Introduction
Macrophages can directly engulf aberrant cells in
normal or non-tumor disease conditions and are therefore
conventionally regarded as anti-carcinogenesis (1). Due to their extremely plastic
phenotypes and highly dynamic functions, macrophages can be
transformed into different subtypes that have been reported to
differentially regulate tumor progression in the tumor
microenvironment (TME) (2).
According to the similarities in gene expression patterns and
functions between TAMs and traditional M1- and M2-type macrophages,
TAMs with mixed phenotypes are broadly divided into M1-like and
M2-like TAMs. M2 macrophages can be further subdivided into M2a,
M2b, M2c and M2d subtypes in response to different inducers in the
TME (3) (Fig. 1).
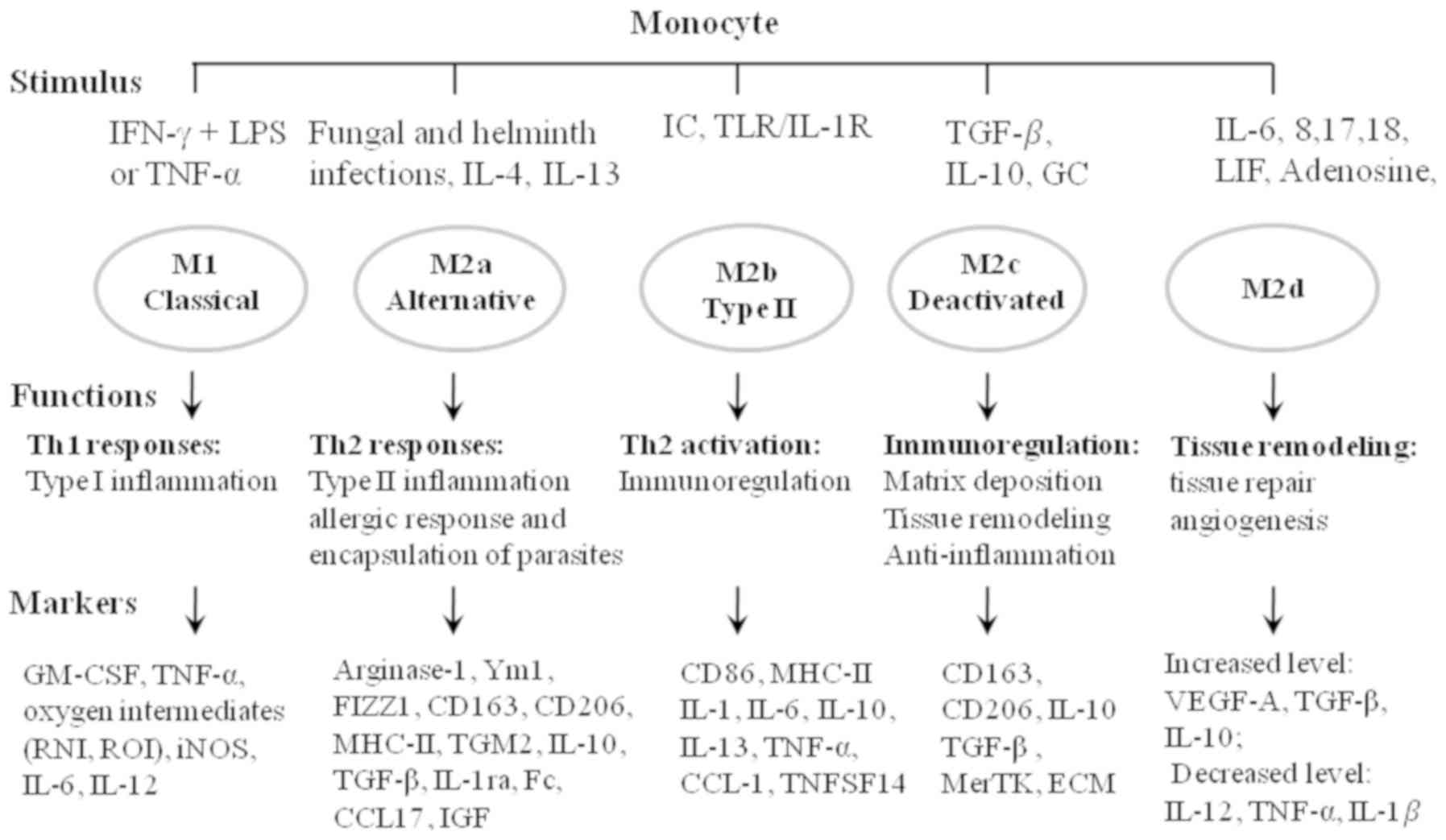 | Figure 1.Overview of macrophage activation.
Stimuli induce the polarization of different macrophage
sub-phenotypes with different functional properties. Markers can be
used to identify the distinct sub-phenotypes. IFN, interferon; LPS,
lipopolysaccharide; TNF, tumor necrosis factor; IL, interleukin;
TLR, toll-like receptor; TGF, tumor growth factor; LIF, leukemia
inhibitory factor; GM-CSF, granulocyte macrophage
colony-stimulating factor; RNI, reactive nitrogen intermediate;
ROI, reactive oxygen intermediate; iNOS, inducible nitric oxide
synthase; FIZZ1, found in inflammatory zone protein 1; MHC, major
histocompatibility complex; TGM2, transglutaminase 2; IGF,
insulin-like growth factor; ECM, extracellular matrix; VEGF,
vascular endothelial growth factor; IC, immune complex; TNFSF14,
TNF superfamily member 14; GC, glucocorticoid; MerTK, Mer receptor
tyrosine kinase. |
M2a macrophages, induced by interleukin (IL)-4 and
IL-13, are characterized by the increased expression of mannose
receptor CD206, decoy IL-1 receptor (IL-R) and C-C motif chemokine
ligand (CCL)17 (4). M2a macrophages
associated with wound healing can contribute to tissue repair by
secreting pro-fibrotic factors, such as transforming growth factor
(TGF)-β, insulin-like growth factor (IGF) and fibronectin (5) (Fig. 1).
M2b macrophages induced by immune complexes (IC) and toll-like
receptor (TLR)/IL-1R ligands are immune regulators (6). By inhibiting immune and inflammatory
responses in cancer and infectious diseases, M2b can promote tumor
development and parasite, bacterial and fungal infections (Fig. 1) (6).
In addition, M2b can attenuate spinal cord injury and early
reperfusion injury after myocardial ischemia in mice, and support
recovery from the injuries (7). M2c
induced by IL-10, TGF-β and glucocorticoids are acquired for the
deactivation of macrophages, and participate in anti-inflammation
responses, matrix deposition and tissue remodeling (8). The increased expression of the Mer
receptor tyrosine kinase on M2c enables more efficient uptake of
apoptotic cells compared with other macrophage subsets in an
anti-inflammatory and pro-tumor manner (Fig. 1). M2d induced by IL-6, 8, 17, 18,
leukemia inhibitory factor and adenosine are characterized by
increased expression of IL-10, TGF-β inducible nitric oxide
synthase (iNOS) and vascular endothelial growth factor (VEGF), and
decreased expression of IL-12, TNF-α and IL-1β (9). The increased levels of VEGF, IL-10 and
iNOS in M2d endow its abilities of tissue repair and angiogenesis
that benefit the metastasis of tumor cells (10) (Fig.
1).
M2-like TAMs are the major component of
tumor-infiltrating immune cells in the TME. The predominate
presence of M2-like TAMs in the TME is partially responsible for
immunosuppression in tumor growth. Individuals with a high M1 to M2
ratio are less susceptible to tumors (11). The ratio of M1-like and M2-like TAMs
in the TME is positively associated with the overall survival and
prognosis of patients with uveal melanomas (12). By inhibiting the secretion of M1-like
TAM-induced factor by epigenetically silencing aberrant DNA
methylation, gastric cancer cells evade tumor immune surveillance
during the transformation of benign cells into invasive cancer
cells (13). The transformation from
M2-like to M1-like TAMs under specific conditions results in the
recession of lung cancer (14).
Therefore, blocking the infiltration of macrophages, eliminating
the accumulation of M2-like TAMs, re-polarizing predominant M2-like
into M1-like TAMs or epigenetically silencing the secretion of
M2-like TAM-induced factors in the TME may be potential candidate
mechanisms for TAMs-targeted cancer immunotherapy. The present
review summarizes current knowledge regarding the recruitment of
macrophages, their polarization into M1-like or M2-like TAMs, and
their differential roles in angiogenesis, angiostasis, invasion,
metastasis and immune activity in the TME, which may offer valuable
insight into how to improve the design of TAMs-targeted cancer
immunotherapies. Some of these strategies show promising effects;
however, challenges still remain (15).
The recruitment of macrophages into the
TME
In terms of anatomy, macrophages can be divided into
tissue-resident and circulating monocytes-derived macrophages.
Tissue-resident macrophages are hypothesized to be the first being
reprogrammed by tumor cells into pro-tumoral M2-like TAMs.
Subsequently, monocyte-derived macrophages are recruited and
polarized into M2-like TAMs in the TME, which is critical for the
establishment of metastatic and malignant tumors (16). Chemoattractants and their receptors
that have been identified to recruit macrophages into the TME
include: CCL2/CC receptor (R)2+, CCL2/CCR5+,
IL-1β/IL-1R, VEGFA/VEGFR, colony-stimulating factor (CSF)1/CSFR and
tyrosine-protein kinase receptor (Tie)/Angiogenin 2 (ANG2)
(17). For example, CCL2 expressed
by tumor cells and macrophages promotes the recruitment of
CCR2+ monocytes and CCR5+ granulocytes into
the TME, promoting tumor growth and metastasis (18). Among the chemoattractants CCL5, C-X-C
motif chemokine ligand (CXCL) 10, CXCL12 and complement C1q, the
latter is the most potent attractant promoting M2-like TAMs
recruitment (19).
Signaling pathways that function downstream of
chemoattractant-receptor pairs for recruiting macrophages into TME
include the TGF-β, PI3Kγ, TLR and mTOR pathways. TGF-β mediates the
recruitment of macrophages that compete with dendritic cells (DCs),
and decreases the antigen-presenting ability of DCs in the adaptive
immune system in skin cancer, which result in the transformation of
a regressing tumor into a progressing tumor (20). Periostin (POSTN), secreted by ovarian
cancer cells, recruits macrophages into the tumor tissue, where
macrophages further increase the expression of POSTN in ovarian
cancer cells through the production of TGF-β. In turn, the
increased expression of POSTN facilitates the recruitment of
macrophages into the TME (21).
Activation of the CSF1/CSF1R signaling axis enhances epidermal
growth factor (EGF) expression, and promotes the recruitment of
macrophages and the migration of epithelial cells in tumor tissues.
Conversely, the inhibition of CSF1 receptor signaling abolishes TAM
infiltration, enhances the recruitment of CD8+ T cells
and reduces the growth of cervical and breast tumors (22).
The tropism of macrophages to hypoxia drives the
migration and infiltration of macrophages into the hypoxic tumor
compartments (23). This migration
is halted by hypoxia-inducible transcription factor (HIF)-1α, when
macrophages arrive in tumor compartments, where macrophages are
polarized into hypoxic TAMs in order to promote tumor growth and
metastasis (23). The pro-tumor
function of hypoxic TAMs was confirmed by macrophage-specific
genetic deletion of neuropilin (Nrp)-1, a binding partner of
hypoxia-induced TAM attractant semaphorin 3A (Sema3A). The deletion
of Nrp-1 impedes macrophage entry into hypoxic tumor compartments
by blocking the Sema3A/Nrp1 signaling cascade, inhibiting
angiogenesis and restoring antitumor activity (24). Hypoxia stimulates the production of
VEGF and induces skin carcinogenesis through the recruitment and
alternative activation of macrophages. Hypoxia-associated
chemoattractant endothelial monocyte-activating polypeptide (EMAP)
2 also stimulates macrophage recruitment under hypoxic conditions
(25). In hypoxic melanoma cells,
HIF-1α induces the translocation and secretion of high-mobility
group box-1 (HMGB1) that increases IL-10 production and M2-like TAM
activation (26). HIF-1α increases
forkhead box protein M1 expression and mediates hypoxia-inducible
epithelial-mesenchymal transition (EMT) in prostate cancer
(27). In addition, extracellular
matrix (ECM) components, integrins, and immunoglobulins in the TME
can promote the infiltration of monocytes and macrophages. For
example, p110γ activated by tumor-derived chemoattractants, such as
receptor tyrosine kinases (RTKs), TLR/IL1Rs or G protein-coupled
receptors (GRCRs), selectively promotes the infiltration of myeloid
cells into the TME (28). Matrix
metalloproteinases (MMPs) can degrade the ECM and regulate
signaling pathways that control cell growth, inflammation and
angiogenesis, and can even work in a non-proteolysis manner
(29). Table I summarizes the cytokines,
chemokines, growth factors, metabolites, integrins, immunoglobulins
and selectins that are associated with M2-like TAMs recruitment in
the TME.
 | Table I.Cytokines, chemokines and metabolites
identified to be associated with M2-like TAMs recruitment in the
TME. |
Table I.
Cytokines, chemokines and metabolites
identified to be associated with M2-like TAMs recruitment in the
TME.
| Cytokines | Chemokines | Metabolites | Cells |
|---|
| FGF-2,VEGF, EMAP2,
CSF1, PDGF, IL-25, TGF-β, HIF-1α, CSF2, IL-4, IL-6, IL-10, TNF-α,
NF-κB, EGF, P2Y2, HMGB1, Fizz1, ADM, SEMA3A, TLR-4, MMPs, STAT3 and
STAT6, PG, serine protease, COX2, cathepsins B and S, endothelin,
oncostatin M, eotaxin, C1q, arginase | CCL2, CCL3, CCL5,
CCL4, CCL22, CCL17, CCL18, CCL20, CCR5, CX3CL1, CX3CL5, CX3CL6,
CXCL8, CX3CL8, CX3CL, CX3CR1 CX3CL16, CXCL12 | High lactate, low
pH, succinate, LPC, S1P, high kynurenine, lipid accumulation,
ribosomal protein S19, and nucleotides such as ATP and UTP | Apoptotic tumor
cells, adipocyte, tumor cell, endothelial cell, fibroblasts and
cancer stem cells |
Apart from the recruitment of macrophages mediated
by signaling molecules, the interactions between TAMs and other
cells, such as apoptotic and non-apoptotic tumor cells, adipocytes,
endothelial cell (ECs) and fibroblasts, also recruit macrophages
into the TME (Table I). Through the
CCL2/IL-1β/CXCL12 signaling pathway, adipocytes recruit and
activate macrophages to promote stromal vascularization and
angiogenesis before the formation of a tumor (30). Adipocyte fatty acid-binding protein
(AFABP) secreted by TAMs and adipocytes facilitates pro-tumor
IL-6/STAT3 signaling through the NFκB/microRNA (miR)-29b pathway in
TAMs with the CD11b+F4/80+ major
histocompatibility (MHC)II-lymphocyte antigen 6 complex phenotype
(31). The number of TAMs settled in
the vicinity of cancer-associated fibroblasts (CAFs) is
significantly correlated with cancer clinical stage. The
interactions between TAMs and CAFs promote the recruitment and
activation of each other, contributing to neuroblastoma progression
(32). Similarly, increased CXCL1
levels in urothelial cancer enhances the recruitment of TAMs and
CAFs, the metastasis of cancer cells, and predicts poor prognosis
(33). TAMs produce IL-6 and signal
via STAT3 to promote the expansion of human hepatocellular cancer
stem cells (CSCs) (34). The
secretion of CSC-derived chemoattractants, such as CCL2, CCL5 and
VEGF-A, are much higher in glioma tissue than in normal tissue,
which facilitates macrophage recruitment and may participate in the
architecture of the glioma-initiating cell niche (35).
The polarization of macrophages in the
TME
Macrophages are highly plastic and have dynamic
phenotypes and functions. Depending on the induction signals,
including hypoxia, malignant cell- or infiltrating T cell-derived
cytokines, chemokines, metabolites and enzymes in the TME,
macrophages are polarized into M1-like or M2-like TAMs that can be
further categorized by Th1/Th2 lymphocyte polarization during
inflammation (36). M1-like TAMs
associated with Th1 are induced by interferon (IFN)-γ, CSF2, TNF-α,
oxygen intermediates released by Th1 immune responses, and possess
pro-inflammatory and cytotoxic antitumor abilities (Fig. 2) (37). Increased iNOS and pro-inflammatory
factors, such as IL-6, IL-12 and IL-4, in M1-like TAMs are
associated with a positive prognosis in patients with non-small
cell lung cancer (38). Conversely,
M2-like TAMs are induced by anti-inflammatory molecules, such as
arginase 1 (ARG1), chitinase 3 like 1 (Ym1), interferon regulatory
factor (IRF)4, peroxisome proliferator-activated receptor (PPAR)γ
or cyclic AMP-responsive element-binding protein (CREB), released
by Th2 immune responses. This is accompanied by increased numbers
of CD4+ T cells and a poor prognosis in patients with
breast cancer (39). TGF-β and IL10
produced by Th2 lymphocytes promote the Th2 response but inhibit
Th1 activity, whilst IFN-γ and IL-4 produced by Th1 lymphocytes
promote the Th1 response but inhibit Th2 activity (40) (Fig.
2). In addition, pro-inflammatory molecules, such as IFNγ,
TLR4, IL12 and NOS that are secreted by the activation of the
STAT1, IRF5, NF-κB or AP1 signaling pathways, promote the Th1
response and the polarization of M1-like TAMs (40). Β-Glucan (a dectin-1 ligand) was also
found to promote M1 polarization via the NFκB/autophagy pathway
(41). The reciprocal regulation
between M1-like and M2-like TAMs are implemented by STAT1/STAT6,
IRF5/IRF4, NF-κB/PPARγ, AP1/CREB, and AP1/PPARγ signaling axes
(40), which are essential for the
initiation, development, and cessation of tumor inflammation, and
may be potential targets for modulating the transformation of
M1-like and M2-like TAMs in clinical cancer immunotherapies
(Fig. 2). However, the importance of
these signaling axes in re-polarizing M2-like to M1-like TAMs
remains to be assessed in TAMs-targeted cancer immunotherapy.
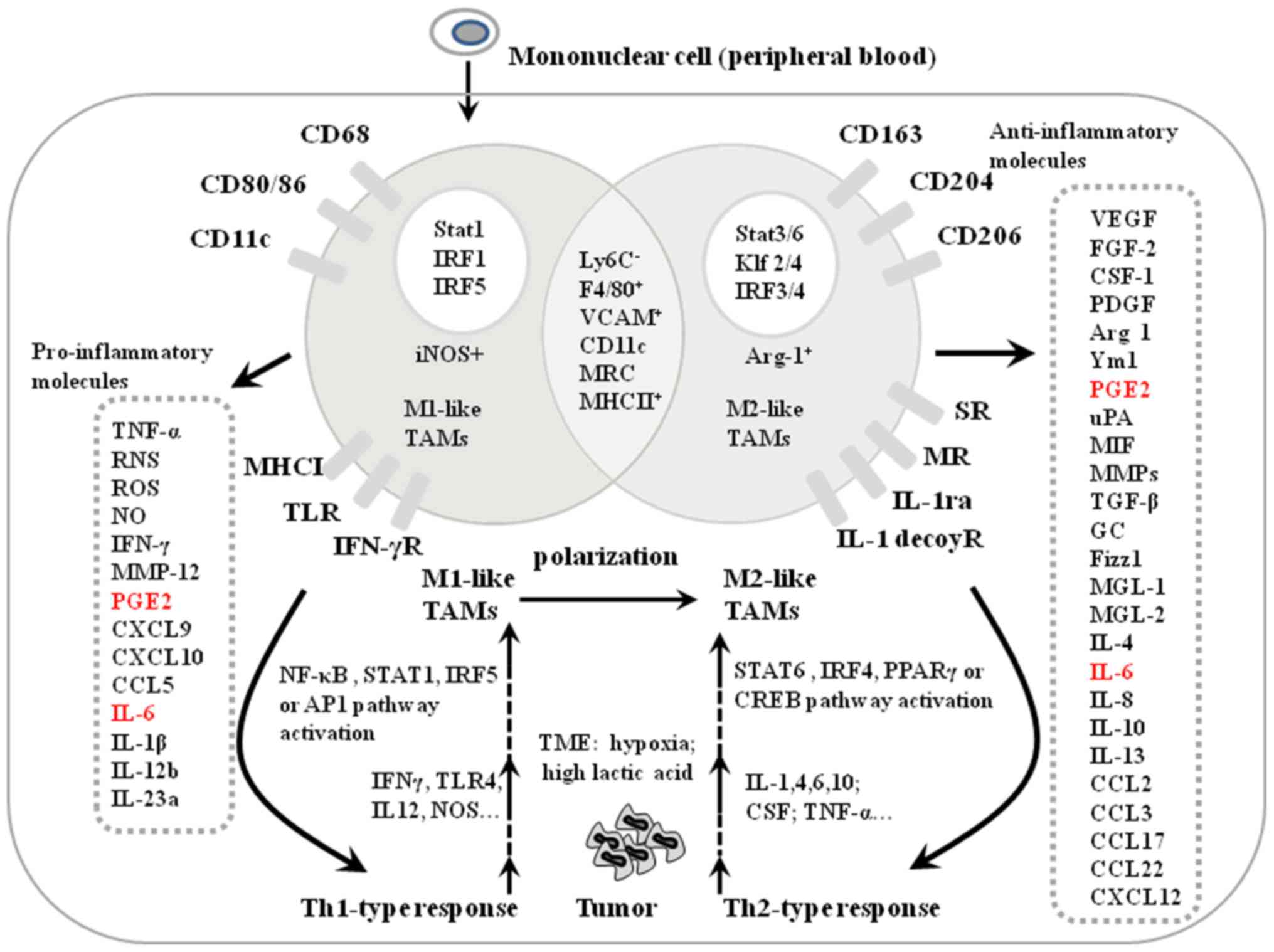 | Figure 2.Markers and polarization of M1-like
and M2-like TAMs in the tumor microenvironment. Markers to
distinguish between M1-like and M2-like TAMs include transcription
factors (white circles), pro-inflammatory molecules secreted by
M1-like TAMs, anti-inflammatory molecules secreted by M2-like TAMs
(grey rectangles) and cell membrane receptors. M1-like and M2-like
TAMs share some common characteristics by secreting the same
factors, indicated by the overlay region of the two circles.
Pro-inflammatory molecules secreted by M1-like TAMs have tumor
cytotoxicity inducing the Th1 response (solid line arrows), and
anti-inflammatory molecules secreted by M2-like TAMs trigger the
Th2 response (solid line arrows). PGE2 and IL-6 secreted by both
M1- and M2-like TAMs are indicated by red letters. Molecules
secreted by Th1 and Th2 cell responses induce the polarization of
M1- and M2-like TAMs, respectively (dotted arrows). TAM,
tumor-associated macrophage; TNF, tumor necrosis factor; RNS,
reactive nitrogen species; ROS, reactive oxygen species; NO, nitric
oxide; MMP, matrix metalloproteinase; PGE2; CXCL, C-X-C motif
chemokine ligand; CCL, C-C motif chemokine ligand; IL, interleukin;
TLR, toll-like receptor; MHC, major histocompatibility complex;
IFN, interferon; IRF, interferon regulatory factor; NOS, nitric
oxide synthase; TME, tumor microenvironment; VCAM, vascular cell
adhesion molecule 1; MRC, CD200; Klf, Kruppel like factor; SR,
secretin receptor; MR, major histocompatibility complex class
I-related; CREB, cyclic AMP-responsive element-binding protein;
CSF, colony stimulating factor; VEGF, vascular endothelial growth
factor; FGF, fibroblast growth factor; PDGF, platelet-derived
growth factor; ARG, Arginase; Ym1, chitinase 3 like 1; PGF2,
prostaglandin F; uPA, plasminogen activator, urokinase; MIF,
macrophage migration inhibitory factor; GC, glucocorticoid; Fizz1,
found in inflammatory zone protein 1; MGL, LLGL scribble cell
polarity complex component 1. |
During the transformation of benign cells into
invasive cancer cells, the TME is dominated by cytokines and growth
factors resulting in the dominance of Th2-like immunosuppression,
rather than a Th1-like pro-inflammatory environment (36). This shift results in M1-like TAMs
polarizing into the M2-like TAMs, and the preferential accumulation
of M2-like TAMs in the TME. These abundant cytokines and growth
factors can be derived from tumor cells and non-tumor cells in the
TME. For example, CD4+ T cell-derived Th2 cytokines,
such as CSF1, IL-4, IL-13 and IL-10, promote the polarization of
M1-like into M2-like TAMs in the TME (42) (Fig.
2). In addition, tumor cell-derived microparticles, lactate and
miRs, such as miR-21-5p, miR-125 and miR-146, also promote M2-like
TAMs polarization in the TME (43).
The TME is dominantly populated by M2-like TAMs, but if M1-like
TAMs activity is enhanced, the inhibition of local tumor growth is
observed. The transformation of M2-like into M1-like TAMs under
specific conditions can result in tumor recession. Therefore,
altering the landscape of M1-like and M1-like TAMs in the TME may
have value as a potential TAMs-targeted cancer immunotherapy
(11). This alteration could be
mediated by blocking macrophage infiltration, selectively killing
M2-like TAMs, re-polarizing M2-like to M1-like TAMs or
epigenetically silencing the secretion of M2-like TAM-induced
molecules in the TME (44).
Markers of M1-like TAMs and M2-like
TAMs
Macrophages are differentiated into either M1-like
or M2-like TAMs, which ensures the intraclonal diversity necessary
to maintain an efficient immune response in the TME. As key
transcription factors for M1 differentiation, STAT1, IRF1 and IRF5
promote the activation of M1-like TAMs by activating the
transcription of NO, IFN-γ, CXCL10 and MMP-12, and the specific
receptors of M1-like TAMs, such as CD68, CD80/86 and CD11c
(45) (Fig. 2). The transcription factors STAT3/6,
Krueppel-like factor 2/4 and IRF3/4 promote the activation of
M2-like TAMs and the ARG1-dependent arginine metabolism (45). Meanwhile, M2-like TAMs secret
pro-tumor factors, such as VEGF, fibroblast growth factor (FGF)-2,
CSF1, and express unique receptors, including scavenger and mannose
receptors CD163, CD206 and CD204 (Fig.
2) (9). Antibodies used for TAMs
identification include CD80/86, CD68, FcγRIII, CD14 and HLA-R for
M1-like TAMs, and CD163, CD204, CD206 and Tim-3 for M2-like TAMs
(Fig. 2) (46). M1-like TAMs with markers of
F4/80+ CD11c+ mannose receptor C-type 1
(MRC1)low can be separated from M2-like TAMs with
markers of F4/80 + CD11c− MRC1high
by flow cytometric analysis (47).
TAMs functions in angiogenesis and
angiostasis
The proportion of M2-like TAMs is associated with
the microvessel density of tumor tissue (12). Quantitative analysis of spatial
associations has demonstrated the co-evolution of TAMs and tumor
neovessels during cervical cancer invasion (48). The center of the tumor tissue is
abundant with disorganized and immature blood vessels, and these
central macrophages are associated with the remolding of blood
vessels (49). Through secreting
pro-angiogenic cytokines and growth factors, such as ornithine,
TGF-β, VEGF, basic fibroblast growth factor (bFGF) and CSF1,
M2-like TAMs provide nutrient factors for tumor angiogenesis
(50). Several paracrine axes of
M2-like TAMs are reported to trigger angiogenesis in the TME,
including EGF/VEGFR2, ANG2/Tie2, CCL-18/ERK, Akt/glycogen synthase
kinase (GSK)-3β/Snail, CSF1/CSFR1 and sphingosine-1-phosphate
receptor 1/NLR family pyrin domain-containing protein 3
(NLRP3)/IL-1β signaling axes (Fig.
3). MMP-9 activity of bone marrow-derived CD45+
myeloid cells containing
Tie2+-VEGFR1+-CD11b+-F4/80+
subpopulations is essential and sufficient to initiate angiogenesis
by making sequestered VEGF bioavailable for interaction with its
receptor VEGFR2 (51). Decreased
levels of macrophage-derived VEGF inhibits angiogenesis in solid
tumors by attenuating the formation of vessels (52). CCL-18 secreted by TAMs promotes
angiogenesis in breast cancer by activating ERK and
Akt/GSK-3β/Snail signaling and induces EMT in human umbilical vein
endothelial cells (53). Excess
blood and lymphatic vessel growth promote tumor progression, while
insufficient growth causes tissue ischemia and lymphedema (54). Lymphatic and blood vascular ECs are
regulated by two endothelial specific receptor tyrosine kinase
systems: VEGF/VEGFR2 And ANG/Tie (54). HIF1α upregulates VEGF, Tie2 and ANG2
expression to promote angiogenesis (55). TAMs expressing Tie2 (ANG receptor)
migrate towards ANG2 expressed by angiogenic vessel cells, which
activates ECs and triggers angiogenesis by establishing an
autocrine loop in vascular ECs (55). As shown in Fig. 3, activated ECs secrete ANG2 that
interacts with the Tie2 receptor expressed by TAMs, mediating
cell-to-cell interactions between ECs and TAMs and recruiting
Tie-2-expressing cells to the vasculature (56). Decreased TAM counts mediated by
inhibiting CSF1 leads to substantial attenuation in tumor
angiogenesis. CSF1 upregulates Tie expression on TAMs. Conditional
Tie2 gene knockdown in MRC1+ Tie2-expressing macrophages
decreases tumor angiogenesis (Fig.
3) (56).
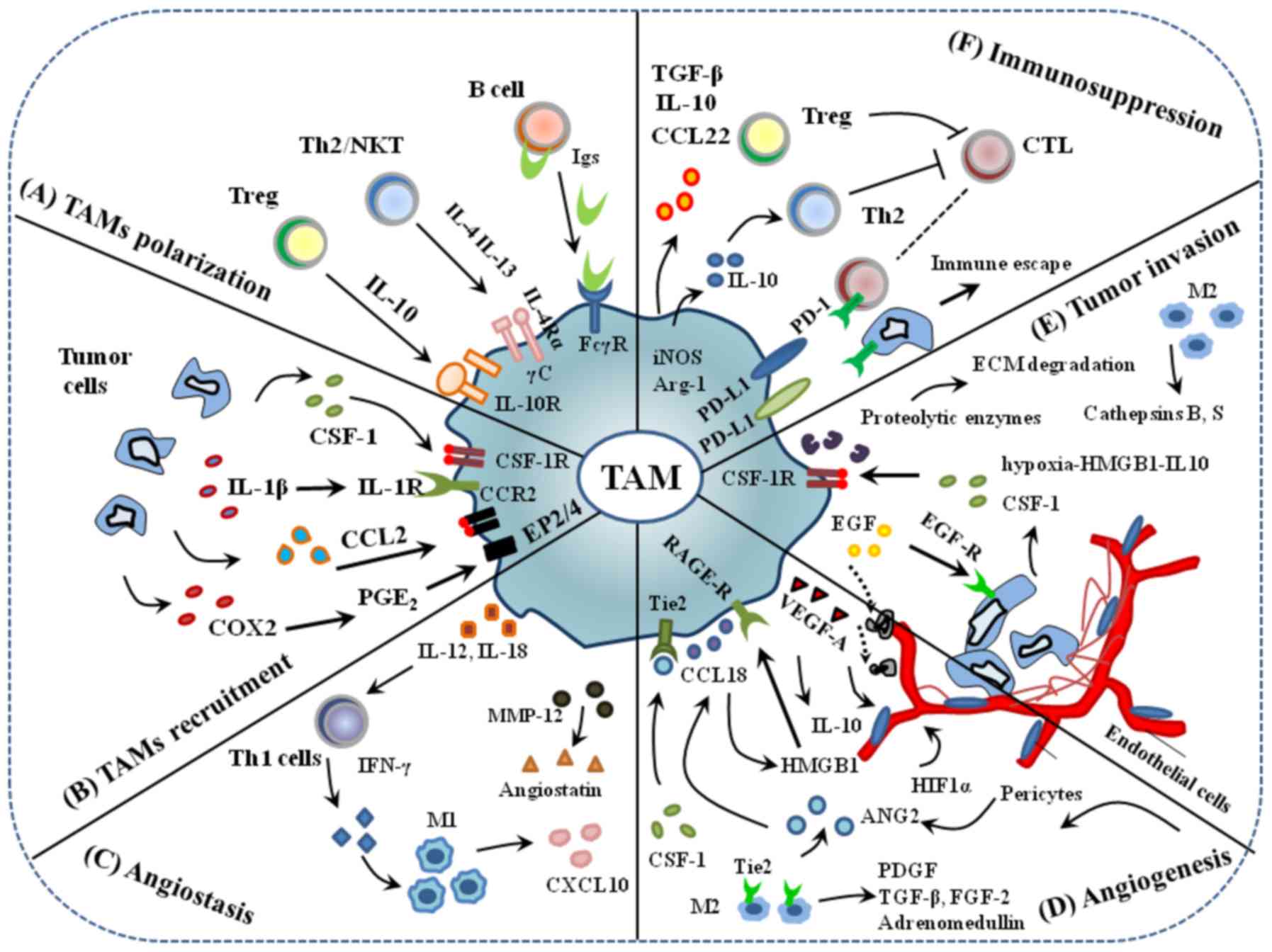 | Figure 3.An overview of TAMs in the TME. (A)
TAM polarization. (B) Tumor cells secrete signal molecules, such as
CSF1 and CCL-2, that interact with their receptors on TAMs to
recruit macrophage into the tumor microenvironment. (C) By
secreting IL-12, 18 and MMP-12 that enhance the levels of IFN-γ and
angiostatic factors, such as CXCL10, and stimulate the
proliferation of M1 macrophages, TAMs possess angiostasis
abilities. (D) TAMs promote angiogenesis through the production of
VEGF-A and other angiogenic factors and express the Tie2 receptor
that can interact with ECs and pericytes to regulate vascular
structure. (E) TAMs secrete proteolytic enzymes, such as MMP and
serine protease, to disrupt the basal layer of extracellular matrix
to disrupt the organization of vascular ECs into blood vessels. (F)
Immunosuppression can occur through soluble or cell surface
mediators, for example TAMs express ligands such as CCL-22, IL-10,
and PD-L1 that interact with immune infiltrating cells and promote
immunosuppression. Immunosuppressive factors TGF-β, TNF-α and IL-10
released by TAMs activate Tregs that cause immunosuppression in the
tumor microenvironment. TAMs, tumor associated macrophages; CSF,
colony stimulating factor; IL, interleukin; MMP, matrix
metalloproteinase; IFN, interferon; CXCL, C-X-C motif chemokine
ligand; VEGF, vascular endothelial growth factor; ECs, endothelial
cells; CCL, C-C motif chemokine ligand; PD-L1, programmed cell
death-ligand 1; TGF, tumor growth factor; TNF, tumor necrosis
factor; B7-H1, B7 homolog 1 or CD274. |
In contrast to M2-like TAM activity in angiogenesis,
M1-like TAMs possess angiostatic properties by secreting IL-12,
IL-18 and MMP-12 (Fig. 3). For
example, the Th1 cell-induced secretion of IFN-γ, IL-12 and IL-18
stimulates the proliferation of M1-like TAMs and the production of
angiostatic factors, such as CXCL10 (57). During tumor progression, PPAR-γ
expression in tumor tissue can switch M1-like TAMs to M2-like TAMs
(12). Selectively blocking PPAR-γ
expression in tumor tissue may be a potential candidate for
TAMs-targeted cancer immunotherapy.
TAMs functions in the invasion and
metastasis of tumor cells
Monitoring tumor metastasis using a multiphoton
microscopy real-time imaging system indicated that a large number
of TAMs are observed at the margin of tumor tissue, with decreasing
numbers in the deeper tumor tissues collected from mouse mammary
tumor virus (MMTV)-polyomavirus middle T antigen (PyMT) and
MMTV-PyMT/c-fms-GFP mice (49). The
observations indicate a critical role of TAMs in the trafficking of
tumor cells into non-tumor tissue, particularly as the invasion and
metastasis of tumor cells are the major cause of cancer-associated
mortality rather than primary tumor growth (58).
Several paracrine signaling axes between tumor cells
and macrophages mediate the migration of both tumor cells and
macrophages, including HMGB1/IL-10, EGF/EGFR, CSF1/CSF1R,
TGF-G/SOX9, CCL4/myosin 3A and TLR4/IL-10 (Fig. 3). Tumor hypoxia increases HMGB1
expression in metastatic melanoma, which promotes IL-10 production
in M2-like TAMs through the receptor for advanced glycation
end-products (26). The paracrine
loop between tumor-synthesized CSF1 and macrophage-produced EGF
mediates the migration and invasion of tumor cells together with
macrophages along collagen fibers, acting as physical pathways
towards blood vessels. Inhibition of either CSF1- or EGF-stimulated
signaling reduces the migration of both tumor cells and macrophages
originating in primary tumor tissues collected from
WAP-Cre/CAG-CAT-EGFP/MMTV-PyMT mice (59) (Fig.
3). IL-4-induced Cathepsin protease activity in TAMs promotes
pancreatic tumor growth and cell invasion in vitro and in
vivo (60). IL-4-expressing
CD4+ T lymphocytes indirectly promote invasion and
subsequent metastasis of mammary adenocarcinomas by directly
regulating the phenotype and effector function of
CD11b+Gr1−F4/80+ TAMs, in turn
enhancing metastasis through activating EGFR signaling in malignant
mammary epithelial cells (61). In
addition, Heregulin B1 and CXCL12 function as tumor metastasis
mediators by controlling the EGF/CSF1 paracrine invasion loop
(62). Large quantities of Versican
produced by Lewis lung cancer cells promote the proliferation of
metastatic tumor cells by activating macrophages through the
TLR2/TLR6 axis (63). The STAT3/6
signaling pathways synergistically increase Cathepsin secretion
from M2-like TAMs, which promote macrophage-mediated pancreatic
cancer cell invasion in a Cathepsin-dependent manner (64). In a triple-negative breast cancer
mouse model, local and systemic levels of TAM-induced MMP-9, VEGF,
Ym1 and Lipocalin-2 mediate metastasis in breast cancer. M2-like
TAMs secrete proteases, such as MMPs and serine protease, to
disrupt cell-cell junctions, the basal membrane and the
organization of vascular ECs into blood vessels, which allows tumor
cells to pass through the ECM and facilitate the migration and
invasion of tumor cells in numerous types of tumor, including
breast and lung cancer (65)
(Fig. 3). Hyaluronic acid (HA), a
major component of the ECM, is specifically recognized by
macrophages expressing the HA receptor CD44 (66). HA-CD44 interactions serve important
roles in monocyte adhesion and recruitment, as well as macrophage
recruitment to support tumor growth and metastasis (66).
CXCL1 secreted by M2-like TAMs promotes breast
cancer invasion and EMT by activating NF-κB/SOX4 signaling
(67). In fact, there are several
CCLR axes that can enhance the migration of cancer cells by
blocking androgen/AR signaling. For example, the CCL5/CCR5 axis
inhibits androgen/AR signaling as an upstream mediator in prostate
cancer cells (68). While the
CCL2/CCR2 axis is negatively regulated by androgen/AR signaling,
with the CCL22/CCR4 axis acting as a further downstream mediator,
both axes promote prostate cancer cell migration (68). CCL4 promotes prostate carcinogenesis
through macrophage androgen/AR signaling. The CCL21/CCR7 axis is
activated by TNF-α and induces lymph node metastasis in prostate
cancer (68). VEGF-A also stimulate
TAMs to produce CXCL1, and elevated CXCL1 in premetastatic liver
tissue promotes the recruitment of CXCR2-positive myeloid-derived
suppressor cells (MDSCs) to form a premetastatic niche, which
ultimately promotes liver metastases (69). Tumor-derived CSF2 induced Bv8
expression in myeloid cells to enhance myelopoiesis and
mobilization of MDSCs from the bone marrow (70). The aforementioned data demonstrate
that TAMs functions in the invasion and metastasis of tumor cells
are complex. A schematic model representing the sensors, effector
molecules and the corresponding functions of the M2-like TAMs are
illustrated in Fig. 4. Some of these
molecules have been defined as M2 markers (such as CD206 and
CD163), while others have not (such as CD209), but they all
function in the activation of M2-like TAMs (Fig. 4) (71).
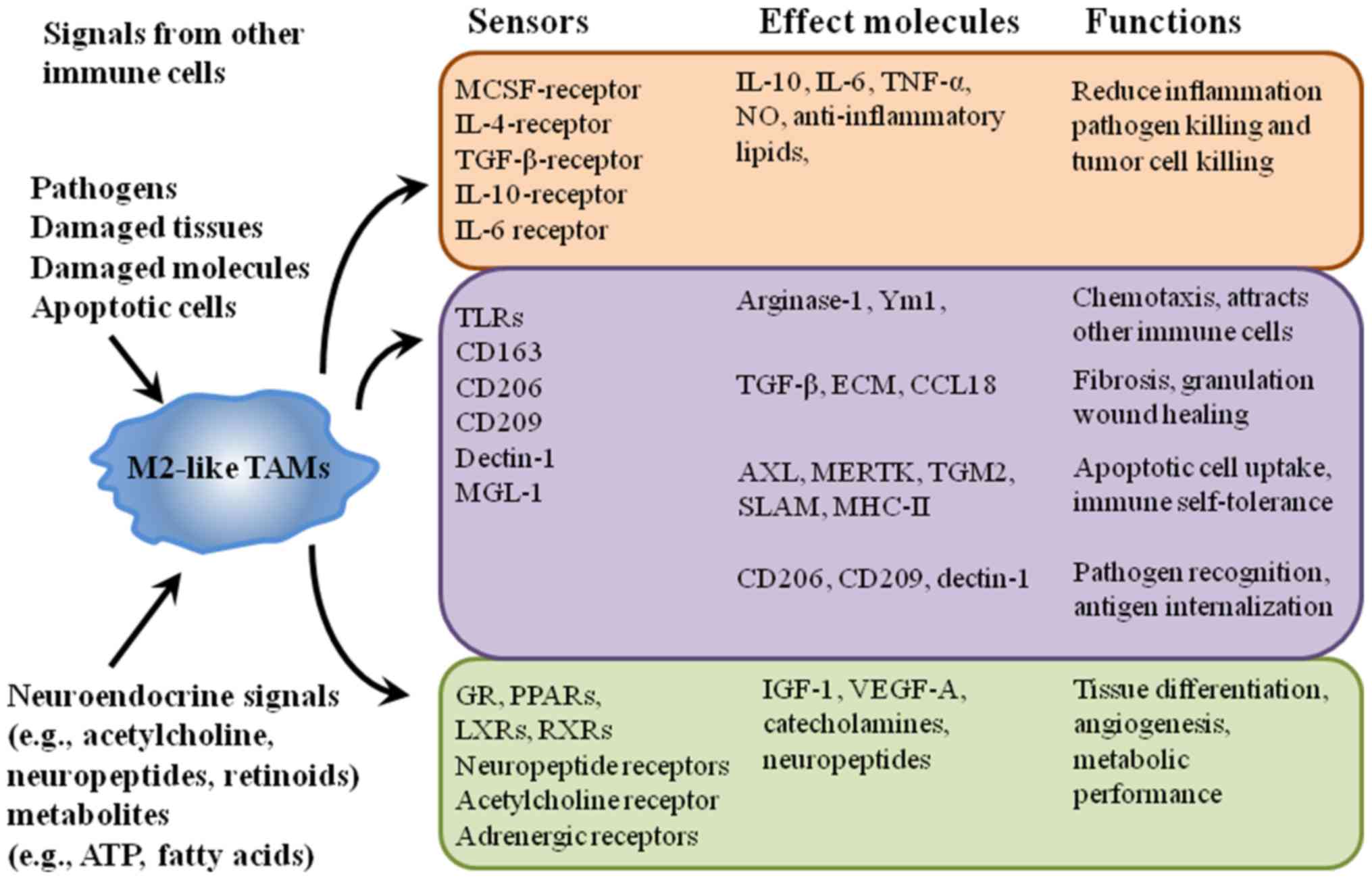 | Figure 4.Alternative model depicting sensor
and functional effects of the M2-like macrophages. The macrophage
phenotype is principally determined by eliciting signals derived
from immune cells, pathogens, apoptotic or damaged cells, and a
wide range of chemical mediators. These signals act through
receptors and signal pathways that elicit a wide range of
functional effects. CSF1, colony stimulating factor 1; IL,
interleukin; TGF, transforming growth factor; TNF, tumor necrosis
factor; Ym1, chitinase 3 like 1; TLR, toll-like receptor; MGL-1,
LLGL scribble cell polarity complex component 1; ECM, extracellular
matrix; CCL, C-C motif chemokine ligand; AXL, AXL receptor tyrosine
kinase; MERTK, MER proto-oncogene tyrosine kinase; TGM2,
transglutaminase 2; SLAM, signaling lymphocytic activation
molecule; MHC-II, major histocompatibility complex class II; IGF,
insulin-like growth factor; VEGF, vascular endothelial growth
factor; GR, glutathione reductase; PPAR, peroxisome proliferator
activated receptor; LXR, nuclear receptor subfamily 1; RXR,
retinoid X receptor. |
TAMs immune effects in the TME
With high phagocytic capacity, macrophages are the
first line of innate immune defense against abnormal cell damage in
tissues. By mediating and providing the required costimulatory
signaling and cytokine secretion, and identifying and presenting
foreign antigens on MHC I and II molecules to T cells, macrophages
also serve a central role in T-cell effective activation of the
adaptive immune response (72).
M1-like TAMs secrete Th1-inducing NO that can directly kill cancer
cells in non-specific manner. In turn, activated Th1 responses
further promote the activation of M1-like TAMs, CD8+ T
cells, IgG B cells and IFN-γ-producing CD4+ T cells
(73). By releasing pro-inflammatory
molecules, such as TNF-α, IFNγ and ROS/RNS, activating TLRs and
decreasing the expression of anti-inflammatory factors, such as
ARG1, TGFβ and IL10, M1-like TAMs promote the inflammatory response
and anti-tumor activity in the TME (Fig.
5). These pro-inflammatory cytokines secreted by M1-like TAMs
trigger the tumoricidal actions of natural killer (NK) cells,
stimulate cytotoxic type Th1 and tumor-specific cytotoxic T cell
responses, and induce the activity of cytotoxic CD8+ T
cells (Fig. 5) (73). M1-like TAMs, but not M2-like TAMs,
are able to release pro-inflammatory IL-12 that is required in
response to the antitumor activities mediated by NK, Th1 and CTL
cells (74).
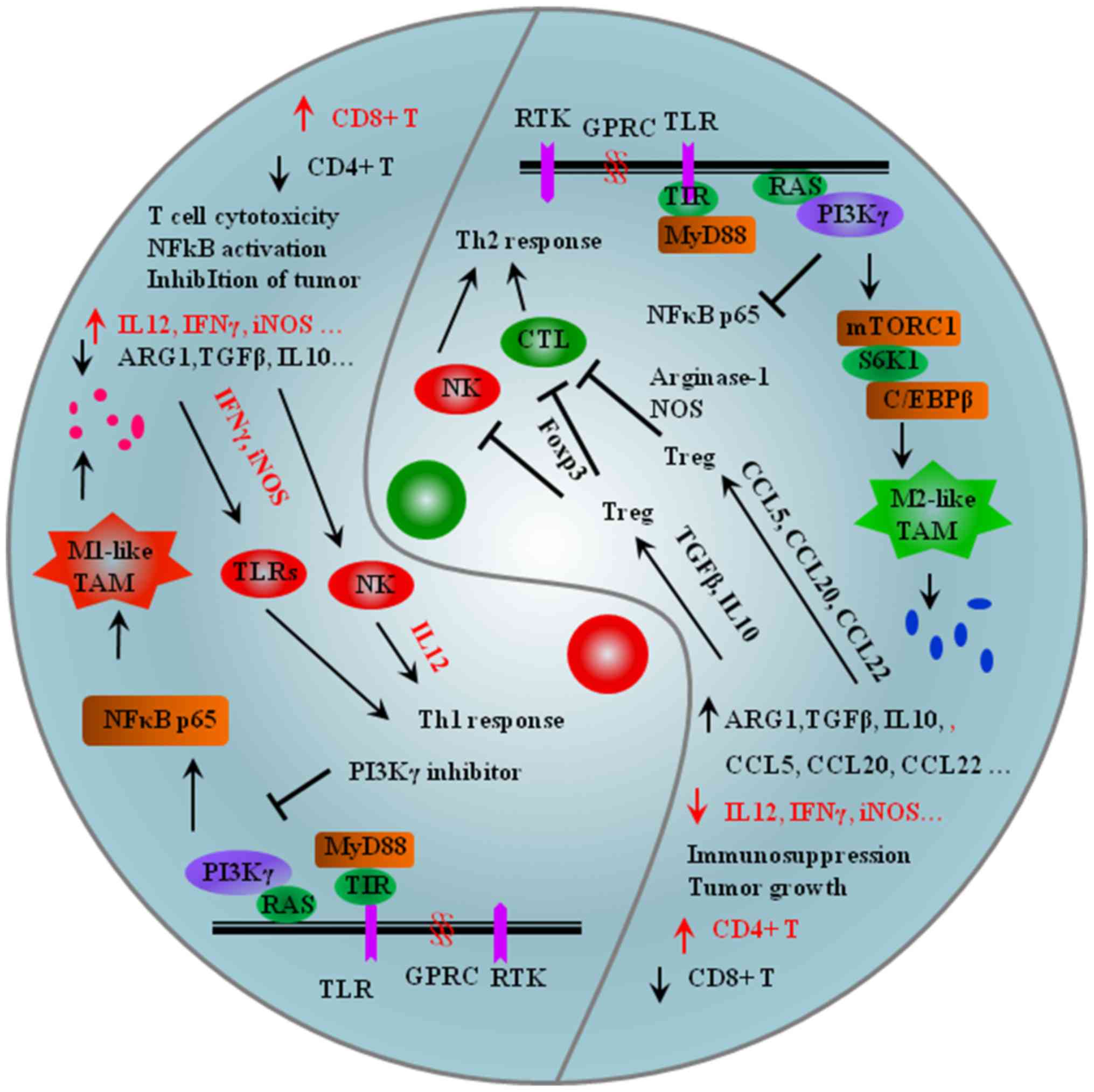 | Figure 5.Opposite immune effects of M1-like
and M2-like TAMs in the tumor microenvironment. Left: IL12, IFNγ
and iNOS derived from M1-like TAMs activate NK cells and TLRs that
cause Th1 pro-inflammatory response and anti-tumor activity.
Inhibition of PI3Kγ strongly activates the NF-κB pathway, causing
increased expression of proinflammatory factors, including IL12 and
IFNγ. Right: TGF-β and IL10 derived from M2-like TAMs activate
Tregs by upregulating Foxp3 in CD4+ T cells that cause Th2
anti-inflammatory response and pro-tumor activity. CCL5, CCL20 and
CCL22 released by M2-like TAMs activate iTregs, Arginase-1 and NOS
to suppress effector T cells to promote immune suppression in the
TME. PI3Kγ works downstream of many chemoattractant-receptors, such
as receptor tyrosine kinases (VEGF-R1 and CSF1R) and G
protein-coupled receptors (CCR2 and CXCR4). Inhibition of the NF-κB
pathway activates PI3Kγ, causing increased expression of
anti-inflammatory factors, including ARG1, TGFβ and CCL5. mTORC1,
mammalian target of rapamycin complex 1; MyD88, Myeloid
differentiation primary response 88 protein; PI3Kγ,
phosphoinositide 3-kinase γ; TIR, toll/interleukin receptor domain;
TLR/ILR, Toll-like receptor/interleukin receptor; ARG, Arginase;
TGFβ, transforming growth factor β; CCL, C-C motif chemokine
ligand; IL, interleukin; IFNγ, interferon γ; iNOS, inducible nitric
oxide synthase; GPCR, G protein-coupled receptor; RTK, receptor
tyrosine kinase; C/EBPβ, enhancer-binding proteins; RAS, KRAS
proto-oncogene, GTPase. |
Several chemoattractant, cytokines and enzymes
derived from M2-like TAMs can stimulate the activation of induced
regulatory T cells (iTregs) and recruit natural Tregs (nTregs),
which exert immunosuppressive effects by directly inhibiting the
function of effector T cells or secreting immunosuppressive
factors, such as CCL5 and CCL20 (Fig.
5). For example, M2-like TAM-derived IL-10, TGF-β, PGE2 and
prostanoids inhibit the cytotoxic function of effector T and NK
cells, promote the development of Tregs and activate iTregs and
ineffective APCs by upregulating Foxp3 in CD4+ T cells
(75). In addition, CCL5, CCL20 and
CCL22 released by M2-like TAMs recruit and activate iTregs to
suppress the function of effector T cells, resulting in immune
suppression in the TME (Fig. 5).
M2-like TAMs secrete CCL22 in human ovarian cancer, mediating Treg
trafficking to tumor tissue through the CCL22/CCR4 axis (76). M2-like TAMs secreted CCL20, which
recruits CCR6+ nTregs to promote immunosuppression in
the TME (55). M2-like TAMs suppress
T cell proliferation in part by expressing ARG1 that catabolizes
arginine necessary for T cell activation and proliferation;
decreased arginine levels results in impaired effector T cell
function in the TME (77). Small
extracellular vesicles containing ARG1 suppress CD4+ and
CD8+ T cell proliferation in vitro and in
vivo in mouse models of ovarian carcinomas (78). Tumor cells use extracellular vesicles
to transport the metabolic checkpoint molecule ARG1 over long
distances to immune cells to weaken antitumor immune responses
(78).
PI3Kγ functions downstream of several
chemoattractant receptors, such as RTKs (including VEGF-R1 and
CSF1R) and GPCR (including CCR2 and CXCR4) (Fig. 5). The inhibition of PI3Kγ activates
the NF-κB pathway, increases the levels of proinflammatory
cytokines, including TNFα, IL-12, NOS2 and MHCII on APCs (including
macrophages and DCs), and inhibits the expression of
immunosuppressive factors, such as IL-10, TGFβ, ARG1 and CCL2
(79). Chemoattractants such as
SDF-1a, VEGFa and IL-1b activate RTKs, GPCRs and TLR/ILR signaling
molecules that initiate tumor inflammation by activating the PI3K
isoform p110γ in Gr1+CD11b+ myeloid cells
(28). M2-like TAMs produce
anti-inflammatory immunosuppression and pro-tumor activity by
releasing growth-promoting molecules like ornithine, which promotes
Th2-type cytotoxic responses, increases the levels of
anti-inflammatory cytokines, such as IL-4, IL-10, IL-13 and IL-10,
decreases levels of pro-inflammatory factors, including IL12,
IFNγ and iNOS, and induces ineffective antigen presentation
(Fig. 5) (79).
Potential strategies for TAMs-targeted
cancer immunotherapy
The predominate presence of M2-like TAMs in the TME
is partially responsible for tumor immune evasion and
chemoresistance, and the potential of TAM-targeted tumor
immunotherapies has received considerable interest. The current
strategies being explored for such cancer treatment include
(Table II): i) blocking the
infiltration of macrophages into the TME (15); ii) depleting/killing dominating
M2-like TAMs in the TME; iii) reprogramming M2-like TAMs into the
M1-like phenotype in the TME (80);
and iv) TAM-mediated delivery of therapeutics (81).
| Table II.Strategies for TAM-targeted antitumor
therapy. |
Table II.
Strategies for TAM-targeted antitumor
therapy.
| A, Blocking TAMs
infiltration to the TME |
|---|
|
|---|
| First author,
year | Potential
agents | Mechanism of
action | (Refs.) |
|---|
| Pathria et
al, 2019 | PF-04136309,
MLN1202, CCX872-B and BMS-813160 | CCR-2 inhibitors
targeting CCL-2/CCR-2 axis | (79) |
|
| B, Depleting
M2-like TAMs in the TME |
|
| First author,
year | Potential
agents | Mechanism of
action | (Refs.) |
|
| Lee et al,
2019 | The hybrid peptide
of MEL-dKLA | Inducing CD206+
M2-like TAMs apoptosis | (83) |
| Opperman et
al, 2019 | Liposomes with
encapsulation of clodronate | Decreases levels of
macrophage-derived insulin-like growth factor 1 | (92) |
| Zhang et al,
2019 | Nanocarriers with a
nanobody specific for CD206 | Anti-CD206
nanobodies inhibit angiogenesis | (93) |
| Zhang et al,
2019 | In
vitro-transcribed mRNA | Switching M1-like
reprogramming | (93) |
|
| C, Reprogramming
TAMs from M2-like to M1-like TAMs |
|
| First author,
year | Potential
agents | Mechanism of
action | (Refs.) |
|
| Wanderley et
al, 2018 | Paclitaxel
(Taxol) | TLR4-dependent
manner | (85) |
| Andersen et
al, 2019 | CD163-targeted
corosolic acid-containing liposomes | Specific inhibition
of STAT3 | (84) |
| Tan et al,
2015 | Baicalin |
Autophagy-associated activation of
RelB/p52 | (87) |
| Locatelli et
al, 2019 | RP6530 | PI3K δ/γ-dependent
pathway | (86) |
| Buhtoiarov et
al, 2011 |
Cyclophosphamide | Up-regulating the
levels of the M1-associated molecules (CD40, CD80, CD86, MHC class
II, IFN-γ, TNF-α, IL-12) and down-regulating the levels of the
M2-associated molecules (IL-4Rα, B7-H1, IL-4, IL-10). | (94) |
| Di Caro et
al, 2016 | Gemcitabine | Improving the
expression of the M1 markers HLA-DR, CD40, CCR7, decreasing the
expression of M2 markers CD163 and CD206 | (95) |
|
| D, TAM-mediated
delivery of therapeutic systems |
|
| First author,
year | Potential
agents | Mechanism of
action | (Refs.) |
|
| Choi et al,
2012 | Liposomal-Dox
delivered by macrophages |
| (91) |
Blocking the signaling axes of chemoattractants and
their receptors (such as CCL2/CCR2+,
CCL2/CCR5+, IL-1β/IL-1R, VEGFA/VEGFR, CSF1/CSFR and
Tie/ANG2) may be potential candidates for targeting
macrophage-recruitment therapy. For example, clinical trials with
several CCR-2 inhibitors, such as PF-04136309 and MLN1202, are
currently ongoing for the treatment of solid tumors, including
pancreatic ductal adenocarcinoma and colorectal cancer (79) (Table
III). Combination of therapy blocking CCL-2 or CCR-2 signaling
with chemo-, radio- or immunotherapy improves antitumor effects by
decreasing the infiltration of myeloid cells in preclinical mouse
models (82). In addition to therapy
blocking macrophage recruitment, selective elimination of the
dominating M2-like TAMs in the TME can inhibit tumor growth and
restore local immune surveillance. For example, a hybrid peptide of
MEL-dKLA selectively triggered M2-like TAM apoptosis without
affecting other leukocytes, such as T cells and DCs, and increased
the M1/M2 ratio, which reduced tumor growth rates, tumor weights
and angiogenesis in a lung cancer mouse model (83) (Table
III).
| Table III.Potential agents to target the
reprogramming TAMs from M2-like to M1-like TAMs in cancer
treatment. |
Table III.
Potential agents to target the
reprogramming TAMs from M2-like to M1-like TAMs in cancer
treatment.
| Agents | Applications | Effects on
TAMs | Mechanism of
action |
|---|
| Paclitaxel
(Taxol) | In vitro and
in vivo models of breast and melanoma tumors | Altered the
signature of TAMs from M2-like to M1-like TAMs | TLR4-dependent
manner (85) |
| CD163-targeted
corosolic acid-containing liposomes | Monocytes and
macrophages | M1-like
reprogramming at the mRNA level | Specific inhibition
of STAT384 |
| RP6530 | Hodgkin lymphoma
in vitro and in vivo | Switching M1-like
reprogramming | PI3Kδ/γ-dependent
pathway (86) |
| The hybrid peptide
of MEL-dKLA | In vitro and
in mouse models of lung carcinoma | Induced apoptosis
in CD206+ M2-like TAMs | Induces
mitochondrial death after cell membrane penetration (68) |
| Baicalin | In vitro and
in vivo hepatocellular carcinoma mouse model | Initiating TAM
reprogramming to an M1-like TAM |
Autophagy-associated activation of
RelB/p52 (87) |
|
Cyclophosphamide | Multiple myeloma,
leukemia, breast cancer, neuroblastoma, lymphoma, ovarian cancer,
retinoblastoma | Altered the
signature of TAMs from M2-like to M1-like TAMs in a manner | Enhances
pro-inflammatory IL-6 and IL-12, and decreases anti-inflammatory
IL-10 and TGF-β (94) |
| Gemcitabine |
| Re-education of
macrophages to M1-like TAMs by upregulating the levels of the M1
markers HLA-DR, CD40, CCR7, downregulating the levels of M2 markers
CD163 and CD206 | Activation of the
pro-inflammatory M1-like TAMs (95) |
| Nanocarrier
encoding M1-polarizing transcription factors |
| Reprogramming
M2-like TAMs to M1-like TAMs | Induces anti-tumor
immunity and tumor regression (93) |
Different studies have shown that genetic or
epigenetic reprogramming of M2-like TAMs into M1-like TAMs in the
TME has promising effects. Specific STAT3 inhibition in human
monocytes and macrophages by CD163-targeted corosolic
acid-containing liposomes promotes M1-like TAMs reprogramming,
inhibits STAT3-regulated IL-10 expression and increases
pro-inflammatory TNFα levels (84).
Paclitaxel alters the signature of TAMs in the TME from a M2-like
pro-tumor profile (CD206, RELMα, MMP9 and ARG1) to a M1-like
antitumor profile (IL12, iNOS and IL6), inducing tumor regression
by reprogramming in a TLR4-dependent manner in mouse models of
breast and melanoma tumors (85)
(Table III). RP6530 re-polarizes
M2-like TAMs into M1-like TAMs to inhibit the growth of tumor
vasculature, leading to tumor regression via the PI3Kδ/γ-dependent
pathway (86). Oral administration
of baicalin mediates the re-polarization of M2-like into M1-like
TAMs in the TME and the inhibition of hepatocellular carcinoma in
an orthotopic mouse model by autophagy-induced RelB/p52 activation
(87). The combination of PMX-53 (a
C5aR1 peptide antagonist) and paclitaxel synergistically inhibits
tumor growth by re-polarizing M2-like towards the M1-like TAM
phenotype, inhibiting angiogenesis and recruiting cytotoxic T
lymphocytes (88). TMP195
re-polarizes M2-like into M1-like TAMs and synergizes with PD-1
antibody to reduce tumor burden and metastasis in an autochthonous
mouse model of breast cancer (89).
In addition, macrophages have great potential in cancer drug
delivery because they can sense chemotactic cues and migrate to
tumors with high efficiency (90).
For example, liposomal-doxorubicin delivered by macrophages
exhibited higher therapeutic efficacy than doxorubicin delivered by
liposome or doxorubicin alone in both subcutaneous and metastasis
xenograft lung tumor models (91)
(Tables II and III). Therapy with Dox-laden nanocapsules
leads to efficient tumor growth suppression, while causing little
systemic toxicity in U87MG tumor bearing nude mice (90).
Conclusion
As summarized in the present review, the
preferential accumulation of M2-like TAMs is a major contributor to
the establishment of metastatic and malignant tumors. TAM-targeted
cancer immunotherapies are now being explored and developed,
including: i) Blocking macrophage recruitment; ii) M2-like
TAM-targeted depletion in the TEM; iii) M2-like TAM-targeted
reprogramming therapy in the TEM; and iv) macrophage-mediated drug
delivery systems. However, there are a number of unsolved
challenges, such as rapid clearance from the blood circulation,
inefficient targeting and cytotoxicity issues, that will limit the
application of TAM-targeted therapy in the clinic. Additionally,
increasing M1-like TAMs through the re-polarization of M2-like TAMs
into M1-like TAMs may induce the infiltration of T lymphocytes into
tumors and increase their ability to kill tumor cells, while
overzealous M1-like TAMs contributing to chronic inflammation may
lead to atherosclerosis and other chronic inflammatory conditions
(11). New technologies, such as
single cell sequencing, and digitalization and visual spatial
analysis models, may help to resolve some of these problems.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
National Nature Science Foundation of China (grant no. 31160233),
the Undergraduate Innovation Program at Nanchang University (grant
no. 20190402294) and the Science and Technology Foundation of
Jiangxi Province (grant no. 20142BAB204013).
Availability of data and materials
Not applicable.
Authors' contributions
MY and KZ were responsible for the writing of the
manuscript. JZ, XP and YZ prepared Figures 1–3.
JW and XC prepared Figure 4 and
Table I. TC modified the writing of
the manuscript and the figures. All authors participated in the
whole process of writing and modifying the manuscript, and read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent to participate
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Zent CS and Elliott MR: Maxed out macs:
Physiologic cell clearance as a function of macrophage phagocytic
capacity. FEBS J. 284:1021–1039. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhang Q, L Y, Bian H, Guo L and Zhu H:
Activation of the α7 nicotinic receptor promotes
lipopolysaccharide-induced conversion of M1 microglia to M2. Am J
Transl Res. 9:971–985. 2017.PubMed/NCBI
|
|
3
|
Yao Y, Xu XH and Jin L: Macrophage
Polarization in physiological and pathological pregnancy. Front
Immunol. 10:7922019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gordon S: Alternative activation of
macrophages. Nat Rev Immunol. 3:23–35. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gensel JC and Zhang B: Macrophage
activation and its role in repair and pathology after spinal cord
injury. Brain Res. 1619:1–11. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lefèvre L, Lugo-Villarino G, Meunier E,
Valentin A, Olagnier D, Authier H, Duval C, Dardenne C, Bernad J,
Lemesre JL, et al: The C-type lectin receptors dectin-1, MR, and
SIGNR3 contribute both positively and negatively to the macrophage
response to Leishmania infantum. Immunity. 38:1038–1049. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang LX, Zhang SX, Wu HJ, Rong XL and Guo
J: M2b macrophage polarization and its roles in diseases. J Leukoc
Biol. 106:345–358. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang Y, Smith W, Hao D, He B and Kong L:
M1 and M2 macrophage polarization and potentially therapeutic
naturally occurring compounds. Int Immunopharmacol. 70:459–466.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang L and Zhang Y: Tumor-associated
macrophages: From basic research to clinical application. J Hematol
Oncol. 10:582017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ferrante CJ, Pinhal-Enfield G, Elson G,
Cronstein BN, Hasko G, Outram S and Leibovich SJ: The
adenosine-dependent angiogenic switch of macrophages to an M2-like
phenotype is independent of interleukin-4 receptor alpha (IL-4Rα)
signaling. Inflammation. 36:921–931. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mills CD, Lenz LL and Harris RA: A
breakthrough: Macrophage-directed cancer immunotherapy. Cancer Res.
76:513–516. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Herwig MC, Bergstrom C, Wells JR, Höller T
and Grossniklaus HE: M2/M1 ratio of tumor associated macrophages
and PPAR-gamma expression in uveal melanomas with class 1 and class
2 molecular profiles. Exp Eye Res. 107:52–58. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang HC, Chen CW, Yang CL, Tsai IM, Hou
YC, Chen CJ and Shan YS: Tumor-associated macrophages promote
epigenetic silencing of gelsolin through DNA methyltransferase 1 in
gastric cancer cells. Cancer Immunol Res. 5:885–897. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sarode P, Zheng X, Giotopoulou GA, Weigert
A, Kuenne C, Günther S, Friedrich A, Gattenlöhner S, Stiewe T,
Brüne B, et al: Reprogramming of tumor-associated macrophages by
targeting β-catenin/FOSL2/ARID5A signaling: A potential treatment
of lung cancer. Sci Adv. 6:eaaz61052020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li X, Liu R, Su X, Pan Y, Han X, Shao C
and Shi Y: Harnessing tumor-associated macrophages as aids for
cancer immunotherapy. Mol Cancer. 18:1772019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Larionova I, Cherdyntseva N, Liu T,
Patysheva M, Rakina M and Kzhyshkowska J: Interaction of
tumor-associated macrophages and cancer chemotherapy.
Oncoimmunology. 8:15960042019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lee HW, Choi HJ, Ha SJ, Lee KT and Kwon
YG: Recruitment of monocytes/macrophages in different tumor
microenvironments. Biochim Biophys Acta. 1835:170–179.
2013.PubMed/NCBI
|
|
18
|
Fantuzzi L, Tagliamonte M, Gauzzi MC and
Lopalco L: Dual CCR5/CCR2 targeting: Opportunities for the cure of
complex disorders. Cell Mol Life Sci. 76:4869–4886. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Vogel DY, Heijnen PD, Breur M, de Vries
HE, Tool AT, Amor S and Dijkstra CD: Macrophages migrate in an
activation-dependent manner to chemokines involved in
neuroinflammation. J Neuroinflammation. 11:232014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Byrne SN, Knox MC and Halliday GM: TGFbeta
is responsible for skin tumour infiltration by macrophages enabling
the tumours to escape immune destruction. Immunol Cell Biol.
86:92–97. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tang M, Liu B, Bu X and Zhao P: Cross-talk
between ovarian cancer cells and macrophages through periostin
promotes macrophage recruitment. Cancer Sci. 109:1309–1318. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ruffell B, Affara NI and Coussens LM:
Differential macrophage programming in the tumor microenvironment.
Trends Immunol. 33:119–126. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Henze AT and Mazzone M: The impact of
hypoxia on tumor- associated macrophages. J Clin Invest.
26:3672–3679. 2016. View Article : Google Scholar
|
|
24
|
Casazza A, Laoui D, Wenes M, Rizzolio S,
Bassani N, Mambretti M, Deschoemaeker S, Van Ginderachter JA,
Tamagnone L and Mazzone M: Impeding macrophage entry into hypoxic
tumor areas by Sema3A/Nrp1 signaling blockade inhibits angiogenesis
and restores antitumor immunity. Cancer Cell. 24:695–709. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Clarijs R, Schalkwijk L, Ruiter DJ and de
Waal RM: EMAP-II expression is associated with macrophage
accumulation in primary uveal melanoma. Invest Ophthalmol Vis Sci.
44:1801–1816. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Huber R, Meier B, Otsuka A, Fenini G,
Satoh T, Gehrke S, Widmer D, Levesque MP, Mangana J, Kerl K, et al:
Tumour hypoxia promotes melanoma growth and metastasis via high
mobility group box-1 and M2-like macrophages. Sci Rep. 6:299142016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tang C, Liu T, Wang K, Wang X, Xu S, He D
and Zeng J: Transcriptional regulation of FoxM1 by HIF-1α mediates
hypoxia-induced EMT in prostate cancer. Oncol Rep. 42:1307–1318.
2019.PubMed/NCBI
|
|
28
|
Schmid MC, Avraamides CJ, Dippold HC,
Franco I, Foubert P, Ellies LG, Acevedo LM, Manglicmot JR, Song X,
Wrasidlo W, et al: Receptor tyrosine kinases and TLR/IL1Rs
unexpectedly activate myeloid cell PI3kγ, a single convergent point
promoting tumor inflammation and progression. Cancer Cell.
19:715–727. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kessenbrock K, Plaks V and Werb Z: Matrix
metalloproteinases: Regulators of the tumor microenvironment. Cell.
141:52–67. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Arendt LM, McCready J, Keller PJ, Baker
DD, Naber SP, Seewaldt V and Kuperwasser C: Obesity promotes breast
cancer by CCL2-mediated macrophage recruitment and angiogenesis.
Cancer Res. 73:6080–6093. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hao J, Yan F, Zhang Y, Triplett A, Zhang
Y, Schultz DA, Sun Y, Zeng J, Silverstein KAT, Zheng Q, et al:
Expression of adipocyte/macrophage fatty acid binding protein in
tumor associated macrophages promotes breast cancer progression.
Cancer Res. 78:2343–2355. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Komohara Y and Takeya M: CAFs and TAMs:
Maestros of the tumour microenvironment. J Pathol. 241:313–315.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Miyake M, Hori S, Morizawa Y, Tatsumi Y,
Nakai Y, Anai S, Torimoto K, Aoki K, Tanaka N, Shimada K, et al:
CXCL1-mediated interaction of cancer cells with tumor-associated
macrophages and cancer-associated fibroblasts promotes tumor
progression in human bladder cancer. Neoplasia. 18:636–646. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wan S, Zhao E, Kryczek I, Vatan L,
Sadovskaya A, Ludema G, Simeone DM, Zou W and Welling TH:
Tumor-associated macrophages produce interleukin 6 and signal via
STAT3 to promote expansion of human hepatocellular carcinoma stem
cells. Gastroenterology. 147:1393–1404. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yi L, Xiao H, Xu M, Ye X, Hu J, Li F, Li
M, Luo C, Yu S, Bian X and Feng H: Glioma-initiating cells: A
predominant role in microglia/macrophages tropism to glioma. J
Neuroimmunol. 232:75–82. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Biswas SK and Mantovani A: Macrophage
plasticity and interaction with lymphocyte subsets: Cancer as a
paradigm. Nat Immunol. 11:889–896. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Murray PJ: Macrophage polarization. Annu
Rev Physiol. 79:541–566. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chen JJ, Yao PL, Yuan A, Hong TM, Shun CT,
Kuo ML, Lee YC and Yang PC: Up-regulation of tumor interleukin-8
expression by infiltrating macrophages: Its correlation with tumor
angiogenesis and patient survival in non-small cell lung cancer.
Clin Cancer Res. 9:729–737. 2003.PubMed/NCBI
|
|
39
|
Noy R and Pollard JW: Tumor-associated
macrophages: From mechanisms to therapy. Immunity. 41:49–61. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Liu YC, Zou XB, Chai YF and Yao YM:
Macrophage polarization in inflammatory diseases. Int J Biol Sci.
10:520–529. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Li X, Luo H, Ye Y, Chen X, Zou Y, Duan J
and Xiang D: β-glucan, a dectin-1 ligand, promotes macrophage M1
polarization via NF-κB/autophagy pathway. Int J Oncol. 54:271–282.
2019.PubMed/NCBI
|
|
42
|
Su S, Liu Q, Chen J, Chen J, Chen F, He C,
Huang D, Wu W, Lin L, Huang W, et al: A positive feedback loop
between mesenchymal-like cancer cells and macrophages is essential
to breast cancer metastasis. Cancer Cell. 25:605–620. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Pan Z, Tian Y, Niu G and Cao C: Role of
microRNAs in remodeling the tumor microenvironment. Int J Oncol.
56:407–416. 2020.PubMed/NCBI
|
|
44
|
Peng D, Kryczek I, Nagarsheth N, Zhao L,
Wei S, Wang W, Sun Y, Zhao E, Vatan L, Szeliga W, et al: Epigenetic
silencing of TH1-type chemokines shapes tumour immunity and
immunotherapy. Nature. 527:249–253. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Lawrence T and Natoli G: Transcriptional
regulation of macrophage polarization: Enabling diversity with
identity. Nat Rev Immunol. 11:750–761. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Heusinkveld M and van der Burg SH:
Identification and manipulation of tumor associated macrophages in
human cancers. J Transl Med. 9:2162011. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Rolny C, Mazzone M, Tugues S, Laoui D,
Johansson I, Coulon C, Squadrito ML, Segura I, Li X, Knevels E, et
al: HRG inhibits tumor growth and metastasis by inducing macrophage
polarization and vessel normalization through downregulation of
PlGF. Cancer Cell. 19:31–44. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Jiang S, Yang Y, Fang M, Li X, Yuan X and
Yuan J: Co-evolution of tumor-associated macrophages and tumor
neo-vessels during cervical cancer invasion. Oncol Lett.
12:2625–2631. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wyckoff JB, Wang Y, Lin EY, Li JF, Goswami
S, Stanley ER, Segall JE, Pollard JW and Condeelis J: Direct
visualization of macrophage-assisted tumor cell intravasation in
mammary tumors. Cancer Res. 67:2649–2656. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Caux C, Ramos RN, Prendergast GC,
Bendriss-Vermare N and Ménétrier-Caux C: A Milestone review on how
macrophages affect tumor growth. Cancer Res. 76:6439–6442. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Du R, Lu KV, Petritsch C, Liu P, Ganss R,
Passegué E, Song H, Vandenberg S, Johnson RS, Werb Z and Bergers G:
HIF1alpha induces the recruitment of bone marrow-derived vascular
modulatory cells to regulate tumor angiogenesis and invasion.
Cancer Cell. 13:206–220. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Stockmann C, Doedens A, Weidemann A, Zhang
N, Takeda N, Greenberg JI, Cheresh DA and Johnson RS: Deletion of
vascular endothelial growth factor in myeloid cells accelerates
tumorigenesis. Nature. 456:814–818. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Lin L, Chen YS, Yao YD, Chen JQ, Chen JN,
Huang SY, Zeng YJ, Yao HR, Zeng SH, Fu YS and Song EW: CCL18 from
tumor-associated macrophages promotes angiogenesis in breast
cancer. Oncotarget. 6:34758–34773. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Saharinen P, Bry M and Alitalo K: How do
angiopoietins Tie with vascular endothelial growth factors? Curr
Opin Hematol. 17:198–205. 2010.PubMed/NCBI
|
|
55
|
Chen Y, Song Y, Du W, Gong L, Chang H and
Zou Z: Tumor-associated macrophages: An accomplice in solid tumor
progression. J Biomed Sci. 26:782019. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Mazzieri R, Pucci F, Moi D, Zonari E,
Ranghetti A, Berti A, Politi LS, Gentner B, Brown JL, Naldini L and
De Palma M: Targeting the ANG2/TIE2 axis inhibits tumor growth and
metastasis by impairing angiogenesis and disabling rebounds of
proangiogenic myeloid cells. Cancer Cell. 19:512–526. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Dirkx AE, Oude Egbrink MG, Wagstaff J and
Griffioen AW: Monocyte/macrophage infiltration in tumors:
Modulators of angiogenesis. J Leukoc Biol. 80:1183–1196. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Chaffer CL and Weinberg RA: A perspective
on cancer cell metastasis. Science. 331:1559–1564. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Wyckoff J, Wang W, Lin EY, Wang Y, Pixley
F, Stanley ER, Graf T, Pollard JW, Segall J and Condeelis J: A
paracrine loop between tumor cells and macrophages is required for
tumor cell migration in mammary tumors. Cancer Res. 64:7022–7029.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Gocheva V, Wang HW, Gadea BB, Shree T,
Hunter KE, Garfall AL, Berman T and Joyce JA: IL-4 induces
cathepsin protease activity in tumor-associated macrophages to
promote cancer growth and invasion. Genes Dev. 24:241–255. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
DeNardo DG, Barreto JB, Andreu P, Vasquez
L, Tawfik D, Kolhatkar N and Coussens LM: CD4(+) T cells regulate
pulmonary metastasis of mammary carcinomas by enhancing protumor
properties of macrophages. Cancer Cell. 16:91–102. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Hernandez L, Smirnova T, Kedrin D, Wyckoff
J, Zhu L, Stanley ER, Cox D, Muller WJ, Pollard JW, Van Rooijen N
and Segall JE: The EGF/CSF-1 paracrine invasion loop can be
triggered by heregulin beta1 and CXCL12. Cancer Res. 69:3221–3227.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Kim S, Takahashi H, Lin WW, Descargues P,
Grivennikov S, Kim Y, Luo JL and Karin M: Carcinoma-produced
factors activate myeloid cells through TLR2 to stimulate
metastasis. Nature. 457:102–106. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Yan D, Wang HW, Bowman RL and Joyce JA:
STAT3 and STAT6 signaling pathways synergize to promote cathepsin
secretion from macrophages via IRE1α activation. Cell Rep.
16:2914–2927. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Quintero-Fabián S, Arreola R,
Becerril-Villanueva E, Torres-Romero JC, Arana-Argáez V,
Lara-Riegos J, Ramírez-Camacho MA and Alvarez-Sánchez ME: Role of
matrix metalloproteinases in angiogenesis and cancer. Front Oncol.
9:13702019. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Jiang D, Liang J and Noble PW: Hyaluronan
as an immune regulator in human diseases. Physiol Rev. 91:221–264.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Wang N, Liu W, Zheng Y, Wang S, Yang B, Li
M, Song J, Zhang F, Zhang X, Wang Q and Wang Z: CXCL1 derived from
tumor-associated macrophages promotes breast cancer metastasis via
activating NF-κB/SOX4 signaling. Cell Death Dis. 9:8802018.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Izumi K and Mizokami A: Suppressive role
of androgen/androgen receptor signaling via chemokines on prostate
cancer cells. J Clin Med. 8:3542019. View Article : Google Scholar
|
|
69
|
Wang D, Sun H, Wei J, Cen B and DuBois RN:
CXCL1 is critical for premetastatic niche formation and metastasis
in colorectal cancer. Cancer Res. 13:3655–3665. 2017. View Article : Google Scholar
|
|
70
|
Kawano M, Mabuchi S, Matsumoto Y, Sasano
T, Takahashi R, Kuroda H, Kozasa K, Hashimoto K, Isobe A, Sawada K,
et al: The significance of G-CSF expression and myeloid-derived
suppressor cells in the chemoresistance of uterine cervical cancer.
Sci Rep. 5:182172015. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Rőszer T: Understanding the mysterious M2
macrophage through activation markers and effector mechanisms.
Mediators Inflamm. 2015:8164602015. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Okeke EB and Uzonna JE: The pivotal role
of regulatory T cells in the regulation of innate immune cells.
Front Immunol. 10:6802019. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Hoves S, Ooi CH, Wolter C, Sade H,
Bissinger S, Schmittnaegel M, Ast O, Giusti AM, Wartha K, Runza V,
et al: Rapid activation of tumor-associated macrophages boosts
preexisting tumor immunity. J Exp Med. 215:859–876. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Ma X, Yan W, Zheng H, Du Q, Zhang L, Ban
Y, Li N and Wei F: Regulation of IL-10 and IL-12 production and
function in macrophages and dendritic cells 4. F1000 Faculty
Rev-1465. 2015.
|
|
75
|
Kanamori M, Nakatsukasa H, Okada M, Lu Q
and Yoshimura A: Induced regulatory T Cells: Their development,
stability, and applications. Trends Immunol. 37:803–811. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Chanmee T, Ontong P, Konno K and Itano N:
Tumor-associated macrophages as major players in the tumor
microenvironment. Cancers (Basel). 6:1670–1690. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Zhang J, Shi Z, Xu X, Yu Z and Mi J: The
influence of microenvironment on tumor immunotherapy. FEBS J.
286:4160–4175. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Czystowska-Kuzmicz M, Sosnowska A, Nowis
D, Ramji K, Szajnik M, Chlebowska-Tuz J, Wolinska E, Gaj P, Grazul
M, Pilch Z, et al: Small extracellular vesicles containing
arginase-1 suppress T-cell responses and promote tumor growth in
ovarian carcinoma. Nat Commun. 10:30002019. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Pathria P, Louis TL and Varner JA:
Targeting tumor-associated macrophages in cancer. Trends Immunol.
40:310–327. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Zhang D, Shi R, Xiang W, Kang X, Tang B,
Li C, Gao L, Zhang X, Zhang L, Dai R and Miao H: The Agpat4/LPA
axis in colorectal cancer cells regulates antitumor responses via
p38/p65 signaling in macrophages. Signal Transduct Target Ther.
5:242020. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Vinogradov S, Warren G and Wei X:
Macrophages associated with tumors as potential targets and
therapeutic intermediate. Nanomedicine (Lond). 9:695–707. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Nywening TM, Belt BA, Cullinan DR, Panni
RZ, Han BJ, Sanford DE, Jacobs RC, Ye J, Patel AA, Gillanders WE,
et al: Targeting both tumour-associated CXCR2+
neutrophils and CCR2+ macrophages disrupts myeloid
recruitment and improves chemotherapeutic responses in pancreatic
ductal adenocarcinoma. Gut. 67:1112–1123. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Lee C, Jeong H, Bae Y, Shin K, Kang S, Kim
H, Oh J and Bae H: Targeting of M2-like tumor-associated
macrophages with a melittin-based pro-apoptotic peptide. J
Immunother Cancer. 7:1472019. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Andersen MN, Etzerodt A, Graversen JH,
Holthof LC, Moestrup SK, Hokland M and Møller HJ: STAT3 inhibition
specifically in human monocytes and macrophages by CD163-targeted
corosolic acid-containing liposomes. Cancer Immunol Immunother.
68:489–502. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Wanderley CW, Colón DF, Luiz JPM, Oliveira
FF, Viacava PR, Leite CA, Pereira JA, Silva CM, Silva CR, Silva RL,
et al: Paclitaxel reduces tumor growth by reprogramming
tumor-associated macrophages to an M1 profile in a TLR4-dependent
manner. Cancer Res. 78:5891–5900. 2018.PubMed/NCBI
|
|
86
|
Locatelli SL, Careddu G, Serio S, Consonni
FM, Maeda A, Viswanadha S, Vakkalanka S, Castagna L, Santoro A,
Allavena P, et al: Targeting cancer cells and tumor
microenvironment in preclinical and clinical models of Hodgkin
lymphoma using the dual PI3Kδ/γ inhibitor RP6530. Clin Cancer Res.
25:1098–1112. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Tan HY, Wang N, Man K, Tsao SW, Che CM and
Feng Y: Autophagy-induced RelB/p52 activation mediates
tumour-associated macrophage repolarisation and suppression of
hepatocellular carcinoma by natural compound baicalin. Cell Death
Dis. 6:e19422015. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Medler TR, Murugan D, Horton W, Kumar S,
Cotechini T, Forsyth AM, Leyshock P, Leitenberger JJ, Kulesz-Martin
M, Margolin AA, et al: Complement C5a fosters squamous
carcinogenesis and limits t cell response to chemotherapy. Cancer
Cell. 34:561–578.e6. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Guerriero JL, Sotayo A, Ponichtera HE,
Castrillon JA, Pourzia AL, Schad S, Johnson SF, Carrasco RD, Lazo
S, Bronson RT, et al: Class IIa HDAC inhibition reduces breast
tumours and metastases through anti-tumour macrophages. Nature.
543:428–432. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Zhang W, Wang M, Tang W, Wen R, Zhou S,
Lee C, Wang H, Jiang W, Delahunty IM, Zhen Z, et al:
Nanoparticle-laden macrophages for tumor-tropic drug delivery. Adv
Mater. 30:e18055572018. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Choi J, Kim HY, Ju EJ, Jung J, Park J,
Chung HK, Lee JS, Lee JS, Park HJ, Song SY, et al: Use of
macrophages to deliver therapeutic and imaging contrast agents to
tumors. Biomaterials. 33:4195–4203. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Opperman KS, Vandyke K, Clark KC, Coulter
EA, Hewett DR, Mrozik KM, Schwarz N, Evdokiou A, Croucher PI,
Psaltis PJ, et al: Clodronate-liposome mediated macrophage
depletion abrogates multiple myeloma tumor establishment in vivo.
Neoplasia. 21:777–787. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Zhang F, Parayath NN, Ene CI, Stephan SB,
Koehne AL, Coon ME, Holland EC and Stephan MT: Genetic programming
of macrophages to perform anti-tumor functions using targeted mRNA
nanocarriers. Nat Commun. 10:39742019. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Buhtoiarov IN, Sondel PM, Wigginton JM,
Buhtoiarova TN, Yanke EM, Mahvi DA and Rakhmilevich AL: Anti-tumour
synergy of cytotoxic chemotherapy and anti-CD40 plus CpG-ODN
immunotherapy through repolarization of tumour-associated
macrophages. Immunology. 132:226–239. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Di Caro G, Cortese N, Castino GF, Grizzi
F, Gavazzi F, Ridolfi C, Capretti G, Mineri R, Todoric J, Zerbi A,
et al: Dual prognostic significance of tumour-associated
macrophages in human pancreatic adenocarcinoma treated or untreated
with chemotherapy. Gut. 65:1710–1720. 2016. View Article : Google Scholar : PubMed/NCBI
|














