Introduction
Gastrointestinal stromal tumors (GISTs) are the most
common pathologic type of mesenchymal tumors in the digestive tract
(1). Most patients face the
challenges of early hematogenous metastasis, postoperative
recurrence and poor clinical outcomes (2). Worldwide, a total of ~20% of patients
are primarily resistant to imatinib mesylate (IM), and 50% of
patients exhibit drug resistance within 18–24 months, which leads
to disease progression and even death (3). A total of 90% of GISTs have a
gain-of-function mutation in KIT proto-oncogene, receptor tyrosine
kinase (KIT) or platelet-derived growth factor receptor α (PDGFRA),
which belong to the receptor tyrosine kinase family and can
self-activate without any tyrosine kinase ligands (4,5). Another
10% of GISTs lack the aforementioned mutation and are referred to
as wild-type GISTs. Succinate dehydrogenase dysfunction occurs in
mitochondria (6). Mitochondrial
dysfunction is considered to be closely associated with tumor
occurrence, development and drug resistance (7,8).
Therefore, the role of mitochondria in tumor formation has become
the frontier of current research.
Mitochondrial Tu translation elongation factor
(TUFM) is the most highly expressed protein in mitochondria
(9). This protein initiates the
polypeptide chain extension cycle and is involved in almost all
mitochondrial protein translation (10). Several studies have demonstrated that
TUFM is highly expressed in gastric cancer, esophageal cancer,
GISTs and other gastrointestinal tumors (11–15).
Related proteomics studies have observed that TUFM expression is
associated with the risk classification and prognosis of GISTs
(13). Therefore, it is reasonable
to hypothesize that TUFM may be related to the occurrence, invasion
and metastasis of GISTs. However, the role of TUFM in GISTs is
still unclear.
IM is the most commonly used target drug for
patients with GIST. IM extends the median survival of patients who
are diagnosed at late stages and thus cannot undergo surgical
resection (16). In the present
study, the relationship among TUFM, GIST-T1 cells and IM-resistant
cells was studied in vitro. By inhibiting TUFM expression in
GIST-T1 cells and IM-resistant cells, novel outlooks for GIST
treatment can be provided.
Materials and methods
Cell lines and culture
The GIST-T1 (passage 20) mutant-type cell line has a
base deletion of 57 bp in KIT exon 11 (PMC GIST01C; Cosmo
Bio Co., Ltd.) (17). GIST-IR cells
(passage 23) were induced by treating GIST-T1 cells with 50 µg/µl
IM (Glivec). GIST-IR drug-resistant cell lines were established by
the gradient method, as previously described (18). No secondary mutations were observed
in KIT/PDGFR exon sequencing analysis (19). GIST-T1 and GIST-IR cells were
cultured in RPMI 1640 medium (Gibco; Thermo Fisher Scientific,
Inc.) with 15% fetal bovine serum (TransGen Biotech Co., Ltd.) in
an incubator at 37°C with 5% CO2 without
antibiotics.
TUFM silencing plasmids and electric
transfection
pcDNA3.1-EGFP-TUFM-shRNA plasmids were constructed
and designed with Primer Premier 5.0 (http://www.premierbiosoft.com/primerdesign) by using
the TUFM fragment of the human gene acquired from NCBI GenBank
(https://www.ncbi.nlm.nih.gov/)(source,
Oryctolagus cuniculus). The primer sequences were as
follows: Forward, 5′-GCAAGCTTGTGGACATCTTCCAGGAGTA-3′ and reverse,
5′-GCGAATTCCTTTGGTCTGCATTCACATT-3′.
Construction and identification of the
pcDNA3.1-EGFR-TUFM-shRNA were performed by Invitrogen; Thermo
Fisher Scientific, Inc. The pcDNA3.1 vector, EcoRI and
HindIII restriction enzymes, and T7-DNA ligase were
purchased from Invitrogen; Thermo Fisher Scientific, Inc. The
length of the target gene was 225 bp, and the results were
confirmed by sequencing (20).
The experiment was divided into three groups:
pcDNA3.1-EGFP-TUFM-shRNA plasmid (1 µg/µl) was transferred in the
TUFM-shRNA group (TUFM shRNA), pcDNA3.1 vector was transfected in
the negative control group (Control) and no transfection was
conducted in the blank group (Blank). Electric transfection
equipment (CTX-1500A; Celetrix Biotechnology Co., Ltd.) was used
for electroporation according to the manufacturer's protocols.
Briefly, 2×106 cells were harvested and resuspended in
120 µl electrotransfer solution (Celetrix Biotechnology Co., Ltd.)
and 2 µl plasmid (1 µg/µl). The cell mixture was immediately added
to the electric tube. The setting procedure of the system was as
follows: 570 volts, 30 msec, 1 time. After electroporation, the
cell mixture was divided into three parts that were resuspended in
RPMI 1640 medium containing 15% FBS and seeded in a 6-well plate
for 24 h at 37°C before subsequent experiments.
Reverse transcription quantitative
(RT-q) PCR
Total RNA was extracted by TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturers' protocol. TaqMan miRNA RT-kit (Invitrogen; Thermo
Fisher Scientific, Inc.) was used to convert total miRNAs into
corresponding cDNAs. The RT reaction conditions were as follows:
37°C for 15 min, 85°C for 5 sec and 4°C for 5 min. Subsequently,
cDNA was used as template for PCR amplification by SYBR™ Green
Master Mix (Thermo Fisher Scientific, Inc.) according to the
manufacturers' instructions. The sequences of the primers were as
follows: TUFM, forward, 5′-ATCCGGGAGCTGCTCACCGA-3′, reverse,
5′-ATGCTGTGGACACTTACATA-3′; and GAPDH, forward,
5′-AGGTCGGTGTGAACGGATTTG-3′ and reverse 5′-GGGGTCGTTGATGGCAACA-3′.
GAPDH was used as the reference gene. The thermocycling conditions
for qPCR were as follows: 94°C for 2 min for 1 cycle, 95°C for 40
sec, 60°C for 35 sec and 72°C for 1 min for 40 cycles. The
expression level of TUFM was analyzed using the 2−ΔΔCq
method (21).
Western blotting
Cells were harvested when confluence reached 90% and
total protein was extracted using Total Protein Extraction Kit
(cat. no. P0033; Beyotime Institute of Biotechnology) according to
the manufacturers' protocol. Total protein concentration was
determined by BCA method (22).
Proteins (20 µg) were separated by 10% SDS-PAGE (cat. no. P0052A;
Beyotime Institute of Biotechnology) and transferred onto PVDF
membranes (cat. no. FFP24; Beyotime Institute of Biotechnology).
Membranes were blocked using Western blocking solution (cat. no.
P0023B; Beyotime Institute of Biotechnology) at 25°C for 60 min and
incubated with primary antibodies against TUFM (cat. no. ab155328;
Abcam; 1:500) and GAPDH (cat. no. ab8254; Abcam; 1:500) at 25°C for
120 min. Membranes were then incubated with Goat polyclonal
secondary antibody to Mouse IgG-H&L (Alexa Fluor®
488; cat. no. ab150113; Abcam; 1:5,000) at 25°C for 60 min.
Proteins were detected using DAB Horseradish Peroxidase Color
Development Kit (cat. no. P00203; Beyotime Institute of
Biotechnology). The data were analyzed via densitometry using
ImageJ 1.52n software (National Institutes of Health).
Cell morphology and fluorescence
assessment
GIST-T1 and GIST-IR cells were plated in 6-well
dishes at a density of 1×105 cells/ml. Cell morphology
and fluorescence were observed under an inverted fluorescence
microscope (magnifications, ×40 and ×100).
Cell proliferation and viability
assays
Cell proliferation was determined by manual cell
counting. Cells were seeded on 24-well plates at a density of
5×104 cells/ml. Live cells were counted every 24 h, and
the average of three wells for 5 days was used to draw cell growth
and inhibition curves. GIST-TI and GIST-IR cells were plated into
96-well plates at a density of 5×103 cells/well in
triplicate. Subsequently, the cells were transfected with
TUFM-shRNA plasmid and pcDNA3.1 vector. Cell viability was
evaluated using CCK-8 assays (Dojindo Molecular Technologies, Inc.)
according to the manufacturer's protocol. The absorbance was
determined at 450 nm.
Wound healing and Transwell
assays
GIST-T1 and GIST-IR cells were plated in 6-well
plates at a density of 1×104 cells/ml and cultured in
RPMI-1640 medium with 15% fetal bovine serum at 37°C with 5%
CO2. After 24 h, when the cells reached confluence in
90%, a scratch was made in the middle of the plate, and the
scratched cells were washed twice with PBS. The cells were cultured
in RPMI-1640 medium with 2.5% fetal bovine serum at 37°C with 5%
CO2. Images of the samples were captured at 0, 24 and 48
h, and cell morphology was observed under an inverted fluorescence
microscope (magnification, ×40). ImageJ 1.52n software was used to
analyze and quantify the range of cell-free regions in the
images.
For Transwell assays, TUFM shRNA and control groups
of both GIST-T1 and GIST-IR cells were inoculated at a density of
5×104 cells/ml in 8 µm-pore Transwell chambers
(Corning). RPMI 1640 medium containing 2.5% FBS was added to the
upper chamber whereas medium containing 15% FBS was added to the
lower chamber. The cells were incubated for 24 h at 37°C with 5%
CO2, and cells in the upper chamber were stained with
eosin at 25°C for 3 min. The cells in five random fields were
counted under an inverted fluorescence microscope (magnification,
×100).
Cell cycle determination
TUFM shRNA and control groups of GIST-T1 and GIST-IR
cells in the logarithmic phase were subjected to trypsin digestion
and centrifugal (4°C, 250 × g, 1 min) precipitation. Subsequently,
1×106 cells/ml cells were fixed with 75% ice-cold
ethanol overnight at 4°C. The cells were centrifuged at 1,000 × g
for 3 min and the supernatant was discarded. The cells were
resuspended in propidium iodide (PI; 10 µg/ml) and RNase (5 µg/ml;
PI Staining kit, Sangon Biotech Co., Ltd.), washed twice with PBS
and incubated at 4°C for 30 min. Subsequently, the supernatants
were discarded after centrifugation (4°C, 1,000 × g, 3 min), and
the cells were washed once with PBS and transferred into a flow
cytometer centrifuge tube to be measured on the machine. Flow
cytometric analysis of 2×104 cells from each sample was
performed on FACSort flow cytometer (BD Biosciences, Inc). FlowJo
VX (BD Biosciences, Inc.) was used for analysis and late apoptosis
assessment.
Statistical analysis
The results are represented as the mean ± standard
deviation of three independent repeats. Unpaired Student's t-test
was performed to compare differences between two groups and one-way
ANOVA followed by Ruhey's post hoc test was performed to compare
differences among multiple groups. Data were analyzed using SPSS
20.0 software (IBM Corp.) or GraphPad Prism 6 (GraphPad Software,
Inc.). P<0.05 was considered to indicate a statistically
significant difference.
Results
TUFM expression is markedly decreased
after transfection of GIST-T1 cells with TUFM shRNA plasmid
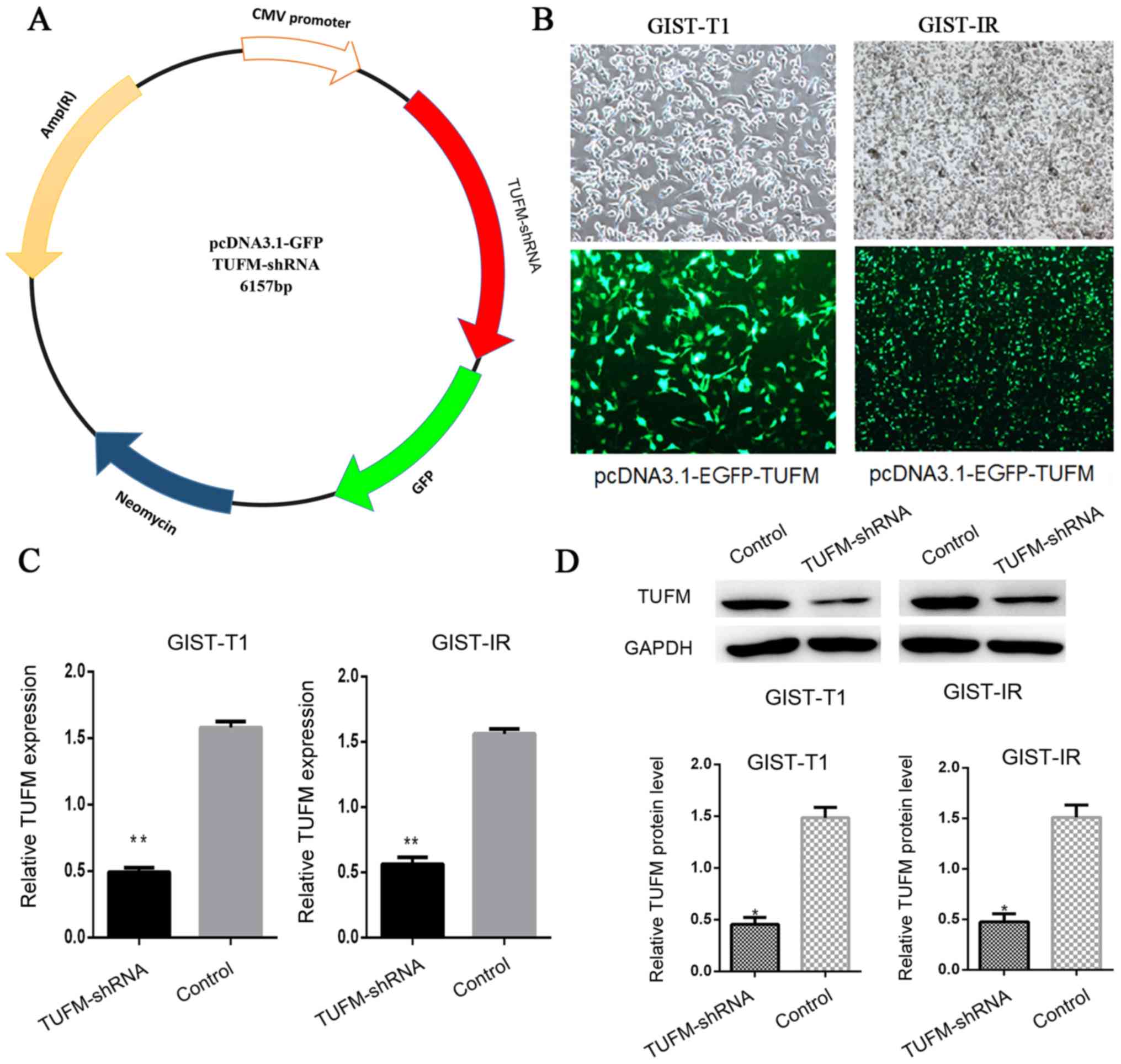
The TUFM shRNA plasmid was constructed to knockdown
TUFM protein expression in cells (Fig.
1A). After the TUFM shRNA plasmid was transfected into GIST-T1
and GIST-IR cells, EGFP-expressing cells were observed and counted
by fluorescence microscopy. The transfection efficiency was ~75%
and no significant cell death was observed (Fig. 1B). Reverse transcription-quantitative
(RT-q)PCR and western blot analyses demonstrated that the
expression levels of TUFM were significantly reduced in the TUFM
shRNA group compared with those in the control group (Fig. 1C and D).
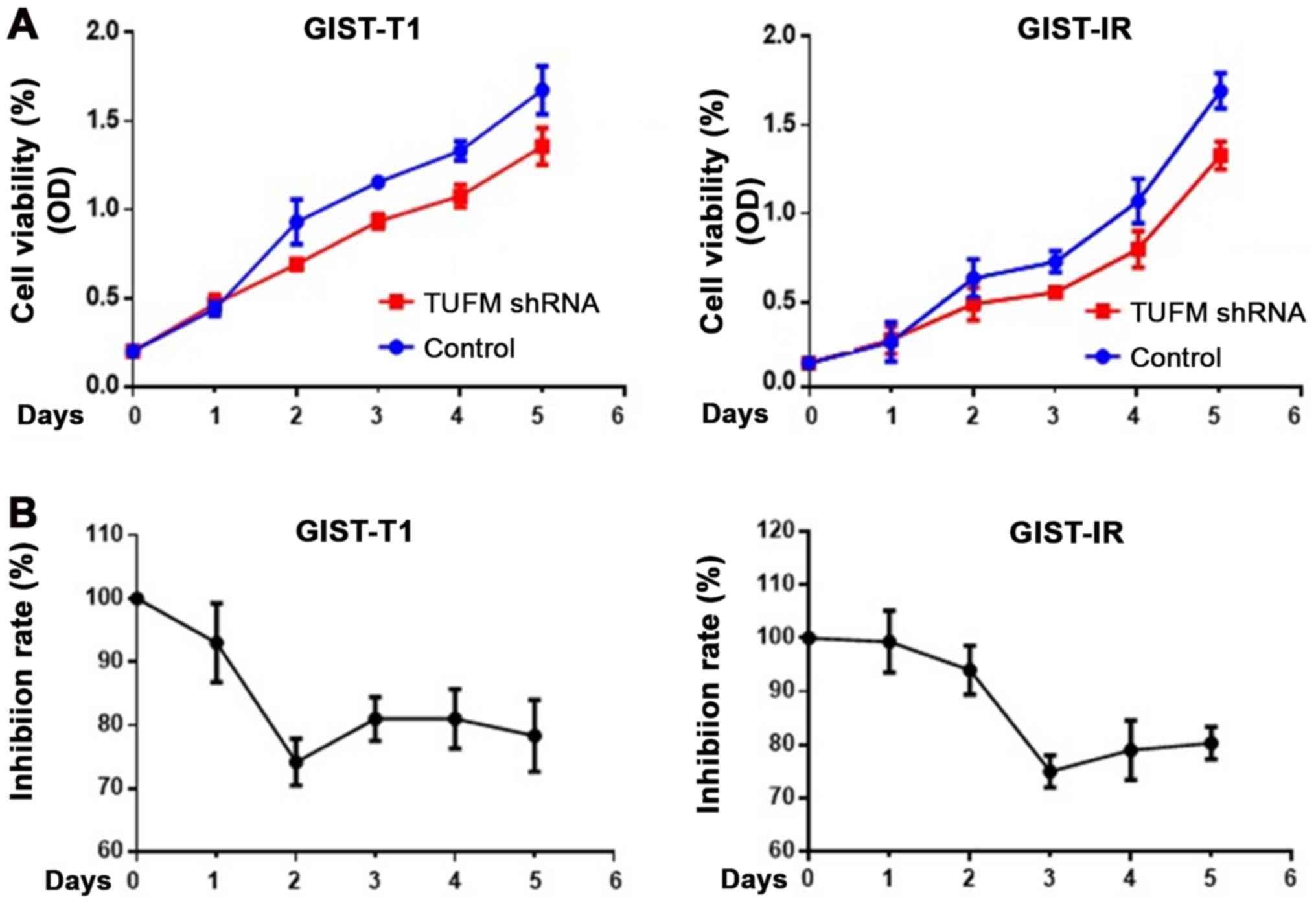
TUFM-knockdown inhibits the viability
and proliferation of GIST-T1 and GIST-IR cells
After TUFM was knocked down, cell viability was
decreased in GIST-T1 and GIST-IR cells (Fig. 2A). Cells were counted for 5
consecutive days to determine cell viability. In addition, CCK-8
assays were used to detect the proliferation rate of GIST-T1 and
GIST-IR cells. The results demonstrated that TUFM silencing
inhibited proliferation of GIST-T1 cells, reaching its maximal
effect at 48 h after transfection, and GIST-IR cells, reaching its
maximal effect at 72 h after transfection (Fig. 2).
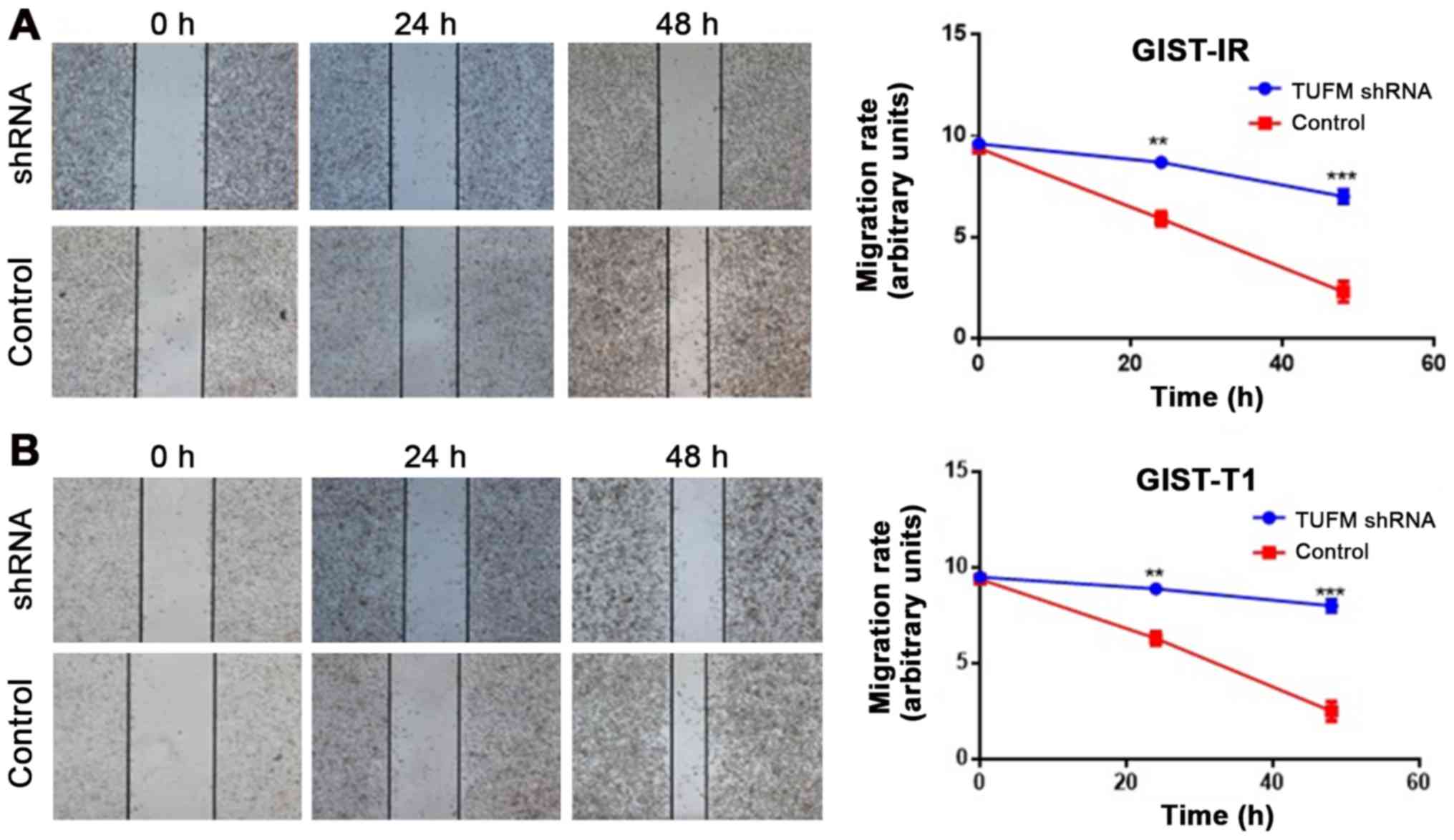
TUFM silencing suppresses the
migration of GIST-T1 and GIST-IR cells
Wound healing assays demonstrated that, compared
with the control groups, TUFM silencing significantly decreased the
migratory rate of GIST-T1 and GIST-IR cells at 48 h after
transfection (Fig. 3A and B).
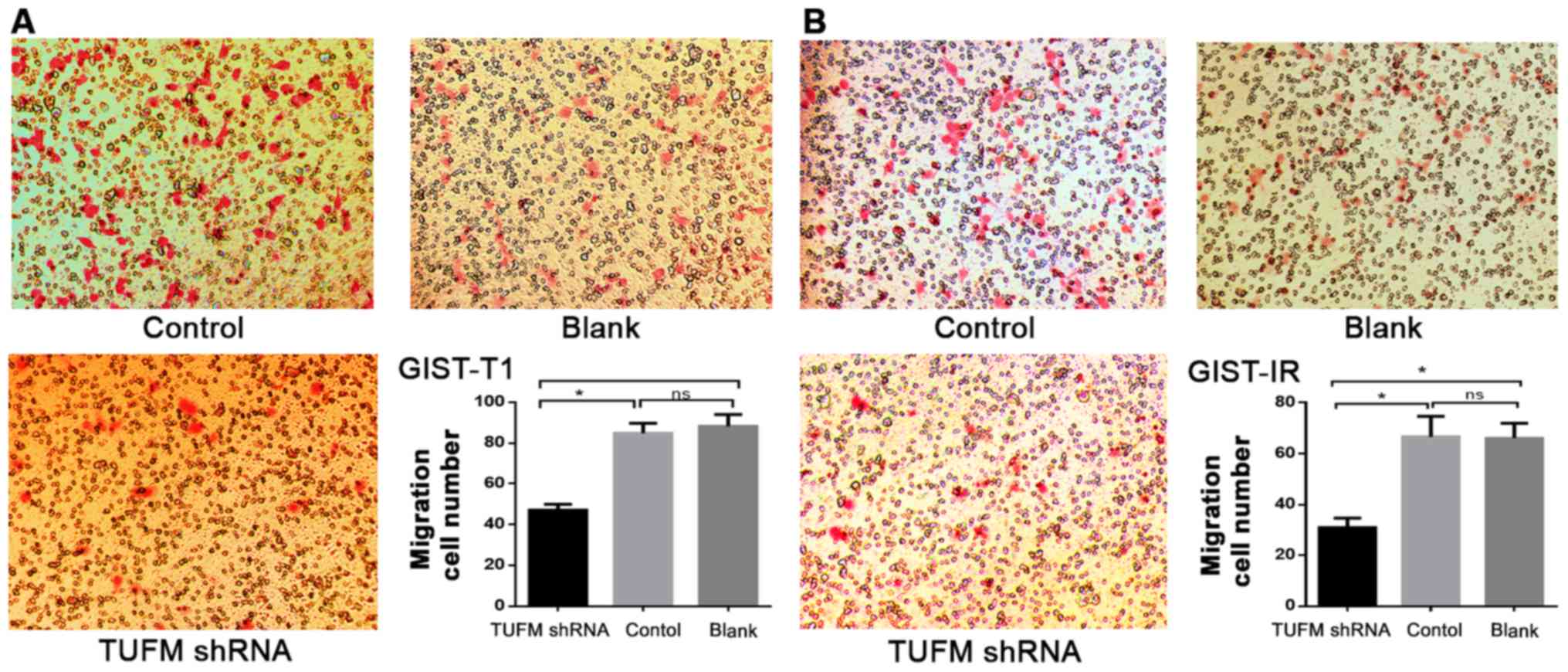
Additionally, Transwell migration assays demonstrated that,
compared with those in the control and blank groups, the number of
GIST-T1 and GIST-IR cells on the underside of the chamber membrane
was significantly decreased after TUFM silencing (P<0.05;
Fig. 4). In addition no significant
differences were observed between the control and blank groups in
GIST-T1 and GIST-IR cells (P>0.05; Fig. 4).
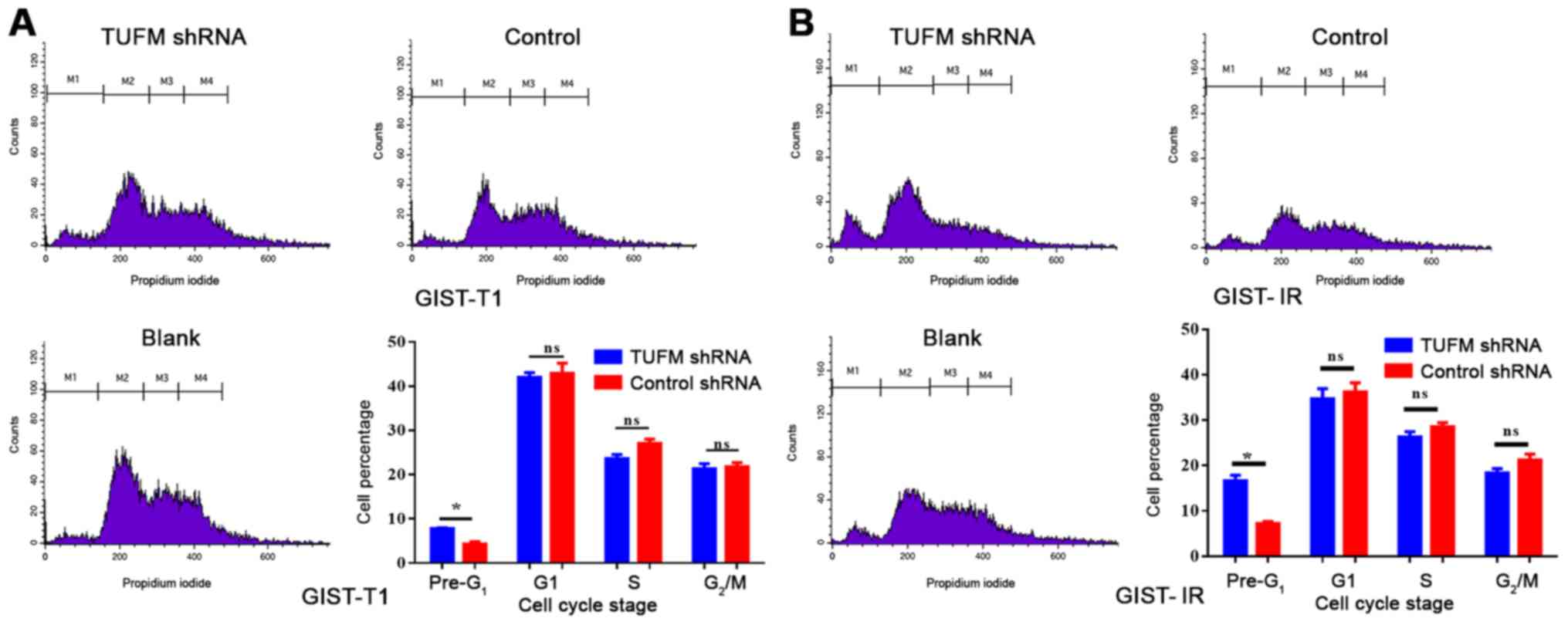
TUFM affects late apoptosis and the
cell cycle of GIST-T1 and GIST-IR cells
Cell cycle and late apoptosis were detected by flow
cytometry. The results demonstrated that the proportion of GIST-T1
and GIST-IR cells in the pre-G1 phase was significantly
higher in the TUFM shRNA group compared with in the GIST-T1 and
GIST-IR cells in the control group (Fig.
5). By contrast, no significant differences were observed in
the proportions of GIST-T1 and GIST-IR cells in the G1,
S and G2/M phases between the two groups (Fig. 5).
Discussion
In 20 years of research, the study of GISTs has
gradually advanced. A variety of molecular pathological mechanisms
are involved in the occurrence, development and drug resistance of
GISTs (23). Currently, GIST has one
of the highest mortality rates among tumors worldwide (22). Genomic and proteomic studies of GIST
tissue have revealed a variety of tumor-related molecules involved
in the occurrence, development and drug resistance of GISTs
(24). The high expression levels of
TUFM in tumor tissues suggest that it may serve an important role
in the process of tumorigenesis and development, and its
association with KIT/PDGFRA remains unclear (25). The present study used an
electroporation method to transfect a TUFM-silencing plasmid, which
is considered to be the most efficient and convenient method of
cell transfection in vitro (26). Combined with RNA interference
technology, the function of TUFM in tumor cells can be evaluated
quickly and can provide an effective platform for further study of
related signaling pathways.
The results of cell counting and CCK-8 assays
demonstrated that cell proliferation was significantly inhibited
after silencing of the TUFM gene in GIST-T1 and GIST-IR cells. This
is consistent with the results of Jhas et al (27), which demonstrated that cell
proliferation was significantly inhibited by silencing of the TUFM
gene in TEX leukemia cells. This effect is considered to be caused
by changes in cell function related to reductions in oxygen
consumption and mitochondrial membrane potential (28). A study by Sajid et al
(29) demonstrated that inhibiting
TUFM phosphorylation through drug treatment inhibited the protein
synthesis and growth of cells. In addition, Ravn et al
(30) observed that TUFM and other
translation protein deficiencies in patients with clinical
mitochondrial protein translation system defects cause lethal
effects during the early neonatal period, whereas, in other
patients, TUFM causes serious defects and oxidation-respiration
chain enzyme dysfunction in the liver and muscle system. The
importance of mitochondrial function in maintaining cell activity
is clear. When TUFM is silenced, the synthesis of mitochondrial
oxidative respiratory chain complex proteins becomes inhibited or
dysfunctional, leading to decreased mitochondrial membrane
potential and disrupted ATP synthesis. Subsequently, the disruption
of energy supply leads to a lack of energy in the cells, and cell
proliferation is inhibited (31).
This may be the main reason why the in vitro silencing of
TUFM inhibits cell proliferation. Therefore, the hypothesis that
the decrease in mitochondrial function after the silencing of TUFM
results in a decrease in cell viability and cell proliferation is
valid.
In proteomic analysis of GISTs, the expression
levels of TUFM in the high-risk group are higher than those in the
low-risk group (13). Furthermore,
GISTs in the high-risk group tend to have more aggressive tumor
cells and worse prognosis (32). The
results of the cell migration experiments performed in the present
study demonstrated that the lateral and vertical migration of cells
was decreased significantly following TUFM-knockdown. Therefore, it
was proposed that interference with TUFM expression may inhibit the
synthesis of adhesion proteins and lead to a decline in cell
migration ability. In a study by Hashim et al (33), Mediterranean patients with colon
cancer were less affected by invasion and metastasis. Notably,
epidemiological analysis observed that the Mediterranean population
consumed more olive oil than individuals in other areas. Through
high-throughput microarray data analysis, it was demonstrated that
olive oil may inhibit the function of TUFM and other related genes
to inhibit colon invasion. Other studies have reported that olive
oil may inhibit TUFM function by inhibiting integrin-α2 and
integrin-α6 expression in tumor cells (34). As a cell surface receptor, integrin
acts as a bridge for cell-cell interactions, and cell and
extracellular matrix interactions. TUFM expression is positively
associated with E-cadherin in lung cancer tissue and serves an
important role in the epithelium during interstitial invasion and
metastasis of lung cancer, possibly by recruiting β-cadherin to
regulate lung cancer cell invasion and metastasis through protein
kinase AMP-activated catalytic subunit α 2-GSK3β signaling
(35). Additionally, abnormal
mitochondrial energy metabolism, mitochondrial dysfunction, and
inadequate ATP energy supply may be reasons for the cell migration
inhibition caused by TUFM silencing.
The results of the cell cycle experiments performed
in the present study demonstrated that following TUFM silencing in
GIST-T1 and GIST-IR cells, there were no significant differences in
the cell cycle distribution among the silenced, negative control
and blank groups; however, the number of cells in the early
G1 phase was significantly increased. This early
G1 phase cell population is often considered to be
associated with reduced PI staining, intracellular DNA degradation
and characteristics of late apoptotic cells (36). These results suggested that TUFM
silencing may promote late apoptosis. Lei et al (37) studied the synergistic effects of TUFM
and NLRX1 (NLR family member X1), and reported that TUFM recruits
autophagosomes, such as ATG5-ATG12, in mitochondria and promotes
the activation of autophagy signaling pathways, leading to
autophagy in mitochondria, and thus altering the dynamic
intracellular mitochondria balance. Furthermore, Rakovic et
al (38) observed that in a
Parkinsonian pathological model, TUFM may be involved in the PTEN
induced kinase 1/Parkin signaling pathway to promote autophagy in
functionally defective mitochondria. It is now hypothesized that
TUFM is involved in initiating mitochondrial autophagy and
scavenging damaged mitochondria, thereby inhibiting tumor cell
apoptosis induced by mitochondrial damage signaling pathways
(39). Therefore, the results of the
present study, which demonstrated that late apoptotic cells were
increased in the TUFM-silenced GIST-T1 cell line, are consistent
with the aforementioned research revealing inhibited mitochondrial
autophagy and damaged mitochondrial scavenging in TUFM stable
knockout mice and TUFM-silenced 293T cells. These changes lead to
considerable mitochondrial disintegration factor release and
increased apoptosis (40). TUFM
silencing directly induces mitochondrial protein synthesis
inhibition, membrane dysfunction and mitochondrial structural
dysfunction, which eventually leads to mitochondrial damage or
disintegration, which is an important contributor to tumor cell
apoptosis (41).
There are a number of challenges in the treatment of
GISTs. TUFM may be involved in numerous tumor cell processes. The
relationship between TUFM expression and the prognosis of patients
with GIST, the relationship between TUFM and cell adhesion
molecules, and the signaling pathway of TUFM require further study.
In addition, as the present study was an in vitro study,
further in vivo evaluation of TUFM function is required.
The present study investigated the effects of TUFM
on the growth, migration and cell cycle of GIST cells in
vitro. TUFM inhibited the growth and migration of GIST cell
lines and promoted the apoptosis of tumor cells. These results
suggested that TUFM may serve a role in reducing postoperative
recurrence and metastasis in patients with GIST. Therefore, TUFM
may become an effective target for inhibiting early hematogenous
metastasis, postoperative recurrence and metastasis in patients
with GIST, even in IM-resistant patients.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Chinese
National Key Clinical College (grant no. 2492012), Fujian
Provincial Health and Family Planning Key Personnel Training
Program (grant no. 1033291) and Quanzhou City Science &
Technology Program of China (grant no. 2019N071S).
Availability of data and materials
The datasets used or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YZ, XW and SZ made substantial contributions to the
conception and design of the study. XW, HS and GS performed the
experiments. XW, HS and GL contributed significantly to data
analysis. XW, YZ and SZ helped performing the analysis with
constructive discussions. XW drafted the manuscript. YZ, XW gave
final approval of the version to be published. YZ and XW agreed to
be accountable for all aspects of the work in ensuring that
questions related to the accuracy and integrity of any part of the
work were appropriately investigated and resolved. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Mazur MT and Clark HB: Gastric stromal
tumors. Reappraisal of histogenesis. Am J Surg Pathol. 7:507–519.
1983. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Roggin KK and Posner MC: Modern treatment
of gastric gastrointestinal stromal tumors. World J Gastroenterol.
18:6720–6728. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Søreide K, Sandvik OM, Søreide JA, Giljaca
V, Jureckova A and Bulusu VS: Global epidemiology of
gastrointestinal stromal tumors (GIST): A systematic review of
population-based cohort studies. Cancer Epidemiol. 40:39–46. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hirota S, Isozaki K, Moriyama Y, Hashimoto
K, Nishida T, Ishiguro S, Kawano K, Hanada M, Kurata A, Takeda M,
et al: Gain-of-function mutations of KIT in human gastrointestinal
stromal tumors. Science. 279:577–580. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hayashi Y, Bardsley MR, Toyomasu Y,
Taguchi T, Rubin BP, Carter M, Ramachandran A and Ordog T: Abstract
1035: Crenolanib, a highly selective platelet-derived growth factor
receptor/(PDGFRA/B) tyrosine kinase inhibitor, destabilizes ETV1
protein in KIT-mutant gastrointestinal stromal tumor (GIST) cells.
Cancer Res. 73 (Suppl 8):S10352013.
|
|
6
|
Gill AJ: Succinate dehydrogenase (SDH) and
mitochondrial driven neoplasia. Pathology. 44:285–292. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yang WL, Addona T, Nair DG, Qi L and
Ravikumar TS: Apoptosis induced by cryo-injury in human colorectal
cancer cells is associated with mitochondrial dysfunction. Int J
Cancer. 103:360–369. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ma L, Wang R, Duan H, Nan Y, Wang Q and
Jin F: Mitochondrial dysfunction rather than mtDNA sequence
mutation is responsible for the multi-drug resistance of small cell
lung cancer. Oncol Rep. 34:3238–3246. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shah ZH, Migliosi V, Miller SCM, Wang A,
Friedman TB and Jacobs HT: Chromosomal locations of three human
nuclear genes (RPSM12, TUFM, and AFG3L1) specifying putative
components of the mitochondrial gene expression apparatus.
Genomics. 48:384–388. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang S, Wei AC, Fu Z, Ruker J, Foster BD,
O'Rourke B and Van Eyk JE: Abstract 16664: Cyclophilin D binds to
tufm and suppresses mitochondrial translation. Circulation.
130:A166642014.
|
|
11
|
Zhang J, Huang JY, Chen YN, Yuan F, Zhang
H, Yan FH, Wang MJ, Wang G, Su M, Lu G, et al: Erratum: Whole
genome and transcriptome sequencing of matched primary and
peritoneal metastatic gastric carcinoma. Sci Rep. 5:137502015.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hou J, Liao LD, Xie YM, Zeng FM, Ji X,
Chen B, Li LY, Zhu MX, Yang CX, Zhao Q, et al: DACT2 is a candidate
tumor suppressor and prognostic marker in esophageal squamous cell
carcinoma. Cancer Prev Res (Phila). 6:791–800. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen DB, Jiang WZ, Hong CY, Wang QL, Hong
ZP, Shi LP and Lin CP: Comparative proteomics study of KIT positive
gastrointestinal stromal tumors based on risk stratification
differences. Chinese J Exper Surg. 34:217–220. 2017.
|
|
14
|
Shi H, Hayes M, Kirana C, Miller R,
Keating J, Macartney-Coxson D and Stubbs R: TUFM is a potential new
prognostic indicator for colorectal carcinoma. Pathology.
44:506–512. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Staab CA, Ceder R, Jägerbrink T, Nilsson
JA, Roberg K, Jörnvall H, Höög JO and Grafström RC: Bioinformatics
processing of protein and transcript profiles of normal and
transformed cell lines indicates functional impairment of
transcriptional regulators in buccal carcinoma. J Proteome Res.
6:3705–3717. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bümming P, Andersson J, Meis-Kindblom JM,
Klingenstierna H, Engström K, Stierner U, Wängberg B, Jansson S,
Ahlman H, Kindblom LG and Nilsson B: Neoadjuvant, adjuvant and
palliative treatment of gastrointestinal stromal tumors (GIST) with
imatinib: A center-based study of 17 patients. Br J Cancer.
89:460–464. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Taguchi T, Sonobe H, Toyonaga SI, Yamasaki
I, Shuin T, Takano A, Araki K, Akimaru K and Yuri K: Conventional
and molecular cytogenetic characterization of a new human cell
line, GIST-T1, established from gastrointestinal stromal tumor. Lab
Invest. 82:663–665. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hiroyuki S, Kazumasa F, Yoshiro S, et al:
Establishment of imatinib-resistant GIST cell lines. PLoS One.
2014.
|
|
19
|
Isakov O, Perrone M and Shomron N: Exome
sequencing analysis: A guide to disease variant detection. Methods
Mol Biol. 1038:137–158. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sanger F and Coulson AR: The use of thin
acrylamide gels for DNA sequencing. FEBS Lett. 87:107–110. 1978.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Pfaffl MW: A new mathematical model for
relative quantification in realtime RT-PCR. Nucleic Acids Res.
29:e452001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Smith PK, Krohn RI, Hermanson GT, Mallia
AK, Gartner FH, Provenzano MD, Fujimoto EK, Goeke NM, Olson BJ and
Klenk DC: Measurment of protein using bicinchoninic acid. Anal
Biochem. 150:76–85. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lasota J and Miettinen M: Clinical
significance of oncogenic KIT and PDGFRA mutations in
gastrointestinal stromal tumors. Histopathology. 53:245–266. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Coe TM, Fero KE, Fanta PT, Mallory RJ,
Tang CM, Murphy JD and Sicklick JK: Population-based epidemiology
and mortality of small malignant gastrointestinal stromal tumors in
the USA. J Gastrointestinal Surg. 6:1132–1140. 2016. View Article : Google Scholar
|
|
25
|
Wilkening A, Rüb C, Sylvester M and Voos
W: Analysis of heat-induced protein aggregation in human
mitochondria. J Biol Chem. 293:11537–11552. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Pavlin D, Tozon N, Sersa G, Pogacnik A and
Cemazar M: Efficient electrotransfection into canine muscle.
Technol Cancer Res Treat. 7:45–54. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jhas B, Sriskanthadevan S, Skrtic M,
Sukhai MA, Voisin V, Jitkova Y, Gronda M, Hurren R, Laister RC,
Bader GD, et al: Metabolic adaptation to chronic inhibition of
mitochondrial protein synthesis in acute myeloid leukemia cells.
PLoS One. 8:e583672013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hershkovitz T, Kurolap A, Gonzaga-Jauregui
C, Paperna T, Mory A, Wolf SE; Regeneron Genetics Center, ; Overton
JD, Shuldiner AR, Saada A, et al: A novel TUFM homozygous variant
in a child with mitochondrial cardiomyopathy expands the phenotype
of combined oxidative phosphorylation deficiency 4. J Hum Genet.
64:589–595. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sajid A, Arora G, Gupta M, Singhal A,
Chakraborty K, Nandicoori VK and Singh Y: Interaction of
mycobacterium tuberculosis elongation factor Tu with GTP is
regulated by phosphorylation. J Bacteriol. 193:5347–5358. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ravn K, Schönewolf-Greulich B, Hansen RM,
Bohr AH, Duno M, Wibrand F and Ostergaard E: Neonatal mitochondrial
hepatoencephalopathy caused by novel GFM1 mutations. Mol Genet
Metab Rep. 3:5–10. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liu J, Fang H, Chi Z, Wu Z, Wei D, Mo D,
Niu K, Balajee AS, Hei TK, Nie L and Zhao Y: XPD localizes in
mitochondria and protects the mitochondrial genome from oxidative
DNA damage. Nuclc Acids Res. 43:5476–5488. 2015. View Article : Google Scholar
|
|
32
|
Xi HQ, Zhang KC, Li JY, Cui JX, Zhao P and
Chen L: Expression and clinicopathologic significance of TUFM and
p53 for the normal-adenoma-carcinoma sequence in colorectal
epithelia. World J Surg Oncol. 15:902017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hashim YZHY, Worthington J, Allsopp P,
Ternan NG, Brown EM, McCann MJ, Rowland IR, Esposto S, Servili M
and Gill CIR: Virgin olive oil phenolics extract inhibit invasion
of ht115 human colon cancer cells in vitro and in vivo. Food Funct.
5:1513–1519. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhang L, Xiao K, Zhao X, Sun X, Zhang J,
Wang X, Zhu Y and Zhang X: Quantitative proteomics reveals key
proteins regulated by eicosapentaenoic acid in endothelial
activation. Biochem Biophys Res Commun. 487:464–469. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
He K, Guo X, Liu Y, Li J, Hu Y, Wang D and
Song J: TUFM induces epithelial-mesenchymal transition and invasion
in lung cancer cells via a mechanism involving AMPK-GSK3β
signaling. Cell Mol Life Sci. 73:2105–2121. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Geido E, Giaretti W and Nüsse M: Detection
of M and early-G1 phase cells by scattering signals combined with
identification of G1, S, and G2 phase cells. Methods Cell Biol.
33:149–156. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lei Y, Wen H, Yu Y, Taxman DJ, Zhang L,
Widman DG, Swanson KV, Wen KW, Damania B, Moore CB, et al: The
mitochondrial proteins NLRX1 and TUFM form a complex that regulates
type 1 interferon and autophagy. Immunity. 36:933–946. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Rakovic A, Grünewald A, Voges L, Hofmann
S, Orolicki S, Lohmann K and Klein C: PINK1-interacting proteins:
Proteomic analysis of overexpressed PINK1. Parkinsons Dis.
2011:1539792011.PubMed/NCBI
|
|
39
|
Lei Y, Kansy BA, Li J, Cong L, Liu Y,
Trivedi S, Wen H, Ting JPY, Ouyang H and Ferris RL: EGFR-targeted
mAb therapy modulates autophagy in head and neck squamous cell
carcinoma through NLRX1-TUFM protein complex. Oncogene.
35:4698–4707. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zottel A, Jovčevska I, Šamec N, Mlakar J,
Šribar J, Križaj I, Vidmar MS and Komel R: Anti-vimentin,
anti-TUFM, anti-NAP1L1 and anti-DPYSL2 nanobodies display cytotoxic
effect and reduce glioblastoma cell migration. Ther Adv Med Oncol.
12:17588359209153022020. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Samec N, Jovcevska I, Stojan J, Zottel A,
Liovic M, Myers MP, Muyldermans S, Šribar J, Križaj I and Komel R:
Glioblastoma-specific anti-TUFM nanobody for in-vitro immunoimaging
and cancer stem cell targeting. Oncotarget. 9:172822018. View Article : Google Scholar : PubMed/NCBI
|