheng Z, et al: Hinokiflavone induces
apoptosis in melanoma cells through the ROS-mitochondrial apoptotic
pathway and impairs cell migration and invasion. Biomed
Pharmacother. 103:101–110. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Talmadge JE and Fidler IJ: AACR centennial
series: The biology of cancer metastasis: Historical perspective.
Cancer Res. 70:5649–5669. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Fidler IJ: The pathogenesis of cancer
metastasis: The ‘seed and soil’ hypothesis revisited. Nat Rev
Cancer. 3:453–458. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kamran MZ and Gude RP: Preclinical
evaluation of the antimetastatic efficacy of pentoxifylline on A375
human melanoma cell line. Biomed Pharmacother. 66:617–626. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Fietz S, Einspanier R, Hoppner S, Hertsch
B and Bondzio A: Determination of MMP-2 and −9 activities in
synovial fluid of horses with osteoarthritic and arthritic joint
diseases using gelatin zymography and immunocapture activity
assays. Equine Vet J. 40:266–271. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Si L, Yan X, Hao W, Ma X, Ren H, Ren B, Li
D, Dong Z and Zheng Q: Licochalcone D induces apoptosis and
inhibits migration and invasion in human melanoma A375 cells. Oncol
Rep. 39:2160–2170. 2018.PubMed/NCBI
|
|
39
|
Li Y, Gan C, Zhang Y, Yu Y, Fan C, Deng Y,
Zhang Q, Yu X, Zhang Y, Wang L, et al: Inhibition of Stat3
signaling pathway by natural product pectolinarigenin attenuates
breast cancer metastasis. Front Pharmacol. 10:11952019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhang T, Li S, Li J, Yin F, Hua Y, Wang Z,
Lin B, Wang H, Zou D, Zhou Z, et al: Natural product
pectolinarigenin inhibits osteosarcoma growth and metastasis via
SHP-1-mediated STAT3 signaling inhibition. Cell Death Dis.
7:e24212016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Gan C, Li Y, Yu Y, Yu X, Liu H, Zhang Q,
Yin W, Yu L and Ye T: Natural product pectolinarigenin exhibits
potent anti-metastatic activity in colorectal carcinoma cells in
vitro and in vivo. Bioorg Med Chem. 27:1150892019. View Article : Google Scholar : PubMed/NCBI
|