Introduction
Globally, liver cancer was the sixth most commonly
diagnosed cancer and the fourth leading cause of cancer-associated
mortality in 2018 (1). Early tumor
recurrence and metastasis are the main causes of high mortality;
however, the signaling pathways that regulate these processes
remain unclear (2).
FXR was initially regarded as a multifunctional
metabolic regulator involved in maintaining the hepatic homeostasis
of bile acid, glucose and lipids (3,4). It has
been well documented that FXR also suppresses liver carcinogenesis
and progression (5–8). FXR may serve an indispensable role in
inhibiting the activity of signaling pathways that are dysregulated
in liver cancer cells (9).
The Wnt/β-catenin pathway serves a key role in liver
biology and liver cancer pathogenesis (10). Aberrant activation of Wnt/β-catenin
signaling has been observed in the majority of hepatocellular
carcinomas (HCCs) and in hepatoblastomas, and is closely associated
with HCC metastasis and prognosis (11,12). The
canonical Wnt/β-catenin pathway interacts with the
epithelial-mesenchymal transition (EMT) signaling network. The EMT
is considered one of the most important initial steps of cancer
invasion (13,14). Wolfe et al (15) demonstrated that the Wnt/β-catenin
pathway is activated in the liver of FXR-knockout mice during
spontaneous HCC development and that FXR deletion may result in the
loss of E-cadherin expression.
It has been previously reported that FXR
downregulation was correlated with the presence of multiple
malignant clinicopathological features, including tumor size,
advanced BCLC stage, poor differentiation and absence of
encapsulation, in patients with HCC (7). FXR inhibited liver cancer cell
proliferation in vivo and in vitro via inactivation
of the mammalian target of rapamycin (mTOR)/ribosomal S6 kinase
(S6K) pathway (9). The current study
aimed to investigate whether FXR influenced the regulation liver
cancer metastasis and the possible underlying mechanism.
Materials and methods
Cell culture
SK-Hep-1 cells were obtained from the cell bank of
the Chinese Academy of Sciences. Mycoplasma detection and cell line
authentication indicating no mycoplasma or other human cell line
contamination were performed using quantitative PCR detection for
mycoplasma and 10 short tandem repeat identification via capillary
electrophoresis for cell line authentication. SK-Hep-1 cells were
cultured in DMEM supplemented with 10% FBS (both Gibco; Thermo
Fisher Scientific, Inc.) and 1% penicillin-streptomycin (HyClone;
Cytiva) in a humidified atmosphere at 37°C with 5% CO2.
The FXR agonist GW4064 was obtained from Sigma-Aldrich (Merck
KGaA).
Construction of FXR recombinant
lentiviral vectors and transduction of the lentivirus into SK-Hep-1
cells
The construction of the human FXR overexpression
lentiviral vectors and negative controls, and transducing
lentivirus into SK-Hep-1 cells were performed as previously
described (7). Briefly, the human
FXR gene NR1H4 PCR-amplified from plasmid pCR4-FXR-TOPO (Open
BioSystems, Inc.) was cloned into the pGC-FU-GFP lentiviral vector
(Shanghai GeneChem Co., Ltd.). A total of 10 µg of the resulting
pGC-FU-FXR-GFP vector or the control empty vector pGC-FU-GFP was
co-transfected with the packaging vectors using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) into 293T cells to produce the lentiviruses that
were used to infect SK-Hep-1 cells. The cells were used for
subsequent experimentation 48 h post infection. SK-Hep-1 cells
transfected with recombinant vectors that overexpressed FXR or the
corresponding controls were designated as SK-Hep-1-FXR or
SK-Hep-1-NC, respectively.
RNA isolation and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was extracted from SK-Hep-1-FXR and
SK-Hep-1-NC cells using TriPure Isolation reagent (Roche Applied
Science) and converted to cDNA using a PrimeScript RT reagent kit
(Takara Bio, Inc.), according to the manufacturer's protocol.
RT-qPCR was performed using SYBR Premix Ex Taq (Takara Bio, Inc.)
and the Applied Biosystems 7500 Fast Real-Time PCR System (Applied
Biosystems; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol, with the following thermocycling
conditions: Denaturation at 95°C for 3 min, followed by 40 cycles
of denaturation at 95°C for 10 sec, annealing at 60°C for 30 sec
and elongation at 72°C for 30 sec, and a final extension step at
72°C for 30 sec. β-actin was used as the internal reference gene.
Relative mRNA levels to β-actin were analyzed using the
2−ΔΔCq method (16). The
paired forward and reverse primers for the amplification of the
specific genes were as follows: FXR forward,
5′-ATGCCTGTAACAAAGAAGCCCC-3′ and reverse,
5′-CACACAGTTGCCCCCGTTTTTAC-3′; low-density lipoprotein
receptor-related protein 5 (LRP5) forward,
5′-GTACCCGCCGATCCTGAAC-3′ and reverse, 5′-TGTAGTCGCTGTCACACACG-3′;
secreted Fzd-related protein (SFRP1) forward,
5′-CTCAACAAGAACTGCCACGC-3′ and reverse, 5′-CTCGTTGTCACAGGGAGGAC-3′;
matrix metalloproteinase 9 (MMP9) forward,
5′-TCTATGGTCCTCGCCCTGAA-3′ and reverse, 5′-CATCGTCCACCGGACTCAAA-3′;
cyclin D1 (CCND1) forward, 5′-CGGTGTCCTACTTCAAATGTGTGC-3′ and
reverse 5′-AGAGGCCACGAACATGCAAGTGG-3′; β-catenin (CTNNB1) forward,
5′-CTGAGGACAAGCCACAAGATTA-3′ and reverse,
5′-ATCCACCAGAGTGAAAAGAACG-3′; c-MYC forward,
5′-AGCTTGTACCTGCAGGATCTGAGC-3′ and reverse,
5′-AGCCTGCCTCTTTTCCACAGAAAC-3′; c-JUN forward,
5′-GTTGAGCTCGGGCTGGATAA-3′ and reverse, 5′-GACTATACTGCCGACCTGGC-3′;
Dishevelled 1 (DVL1) forward, 5′-TGAACCTCAACAGTGGCTCC-3′ and
reverse, 5′-GTGTGATCCGATTCACTGCC-3′; and Frizzled 5 (FZD5)
forward, 5′-TGTGCTTCATCTCCACGTCC-3′ and reverse,
5′-CCGTGGTCTCGTAGTGGATG-3′.
Microarray and bioinformatics
analysis
SK-Hep-1-FXR cells and SK-Hep-1-NC were pretreated
with 2 µM GW4064 for 24 h at 37°C, and total RNA from the cells was
isolated according to the aforementioned method and sent to
Kangchen BioTech Co., Ltd., for mRNA microarray analysis. The
samples were labeled and hybridized using a Whole Human Genome
Microarray kit (cat. no. 4×44K one-color v2; Agilent Technologies,
Inc.), according to the manufacturer's protocol. Raw data was
extracted using Agilent Feature Extraction software (v9.5.3;
Agilent Technologies, Inc.). Gene ontology (http://geneontology.org/) and Kyoto Encyclopedia of
Genes and Genomes databases (https://www.kegg.jp/) were used for bioinformatics
analysis. The microarray data were deposited to the Gene Expression
Omnibus (GEO) repository with the access no. GSE149111 (ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE149111).
Transwell migration and invasion
assays
The migratory and invasive capabilities of
SK-Hep-1-FXR and SK-Hep-1-NC cells were assessed using Transwell
BioCoat™ Matrigel™ Invasion Chambers (BD Biosciences), according to
the manufacturer's protocol. After pretreatment with the FXR
agonist GW4064 for 24 h at 37°C, 2×104 cells in 200 µl
of serum-free DMEM were loaded into the upper chamber of the
Transwell system and 750 µl DMEM containing 10% FBS were added to
the lower chamber. Following incubation for 24 h at 37°C, the cells
that did not migrate through the pores of the Transwell inserts in
the upper chamber were removed with a sterile cotton swab. Cells in
the lower chamber were fixed in 4% paraformaldehyde for 10 min at
room temperature and stained with hematoxylin for 3 min and eosin
for 15 sec at room temperature. Cells were then counted and imaged
in 10 randomly selected microscopic fields under an Olympus BX41
light microscope (Olympus Corporation) at a magnification of ×200.
Experiments were performed in replicate inserts and mean values
were calculated from 3 independent experiments. Inserts coated with
Matrigel at room temperature for 10 min were used in the invasion
assay to determine the invasive abilities of the cells.
Wound healing and scratch assays
After pretreatment with the FXR agonist GW4064 for
24 h at 37°C, a total of 2.5×105 SK-Hep-1-FXR and
SK-Hep-1-NC cells were seeded onto 6-well plates and scratched with
a sterile pipette tip to the confluent monolayer when cells reached
100% confluence. Following washing with PBS, serum-free DMEM was
added and cell migration was observed at 0 and 24 h. Images of
cells migrating across the scratched field were captured using an
Olympus fluorescence inverted microscope (Olympus Corporation) at
×100 magnification, and the number of cells that migrated into the
scratched area was counted (17) and
statistical analysis was performed using SPSS software (v22.0; IBM
Corp.) for 10 randomly selected fields within the scratched area
from 3 independent experiments.
Preparation of protein extracts and
western blotting
To prepare the total cellular proteins, SK-Hep-1-FXR
and SK-Hep-1-NC cell pellets were lysed in Cell Lysis Buffer
(Beyotime Institute of Biotechnology) with 1% phenylmethanesulfonyl
fluoride (PMSF; Beyotime Institute of Biotechnology). Nuclear
proteins were isolated using a Nuclear and Cytoplasmic Protein
Extraction kit (Beyotime Institute of Biotechnology), according to
the manufacturer's protocol. Briefly, cells were harvested and the
pellets were dissolved with cytoplasmic protein extraction agent A
(Beyotime Institute of Biotechnology) with PMSF. Cells were
vortexed at room temperature for 5 sec and incubated for 10–15 min
at 4°C on ice. Cytoplasmic protein extraction agent B (Beyotime
Institute of Biotechnology) was then added and the lysates were
centrifuged at 14,000 × g for 10 min at 4°C. The insoluble pellet
fraction was resuspended in nuclear protein extraction agent
supplemented with PMSF and the supernatants of the nuclear extract
were collected immediately. Total and nuclear proteins were
quantified using a bicinchoninic acid (BCA) protein assay (Beyotime
Institute of Biotechnology), aliquoted and stored at −80°C until
use. For western blotting, 30–70 µg of the proteins were separated
via 12% SDS-PAGE, transferred to the PDVF membranes (Beyotime
Institute of Biotechnology), blocked at room temperature with 5%
non-fat milk for 1 h. The members were incubated overnight at 4°C
with diluted mouse anti-human FXR monoclonal antibodies (1:500;
R&D Systems, Inc.; cat. no. PP-A9033A-00), mouse anti-human
monoclonal β-actin antibodies (1:2,500; Beyotime Institute of
Biotechnology; cat. no. AF0003), rabbit anti-human polyclonal
β-catenin (1:2,000; Abcam; cat. no. ab6302), rabbit anti-human
cyclin D1 monoclonal antibodies (1:1,000; Cell Signaling
Technology, Inc.; cat. no. 55506) and mouse anti-human Histone H3
monoclonal antibodies (1:1,000; Beyotime Institute of
Biotechnology; cat. no. AF0009). The membranes were washed with
TBS-Tween (0.1% Tween 20; Beyotime Institute of Biotechnology)
three times for 10 min each and incubated with HRP-labeled
secondary antibodies (goat anti-mouse, cat. no. 14709; or goat
anti-rabbit, cat. no. 14708; both 1:1,000 dilution; Cell Signaling
Technology, Inc.) at room temperature for 1 h. The bands were
detected using a BeyoECL Plus kit (Beyotime Institute of
Biotechnology) according to the manufacturer's protocol. Image Lab
software (version 5.0; Bio-Rad Laboratories, Inc.) was used for the
quantitative analysis. β-actin and Histone H3 were used as loading
controls for whole cell extracts and nuclear extracts,
respectively.
Animal studies
A total of 40 male BALB/c nu/nu mice (age, 4–6
weeks; weight, 16–18 g) were purchased from the Shanghai Laboratory
Animals Center and housed under controlled illumination (12 h light
and 12 h dark cycle), at a temperature of 22±2°C and humidity of
40–60% for 7 days with food and water available ad libitum.
To observe local invasion, 6 mice per group were injected
subcutaneously with 5×106 SK-Hep-1-FXR or SK-Hep-1-NC
cells resuspended in serum-free DMEM into the flank. Tumors were
palpable on day 7 post-transplantation. The mice were sacrificed on
day 21 post-inoculation by cervical dislocation after anesthesia
via intraperitoneal injection of a mixture of ketamine (100 mg/kg)
and xylazine (10 mg/kg). Tumors (1,210±120 mm3 for
SK-Hep-1-NC and 678±91 mm3 for SK-Hep-1-NC) and the
surrounding tissues deep into the muscle wall (~0.5 cm from the
tumor) were harvested and fixed in 10% neutral buffered formalin at
room temperature for 24 h. Sections of 3-µm thickness were then
stained at room temperature with hematoxylin for 5 min and eosin
for 1 min. Images were captured using an Olympus BX41 light
microscope (Olympus Corporation) at ×40 magnification.
For the in vivo lung metastasis study,
5×106 SK-Hep-1-FXR or SK-Hep-1-NC cells were injected
into mice through the caudal vein (n=4/group). Lungs were harvested
30 days later. The lung tissues were fixed in 10% neutral buffered
formalin at room temperature for 24 h, embedded into paraffin and
stained with hematoxylin and eosin as aforementioned.
Immunohistochemistry staining was performed in the lung tissues
containing metastatic foci. The histopathologically defined
metastatic foci were analyzed under a light microscope (Olympus
BX41; Olympus Corporation; magnification, ×100) after the lung
tissues were fixed and stained with hematoxylin and eosin, as
aforementioned. The number of the metastases with a maximum
diameter >0.2 mm was counted.
All experimental procedures involving the animals
were performed in accordance with the National Institutes of Health
Guide for the Care and Use of Animals (18) and were approved by the Institutional
Animal Care and Use Committee of Fujian Medical University (Fuzhou,
China).
Immunohistochemistry (IHC)
IHC staining was performed using an Envision Plus
System (Fuzhou Maixin Biotech Co., Ltd.) according to the
manufacturer's protocol. Briefly, the paraffin-embedded lung
tissues were cut into 3-µm-thick sections that were heated at 67°C
for 2 h. Sections were then deparaffinized twice with xylene for 5
min, rehydrated using a gradient ethanol series (100% for 5 min
twice, 95% for 5 min twice, 90% for 5 min, 85% for 5 min and 80%
for 5 min) and washed with double-distilled water for 1 h (all at
room temperature). Subsequently, sections were pretreated with 3%
H2O2 for 15 min at 4°C to block endogenous
peroxidase activity. Antigens were retrieved by pressure cooker
treatment at 120°C for 100 sec in 0.01 mmol/l citrate buffer (pH
6). To block non-specific antibody binding, sections were treated
with 10% goat serum (cat. no. ZLI-9021; OriGene Technologies, Inc.)
for 30 min at room temperature. The primary antibodies against
β-catenin (1:2,000; Abcam; cat. no. ab6302) and FXR (1:500; R&D
Systems, Inc.; PP-A9033A-00) were incubated with tissue sections
overnight at 4°C. The next day, tissue sections were incubated with
horseradish peroxidase-conjugated secondary antibodies (1:50;
Beyotime Institute of Technology; cat. no. AF0216) for 2 h at room
temperature. Images were captured using a fluorescence inverted
microscope (Olympus Corporation) at ×100 magnification. For
β-catenin, nuclear staining with or without cytoplasmic staining
were considered as positive. For FXR, cytoplasmic plus nuclear
staining was considered as positive. A 5-tiered scale was used to
assess the degree of FXR and β-catenin staining based on the
average percentage of positively stained cells (0, ≤5%; 1, 6–25%;
2, 26–50%; 3, 51–75%; 4, 76–100%) multiplied by the staining
intensity judged by two experienced pathologists [0, no staining;
1, weak staining (light yellow); 2, moderate staining
(yellow-brown); and 3, strong staining (brown)] to calculate a
final score ranging 0–12.
Statistical analysis
Statistical analysis was performed using SPSS
software (version 19; IBM Corp.). Values were expressed as the mean
± standard error of the mean of ≥3 independent experiments.
Unpaired Student's t-test or χ2 test was used to
determine the difference between groups. P<0.05 was considered
to indicate a statistically significant difference.
Results
FXR suppresses liver cancer cell
migration and invasion in vitro
To investigate the effect of FXR on liver cancer
cell migration and invasion in vitro, SK-HEP-1 cell line was
used as the cell model since SK-HEP-1 expresses FXR at very low
levels (7,9) and has a high metastatic capacity
(19). Therefore, the metastatic
suppressive effect of FXR could be determined when it is
overexpressed. SK-HEP-1 is an immortal human cell line derived from
the ascitic fluid of a patient with liver adenocarcinoma (20). Although these cells are of
endothelial origin and do not exhibit hepatocyte properties, they
have been generally considered as a human hepatoma cell line with
mesenchymal characteristics (19,20). The
isogenic paired FXR-overexpressing SK-HEP-1-FXR cell line was
established from SK-HEP-1 using a lentivirus expression system as
previously described (7). FXR
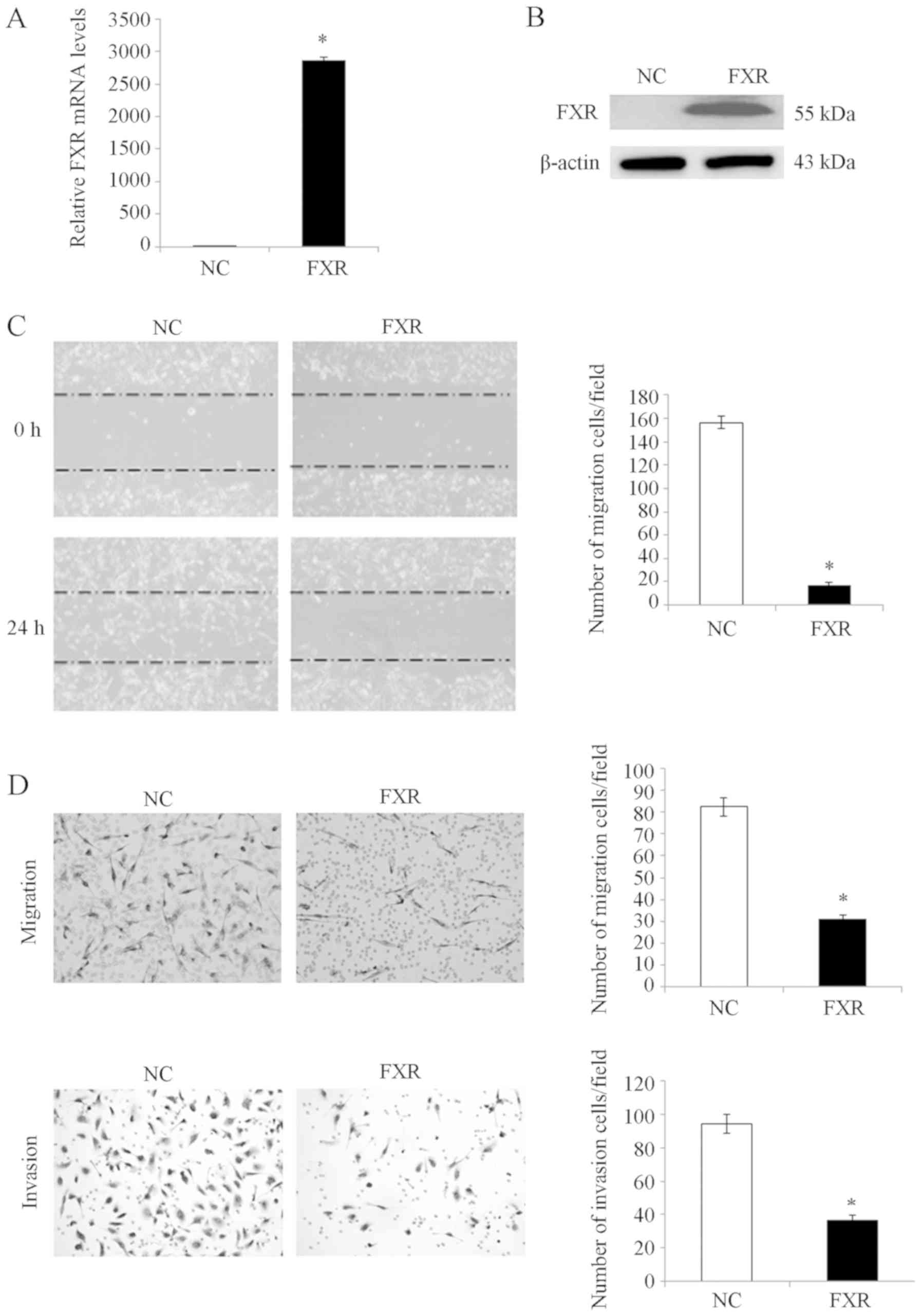
overexpression in SK-HEP-1-FXR cells was confirmed using RT-qPCR
(Fig. 1A) and western blotting
analysis (Fig. 1B). After
pretreating SK-Hep-1-FXR and control cells with the FXR agonist
GW4064 for 24 h at 37°C, wound healing and Transwell assays were
performed to determine cell migratory and invasive capacities,
respectively. SK-Hep-1-FXR cells demonstrated significantly
impaired wound healing ability (Fig.
1C; P<0.05) and significantly inhibited migration and
invasion abilities (Fig. 1D;
P<0.05), compared with controls. These data indicated that FXR
negatively regulated liver cancer cell migration and invasion in
vitro.
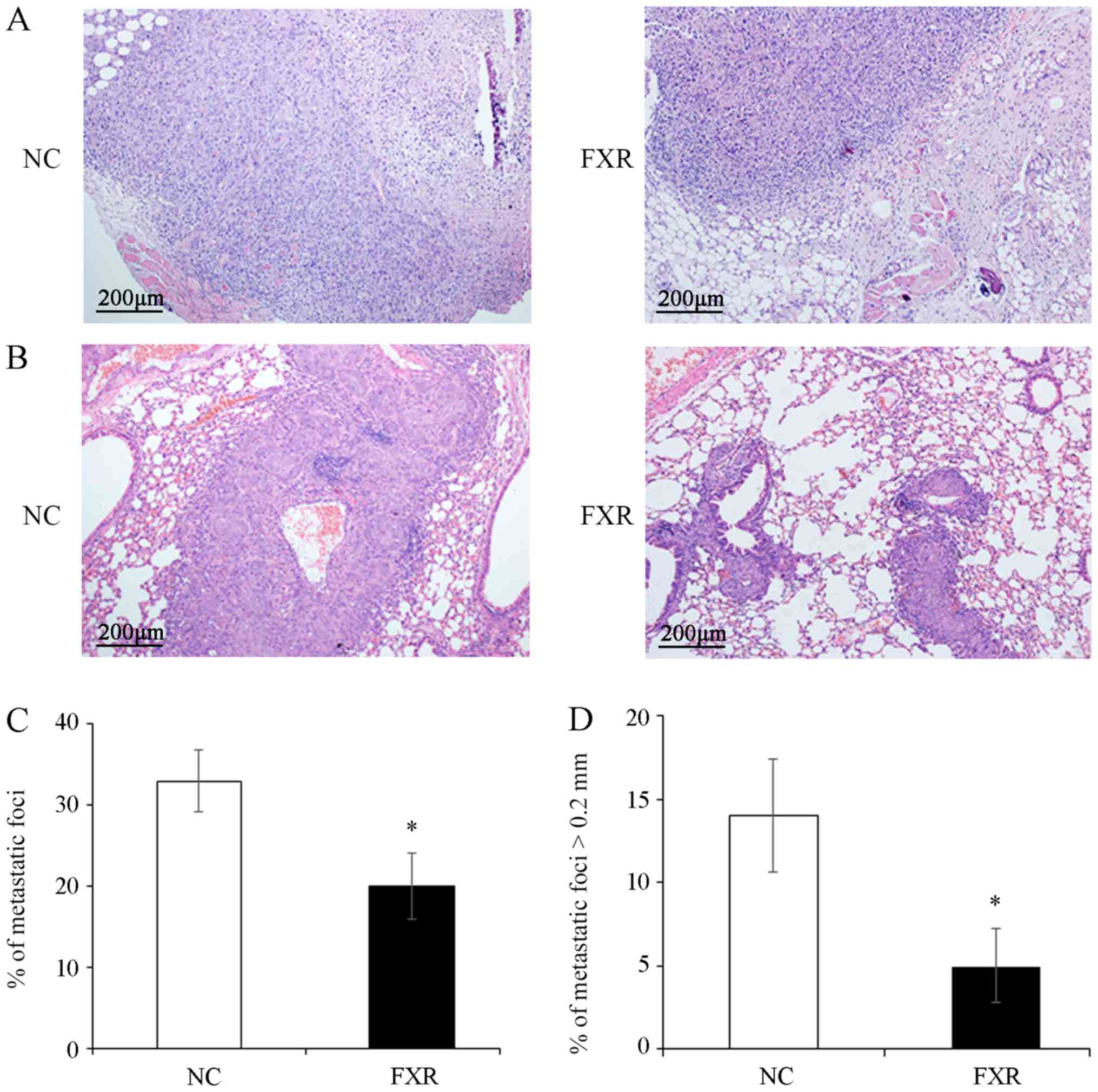
FXR inhibits local invasion and
metastasis of SK-Hep-1 ×enografts in vivo
The effect of FXR overexpression on the metastatic
potential in vivo was examined by subcutaneous injection of
the SK-Hep-1-FXR or SK-Hep-1-NC cells into nude mice to observe the
depth of local invasion or by tail vein injection to detect lung
metastasis. The results demonstrated that the invasion depth of
SK-Hep-1-FXR xenograft was limited to the subcutaneous loose
connective tissue layer (Fig. 2A,
right panel). By contrast, the tumors xenografted from controls
invaded deeply into the muscle layer (Fig. 2A, left panel). Furthermore,
significantly fewer and smaller pulmonary metastatic nodules, as
shown in the representative images of hematoxylin and eosin
staining on the lung tissue sections (Fig. 2B) and evidenced by the total number
of metastatic foci (Fig. 2C) and the
number of metastases with the maximum diameter >0.2 mm (Fig. 2D), were observed in the lungs of mice
injected with SK-Hep-1-FXR cells compared with controls (both
P<0.05). However, since most of lung metastatic foci are tiny
and not located on the surface of the lung (21), it is a technical challenge to measure
the volume or weight of such micro-metastases. These results
demonstrated that FXR inhibited local invasion and lung metastasis
of SK-Hep-1 ×enografts in vivo.
FXR may inhibit Wnt/β-catenin pathway
activation
To explore possible molecular mechanisms underlying
the metastatic suppressive effect of FXR, SK-Hep-1-FXR and
SK-Hep-1-NC cells were pretreated with the FXR agonist GW4064 and
microarray analysis was conducted to identify differentially
expressed genes. Gene ontology analysis demonstrated that
differentially expressed genes resulting from FXR overexpression in
SK-Hep-1-FXR cells were involved in diverse biological processes
including ‘cell migration’ and ‘cell proliferation’ (Fig. 3A). Pathway analysis demonstrated that
FXR upregulation in SK-Hep-1 cells altered the activity of
different cellular signaling pathways, including ‘cell cycle’,
‘mismatch repair’, ‘transforming growth factor β’ and ‘Wnt’ and
‘Janus kinase/signal transducers and activators of transcription’
(JAK-STAT; Fig. 3B). The Wnt pathway
was selected for further study due to its well-established role in
liver cancer cell proliferation and migration (22). The microarray data screened out
several differentially expressed genes of Wnt pathway including
FZD5, LRP5, sFRP1, DVL1 and CTNNB1. RT-qPCR was used to confirm
microarray data. The results demonstrated that FXR overexpression
downregulated Wnt/β-catenin upstream genes, including FZD5 and LRP5
(P<0.05; Fig. 3C). However, the
mRNA levels of sFRP and DVL were upregulated in SK-Hep-1-FXR cells,
while CTNNB1 expression was downregulated (P<0.05; Fig. 3C). sFRP is an antagonist of the Wnt
pathway which binds directly to Wnt ligands, preventing Wnt from
binding its receptors (23). DVL was
considered as a dual function adaptor and has been reported to be a
negative regulator of cell surface FZD level (24). Furthermore, β-catenin target genes
CCND1, c-MYC, c-JUN and MMP9 were downregulated in response to FXR
overexpression, as assessed via RT-qPCR (P<0.05; Fig. 3D) and western blotting (Fig. 3E). These results indicated that FXR
may function as a suppressor of the Wnt/β-catenin
pathway.
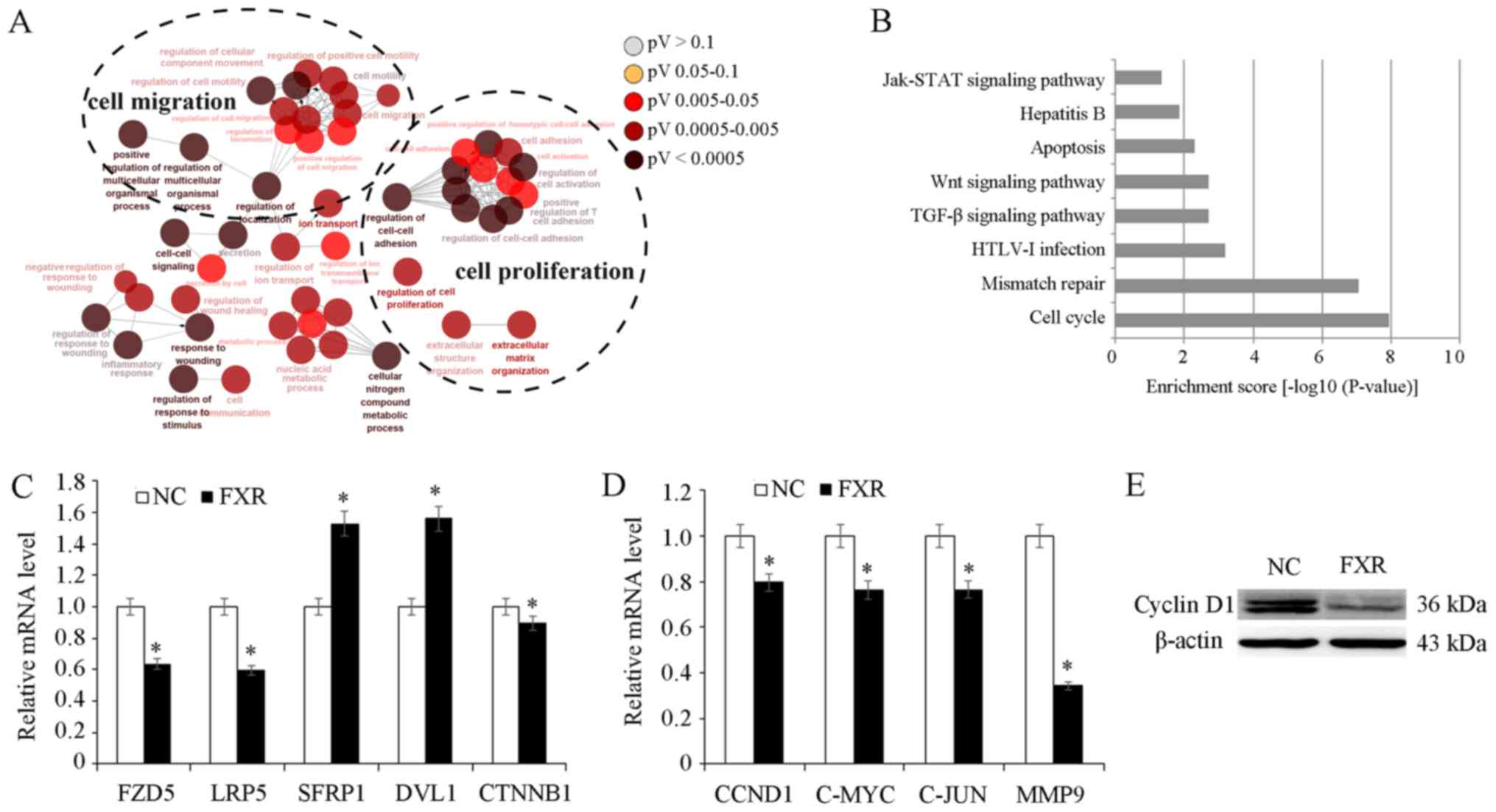 | Figure 3.Role of FXR in Wnt/β-catenin pathway.
SK-Hep-1-FXR and SK-Hep-1-NC cells were pretreated with GW4064 2 µM
for 24 h at 37°C, and harvested for microarray analysis. (A) Gene
ontology analysis demonstrated that the differentially expressed
genes resulting from FXR upregulation in SK-Hep-1-FXR cells were
involved in diverse biological processes. (B) Kyoto Encyclopedia of
Genes and Genomes pathway analysis revealed the alteration of
diverse cellular signaling pathways due to the FXR overexpression.
(C) mRNA expression levels of CTNNB1 and its upstream genes in
SK-Hep-1-FXR and SK-Hep-1-NC was detected via RT-qPCR. (D) CTNNB1
target genes were detected using RT-qPCR. (E) Cyclin D1 protein
expression was determined via western blotting. Results are
presented as mean ± standard error of the mean from three
independent experiments. *P<0.05 vs. NC. The ‘FXR’ group
represents FXR-overexpressing SK-Hep-1-FXR cells and the ‘NC’ group
represents the control SK-Hep-1-NC cells. FXR, farnesoid X
receptor; NC, negative control; RT-qPCR, reverse
transcription-quantitative PCR; Jak/STAT, Janus kinase/signal
transducers and activators of transcription; TGF-β, transforming
growth factor β; HTLV-1, human T-cell leukemia virus type 1; FZD5,
Frizzled 5; LRP5, low-density lipoprotein receptor-related protein
5; SFRP1, secreted Fzd-related protein; DVL1, Dishevelled 1; CCND1,
cyclin D1; CTNNB1, β-catenin. |
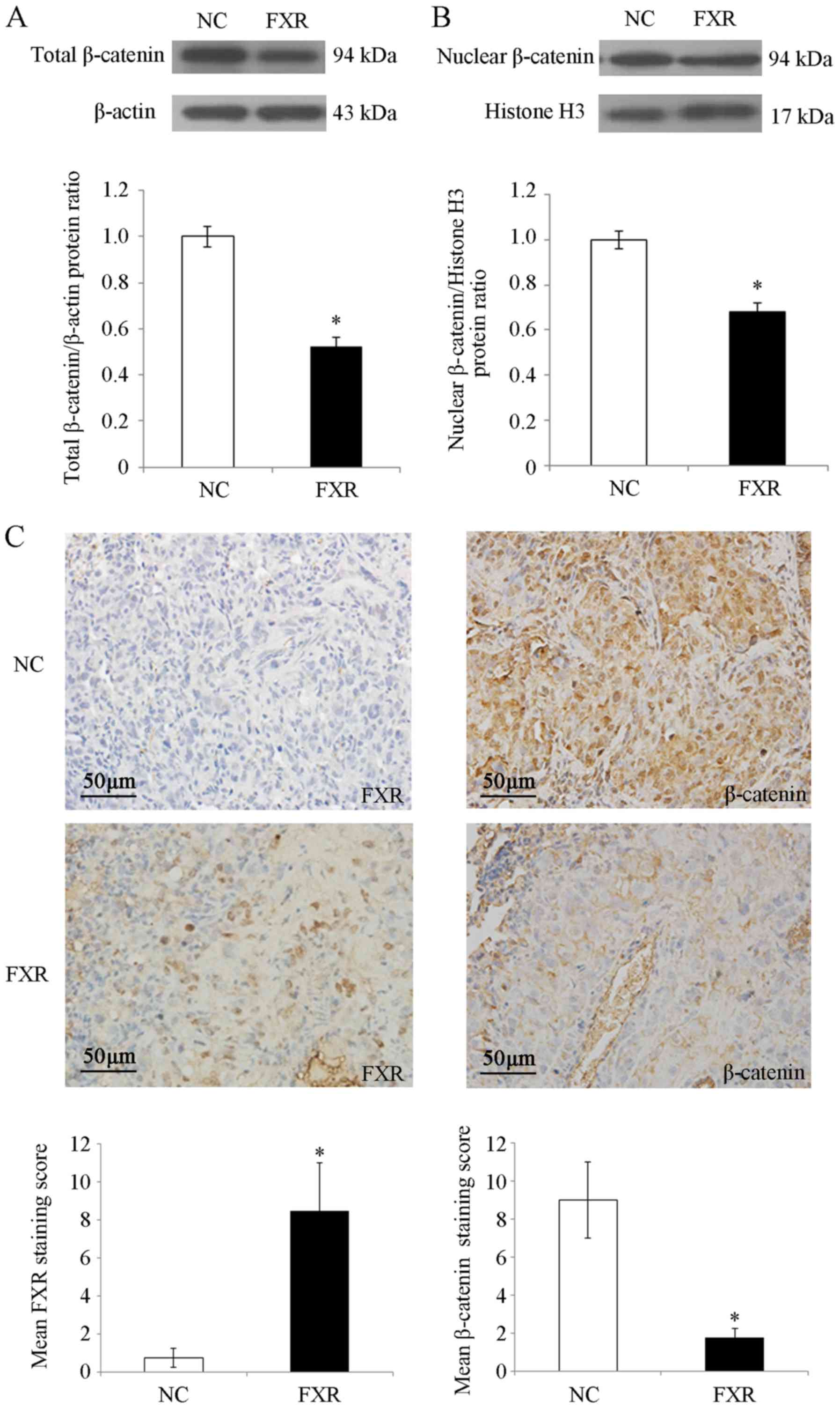
FXR blocks the nuclear translocation
of β-catenin in SK-Hep-1 cells in vitro and in vivo
β-catenin is the nuclear effector of the
Wnt/β-catenin pathway (25). To
investigate the possible mechanisms by which FXR inhibited
Wnt/β-catenin signaling, whether FXR blocked β-catenin nuclear
translocation in SK-Hep-1 cells was investigated. The results
demonstrated that total β-catenin protein levels were significantly
decreased by FXR overexpression (P<0.05; Fig. 4A), which was consistent with the
reduced CTNNB1 (the gene encoding β-catenin) mRNA levels in
SK-Hep-1-FXR cells (P<0.05; Fig.
3C). Furthermore, nuclear β-catenin levels were significantly
decreased in SK-Hep-1-FXR cells compared with controls (P<0.05;
Fig. 4B). As shown in Fig. 4C, IHC staining demonstrated strong
nuclear and moderate cytoplasmic staining for β-catenin protein in
the lung metastases of nude mice injected with SK-Hep-1-NC cells,
whereas β-catenin exhibited moderate membrane staining and weak
nuclear and cytoplasmic staining in the lung metastases of the mice
injected with SK-Hep-1-FXR cells (staining score, 9±2 for
SK-Hep-1-NC vs. 1.75±0.5 for SK-Hep-1-FXR; P<0.05). These
results indicated that when FXR was upregulated and activated,
total and nuclear β-catenin protein levels were concurrently
reduced in vitro and in vivo. FXR may inactivate the
Wnt/β-catenin pathway by blocking the nuclear translocation of
β-catenin, which may have been caused by the decreasing expression
or increasing degradation of β-catenin.
Discussion
Nuclear receptor FXR is a ligand-mediated
transcription factor (26). FXR was
first identified as an endogenous bile acid sensor in the liver
(27) and its role in protecting the
liver via metabolic regulation and tumor inhibition is well
established (5). However, the
molecular mechanisms by which FXR protects against liver cancer are
yet to be elucidated.
A previous study reported that FXR-knockout mice
spontaneously developed liver tumors as they aged (28). Moreover, FXR re-expression or
treatment with FXR agonists resulted in the repression of liver
cancer cell proliferation and xenograft growth in nude mice
(7,9). The results of the current study
indicated that, in addition to suppressing liver cancer cell
proliferation, FXR also inhibited migration and invasion in
vitro. Furthermore, the results demonstrated that FXR inhibited
local subcutaneous invasion and lung metastasis in the SK-Hep-1
×enograft models. Impaired cell migration and reduced cell growth
rate induced by FXR overexpression may contribute to the less
aggressive response in these cells. These results are consistent
with previous findings in human HCC specimens (7). Notably, whether other organs, in
addition to the lungs, had metastases in the in vivo mouse
model remains unclear. The site of injection largely defines where
metastases will develop. For instance, tail vein injections of
tumor cells results primarily in pulmonary metastases, while
intrasplenic or portal vein injections are the most common sites
employed to develop metastasis in the liver (29). Therefore, in the current study, the
lungs were the major site of metastasis development following tail
vein injections of SK-Hep-1-NC or SK-Hep-1-FXR cells; however, it
would be worthwhile to investigate the other organs in future
studies.
To further explore the possible molecular mechanism
by which FXR exerts its metastatic suppressor function in liver
cancer cells, gene expression profiles were analyzed in
SK-Hep-1-FXR and SK-Hep-1-NC cells. Pathway analysis on
differentially expressed genes revealed that FXR-upregulation in
SK-Hep-1 cells significantly enriched various cellular signaling
pathways, including ‘cell cycle’, ‘mismatch repair’, ‘TGF-β’,
‘JAK-STAT’ and ‘Wnt’. The role of FXR on the regulation of liver
cancer cell cycle has been previously studied and the results
demonstrated that FXR prevented the growth of liver cancer cells by
inhibiting the mTOR-s6K pathway (9).
The current study selected the Wnt/β-catenin pathway for further
investigation, as the dysregulation of this pathway is a key
molecular event in liver carcinogenesis and liver cancer
progression (30).
The canonical Wnt-β/catenin network controls various
biological processes in the liver. For example, when Wnt ligands
bind the FZD and LRP 5/6 coreceptor complex, β-catenin
phosphorylation and subsequent degradation is inhibited, resulting
in β-catenin accumulation in the cytoplasm and translocation into
the nucleus (21). In the nucleus,
β-catenin binds lymphoid enhancing factor/T cell factor to form a
transcriptional complex that promotes the transcription of its
target genes (31). In the current
study, mRNA microarray data revealed that FXR overexpression
resulted in significant changes of mRNA levels of certain core
components of this pathway, including downregulation of FZD5 and
LRP5, upregulation of sFRP and DVL, and significantly decreased
expression of β-catenin target genes (including CCND1, c-MYC, c-JUN
and MMP9). RT-qPCR and western blotting confirmed the results of
the microarray. These data support the hypothesis that FXR may
represent an inhibitor of the Wnt/β-catenin pathway. Furthermore,
the results demonstrated that FXR overexpression resulted in the
concurrent reduction of total and nuclear β-catenin proteins in
vitro and in vivo. Additionally, β-catenin mRNA levels
were downregulated in SK-Hep-1-FXR cells. It should be noted that a
pharmacological intervention on FXR expression to confirm these
results was not performed since there is not a specific and potent
FXR antagonist that is commercially available. Regardless, the
impaired nuclear translocation of β-catenin by FXR upregulation may
be caused by decreased expression or increased degradation of
β-catenin; however, this requires further research. In summary, the
results of the current study indicated that FXR inhibited the
Wnt/β-catenin pathway, which may lead to a reduction of the
migratory and invasive capabilities of Sk-Hep-1 cells.
Aberrant activation of the Wnt-β/catenin pathway has
been reported in a variety of liver diseases, such as liver
malignancies (hepatoblastoma, hepatocellular carcinoma and
cholangiocarcinoma) and non-tumor liver diseases (liver fibrosis,
liver steatosis, cholestasis and cystogenesis) (22). Increased nuclear β-catenin and c-MYC
expression has been revealed to be associated with vascular
invasion, which promotes HCC recurrence (11). Wnt/β-catenin signaling drives EMT,
which is considered the key initial event in metastasis (32). Therefore, further investigation into
the association between FXR, Wnt/β-catenin signaling and EMT is
required. In addition to the Wnt/β-catenin pathway, the JAK-STAT
pathway is a well-known oncogenic signaling pathway that promoted
the invasive ability of HCC cells by modulating the expression of
MMPs, including MMP-9 (10).
Additionally, MMP-9 is a target gene of β-catenin (33). The results of the current study
demonstrated that MMP9 mRNA was downregulated in response to FXR
overexpression in SK-Hep-1 cells, which indicated a potential
crosstalk between FXR, Wnt/β-catenin and JAK-STAT. Furthermore, the
TGF-β pathway maintains liver homeostasis and participates in
various liver diseases, including liver fibrosis and liver cancer
(34). TGF-β serves a dual role in
liver carcinogenesis, serving as a suppressor in early stages and a
contributor to later progression by enhancing the migratory and
invasive capacities of tumors (35).
Whether other pathways such as TGF-β, JAK-STAT and mismatch repair
also participate in FXR-regulated liver cancer cell invasion and
metastasis will be researched in future studies.
Tumor cell dissemination via invasion or metastasis
is the primary cause for the early recurrence of liver cancer in
patients following curative resection, and is also the main
obstacle to improving the prognosis of this disease (11). Due to the critical roles of FXR and
the Wnt/β-catenin pathway in liver cancer tumorigenesis and
progression, a more detailed understanding of the crosstalk between
these factors is required for the development of effective liver
cancer therapies.
Acknowledgements
Not applicable.
Funding
The current work was supported by the Natural
Science Foundation of Fujian Province (grant no. 2018J01827).
Availability of data and materials
Raw mRNA microarray data on the differentially
expressed genes resulting from FXR overexpression was deposited
into the Gene Expression Omnibus repository and available with the
access no. ‘GSE149111’ via the following link: https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE149111.
Authors' contributions
XH designed the study, interpreted data and wrote
the manuscript. QL, NL, JL, HS and MG performed the experiments,
analyzed the data and helped with the critical revision for
important intellectual content. YZ and XW analyzed the data and
prepared the figures. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The current study was approved by the Institutional
Animal Care and Use Committee of Fujian Medical University
(approval no. FJMUIRB-2011007).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
FXR
|
farnesoid X receptor
|
|
EMT
|
epithelial- mesenchymal transition
|
|
HCC
|
hepatocellular carcinoma
|
|
LRP
|
low-density lipoprotein
receptor-related protein
|
|
FZD
|
frizzled
|
|
sFRP
|
secreted FZD-related protein
|
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Shang N, Wang H, Bank T, Perera A, Joyce
C, Kuffel G, Zilliox MJ, Cotler SJ, Ding X, Dhanarajan A, et al:
Focal adhesion kinase and β-catenin cooperate to induce
hepatocellular carcinoma. Hepatology. 70:1631–1645. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Massafra V, Pellicciari R, Gioiello A and
van Mil SW: Progress and challenges of selective farnesoid X
receptor modulation. Pharmacol Ther. 191:162–177. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tran M, Liu Y, Huang W and Wang L: Nuclear
receptors and liver disease: Summary of the 2017 basic research
symposium. Hepatol Commun. 2:765–777. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Huang XF, Zhao WY and Huang WD: FXR and
liver carcinogenesis. Acta Pharmacol Sin. 36:37–43. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jiang Y, Iakova P, Jin J, Sullivan E,
Sharin V, Hong IH, Anakk S, Mayor A, Darlington G, Finegold M, et
al: Farnesoid X receptor inhibits gankyrin in mouse livers and
prevents development of liver cancer. Hepatology. 57:1098–1106.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Su H, Ma C, Liu J, Li N, Gao M, Huang A,
Wang X, Huang W and Huang X: Downregulation of nuclear receptor FXR
is associated with multiple malignant clinicopathological
characteristics in human hepatocellular carcinoma. Am J Physiol
Gastrointest Liver Physiol. 303:G1245–1253. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu N, Meng Z, Lou G, Zhou W, Wang X,
Zhang Y, Zhang L, Liu X, Yen Y, Lai L, et al: Hepatocarcinogenesis
in FXR−/− mice mimics human HCC progression that
operates through HNF1alpha regulation of FXR expression. Mol
Endocrinol. 26:775–785. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Huang X, Zeng Y, Wang X, Ma X, Li Q, Li N,
Su H and Huang W: FXR blocks the growth of liver cancer cells
through inhibiting mTOR-s6K pathway. Biochem Biophys Res Commun.
474:351–356. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Aravalli RN, Steer CJ and Cressman EN:
Molecular mechanisms of hepatocellular carcinoma. Hepatology.
48:2047–2063. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen J, Rajasekaran M, Xia H, Zhang X,
Kong SN, Sekar K, Seshachalam VP, Deivasigamani A, Goh BK, Ooi LL,
et al: The microtubule-associated protein PRC1 promotes early
recurrence of hepatocellular carcinoma in association with the
Wnt/β-catenin signalling pathway. Gut. 65:1522–1534. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dong LW, Yang GZ, Pan YF, Chen Y, Tan YX,
Dai RY, Ren YB, Fu J and Wang HY: The oncoprotein p28GANK
establishes a positive feedback loop in β-catenin signaling. Cell
Res. 21:1248–1261. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fu Y, Zheng S, An N, Athanasopoulos T,
Popplewell L, Liang A, Li K, Hu C and Zhu Y: β-catenin as a
potential key target for tumor suppression. Int J Cancer.
129:1541–1551. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Schmalhofer O, Brabletz S and Brabletz T:
E-cadherin, beta-catenin, and ZEB1 in malignant progression of
cancer. Cancer Metastasis Rev. 28:151–166. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wolfe A, Thomas A, Edwards G, Jaseja R,
Guo GL and Apte U: Increased activation of the Wnt/β-catenin
pathway in spontaneous hepatocellular carcinoma observed in
farnesoid X receptor knockout mice. J Pharmacol Exp Ther.
338:12–21. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Meng Z, Fu X, Chen X, Zeng S, Tian Y, Jove
R, Xu R and Huang W: miR-194 is a marker of hepatic epithelial
cells and suppresses metastasis of liver cancer cells in mice.
Hepatology. 52:2148–2157. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
National Research Council (US) Committee
for the Update of the Guide for the Care and Use of Laboratory
Animals, . The National Academies Collection: Reports Funded by
National Institutes of Health: Guide for the Care and Use of
Laboratory Animals. 8th. National Academies Press; Washington, DC:
2011
|
|
19
|
Eun JR, Jung YJ, Zhang Y, Zhang Y,
Tschudy-Seney B, Ramsamooj R, Wan YJ, Theise ND, Zern MA and Duan
Y: Hepatoma SK Hep-1 cells exhibit characteristics of oncogenic
mesenchymal stem cells with highly metastatic capacity. PLoS One.
9:e1107442014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Heffelfinger SC, Hawkins HH, Barrish J,
Taylor L and Darlington GJ: SK HEP-1: A human cell line of
endothelial origin. In Vitro Cell Dev Biol. 28A:136–142. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shang X, Lin X, Alvarez E, Manorek G and
Howell SB: Tight junction proteins claudin-3 and claudin-4 control
tumor growth and metastases. Neoplasia. 14:974–985. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Perugorria MJ, Olaizola P, Labiano I,
Esparza-Baquer A, Marzioni M, Marin JJG, Bujanda L and Banales JM:
Wnt-β-catenin signalling in liver development, health and disease.
Nat Rev Gastroenterol Hepatol. 16:121–136. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kawano Y and Kypta R: Secreted antagonists
of the Wnt signalling pathway. J Cell Sci. 116:2627–2634. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jiang X, Charlat O, Zamponi R, Yang Y and
Cong F: Dishevelled promotes Wnt receptor degradation through
recruitment of ZNRF3/RNF43 E3 ubiquitin ligases. Mol Cell.
58:522–533. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Clevers H and Nusse R: Wnt/β-catenin
signaling and disease. Cell. 149:1192–1205. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Koutsounas I, Giaginis C and Theocharis S:
Farnesoid X receptor (FXR) from normal to malignant state. Histol
Histopathol. 27:835–853. 2012.PubMed/NCBI
|
|
27
|
Makishima M, Okamoto AY, Repa JJ, Tu H,
Learned RM, Luk A, Hull MV, Lustig KD, Mangelsdorf DJ and Shan B:
Identification of a nuclear receptor for bile acids. Science.
284:1362–1365. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang F, Huang X, Yi T, Yen Y, Moore DD and
Huang W: Spontaneous development of liver tumors in the absence of
the bile acid receptor farnesoid X receptor. Cancer Res.
67:863–867. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Khanna C and Hunter K: Modeling metastasis
in vivo. Carcinogenesis. 26:513–523. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liang Y, Feng Y, Zong M, Wei XF, Lee J,
Feng Y, Li H, Yang GS, Wu ZJ, Fu XD and Feng GS: β-catenin
deficiency in hepatocytes aggravates hepatocarcinogenesis driven by
oncogenic β-catenin and MET. Hepatology. 67:1807–1822. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Macdonald BT, Semenov MV and He X:
SnapShot: Wnt/beta-catenin signaling. Cell. 131:12042007.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gonzalez DM and Medici D: Signaling
mechanisms of the epithelial-mesenchymal transition. Sci Signal.
7:re82014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yu W, Li L, Zheng F, Yang W, Zhao S, Tian
C, Yin W, Chen Y, Guo W, Zou L and Deng W: β-catenin cooperates
with CREB binding protein to promote the growth of tumor cells.
Cell Physiol Biochem. 44:467–478. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Fabregat I, Moreno-Càceres J, Sánchez A,
Dooley S, Dewidar B, Giannelli G and Ten Dijke P: TGF-β signalling
and liver disease. FEBS J. 283:2219–2232. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Fabregat I and Caballero-Díaz D:
Transforming growth factor-β-induced cell plasticity in liver
fibrosis and hepatocarcinogenesis. Front Oncol. 8:3572018.
View Article : Google Scholar : PubMed/NCBI
|