Introduction
Colorectal cancer (CRC) is one of the most common
malignant diseases worldwide. A previous study showed that are high
rates of incidence and disease-associated mortality for CRC
(1). The development of effective
screening methods to detect cancer is vital as it is necessary for
early diagnosis and therapy. To provide novel biological insight
and advances in therapy, the molecular mechanisms underlying
malignant tumor occurrence and progression are being investigated,
with the aim of developing more effective therapies for CRC
(2).
MicroRNAs (miRNAs or miRs) are small non-coding RNA
molecules, which are endogenous and 18–25 nucleotides in length.
The function of miRNAs is to modulate the expression of target
mRNAs at a post-transcriptional level and the mechanism of action
is by inhibiting mRNA translation or inducing mRNA degradation by
binding to a target site (3,4). The target sites may be in the 3′
untranslated region (3′-UTR), coding region and the promoter
(5,6). It has been reported that miRNAs
regulate the expression of >60% of human protein-encoding genes
(7). These genes take part in
numerous biological processes, including cell proliferation,
differentiation, apoptosis, invasion and autophagy (8–10).
miR-424-5p, located at Xq26.3, it is differentially
expressed in tumor tissues and normal tissues adjacent to tumors,
and it serves different biological functions (11). The expression levels of miR-424-5p
are increased in pancreatic (12)
and gastric cancer (13); however,
the expression levels are decreased in hepatocellular carcinoma,
esophageal squamous cell carcinoma and cervical cancer (14–16). The
expression and function of miR-424-4p in CRC requires to be
reported.
The present study analyzed the miRNA expression
profiles using miRNA sequencing data from The Cancer Genome Atlas
(TCGA; http://cancergenome.nih.gov/) and
determined the cyto-biological effects of miR-424-5p on human CRC
cells.
Materials and methods
miRNA array in TCGA for heatmap
analysis
Data from TCGA, which is a large-scale cancer genome
project that provides genome sequencing data of 33 types of cancer
(http://cancergennme.nih.gov/) (17), were analyzed. The data included
microRNA sequencing, RNA-sequencing and clinical datasets for
various cancer types. In the present study, miRNA array data were
obtained from TCGA for heatmap analysis. The sequencing data of
miRNAs from 11 adjacent control samples and 619 CRC tissue samples
were downloaded from TCGA database and the expression matrix of the
miRNAs was aggregated.
Patients and samples
CRC and paired non-tumor adjacent tissue samples, as
well as clinicopathological data were collected from patients who
underwent radical resection between July 2016 and March 2018 at The
First Hospital of Hebei Medical University (Shijiazhuang, China). A
total of 59 pairs of CRC and non-tumor adjacent tissues were used
for the experiments. The adjacent tissue samples were obtained ≥5
cm from the edge of the tumor. The present study was approved by
the Ethics Committee of the First Affiliated Hospital of Hebei
Medical University (Shijiazhuang, China; approval nos. 2013106 and
2016004). All patients provided written informed consent for the
use of their samples. Following surgical removal, tissues were
immediately frozen in liquid nitrogen and stored at −80°C. The
pathological diagnosis of all cancer tissue samples was
adenocarcinoma or mucinous carcinoma, all the tissues were
pathologically confirmed as cancerous or normal. The
clinicopathological data of the patients are presented in Table I.
 | Table I.Clinicopathological characteristics
of the patients with colorectal cancer (n=59). |
Table I.
Clinicopathological characteristics
of the patients with colorectal cancer (n=59).
| Variable | Value, n | miR-424-5p
expression |
P-valuea |
|---|
| Age, years |
|
| 0.4995 |
|
<65 | 36 | 11.505 (1.895,
105.238) |
|
|
≥65 | 23 | 7.311 (1.986,
58.081) |
|
| Sex |
|
| 0.3716 |
|
Male | 35 | 12.381 (2.042,
105.420) |
|
|
Female | 24 | 6.503 (1.618,
97.400) |
|
| Dukes' stage |
|
| 0.0159 |
|
A,B | 32 | 2.695 (1.518,
43.650) |
|
|
C,D | 27 | 32.450 (5.696,
117.800) |
|
| Pathological
type |
|
| 0.0451 |
|
Adenocarcinoma | 39 | 12.380 (1.905,
170.100) |
|
|
Mucinous carcinoma | 20 | 4.848 (1.641,
26.720) |
|
| Depth of
invasion |
|
| 0.0450 |
|
T1,T2 | 23 | 3.770 (1.470,
14.670) |
|
|
T3,T4 | 36 | 33.650 (2.191,
117.400) |
|
| Location |
|
| 0.5943 |
|
Colon | 32 | 7.090 (1.916,
97.400) |
|
|
Rectum | 27 | 9.318 (1.905,
117.800) |
|
| Lymph node
metastasis |
|
| 0.6488 |
|
Absent | 30 | 7.075 (1.811,
104.900) |
|
|
Present | 29 | 9.318 (1.967,
108.600) |
|
| Distant
metastasis |
|
| 0.1495 |
|
Absent | 49 | 6.635 (1.854,
95.460) |
|
|
Present | 10 | 33.490 (8.706,
116.200) |
|
Cell culture and transfection
The human CRC cell line SW480 was obtained from
Professor Xiao-feng Sun (American Type Culture Collection).
Detection of mycoplasma free cell was performed using a Mycoplasma
Monitoring kit (Vazyme Biotech Co., Ltd.). Cells were cultured in
Dulbecco's modified Eagle's medium (DMEM; Gibco; Thermo Fisher
Scientific, Inc.) containing 10% fetal bovine serum (FBS; Gibco;
Thermo Fisher Scientific, Inc.) and 1% penicillin-streptomycin. All
cells were cultured in standard growth medium at 37°C in a
humidified atmosphere with 5% CO2. miR-424-5p mimic (50
nM; cat. no. HmiR-SN0494), miR-424-5p inhibitor (50 nM; cat. no.
HmiR-AN0494-SN-10), mimic control (50 nM; cat. no. CmiR-SN0001-SN)
and inhibitor control (50 nM; cat. no. CmiR-AN0001-SN) were
purchased from GeneCopoeia, Inc. Lipofectamine 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.) was used to perform transfection
according to the manufacturer's protocol.
Cell proliferation and colony
formation assay
To investigate the role of miR-424-5p in the
development of CRC, the cell viability was evaluated using a Cell
Counting Kit (CCK)-8 assay (Vazyme Biotech Co., Ltd.), according to
the manufacturer's instructions. SW480 cells were transfected with
miR-424 mimic, mimic control, miR-424 inhibitor or inhibitor
control for 48 h. Logarithmically growing cells were then seeded in
10-cm normal cell culture dishes (3,000 cells/dish) and incubated
for 14 days in medium with 10% FBS that was changed every week, and
cells were cultured at 37°C. Subsequently, the colonies were
stained with crystal violet (0.5%) for 1 h at room temperature and
the number of colonies was counted manually. The colony formation
ability was determined in triplicate in tissue culture dishes.
Cell migration and invasion assay
The cell motility capacities were determined using a
wound-healing assay and Transwell assay. The wound-healing assay
was performed in 6-well culture plates; SW480 cells were seeded in
the plate at a density of 50×105 cells/well in culture
medium. Following transfection for 48 h, the cells reached 100%
confluence and confluent monolayers were scratched with a sterile
micro-pipette tip (200 µl), followed by washing with PBS to remove
floating cells. The width of the scratch was then measured at 0,
24, 48 and 72 h of incubation in serum-free medium. A total of
three independent experiments were performed.
For migration assays, transfected cells
(20×105 per well) in 100 µl serum-free DMEM were plated
in the top chamber of a Transwell system (Corning, Inc.), which
consisted of 12 wells with an 8.0-µm pore diameter polycarbonate
membrane insert. The insert was placed onto a 24-well plate, the
bottom chambers of which were filled with 600 µl DMEM with 10% FBS
as a chemoattractant. Following 48 h of incubation, the cells
remaining on the upper surface of the filter were removed by
scraping with a cotton swab, while the cells that had transgressed
to the lower surface of the filter were fixed with Diff Quik Fix
(mainly methanol) for 2 min, followed by staining with Diff Quik I
(eosin) for 2 min and staining with Diff Quik II (mainly methylene
blue) for 45 sec. All Diff Quik products were purchased from Zhuhai
Beisuo Biology Co, Ltd (BA-4150; www.baso.com.cn). A total of three independent
experiments were performed.
For invasion experiments, transfected cells
(20×105 per well) in 200 µl FBS-free DMEM were cultured
in the top chamber of a Transwell system that included a Matrigel
Invasion Chambers (cat. no. 354480; Corning, Inc.), while the
bottom well contained 800 µl medium with FBS. Following incubation
for 48 h, cells that had migrated to the lower side of the chamber
were stained as specified above. A total of three independent
experiments were performed.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from the frozen tissues
using TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol. The concentration and
purity of all RNA samples were detected using a NanoDrop ND-2000
spectrophotometer (NanoDrop Technologies; Thermo Fisher Scientific,
Inc.).
cDNA synthesis of miRNA was performed using the
All-in-One miRNA First-Strand cDNA Synthesis Kit (QP013;
GeneCopoeia, Inc.), according to the manufacturer's protocol. qPCR
was performed using an All-in-One™ miRNA qPCR kit (QP015;
GeneCopoeia, Inc.) with a Roche LightCycler 480 II Real-Time PCR
system (Roche Diagnostics) according to the manufacturer's
protocol. The sequence-specific primers for mature miR-424-5p (cat.
no. HmiRQP0494) and the U6 internal control (cat. no. HmiRQP9001),
as well as the Uni-miR qPCR Primer were purchased from GeneCopoeia,
Inc. The relative expression of miR-424-5p was calculated using the
2−ΔΔCq method with normalization to the expression of U6
(18).
A total of 1 µg RNA was reverse-transcribed into
cDNA using the PrimeScript RT kit (Takara Bio Inc.) for mRNA
detection, at 37°C for 15 min, and 85°C for 5 sec. qPCR was
subsequently performed using the Power SYBR® Green
Master mix (Applied Biosystems; Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol (step 1: 95°C for 10 min;
step 2 for 40 cycles: 95°C for 15 sec, 60°C for 1 min). The
following primer pairs were used for the qPCR: WNT6, Forward:
5′-CGTAGGGCGGTCACGATG-3′ and reverse: 5′-AACTGGAACTGGCACTCTCG-3′;
WNT11, forward: 5′-TCTTTGGGGTGGCACTTCTC-3′ and reverse:
5′-TGCCGAGTTCACTTGACGAG-3′; DVL1, forward:
5′-CCTCACTAACCAGCTCCGTG-3′ and reverse: 5′-ACGTGTGTGTACAGCCAGTC-3′;
RAC3, forward: 5′-CCTTCGAGAATGTTCGTGCC-3′ and reverse:
5′-TTCACAGAGCCAATCTCCCG-3′; FZD9, forward:
5′-GAACCCCACACACCTCTAGC-3′ and reverse: 5′-CTAATGAGCCTCACGGGGTC-3′;
SFRP, forward: 5′-GCTGCACATGAAGAATGGCG-3′ and reverse:
5′-GTAGTAGAGGGAGCAGGGGT-3′; FZD2, forward:
5′-GCGAAGCCCTCATGAACAAG-3′ and reverse: 5′-CTCCGTCCTCGGAGTGGTTC-3′
and GAPDH, forward: 5′-ACCCACTCCTCCACCTTTG-3′ and reverse:
5′-CTCTTGTGCTCTTGCTGGG-3′. The expression levels were normalized to
the internal reference gene GAPDH and quantified using the
2−ΔΔCq method (18).
Transcriptome sequencing and
differential expression analysis
To identify small RNAs and the target genes, SW480
colon cancer cells were transfected with miR-424-5p mimic and mimic
control, and following transfection for 48 h, the total RNA was
extracted for transcriptome sequencing. A total amount of 3 µg RNA
per sample was used for the RNA sample preparations. Sequencing
libraries were generated using the NEBNext® Ultra™ RNA
Library Prep kit for Illumina® (New England Biolabs)
following the manufacturer's protocol and index codes were added to
assign sequences to each sample. Library quality was assessed on
the Agilent Bioanalyzer 2100 system (Agilent Technologies, Inc.).
The clustering of the index-coded samples was performed on a cBot
Cluster Generation system using the TruSeq PE Cluster kit
v3-cBot-HS (Illumia, Inc.) according to the manufacturer's
protocol. Following cluster generation, the library preparations
were sequenced on an Illumina Hiseq platform (Illumina, Inc.).
Raw data (raw reads) in fastq format were first
processed through in-house perl scripts. Reference genome and gene
model annotation files were downloaded from the genome website
directly. An index of the reference genome was built using Hisat2
v2.0.5 and the Fragments Per Kilobase of transcript per Million
mapped reads (FPKM) of each gene were calculated based on the
length of the gene and reads count mapped to this gene. The FPKM
results were uploaded to Baidu (https://pan.baidu.com/s/1PxiK87dIbhfgZUL2y9nqZg)
(extraction code, djau). Differential expression analysis of
miR-424-5p mimic/mimic control was performed using the DESeq2
package of R (1.16.1). The resulting P-values were adjusted using
the Benjamini and Hochberg approach for controlling the false
discovery rate. DEGs were screened from three independent
transcriptome sequencing by DESeq2 and genes with an adjusted
P<0.05 were considered as DEGs.
Subsequently, disease oncology analysis using the
Database for Annotation, Visualization and Integrated Discovery
(DAVID, http://david.ncifcrf.gov/) and Kyoto
Encyclopedia of Genes and Genomes Orthology Based Annotation System
(http://kobas.cbi.pku.edu.cn/index.php), gene ontology
(GO) enrichment and Kyoto Encyclopedia of Genes and Genomes (KEGG)
pathway analyses of the DEGs were performed using the
clusterProfiler package of R.
Statistical analysis
For continuous variables, values are expressed as
the median ± standard deviation or the median and 25th and 75th
percentiles (interquartile range). All experiments were performed
in triplicates. Differences between groups were analyzed using the
Mann-Whitney U-test, two-way analysis of variance and correction
for multiple comparisons using the Sidak statistical hypothesis
test or an unpaired Student's t-test. P<0.05 was considered to
indicate a statistically significant difference. SPSS v21.0 (IBM,
Corp.) and GraphPad Prism v7.0 (GraphPad Software, Inc.) were used
for statistical analysis.
Results
miR-424-5p is upregulated in human CRC
tissues
Heatmap analysis of the miRNA expression profiles of
619 CRC and 11 normal colorectal tissues was first performed using
the TCGA database. From the series matrix files, 276 DEGs were
identified, among which 180 were upregulated and 96 were
downregulated (Table SI). The top
30 upregulated miRNAs are presented in a heatmap in Fig. 1A. To further confirm these results,
the expression of miR-424-5p in 59 pairs of CRC tissue and
corresponding non-tumor colorectal tissue samples was analyzed. The
expression of miR-424-5p in the CRC tissues was significantly
higher compared with that in the corresponding non-tumor colorectal
tissues (P=0.0056; Fig. 1B). The
clinicopathological characteristics [age (median=62, 25% percentile
(P25)=50.5, P75=72.5); sex (male:female=35:24); Dukes' stage
(A,B:C,D=32:27); pathological type (adenocarcinoma: Mucinous
carcinoma=39:20); depth of invasion (T1,T2:T3,T4=23:36); location
(colon:rectum=32:27); lymph node metastasis (absent:present=30:29);
distant metastasis (absent:present=49:10)] of the patients are
presented in Table I. In summary,
the results revealed that mR-424-5p is upregulated in CRC
tissues.
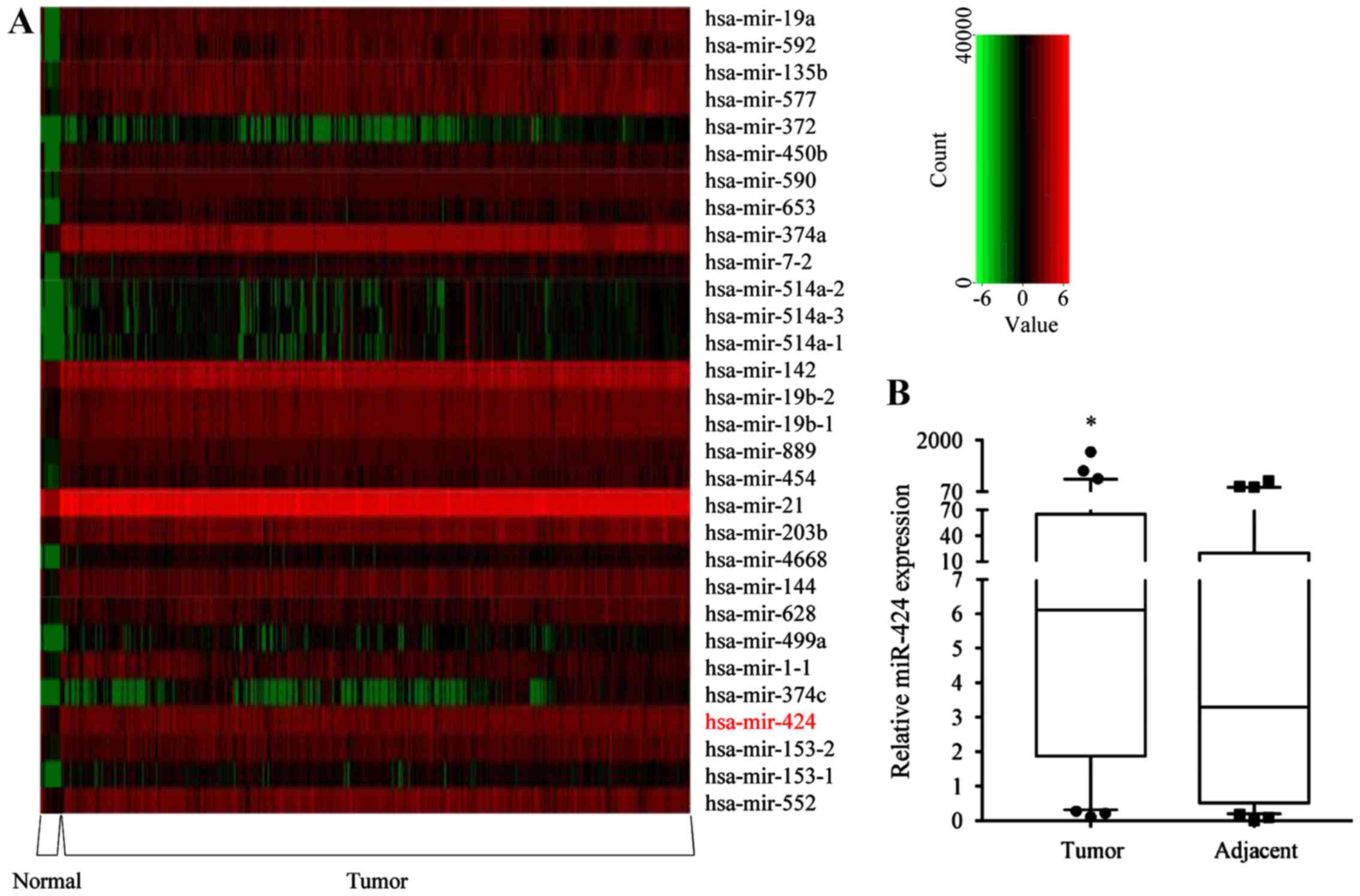 | Figure 1.Expression of miR-424-5p is
upregulated in human CRC tissues. (A) Heat-map analysis of the top
30 upregulated miRNAs in 619 CRC and 11 normal colorectal tissues
from The Cancer Genome Atlas database. The green/black/red color
indicates low/medium/high miRNA expression. (B) miR-424-5p
expression was determined in 59 CRC tissues (median=6.12, P25=1.87,
P75=65.12) and adjacent normal tissues (median=3.29, P25=0.51,
P75=19.84) by RT-qPCR analysis. The inter-quartile ranges were
shown in the box diagram. *P<0.05 vs. adjacent normal
tissues, according to the Mann-Whitney U-test. miRNA/miR, microRNA;
CRC, colorectal cancer; hsa, Homo sapiens; P25, 25%
percentile; P75, 75% percentile. |
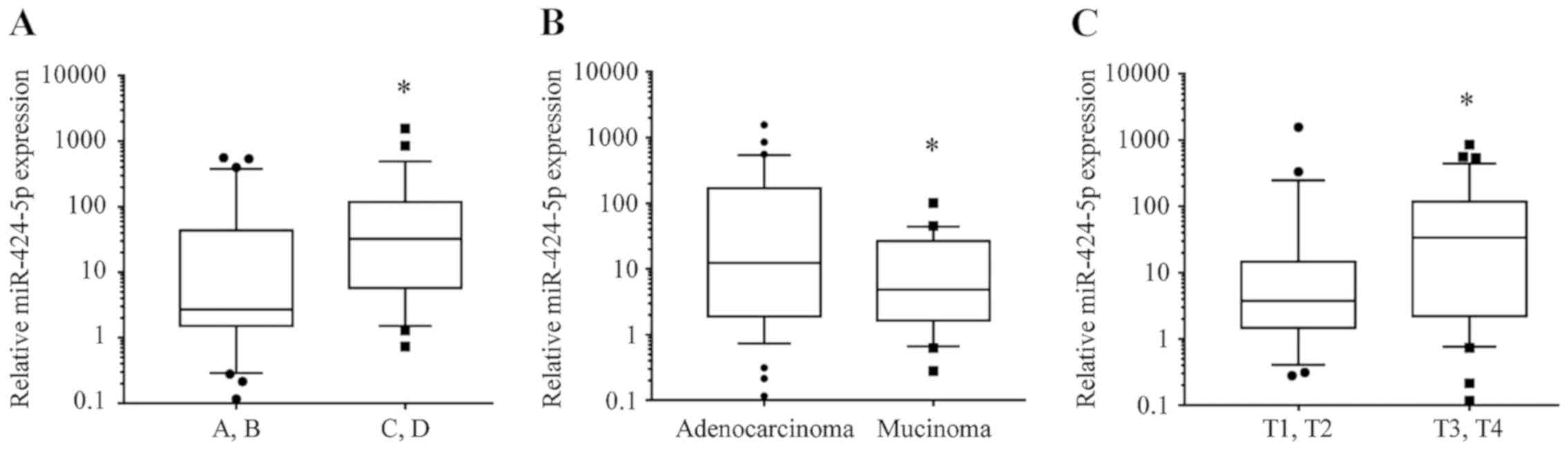
miR-424-5p expression is associated
with Dukes' stage, depth of invasion and pathological type of
CRC
The expression levels of miR-424-5p in CRC tissues
were significantly increased in Dukes' stage C.D compared with
those in Dukes' stage A.B [median (interquartile range), 32.45
(5.696-117.8) vs. 2.695 (1.518-43.65); P=0.0159; Fig. 2A]. The expression of miR-424-5p in
CRC tissues was also increased in adenocarcinoma compared with
mucinous carcinoma [12.38 (1.905-170.1) vs. 4.848 (1.641-26.72);
P=0.0451; Fig. 2B]. Regarding
infiltration depth, the expression levels of miR-424-5p in CRC
tissues were upregulated in T3.T4 stage [33.65 (2.191-117.4)]
compared with T1.T2 stage [3.77 (1.47-14.67); P=0.0450; Fig. 2C].
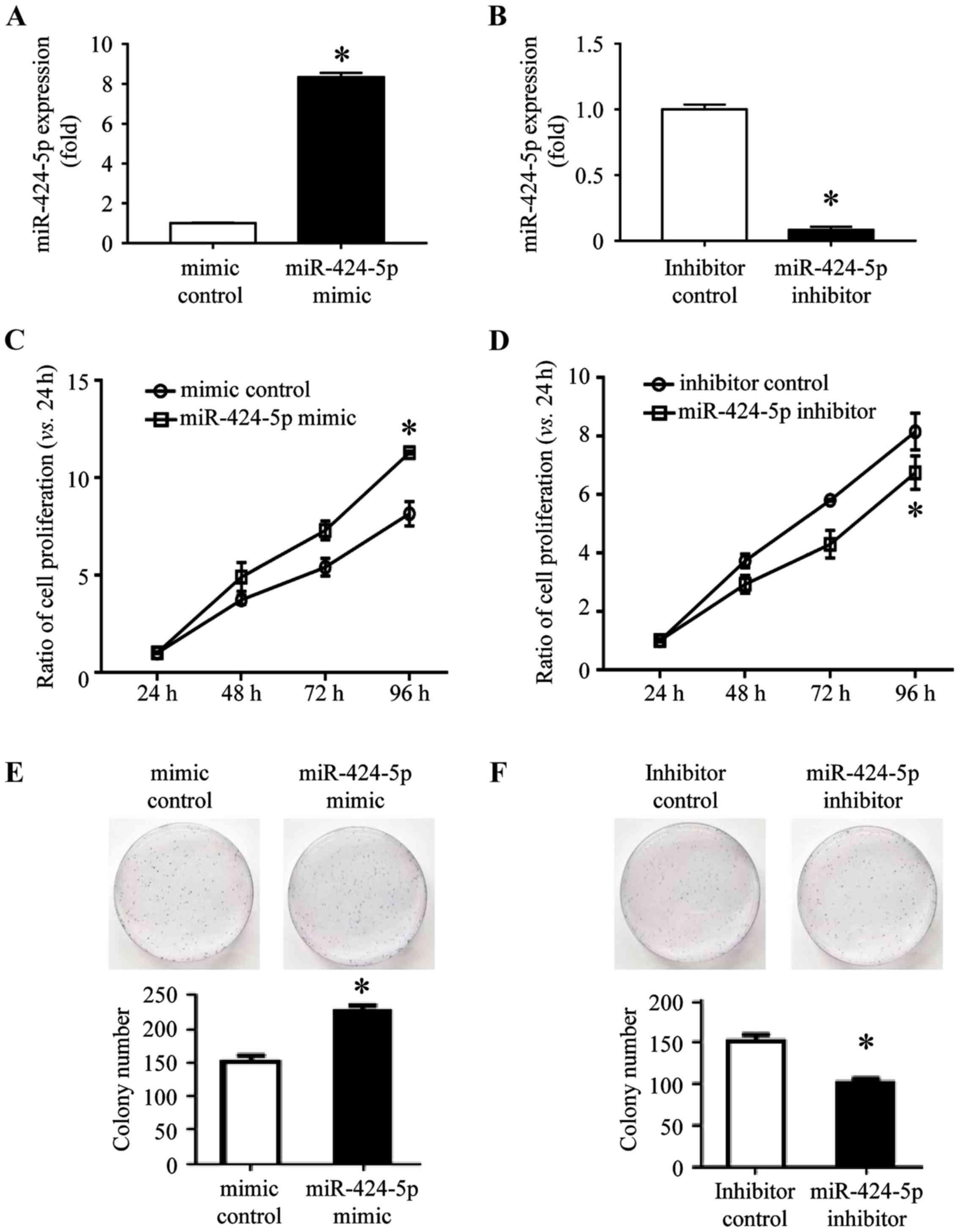
Overexpression of miR-424-5p promotes
and suppression of miR-424-5p inhibits CRC cell proliferation
To investigate the role of miR-424-5p in the
development of CRC, the cell viability was evaluated using a Cell
Counting Kit (CCK)-8 assay. The SW480 cells were successfully
transfected with miR-424-5p mimic (8.35±0.086) or mimic control
(1±0.015), and miR-424-5p inhibitor (0.083±0.009) or inhibitor
control (1±0.015; Fig. 3A and B).
Following transfection for 24 h, a CCK-8 assay was performed and
miR-424-5p expression was measured. The expression of miR-424-5p
and cell viability was increased following transfection of
miR-424-5p mimic as compared that in the mimic control-transfected
SW480 cells at 24, 48, 72 and 96 h (Fig.
3C). Following transfection with miR-424-5p inhibitor, the
expression of miR-424-5p and the viability of SW480 cells was
significantly decreased as compared with that in the inhibitor
control-transfected group at 48, 72 and 96 h (Fig. 3D). A colony formation assay was also
performed with SW480 cells transfected with miR-424-5p mimic or
mimic control and miR-424-5p inhibitor or inhibitor control. Colony
formation by SW480 cells transfected with miR-424-5p mimic was
significantly increased compared with that in the mimic control
group (Fig. 3E). In the miR-424-5p
inhibitor-transfected SW480 group, the colony formation was
significantly decreased as compared with that in the inhibitor
control group (Fig. 3F). These
results indicated that the miR-424-5p expression is positively
associated with CRC cell proliferation.
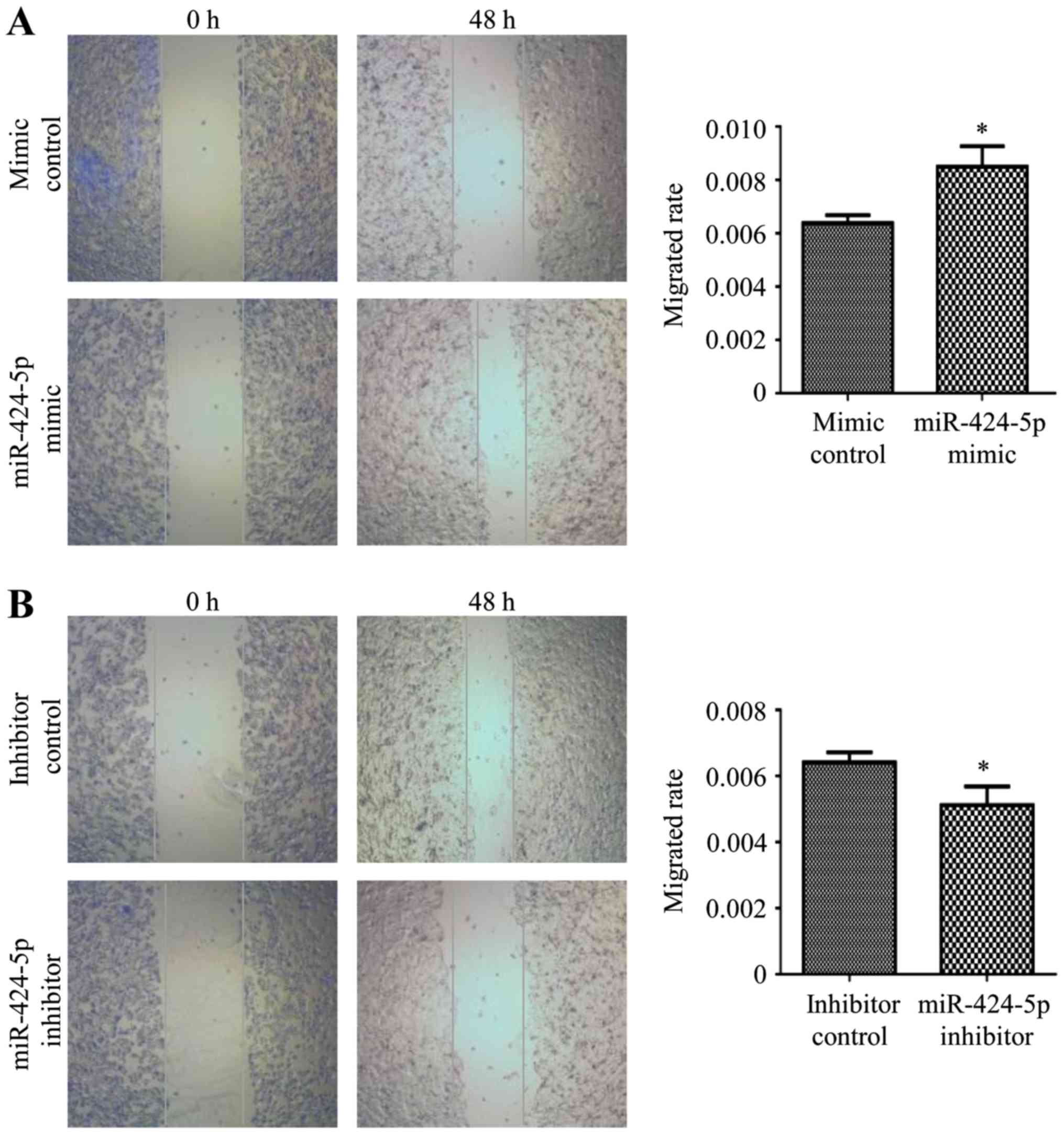
Overexpression of miR-424-5p promotes
and suppression of miR-424-5p inhibits the migration and invasion
of CRC cells
SW480 cells were transfected with miR-424-5p mimic
or mimic control and inhibitor or inhibitor control for 48 h and
then subjected to a wound-healing assay. The results revealed that
miR-424-5p overexpression promoted CRC cell migration, while
silencing of miR-424-5p markedly inhibited CRC cell migration
(P<0.05; Fig. 4A and B).
Subsequently, Transwell assays were used to examine the effects of
miR-424-5p on SW480 cell migration and invasion. Overexpression of
miR-424-5p significantly promoted, while silencing of miR-424-5p
significantly inhibited the migration of CRC cells (P<0.05;
Fig. 4C and D). Blue staining
represents migrating/invading cells. As presented in Fig. 4E, SW480 cells transfected with
miR-424-5p mimic exhibited a significantly higher invasive
capacity. However, those transfected with miR-424-5p inhibitor had
a significantly lower invasive capacity compared with those
transfected with the control (Fig.
4F). Therefore, miR-424-5p was indicated to have a stimulatory
effect on the migration and invasion of CRC cells.
Identification and functional
characterization of DEGs from the transcriptome sequencing of SW480
cells transfected with miR-424-5p mimic and control
Transcriptome sequencing revealed DEGs between SW480
cells transfected with miR-424-5p mimic and those transfected with
the control. These DEGs comprised 1,322 upregulated genes (Table SII) and 980 downregulated genes
(Table SIII), including
protein-coding genes and non-coding genes (Fig. 5A). The top 20 upregulated
protein-coding genes are presented in Table II and the top 20 downregulated
protein-coding genes are presented in Table III. GO analyses revealed that the
majority of the DEGs were significantly enriched in ‘serine
hydrolase activity’, ‘serine-type peptidase’ and ‘endopeptidase
activity’ (Fig. 5B). Disease
ontology functional enrichment analysis of these DEGs revealed that
they were accumulated in different diseases, including skin
disease, infertility and colonic diseases (Fig. 5C). Signaling pathway analysis
indicated that the DEGs were enriched in the ‘Wnt signaling
pathway’ (Fig. 5D).
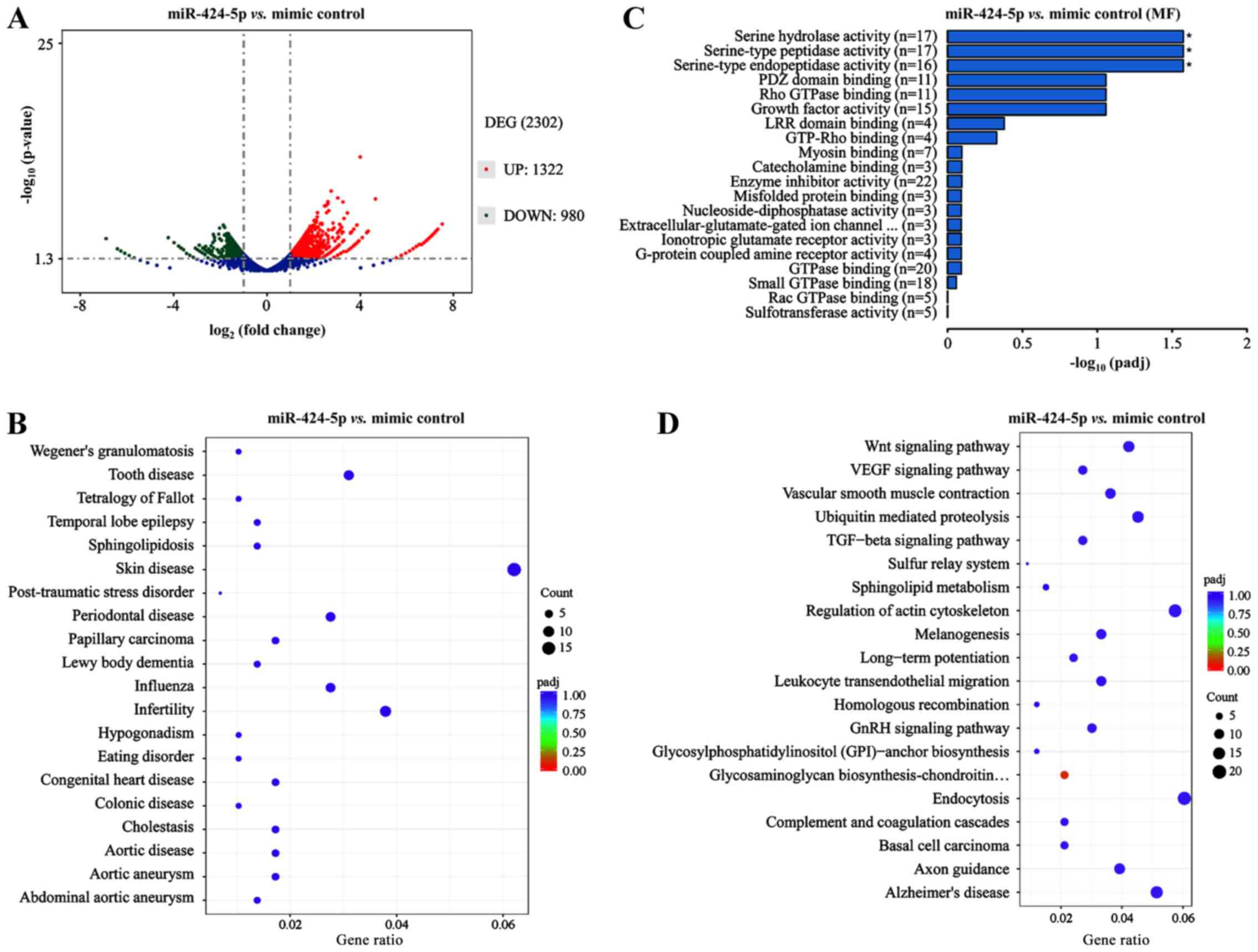 | Figure 5.Identification and functional
characterization of DEGs from the transcriptome sequencing in the
SW480 cells transfected with miR-424-5p mimic and mimic control.
(A) Volcano plot of the DEGs between the miR-424-5p mimic- and
mimic control-transfected SW480 cells from transcriptome
sequencing, including 1,322 upregulated genes (red dots, fold
change >2 and P<0.05) and 980 downregulated genes (green
dots, fold change ≤2 and P<0.05). (B) Gene ontology analyses
revealed that most of the DEGs were significantly enriched in
serine hydrolase activity, serine-type peptidase and endopeptidase
activity. (C) Disease ontology analysis of the DEGs. The DEGs were
enriched in different diseases, including skin disease, infertility
and colonic disease. The size of dots indicates the counts of DEGs.
(D) Signaling pathway analyses using the Kyoto Encyclopedia of
Genes and Genomes indicated that DEGs were enriched in signal
transduction MAPK signaling pathway. The size of the dots indicates
the counts of DEGs. miR, microRNA; DEG, differentially expressed
gene; VEGF, vascular endothelial growth factor; TGF, transforming
growth factor. |
| Table II.Top 20 upregulated protein-coding
differentially expressed genes (miR-424-5p mimic/mimic control,
P<0.05). |
Table II.
Top 20 upregulated protein-coding
differentially expressed genes (miR-424-5p mimic/mimic control,
P<0.05).
| Gene name | Log2
(fold-change) | Chromosome | Gene
description |
|---|
| SLC10A5 | 6.529 | 8 | Solute carrier
family 10 member 5 |
| C11orf42 | 6.404 | 11 | Chromosome 11 open
reading frame 42 |
| FGL2 | 6.404 | 7 | Fibrinogen-like
2 |
| B3GAT2 | 6.269 | 6 |
Beta-13-glucuronyltransferase 2 |
| PSG9 | 6.119 | 19 | Pregnancy-specific
beta-1-glycoprotein 9 |
| PKD2L2 | 5.951 | 5 | Polycystic kidney
disease 2-like 2 |
| GCKR | 5.951 | 2 | Glucokinase
(hexokinase 4) regulator |
| VWA5A | 5.951 | 11 | Von Willebrand
factor A domain containing 5A |
| FRRS1 | 5.951 | 1 | Ferric-chelate
reductase 1 |
| SENP3-EIF4A1 | 5.951 | 17 | SENP3-EIF4A1
readthrough (NMD candidate) |
| GREB1L | 5.762 | 18 | Growth regulation
by estrogen in breast cancer-like |
| COL23A1 | 5.762 | 5 | Collagen type XXIII
α 1 |
| MRAP | 5.762 | 21 | Melanocortin 2
receptor accessory protein |
| SLC22A9 | 5.762 | 11 | Solute carrier
family 22 (organic anion transporter) member 9 |
| CYP26C1 | 5.762 | 10 | Cytochrome P450
family 26 subfamily C member 1 |
| SLC46A2 | 5.762 | 9 | Solute carrier
family 46 member 2 |
| RP11-166B2.1 | 5.762 | 16 | Putative NPIP-like
protein LOC729978 |
| GPR182 | 5.544 | 12 | G protein-coupled
receptor 182 |
| PCDHGB5 | 5.544 | 5 | Protocadherin gamma
subfamily B 5 |
| TAL1 | 5.544 | 1 | T cell acute
lymphocytic leukemia 1 |
| Table III.Top 20 downregulated protein-coding
differentially expressed genes (miR-424-5p mimic/mimic control,
P<0.05). |
Table III.
Top 20 downregulated protein-coding
differentially expressed genes (miR-424-5p mimic/mimic control,
P<0.05).
| Gene name | Log2
(fold-change) | Chromosome | Gene
description |
|---|
| CGB5 | −6.41484 | 19 | Chorionic
gonadotropin beta polypeptide 5 |
| FAM209A | −6.26472 | 20 | Family with
sequence similarity 209 member A |
| ABI3 | −6.09713 | 17 | ABI family member
3 |
| SLC6A4 | −5.9075 | 17 | Solute carrier
family 6 member 4 |
| NANOS2 | −5.9075 | 19 | Nanos homolog 2
(Drosophila) |
| AGXT | −5.9075 | 2 | Alanine-glyoxylate
aminotransferase |
| CLEC17A | −5.9075 | 19 | C-type lectin
domain family 17 member A |
| IL1R2 | −5.68911 | 2 | Interleukin 1
receptor type II |
| SPATA3 | −5.68911 | 2 | Spermatogenesis
associated 3 |
| REG3A | −5.68911 | 2 | Regenerating family
member 3 α |
| PCED1B | −3.98055 | 12 | PC-esterase domain
containing 1B |
| OMP | −3.78942 | 11 | Olfactory marker
protein |
| TPO | −3.68344 | 2 | Thyroid
peroxidase |
| ERN2 | −3.30883 | 16 | Endoplasmic
reticulum to nucleus signaling 2 |
| BEND6 | −3.30883 | 6 | BEN domain
containing 6 |
| ADGRD1 | −3.15871 | 12 | Adhesion G
protein-coupled receptor D1 |
| GSTM3 | −3.06686 | 1 | Glutathione
S-transferase mu 3 (brain) |
| ADGRE1 | −3.06686 | 19 | Adhesion G
protein-coupled receptor E1 |
| USHBP1 | −2.97446 | 19 | Usher syndrome 1C
binding protein 1 |
| CCL3 | −2.76975 | 17 | Chemokine (C-C
motif) ligand 3 |
Nuclear factor of activated T cells 5, Wnt family
member 6 (WNT6), adenomatous polyposis coli, nuclear factor of
activated T cells 3, low-density lipoprotein receptor-related
protein 6, WNT2B, WNT11, dishevelled segment polarity protein 1
(DVL1), Rac family small GTPase 3 (RAC3), frizzled class receptor 9
(FZD9), secreted frizzled-related protein 5 (SFRP5), frizzled class
receptor 2 (FZD2), Rho-associated coiled-coil containing protein
kinase 1 (ROCK1) and ROCK2 were enriched in the Wnt signaling
pathway; therefore, the expression of these genes was analyzed by
RT-qPCR. The expression levels of WNT6, WNT11, DVL1, RAC3, FZD9,
SFRP5 and FZD2 were downregulated in miR-424-5p mimic-transfected
cells compared with the control cells (Table IV).
| Table IV.Expression of genes of the Wnt
signaling pathway. |
Table IV.
Expression of genes of the Wnt
signaling pathway.
| Gene name | Fold-change | P-value | Gene
description |
|---|
| WNT6 | 0.2845772 | 0.00002 | Wnt family member
6 |
| WNT11 | 0.43327208 | 0.00421 | Wnt family member
11 |
| DVL1 | 0.45380582 | 0.00614 | Dishevelled segment
polarity protein 1 |
| RAC3 | 0.45782965 | 0.00737 | Rac family small
GTPase 3 |
| FZD9 | 0.46584114 | 0.00921 | Frizzled class
receptor 9 |
| SFRP5 | 0.47657406 | 0.01022 | Secreted
frizzled-related protein 5 |
| FZD2 | 0.48708141 | 0.01351 | Frizzled class
receptor 2 |
Discussion
The expression of miR-424-5p has been reported to be
dysregulated in human cancer and may either be up- or downregulated
in tumor tissues depending on the tumor type (19). miR-424-5p was observed to be
upregulated in certain types of cancer, including esophageal
squamous cell carcinoma (15),
gastric cancer (13), pancreatic
cancer (12) and tongue squamous
cell carcinoma (20), while it was
downregulated in others, including epithelial ovarian cancer
(11), endometrial carcinoma
(21), cervical cancer (22), lung cancer (23), chronic myeloid leukemia (24), hepatocellular carcinoma (25) and bladder cancer (26). The present study indicated that
miR-424-5p was increased in CRC tissues and its association with
clinicopathological data were determined. The expression of
miR-424-5p was higher in advanced Dukes' stages of CRC (compared
with less advanced stages of CRC), deeper invasion depth of tumors
(compared with T1&T2 invasion depth of CRC) and adenocarcinoma
(compared with mucinous carcinoma). miR-424-5p expression was
previously reported to be associated with muscle invasion, high
pathological grade, lymph node involvement and distant metastasis
in bladder cancer (26). Various
studies have reported that the abnormal expression of miR-424-5p is
associated with different clinicopathological characteristics in
different tumor types. All of these studies suggested that
miR-424-5p has an important role in tumor progression.
A large number of studies have indicated that the
same miRNA may serve different biological roles in different types
of cancer cell by acting as either an oncogene or tumor suppressor.
miR-424-5p has been demonstrated to serve as a tumor suppressor in
most types of human cancer, including hepatocellular carcinoma
(27,28). miR-424-5p has been reported to
suppress tumor cell proliferation, migration and invasion through
downregulating the expression of c-Myb, catenin beta interacting
protein 1 (ICAT) and Akt3. miR-424-5p may function as a tumor
suppressor in endometrial carcinoma cells by targeting E2F
transcription factor (E2F)7 (21).
Similarly, miR-424-5p acts as an oncogene by targeting various
genes in cervical cancer (16,29) and
breast cancer (19). miR-424-5p has
been reported to be aberrantly expressed in several cancer types
and to act as a tumor suppressor when downregulated in these cancer
types. However, miR-424-5p promotes cell proliferation by targeting
Smad3 via the transforming growth factor-β signaling pathway in
gastric cancer (13). Various
studies have indicated that RNA exhibits different expression and
biological functions in different tumor tissue types. The present
study demonstrated that miR-424-5p promotes the migration and
invasion of the SW480 CRC cell line.
It is understood that miRNA leads to the alteration
of mRNA by binding to the 3′-UTR of the gene, leading to the
regulation of posttranscriptional gene expression by inhibiting the
translation or degradation of mRNA. However, the exact mechanisms,
including all of the target genes and targeting interactions, of
how miRNAs regulate their target genes remain to be fully
elucidated (30). miR-424-5p were
also able to supress thousands of target genes expression,
including the famous cancer related genes c-Myb, ICAT, Akt3 and
E2F3 etc., downregulating of those genes could suppress
hepatocellular carcinoma cell proliferation, migration and invasion
(19). miRNAs are involved in a
number of other oncogenic or carcinogenic networks, which leads to
the regulation of several mRNAs. For instance, RAS-responsive
element-binding protein (RREB1) and mutant KRAS repress miR-143-5p
and the miR-145-5p promoter, and KRAS and RREB1 are targets of
miR-145, revealing a feedback mechanism that increases the effect
of RAS signaling (31). By
performing a sequencing analysis and bioinformatics analysis of the
results, the present study demonstrated that miR-424-5p regulates
multiple genes. These genes are enriched in different signaling
pathways and located in different cell types. Signaling pathway
analysis indicated that the DEGs were enriched in ‘endocytosis’,
‘regulation of actin cytoskeleton’ and ‘Wnt signaling pathway’.
Of note, the present study had certain limitations.
For instance, only real-time PCR was used to detect the expression
of miR-424-5p and no in situ hybridization (ISH) was
employed. ISH is a complex technique and its advantage is that
paraffin sections may be used. This technology is frequently used
when paraffin sections are available (32). When total RNA is extracted from fresh
tissue, PCR is more convenient than that ISH to examine the
expression of miRNA. However, the intracellular position of
miR-424-5p expression cannot be indicated using RT-qPCR.
In conclusion, the present study demonstrated that
miR-424-5p, as a key candidate miRNA, was involved in the
development of CRC. In addition, miR-424-5p regulated gene
expression profiles and pathways of CRC, including the WNT6,
WNT11, DVL1, RAC3, FZD9, SFRP5 and FZD2 genes. It was
also demonstrated to a certain extent that the occurrence and
development of tumors is the result of the common action of
multiple genes. These candidate genes and pathways may be novel
therapeutic targets and biomarkers for CRC.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank Professor Xiao-feng
Sun (Division of Oncology, Department of Clinical and Experimental
Medicine, Faculty of Health Sciences, Linköping University,
Linköping, Sweden) for providing materials.
Funding
This study was supported by the National Natural
Science Foundation of China (grant no. 81572758), International
Science and Technology Cooperation Program of Hebei (grant nos.
2019YX006A and YZ201802), Natural Science Foundation of Hebei
(grant no. H2017206286), Foundation for Distinguished Young Talents
in Higher Education of Hebei (grant no. BJ2018042) and The Project
of Hebei Province Science and Technology Plan from Hebei province
(grant nos. zh2018002, LNB201911, 18077741D, XH201701 and
201805A049).
Availability of data and materials
miRNA array data are available from The Cancer
Genome Atlas repository (http://cancergennme.nih.gov/). The datasets used
and/or analyzed during the current study are available from the
corresponding author on reasonable request.
Authors' contributions
XJ, WFY and ZZ conceived and designed the
experiments. WTY, GW, WL, WFY, CZ and SC performed the experiments.
WTY, FS and WL collected and analyzed the data. WTY and XJ
interpreted the results and wrote the manuscript. All authors read
and approved the manuscript and agree to be accountable for all
aspects of the research and to guarantee for the accuracy and
integrity of any part of the work.
Ethics approval and consent to
participate
This study was performed in accordance with standard
guidelines and was approved by the Ethics Committee of The First
Hospital of Hebei Medical University (Shijiazhuang, China; approval
nos. 2013106 and 2016004). All patients provided written informed
consent prior to the study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Cao L, Liu Y, Wang D, Huang L, Li F, Liu
J, Zhang C, Shen Z, Gao Q, Yuan W and Zhang Y: MiR-760 suppresses
human colorectal cancer growth by targeting BATF3/AP-1/cyclinD1
signaling. J Exp Clin Cancer Res. 37:832018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Huang Q and Ma Q: MicroRNA-106a inhibits
cell proliferation and induces apoptosis in colorectal cancer
cells. Oncol Lett. 15:8941–8944. 2018.PubMed/NCBI
|
|
3
|
Gu S, Jin L, Zhang F, Sarnow P and Kay MA:
Biological basis for restriction of microRNA targets to the 3′
untranslated region in mammalian mRNAs. Nat Struct Mol Biol.
16:144–150. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lu J, Getz G, Miska EA, Alvarez-Saavedra
E, Lamb J, Peck D, Sweet-Cordero A, Ebert BL, Mak RH, Ferrando AA,
et al: MicroRNA expression profiles classify human cancers. Nature.
435:834–838. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ye JJ and Cao J: MicroRNAs in colorectal
cancer as markers and targets: Recent advances. World J
Gastroenterol. 20:4288–4299. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fayyad-Kazan H, Bitar N, Najar M, Lewalle
P, Fayyad-Kazan M, Badran R, Hamade E, Daher A, Hussein N, ElDirani
R, et al: Circulating miR-150 and miR-342 in plasma are novel
potential biomarkers for acute myeloid leukemia. J Transl Med.
11:312013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Friedman RC, Farh KK, Burge CB and Bartel
DP: Most mammalian mRNAs are conserved targets of microRNAs. Genome
Res. 19:92–105. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang B, Yin M, Cheng C, Jiang H, Jiang K,
Shen Z, Ye Y and Wang S: Decreased expression of miR4903p in
colorectal cancer predicts poor prognosis and promotes cell
proliferation and invasion by targeting RAB14. Int J Oncol.
53:1247–1256. 2018.PubMed/NCBI
|
|
9
|
Kassambara A, Jourdan M, Bruyer A, Robert
N, Pantesco V, Elemento O, Klein B and Moreaux J: Global miRNA
expression analysis identifies novel key regulators of plasma cell
differentiation and malignant plasma cell. Nucleic Acids Res.
45:5639–5652. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gozuacik D, Akkoc Y, Ozturk DG and Kocak
M: Autophagy-regulating microRNAs and Cancer. Front Oncol.
7:652017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu J, Gu Z, Tang Y, Hao J, Zhang C and
Yang X: Tumour-suppressive microRNA-424-5p directly targets CCNE1
as potential prognostic markers in epithelial ovarian cancer. Cell
Cycle. 17:309–318. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wu K, Hu G, He X, Zhou P, Li J, He B and
Sun W: MicroRNA-424-5p suppresses the expression of SOCS6 in
pancreatic cancer. Pathol Oncol Res. 19:739–748. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wei S, Li Q, Li Z, Wang L, Zhang L and Xu
Z: MiR-424-5p promotes proliferation of gastric cancer by targeting
Smad3 through TGF-β signaling pathway. Oncotarget. 7:75185–75196.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang Y, Li T, Guo P, Kang J, Wei Q, Jia
X, Zhao W, Huai W, Qiu Y, Sun L and Han L: MiR-424-5p reversed
epithelial-mesenchymal transition of anchorage-independent HCC
cells by directly targeting ICAT and suppressed HCC progression.
Sci Rep. 4:62482014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang F, Wang J, Yang X, Chen D and Wang L:
MiR-424-5p participates in esophageal squamous cell carcinoma
invasion and metastasis via SMAD7 pathway mediated EMT. Diagn
Pathol. 11:882016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhou Y, An Q, Guo RX, Qiao YH, Li LX,
Zhang XY and Zhao XL: MiR424-5p functions as an anti-oncogene in
cervical cancer cell growth by targeting KDM5B via the Notch
signaling pathway. Life Sci. 171:9–15. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lee JS: Exploring cancer genomic data from
the cancer genome atlas project. BMB Rep. 49:607–611. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang J, Wang S, Zhou J and Qian Q:
MiR-424-5p regulates cell proliferation, migration and invasion by
targeting doublecortin-like kinase 1 in basal-like breast cancer.
Biomed Pharmacother. 102:147–152. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li D, Liu K, Li Z, Wang J and Wang X:
MiR-19a and miR-424 target TGFBR3 to promote
epithelial-to-mesenchymal transition and migration of tongue
squamous cell carcinoma cells. Cell Adh Migr. 12:236–246. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li Q, Qiu XM, Li QH, Wang XY, Li L, Xu M,
Dong M and Xiao YB: MicroRNA-424 may function as a tumor suppressor
in endometrial carcinoma cells by targeting E2F7. Oncol Rep.
33:2354–2360. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xu J, Li Y, Wang F, Wang X, Cheng B, Ye F,
Xie X, Zhou C and Lu W: Suppressed miR-424 expression via
upregulation of target gene Chk1 contributes to the progression of
cervical cancer. Oncogene. 32:976–987. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang M, Zeng J, Zhao Z and Liu Z: Loss of
MiR-424-3p, not miR-424-5p, confers chemoresistance through
targeting YAP1 in non-small cell lung cancer. Mol Carcinog.
56:821–832. 2017. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hershkovitz-Rokah O, Modai S,
Pasmanik-Chor M, Toren A, Shomron N, Raanani P, Shpilberg O and
Granot G: Restoration of miR-424 suppresses BCR-ABL activity and
sensitizes CML cells to imatinib treatment. Cancer Lett.
360:245–256. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lu M, Kong X, Wang H, Huang G, Ye C and He
Z: A novel microRNAs expression signature for hepatocellular
carcinoma diagnosis and prognosis. Oncotarget. 8:8775–8784. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wu CT, Lin WY, Chang YH, Lin PY, Chen WC
and Chen MF: DNMT1-dependent suppression of microRNA424 regulates
tumor progression in human bladder cancer. Oncotarget.
6:24119–24131. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yang H, Zheng W, Shuai X, Chang RM, Yu L,
Fang F and Yang LY: MicroRNA-424 inhibits Akt3/E2F3 axis and tumor
growth in hepatocellular carcinoma. Oncotarget. 6:27736–27750.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yu L, Ding GF, He C, Sun L, Jiang Y and
Zhu L: MicroRNA-424 is down-regulated in hepatocellular carcinoma
and suppresses cell migration and invasion through c-Myb. PLoS One.
9:e916612014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang X, Li Q, Jin H, Zou H, Xia W, Dai N,
Dai XY, Wang D, Xu CX and Qing Y: MiR-424 acts as a tumor
radiosensitizer by targeting aprataxin in cervical cancer.
Oncotarget. 7:77508–77515. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kent OA, Fox-Talbot K and Halushka MK:
RREB1 repressed miR-143/145 modulates KRAS signaling through
downregulation of multiple targets. Oncogene. 32:2576–2585. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Resende TP, Marshall Lund L, Rossow S and
Vannucci FA: Next generation sequencing coupled with in situ
hybridization: A novel diagnostic platform to investigate swine
emerging pathogens and new variants of endemic viruses. Front Vet
Sci. 6:4032019. View Article : Google Scholar : PubMed/NCBI
|