Gastric carcinoma is the fifth most frequently
diagnosed cancer and the third leading cause of cancer-associated
death worldwide (1). In 2018 alone,
1,033,701 new cases and 782,685 deaths from gastric cancer were
expected globally (1). The 5-year
overall survival rate of patients with metastatic gastric cancer is
only 2%, with a median survival time of 8.6 months (2). Several studies have identified various
oncogenes and tumor suppressors that regulate the tumorigenesis of
gastric cancer (3,4). For instance, TP53 regulates target
genes in response to cellular stress and BRCA2 is involved in DNA
repair, which are both major genes that are frequently mutated in
gastric cancer (5,6). However, no well-established targets
besides human epidermal growth factor receptor 2 (7,8) have
been shown to modify the outcomes of patients with gastric cancer.
Therefore, it is essential to develop novel targets and therapeutic
approaches.
Transforming growth factor (TGF)-β is one of the
most extensively expressed cytokines in the tumor microenvironment,
and it plays an important role in tumor initiation and progression
(9). TGF-β is produced in large
amounts by numerous tumor types and is known to be pro-oncogenic
(10,11). The activated TGF-β receptor
transduces its signal via the phosphorylation of SMAD2/3 and
subsequent recruitment of SMAD4 intracellularly. This protein
complex then enters the nucleus and initiates the transcription of
the mesenchymal markers SNAI1, SNAI2, TWIST1 and ZEB1 (12–14),
eventually promoting the migration and invasion of tumor cells
(15). It has been reported that
high expression of TGF-β1 decreases the overall survival rate of
patients with gastric cancer (16).
Serpin peptidase inhibitor clade A member 1
(SERPINA1), a member of the protease inhibitor family of proteins,
is primarily synthesized in the liver. It is also produced in
certain neoplastic cells, such as those of colon, ovarian and lung
cancer (17–19). Tumor cells synthesize and release
SERPINA1, which plays a major role in physiological and
pathological processes, such as angiogenesis, wound healing, and
tumor invasion and metastasis (20).
The expression of SERPINA1 has been reported to be
correlated with poor prognoses in terms of metastasis among
patients with lung, colon and skin cancer (18,19,21,22).
However, details regarding the mechanism underlying the role of
SERPINA1 in the progression and metastasis of gastric cancer
remains unknown.
The present study aimed to provide important
insights into the mechanism underlying the pathogenesis of gastric
cancer and to evaluate SERPINA1 as a potential prognostic
biomarker.
The AGS gastric cancer cell line was purchased from
the American Type Culture Collection and authenticated using short
tandem repeat analysis (Beyotime Institute of Biotechnology) in
2020. The cells were maintained in Dulbecco's modified Eagle's
medium (DMEM) and 10% fetal calf serum (both Invitrogen; Thermo
Fisher Scientific, Inc.) in an incubator at 37°C with 5% carbon
dioxide. For SERPINA1-knockdown analyses, AGS cells were
transfected with 10 nM SERPINA1 small interfering (si)RNA
(siSERPINA1) or non-targeting negative control siRNA (siCONTROL)
using RNAiMAX transfection reagent (all Invitrogen; Thermo Fisher
Scientific, Inc.). For SERPINA1-overexpression experiments,
AGS cells were transfected with 1 µg SERPINA1 overexpression
vector pcDNA3.1(+)-SERPINA1 (pSERPINA1) plasmid (GenScript) or 1 µg
empty control vector pcDNA3.1(+)-SERPINA1_del (pCONTROL) plasmid
(GenScript) and selected using 1,200 µg/ml of G418 for 4 weeks.
Cells were fixed with 4% paraformaldehyde (PFA) in
phosphate-buffered saline (PBS) at room temperature for 15 min and
permeabilized in 0.1% Triton X-100 in PBS. Filamentous (F)-actin
was labeled with Alexa Fluor 555 phalloidin (165 nM; Invitrogen;
Thermo Fisher Scientific, Inc.) for 1 h at room temperature and
nuclei were stained with DAPI (1:10; PerkinElmer, Inc.) for 15 min
at room temperature. Images were captured using the Mantra
quantitative pathology imaging system (v1.03; PerkinElmer,
Inc.).
An ibidi culture-insert (Ibidi GmBH) was placed in
one well of a 6-well cell culture plate. Cells (70 µl/well;
1×106 cells/ml) were seeded into both wells and
incubated at 37°C in 5% carbon dioxide. After 24 h, the insert was
removed, creating a 500-µm cell-free gap, and subsequently 2
ml/well serum-free cell culture medium (DMEM) was added. Gap
closure was tracked and images were captured using an inverted
light microscope with a digital camera under ×10 magnification
(Olympus Corporation). Quantification of gap closure was performed
using ImageJ software (v1.52; National Institutes of Health).
Apoptosis was determined using Alexa Fluor
488-labeled Annexin V and the PI apoptosis detection kit
(Invitrogen; Thermo Fisher Scientific, Inc.). Cells were seeded in
a 6-well plate (2 ml/well; 0.15×106 cells/ml) and
cultured at 37°C for 48 h. Cells were then harvested and
resuspended, stained with Annexin V and propidium iodide (PI), and
analyzed using flow cytometry (BD LSRFortessa, BD Biosciences) and
FlowJo software (v10.6.1; BD Biosciences) within 1 h.
Cells were harvested, resuspended, and fixed in 70%
cold ethanol at 4°C for 12 h. The cells were then treated with DAPI
(BD Biosciences) in PBS for 30 min at room temperature in the dark.
The cell cycle was analyzed using the BD LSRFortessa flow cytometer
and FlowJo software (v10.6.1; BD Biosciences).
Cultured cells were lysed in a
radioimmunoprecipitation assay buffer (Thermo Fisher Scientific,
Inc.) containing a protease and phosphatase inhibitor cocktail
(Thermo Fisher Scientific, Inc.). Lysates were sonicated at 20 kHz
for 15 sec on ice and centrifuged at 10,000 × g for 10 min at 4°C.
Whole-lysate proteins (15–25 µg) were loaded in each lane. Gel
electrophoresis was performed using a 4–12% gradient polyacrylamide
gel (Invitrogen; Thermo Fisher Scientific, Inc.) and the proteins
were transferred to polyvinylidene difluoride membranes
(Invitrogen; Thermo Fisher Scientific, Inc.) according to standard
protocols (37). The membranes were
then cut and incubated with SERPINA1 (1:1,500, cat. no. ab207303,
Abcam), SMAD4 (1:1,500, cat. no. 46535S) or GAPDH (1:3,000, cat.
no. 5174S) (both Cell Signaling Technology, Inc.) antibodies
according to the known molecular weights of the proteins.
HRP-coupled goat anti-rabbit secondary antibody (cat. no. 7074S,
Cell Signaling Technology, Inc.) was used at 1:3,000 dilution.
Enhanced chemiluminescence signals were recorded and quantified
using the ChemiDoc MP imaging system and Image Lab v5.0 software
(both Bio-Rad Laboratories, Inc.).
Total RNA was isolated from cultured cell lines
using the RNeasy Plus Mini kit (Qiagen GmBH). RNA was reverse
transcribed into cDNA using the SuperScript IV First-Strand
Synthesis System kit (Invitrogen; Thermo Fisher Scientific, Inc.)
at 50°C for 20 min and then 80°C for 10 min. RT-qPCR was performed
using the QuantStudio 7 Real-Time PCR system (Applied Biosystems;
Thermo Fisher Scientific, Inc.) and the PowerUp SYBR Green Master
mix (Invitrogen; Thermo Fisher Scientific, Inc.). The PCR primer
sequences were as follows: SERPINA1, Forward:
5′-GAAGAGCGTCCTGGGTCAAC-3′ and reverse: 5′-TGGTCAGCACAGCCTTATGC-3′;
SMAD4, forward: 5′-CCCATCCCGGACATTACTGG-3′ and reverse:
5′-TAGGGCAGCTTGAAGGAACC-3′; PAI-1, forward:
5′-GCAAGGCACCTCTGAGAACT-3′ and reverse: 5′-GGGTGAGAAAACCACGTTGC-3′;
ACTB, forward: 5′-TGACATTAAGGAGAAGCTGTGCTA-3′ and reverse:
5′-GAGTTGAAGGTAGTTTCGTGGATG-3′. The relative expression of genes
was assessed using the 2−ΔΔCq method (38), and the expression of ACTB was used as
a reference.
Statistical analyses were carried out using GraphPad
Prism version 7 (GraphPad Software, Inc.). Data are presented as
the mean ± standard deviation and were compared using unpaired
Student's t-tests. The association between SERPINA1
expression and tumor stages in digestive system cancer datasets was
analyzed using one-way ANOVA. P<0.05 was considered to indicate
a statistically significant difference.
The effect of SERPINA1 on survival among patients
with gastric cancer was further analyzed by examining the
association between high SERPINA1 mRNA expression and poor
prognosis using the Kaplan-Meier Plotter online survival analysis
tool (31). The overall survival
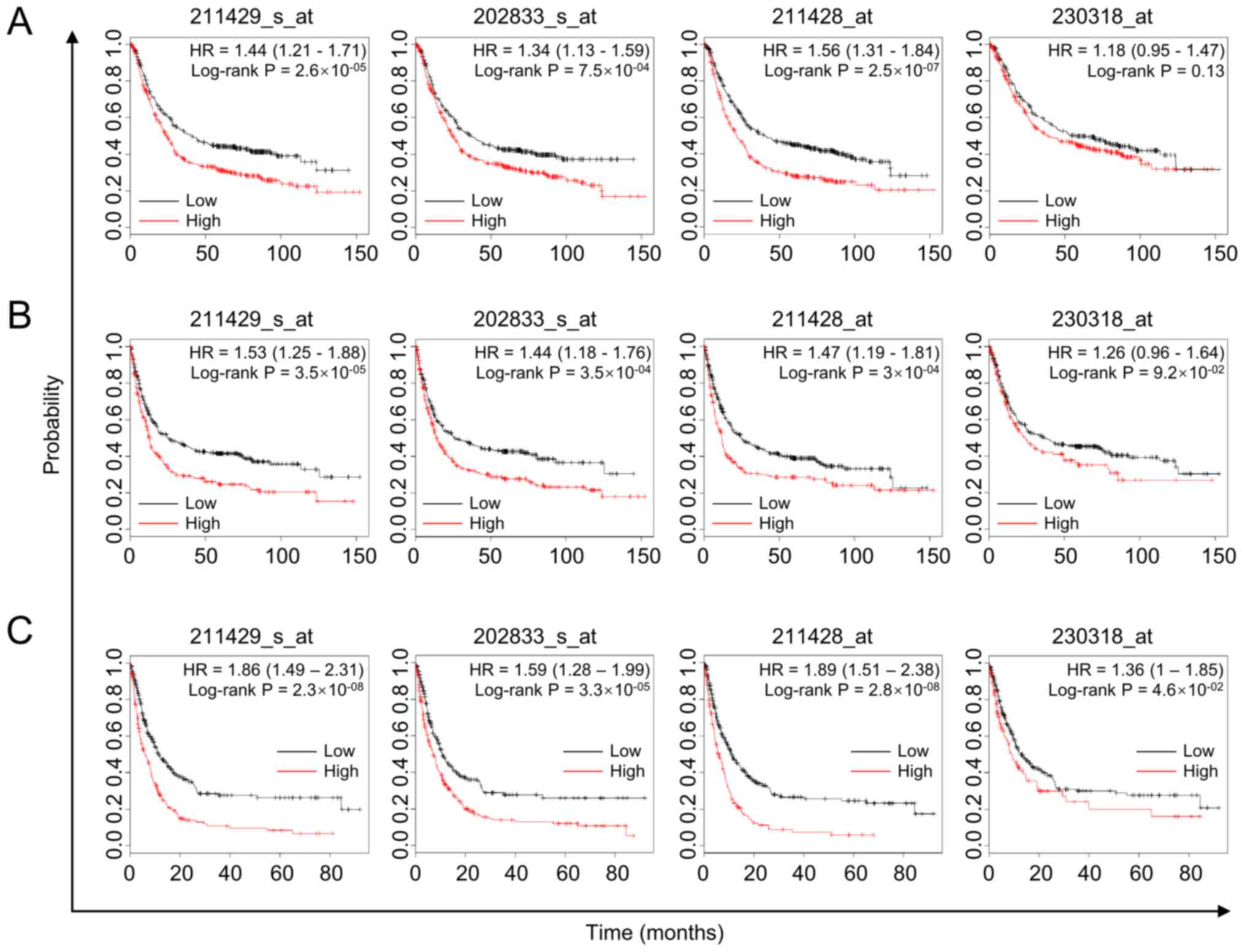
(OS) of 876 patients with gastric cancer showed that high
SERPINA1 expression was associated with shorter survival
time in probe 211429_s_at, 202833_s_at and 211428_at (log-rank
P=2.6×10−5, 7.5×10−4 and 2.5×10−7,
respectively; Fig. 2A). The
progression-free survival (PFS) time of 646 patients with gastric
cancer also showed the same outcomes in probe 202833_s_at,
211428_at and 211429_s_at (log-rank P=3.5×10−4,
3.0×10−4 and 3.5×10−5, respectively; Fig. 2B), as did the post-progression
survival time of 499 patients in all probes (Fig. 2C). In the analyses of OS and PFS, the
difference obtained for probe 230318_at did not reach statistical
significance (P<0.05). However, a similar tendency was observed
with the probes mentioned above (log-rank P=0.13 and 0.09,
respectively; Fig. 2A and B).
Overall, SERPINA1 is associated with clinical outcome of
gastric cancer patients and high SERPINA1 expression
indicates a short lifespan.
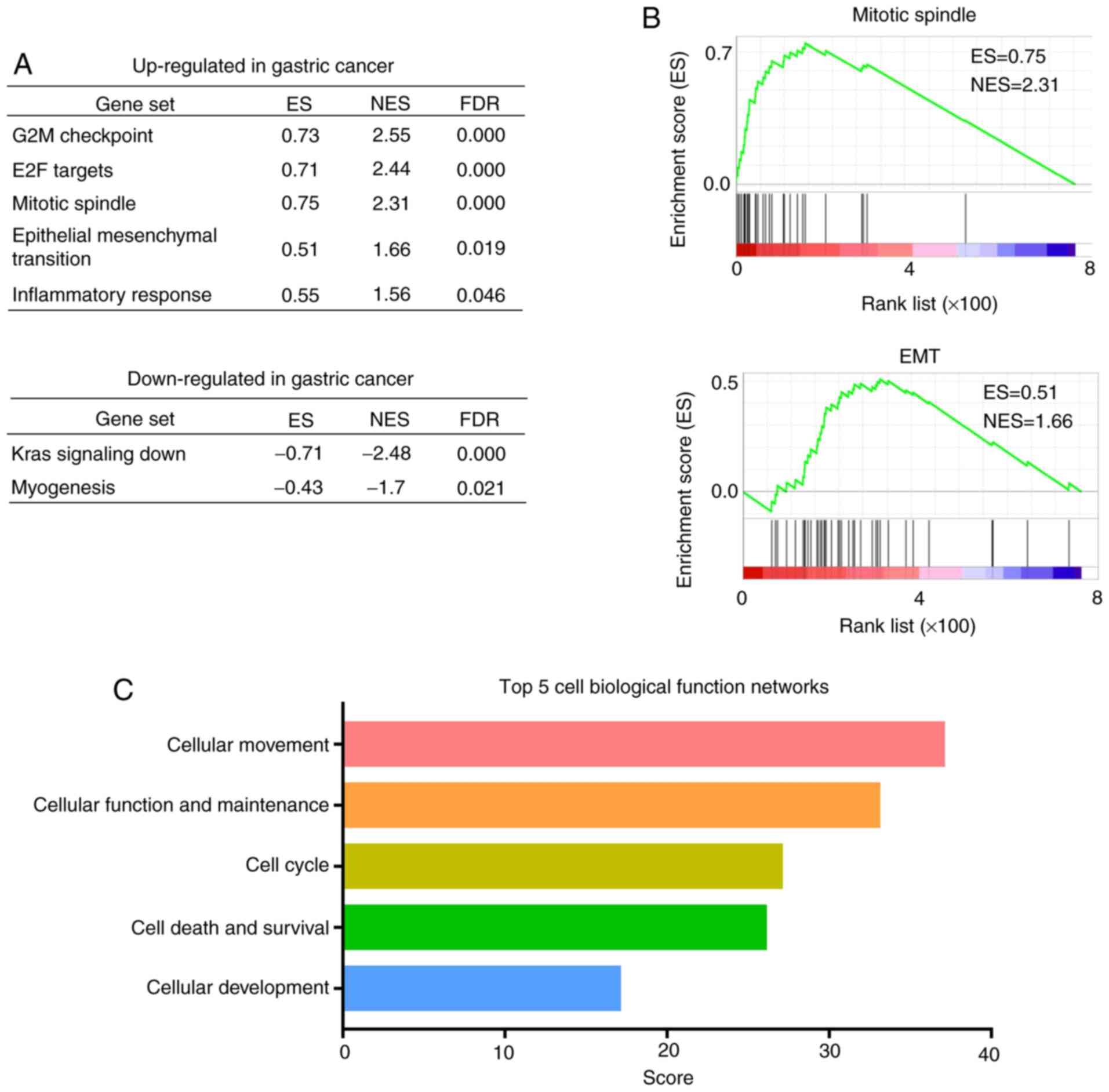
Employing GEPIA2, the DEGs in the gastric
adenocarcinoma TCGA-STAD dataset were analyzed. SERPINA1
(Log2FC=2.374, adjusted P=3.34×10−19) was
identified as one of 843 DEGs (Table
SII). A subsequent GSEA (32)
was performed that focused on hallmark gene sets representing 50
specific well-defined, large-scale biological processes and
displaying coherent expression (46). By analyzing the TCGA-STAD dataset,
five hallmark gene sets (G2M checkpoint, E2F targets, mitotic
spindle, epithelial mesenchymal transition and inflammatory
response) were identified in which DEGs were upregulated, and two
hallmark gene sets (Kras signaling down and myogenesis) in which
the DEGs were downregulated [all false discovery rate (FDR) q-value
<0.05; Fig. 3A]. Furthermore, two
of the upregulated gene sets, ‘mitotic spindle’ [normalized
enrichment score (NES)=2.31] (Fig.
3B) and ‘epithelial-mesenchymal-transition’ (EMT; NES=1.66)
(Fig. 3B) showed an association with
tumor progression.
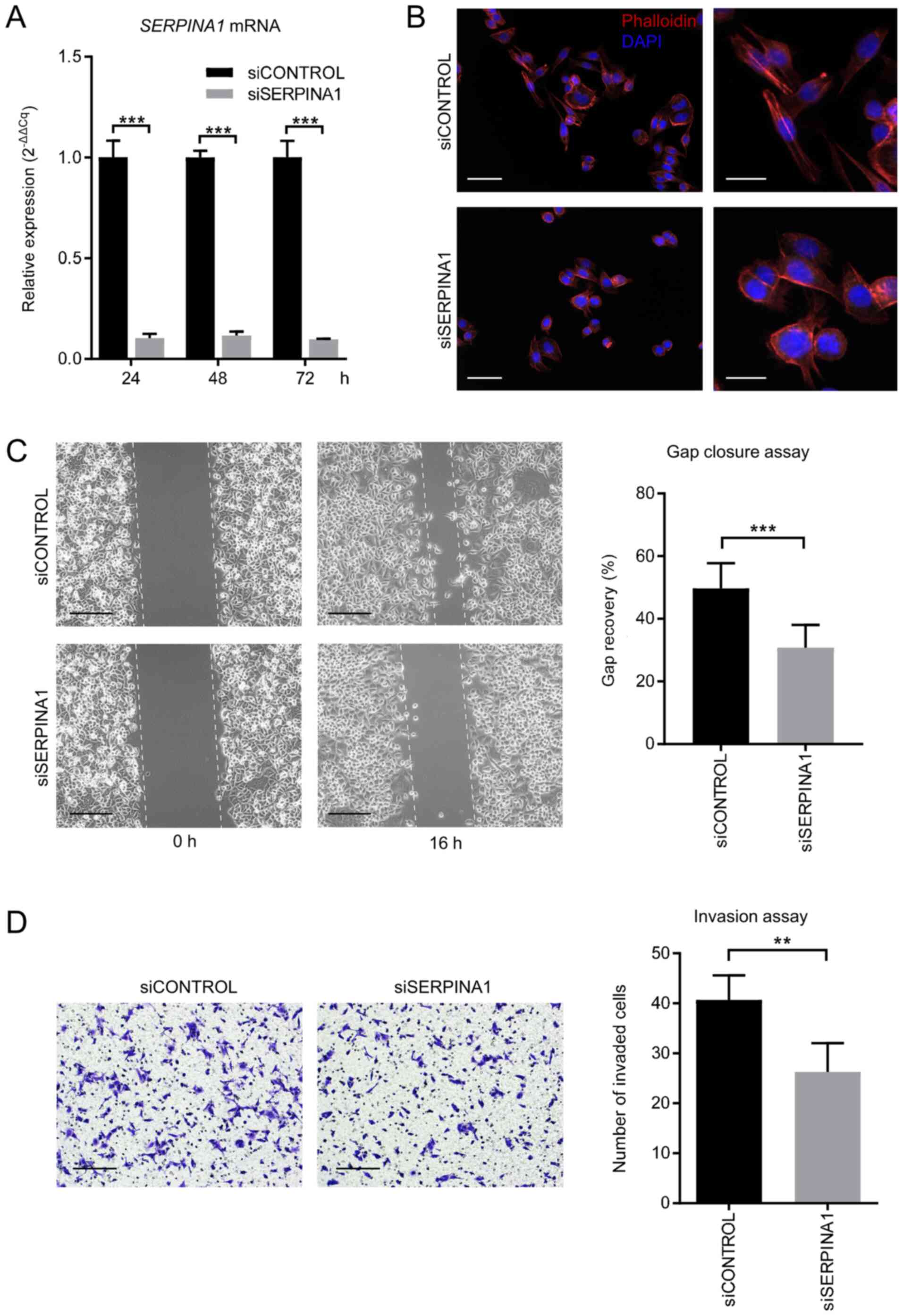
To exclude the possibility that the inhibition of
cell migration and invasion was due to an increased rate of
apoptosis and/or a state of proliferation arrest among cells, these
effects in the AGS cell line were analyzed after
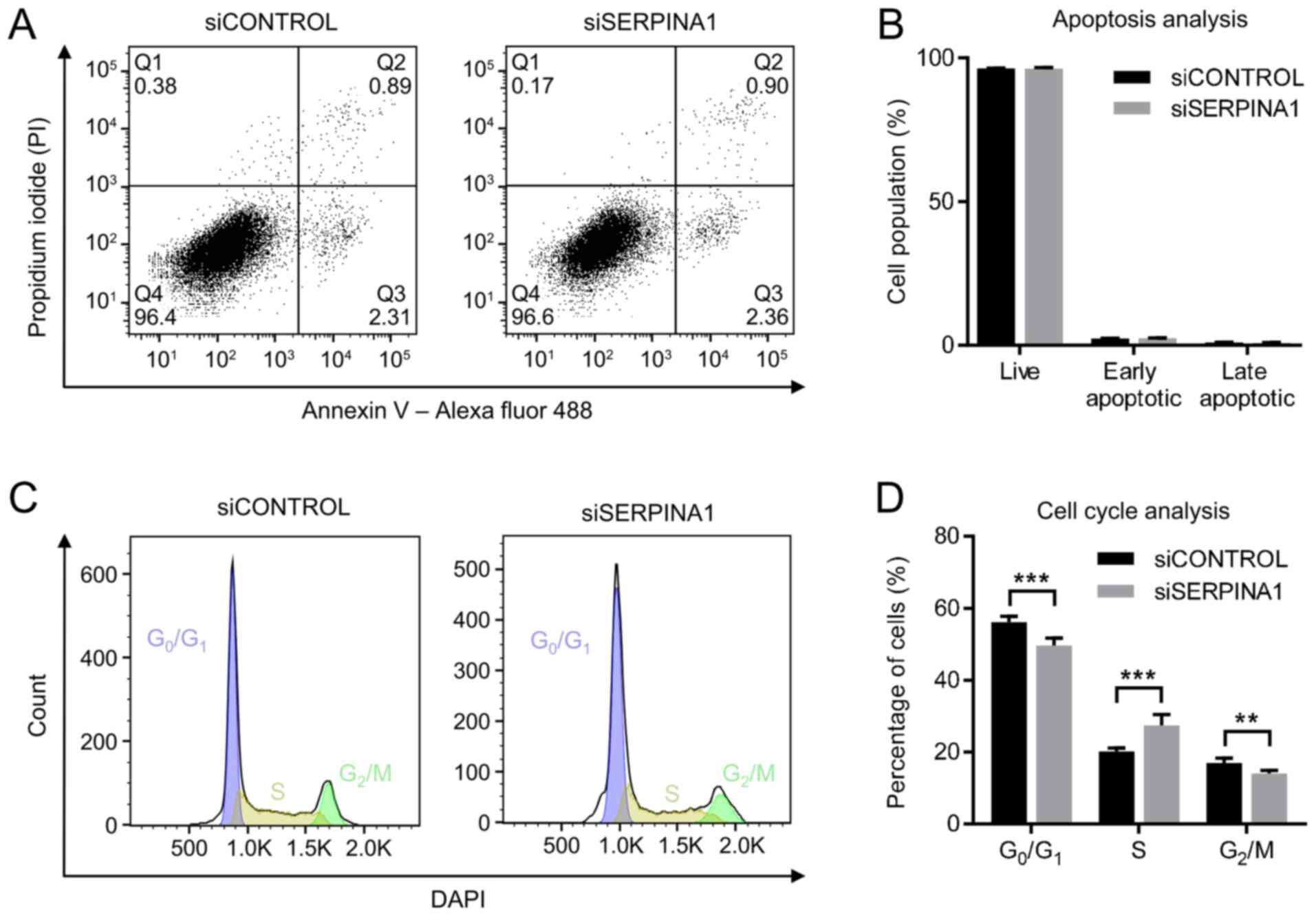
SERPINA1-knockdown. Dual staining of the
SERPINA1-downregulated AGS cells with PI and Annexin V-Alexa Fluor
488 was used to assess apoptosis. Flow cytometry revealed no
significant difference between SERPINA1 siRNA-treated AGS cells and
control siRNA-treated cells regarding the proportions of live
cells, early apoptotic cells and late apoptotic cells (Fig. 5A and B). Cell cycle distribution was
also evaluated by flow cytometric analysis using DAPI staining. No
significant shifts in the G1 and G2 peak
positions were observed among AGS cells after SERPINA1 siRNA
transfection (Fig. 5C). However, a
significantly increased proportion of S phase cells (from 20.2 to
27.4%), decreased G0/G1-phase (from 56.2 to
49.6%) and G2/M-phase (from 17.0 to 14.1%) cells were
observed (Fig. 5D) after
SERPINA1-knockdown. This suggested that gene silencing of
SERPINA1 promoted S phase entry in the AGS cell line.
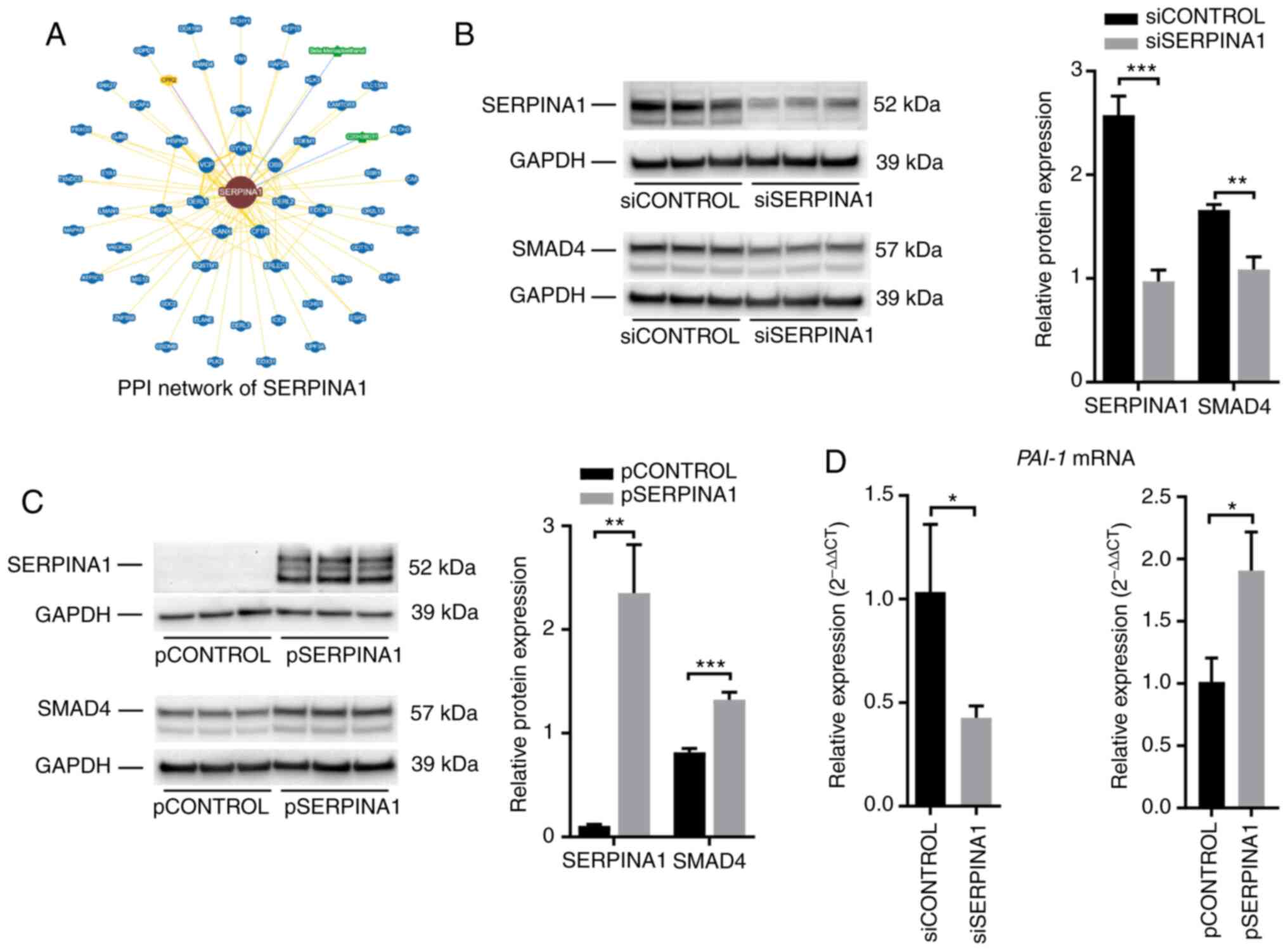
To elucidate the mechanism underlying the effects of
SERPINA1, protein-protein interaction data of SERPINA1 were queried
using the BioGRID (35) and Wiki-Pi
(36) databases. Seventy-three
interactions involving SERPINA1 were identified in the BioGRID
database (Fig. 7A and Table SIV), and 26 interactions were
reported in the Wiki-Pi database (Table
SV). SMAD4, which plays a role in regulating TGF-β-mediated EMT
in breast cancer (48),
nasopharyngeal carcinoma (49) and
squamous cell carcinoma of the head and neck (50), was found to interact with SERPINA1 in
both databases.
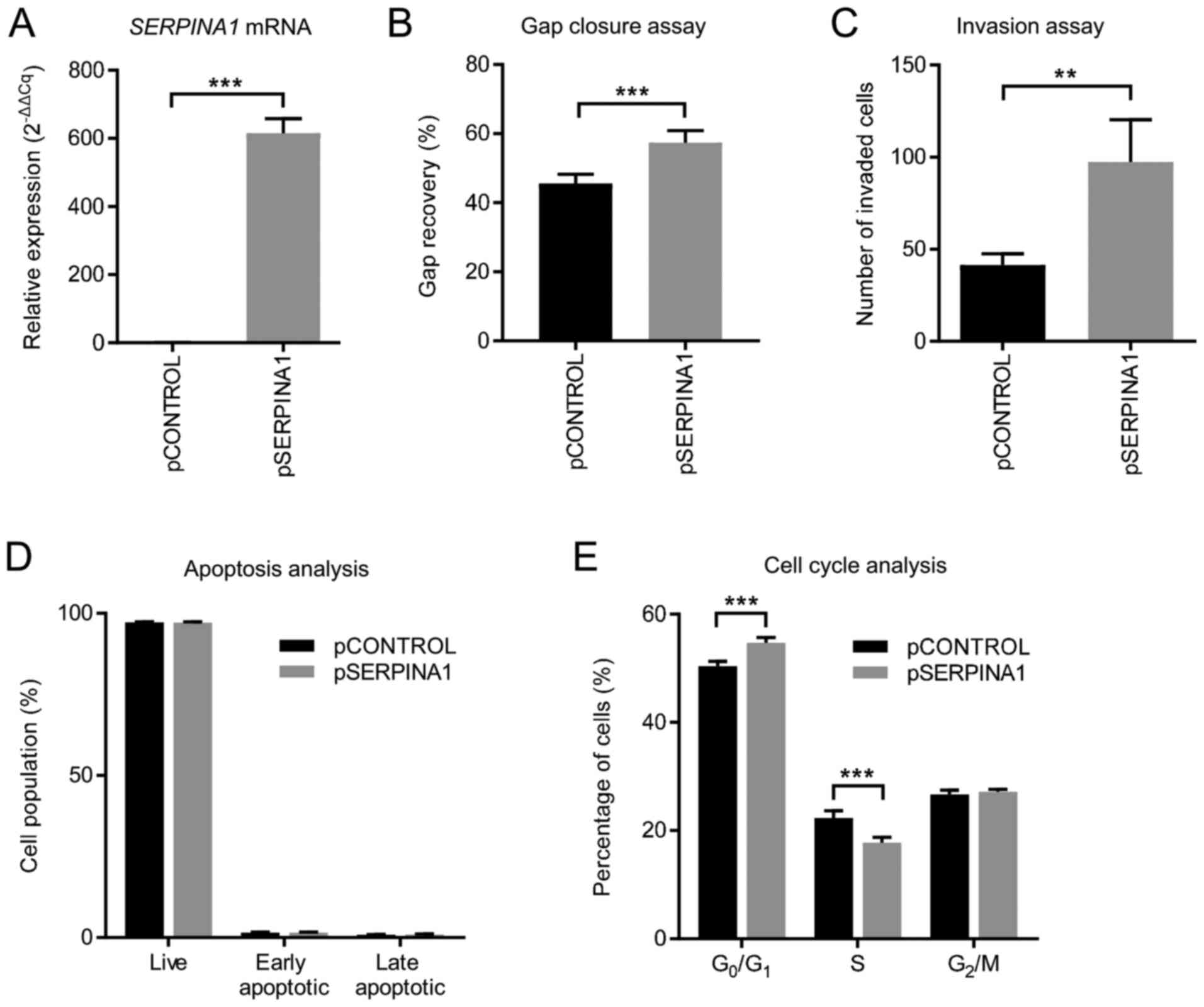
To further assess whether SERPINA1 regulated the
TGF-β signaling pathway via SMAD4, the expression of SMAD4 protein
in both SERPINA1-knockdown and
SERPINA1-overexpression AGS cells was analyzed. Western
blotting showed that the SMAD4 protein levels were downregulated
after silencing SERPINA1 (Fig.
7B), whereas overexpression of SERPINA1 led to an
upregulation of SMAD4 protein (Fig.
7C). In addition, changes in the mRNA levels of plasminogen
activator inhibitor 1 (a SMAD4-dependent TGF-β signaling target
gene) (51) (Fig. 7D) were consistent with changes in
SMAD4 protein expression in AGS cells subjected to
SERPINA1-knockout and overexpression. These findings
suggested that SERPINA1 might regulate the TGF-β signaling pathway
through interaction with SMAD4.
In several Western Asian countries, gastric cancer
is the most commonly diagnosed cancer and, in Eastern Asia,
incidence rates have increased markedly in the past few decades;
for example, the crude incidence rate in the Republic of Korea was
78.3 per 100,000 among males and 37.2 among females in 2012,
compared with a crude incidence rate of 49.2 among males and 27.3
among females in 1999 (1,52,53).
However, the underlying mechanisms driving poor clinical outcomes
are not understood. Therefore, the identification of novel
biomarkers and therapeutic targets for gastric cancer are essential
to improve prognosis determination and treatment.
SERPINA1 is a member of the serpins superfamily of
protease inhibitors, which play a crucial physiological role in
hormone transport, blood clotting, corticosteroid binding and blood
pressure regulation (54). However,
serpins have also been found to function in tumorigenesis and
cancer metastasis (54).
Particularly, SERPINA1 has been reported to be overexpressed
in various malignant tumors. High expression of SERPINA1 has
been observed in prostate (55),
lung (55) and colorectal (18) cancer. SERPINA1 was also
reported to be upregulated in serum samples of patients with
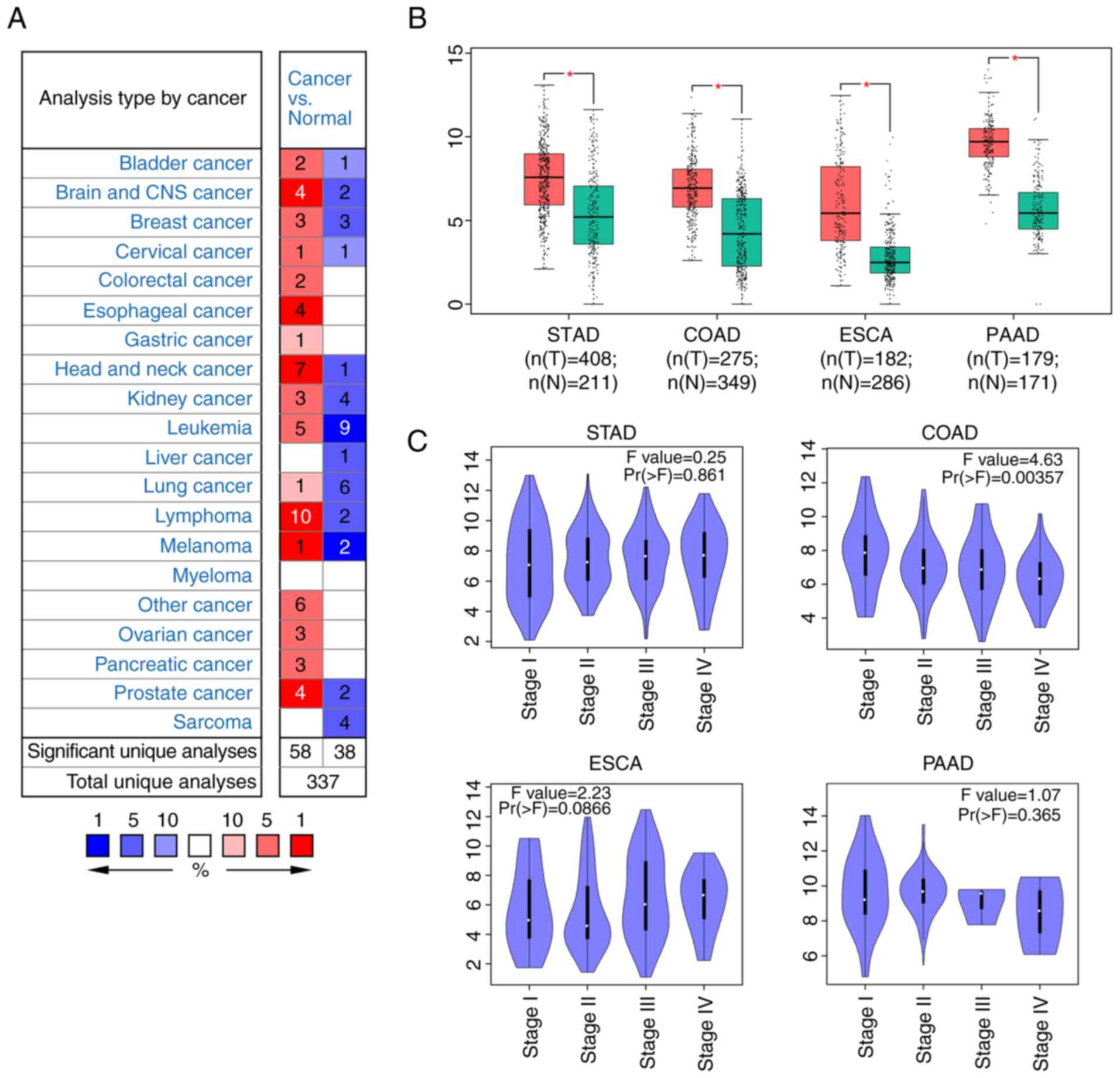
gastric cancer compared with healthy individuals (56). The present study demonstrated that
SERPINA1 was overexpressed in colorectal, esophageal,
gastric and pancreatic cancer, which indicated a strong association
between a high SERPINA1 mRNA levels and digestive system
tumorigenesis. In addition, it was reported that overexpression of
SERPINA1 was associated with a shorter lifespan in four
gastric cancer datasets. Consistent with the present results, Kwon
et al (57) also reported an
inverse correlation between SERPINA1 expression and survival
time in the Korean population.
Despite its clinical relevance, the functional role
of SERPINA1 in tumor cells remains unknown. Using bioinformatics
analyses of the TCGA-STAD database, the present study revealed that
the expression of SERPINA1 was significantly upregulated compared
with normal tissues. Further GSEA revealed that the ‘mitotic
spindle’ and ‘EMT’ gene sets were enriched in cancer tissues, which
were associated with the function of the cytoskeleton. In addition,
IPA analysis demonstrated that the top biological function network
of SERPINA1-enriched genes was ‘cellular movement’. All these
findings suggested that SERPINA1 may be involved in cellular
movement, which is mediated by the cytoskeleton (58). To resolve the underlying molecular
mechanisms of SERPINA1 in gastric cancer progression, the
expression of the SERPINA1 gene was manipulated in
vitro. A series of investigations revealed that SERPINA1
regulated cell morphology, migration and invasion and the number of
S phase cells, but had no impact on apoptosis in the AGS gastric
cell line. Similarly, the regulation of SERPINA1 in cell migration
and invasion has also been reported in ovarian (17) and colon cancer (18) cell lines.
Although the mechanisms underlying the role of
SERPINA1 in tumor cell migration/invasion and cell cycle have not
been fully elucidated, it has been shown that fibronectin is
upregulated by SERPINA1 (18). The
upregulation of fibronectin promotes cell migration and invasion in
colorectal cancer (18). Byon et
al (59) and Hernanda et
al (60) observed enhanced cell
migration through SMAD4 in breast cancer cells and hepatoma cells,
respectively. The present study demonstrated that overexpression of
SERPINA1 upregulated the expression of SMAD4, and
subsequently activated the SMAD4-dependent TGF-β signaling pathway.
This promoted the migration and invasion of human gastric cancer
cells and reduced the proportion of S phase cells. Based on the
present data, it was hypothesized that SERPINA1 may protect the
intracellular transport of SMAD4 from the cell membrane to the
nucleus (61), as SERPINA1 inhibits
the activity of protease and may prevent SMAD4 complex degradation
(62). However,
co-immunoprecipitation experiments are required to confirm the
binding of SERPINA1 and SMAD4, and further signal transduction
studies may be helpful to elucidate the precise mechanisms
underlying the regulation of SMAD4 expression by SERPINA1.
Not applicable.
The present study was funded by Amgen Asia Research
and Development Center.
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
LJ and LGH conceived and designed this study and
wrote the manuscript. LJ performed the experiments and analyzed the
data. Both authors read and approved the final manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018.PubMed/NCBI
|
|
2
|
Ben Kridis W, Marrekchi G, Mzali R, Daoud
J and Khanfir A: Prognostic factors in metastatic gastric
carcinoma. Exp Oncol. 41:173–175. 2019.PubMed/NCBI
|
|
3
|
Matsuoka T and Yashiro M: Biomarkers of
gastric cancer: Current topics and future perspective. World J
Gastroentero. 24:2818–2832. 2018.
|
|
4
|
Chia NY and Tan P: Molecular
classification of gastric cancer. Ann Oncol. 27:763–769.
2016.PubMed/NCBI
|
|
5
|
Gao J, Aksoy BA, Dogrusoz U, Dresdner G,
Gross B, Sumer SO, Sun Y, Jacobsen A, Sinha R, Larsson E, et al:
Integrative analysis of complex cancer genomics and clinical
profiles using the cBioPortal. Sci Signal. 6:pl12013.PubMed/NCBI
|
|
6
|
Chen K, Yang D, Li X, Sun B, Song F, Cao
W, Brat DJ, Gao Z, Li H, Liang H, et al: Mutational landscape of
gastric adenocarcinoma in Chinese: Implications for prognosis and
therapy. Proc Natl Acad Sci USA. 112:1107–1112. 2015.PubMed/NCBI
|
|
7
|
Boku N: HER2-positive gastric cancer.
Gastric Cancer. 17:1–12. 2014.PubMed/NCBI
|
|
8
|
Oh DY and Bang YJ: HER2-targeted
therapies-a role beyond breast cancer. Nat Rev Clin Oncol.
17:33–48. 2020.PubMed/NCBI
|
|
9
|
Pickup M, Novitskiy S and Moses HL: The
roles of TGFβ in the tumour microenvironment. Nat Rev Cancer.
13:788–799. 2013.PubMed/NCBI
|
|
10
|
Batlle E and Massagué J: Transforming
growth factor-β signaling in immunity and cancer. Immunity.
50:924–940. 2019.PubMed/NCBI
|
|
11
|
Zhao M, Mishra L and Deng CX: The role of
TGF-β/SMAD4 signaling in cancer. Int J Biol Sci. 14:111–123.
2018.PubMed/NCBI
|
|
12
|
Vincent T, Neve EP, Johnson JR, Kukalev A,
Rojo F, Albanell J, Pietras K, Virtanen I, Philipson L, Leopold PL,
et al: A SNAIL1- SMAD3/4 transcriptional repressor complex promotes
TGF-beta mediated epithelial-mesenchymal transition. Nat Cell Biol.
11:943–950. 2009.PubMed/NCBI
|
|
13
|
Bracken CP, Gregory PA, Kolesnikoff N,
Bert AG, Wang J, Shannon MF and Goodall GJ: A double-negative
feedback loop between ZEB1-SIP1 and the microRNA-200 family
regulates epithelial-mesenchymal transition. Cancer Res.
68:7846–7854. 2008.PubMed/NCBI
|
|
14
|
Thuault S, Valcourt U, Petersen M,
Manfioletti G, Heldin CH and Moustakas A: Transforming growth
factor-beta employs HMGA2 to elicit epithelial-mesenchymal
transition. J Cell Biol. 174:175–183. 2006.PubMed/NCBI
|
|
15
|
Zhang H, Liu L, Wang Y, Zhao G, Xie R, Liu
C, Xiao X, Wu K, Nie Y, Zhang H and Fan D: KLF8 involves in
TGF-beta-induced EMT and promotes invasion and migration in gastric
cancer cells. J Cancer Res Clin Oncol. 139:1033–1042.
2013.PubMed/NCBI
|
|
16
|
Hawinkels LJ, Verspaget HW, van Duijn W,
van der Zon JM, Zuidwijk K, Kubben FJ, Verheijen JH, Hommes DW,
Lamers CB and Sier CF: Tissue level, activation and cellular
localisation of TGF-beta1 and association with survival in gastric
cancer patients. Br J Cancer. 97:398–404. 2007.PubMed/NCBI
|
|
17
|
Normandin K, Péant B, Le Page C, de
Ladurantaye M, Ouellet V, Tonin PN, Provencher DM and Mes-Masson
AM: Protease inhibitor SERPINA1 expression in epithelial ovarian
cancer. Clin Exp Metastas. 27:55–69. 2010.
|
|
18
|
Kwon CH, Park HJ, Choi JH, Lee JR, Kim HK,
Jo HJ, Kim HS, Oh N, Song GA and Park DY: Snail and serpinA1
promote tumor progression and predict prognosis in colorectal
cancer. Oncotarget. 6:20312–20326. 2015.PubMed/NCBI
|
|
19
|
Ercetin E, Richtmann S, Delgado BM,
Gomez-Mariano G, Wrenger S, Korenbaum E, Liu B, DeLuca D, Kuhnel
MP, Jonigk D, et al: Clinical significance of SERPINA1 gene and its
encoded alpha1-antitrypsin protein in NSCLC. Cancers (Basel).
11:13062019.
|
|
20
|
Janciauskiene S: Conformational properties
of serine proteinase inhibitors (serpins) confer multiple
pathophysiological roles. Biochim Biophys Acta. 1535:221–235.
2001.PubMed/NCBI
|
|
21
|
Farshchian M, Kivisaari A, Ala-aho R,
Riihilä P, Kallajoki M, Grénman R, Peltonen J, Pihlajaniemi T,
Heljasvaara R and Kähäri VM: Serpin peptidase inhibitor Clade A
member 1 (SerpinA1) is a novel biomarker for progression of
cutaneous squamous cell carcinoma. Am J Pathol. 179:1110–1119.
2011.PubMed/NCBI
|
|
22
|
Chan HJ, Li H, Liu Z, Yuan YC, Mortimer J
and Chen S: SERPINA1 is a direct estrogen receptor target gene and
a predictor of survival in breast cancer patients. Oncotarget.
6:25815–25827. 2015.PubMed/NCBI
|
|
23
|
Rhodes DR, Yu J, Shanker K, Deshpande N,
Varambally R, Ghosh D, Barrette T, Pandey A and Chinnaiyan AM:
ONCOMINE: A cancer microarray database and integrated data-mining
platform. Neoplasia. 6:1–6. 2004.PubMed/NCBI
|
|
24
|
Tang Z, Li C, Kang B, Gao G, Li C and
Zhang Z: GEPIA: A web server for cancer and normal gene expression
profiling and interactive analyses. Nucleic Acids Res.
45W:W98–W102. 2017.
|
|
25
|
Bass AJ, Thorsson V, Shmulevich I,
Reynolds SM, Miller M, Bernard B, Hinoue T, Laird PW, Curtis C,
Shen H, et al: Comprehensive molecular characterization of gastric
adenocarcinoma. Nature. 513:202–209. 2014.PubMed/NCBI
|
|
26
|
Muzny DM, Bainbridge MN, Chang K, Dinh HH,
Drummond JA, Fowler G, Kovar CL, Lewis LR, Morgan MB, Newsham IF,
et al: Comprehensive molecular characterization of human colon and
rectal cancer. Nature. 487:330–337. 2012.PubMed/NCBI
|
|
27
|
Kim J, Bowlby R, Mungall AJ, Robertson AG,
Odze RD, Cherniack AD, Shih J, Pedamallu CS, Cibulskis C, Dunford
A, et al: Integrated genomic characterization of oesophageal
carcinoma. Nature. 541:169–174. 2017.PubMed/NCBI
|
|
28
|
Raphael BJ, Hruban RH, Aguirre AJ, Moffitt
RA, Yeh JJ, Stewart C, Robertson AG, Cherniack AD, Gupta M, Getz G,
et al: Integrated genomic characterization of pancreatic ductal
adenocarcinoma. Cancer Cell. 32:185–203.e13. 2017.PubMed/NCBI
|
|
29
|
Carithers LJ, Ardlie K, Barcus M, Branton
PA, Britton A, Buia SA, Compton CC, DeLuca DS, Peter-Demchok J,
Gelfand ET, et al: A novel approach to high-quality postmortem
tissue procurement: The GTEx project. Biopreserv Biobank.
13:311–319. 2015.PubMed/NCBI
|
|
30
|
Amin MB, Greene FL, Edge SB, Compton CC,
Gershenwald JE, Brookland RK, Meyer L, Gress DM, Byrd DR and
Winchester DP: The eighth edition AJCC cancer staging manual:
Continuing to build a bridge from a population-based to a more
‘personalized’ approach to cancer staging. CA Cancer J Clin.
67:93–99. 2017.PubMed/NCBI
|
|
31
|
Szasz AM, Lanczky A, Nagy A, Forster S,
Hark K, Green JE, Boussioutas A, Busuttil R, Szabo A and Gyorffy B:
Cross-validation of survival associated biomarkers in gastric
cancer using transcriptomic data of 1,065 patients. Oncotarget.
7:49322–49333. 2016.PubMed/NCBI
|
|
32
|
Subramanian A, Tamayo P, Mootha VK,
Mukherjee S, Ebert BL, Gillette MA, Paulovich A, Pomeroy SL, Golub
TR, Lander ES and Mesirov JP: Gene set enrichment analysis: A
knowledge-based approach for interpreting genome-wide expression
profiles. Proc Natl Acad Sci USA. 102:15545–15550. 2005.PubMed/NCBI
|
|
33
|
Cerami E, Gao J, Dogrusoz U, Gross BE,
Sumer SO, Aksoy BA, Jacobsen A, Byrne CJ, Heuer ML, Larsson E, et
al: The cBio cancer genomics portal: An open platform for exploring
multidimensional cancer genomics data. Cancer Discov. 2:401–404.
2012.PubMed/NCBI
|
|
34
|
Kramer A, Green J, Pollard J Jr and
Tugendreich S: Causal analysis approaches in ingenuity pathway
analysis. Bioinformatics. 30:523–530. 2014.PubMed/NCBI
|
|
35
|
Oughtred R, Stark C, Breitkreutz BJ, Rust
J, Boucher L, Chang C, Kolas N, O'Donnell L, Leung G, McAdam R, et
al: The BioGRID interaction database: 2019 update. Nucleic Acids
Res. 47D:D529–D541. 2019.
|
|
36
|
Orii N and Ganapathiraju MK: Wiki-pi: A
web-server of annotated human protein-protein interactions to aid
in discovery of protein function. PLoS One. 7:e490292012.PubMed/NCBI
|
|
37
|
Kim B: Western blot techniques. Methods
Mol Biol. 1606:133–139. 2017.PubMed/NCBI
|
|
38
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI
|
|
39
|
Skrzypczak M, Goryca K, Rubel T, Paziewska
A, Mikula M, Jarosz D, Pachlewski J, Oledzki J and Ostrowski J:
Modeling oncogenic signaling in colon tumors by multidirectional
analyses of microarray data directed for maximization of analytical
reliability. PLoS One. 5:e130912010.PubMed/NCBI
|
|
40
|
Sabates-Bellver J, Van der Flier LG, de
Palo M, Cattaneo E, Maake C, Rehrauer H, Laczko E, Kurowski MA,
Bujnicki JM, Menigatti M, et al: Transcriptome profile of human
colorectal adenomas. Mol Cancer Res. 5:1263–1275. 2007.PubMed/NCBI
|
|
41
|
Kimchi ET, Posner MC, Park JO, Darga TE,
Kocherginsky M, Karrison T, Hart J, Smith KD, Mezhir JJ,
Weichselbaum RR and Khodarev NN: Progression of Barrett's
metaplasia to adenocarcinoma is associated with the suppression of
the transcriptional programs of epidermal differentiation. Cancer
Res. 65:3146–3154. 2005.PubMed/NCBI
|
|
42
|
Kim SM, Park YY, Park ES, Cho JY, Izzo JG,
Zhang D, Kim SB, Lee JH, Bhutani MS, Swisher SG, et al: Prognostic
biomarkers for esophageal adenocarcinoma identified by analysis of
tumor transcriptome. PLoS One. 5:e150742010.PubMed/NCBI
|
|
43
|
Wang Q, Wen YG, Li DP, Xia J, Zhou CZ, Yan
DW, Tang HM and Peng ZH: Upregulated INHBA expression is associated
with poor survival in gastric cancer. Med Oncol. 29:77–83.
2012.PubMed/NCBI
|
|
44
|
Badea L, Herlea V, Dima SO, Dumitrascu T
and Popescu I: Combined gene expression analysis of whole-tissue
and microdissected pancreatic ductal adenocarcinoma identifies
genes specifically overexpressed in tumor epithelia.
Hepatogastroenterology. 55:2016–2027. 2008.PubMed/NCBI
|
|
45
|
Logsdon CD, Simeone DM, Binkley C,
Arumugam T, Greenson JK, Giordano TJ, Misek DE, Kuick R and Hanash
S: Molecular profiling of pancreatic adenocarcinoma and chronic
pancreatitis identifies multiple genes differentially regulated in
pancreatic cancer. Cancer Res. 63:2649–2657. 2003.PubMed/NCBI
|
|
46
|
Liberzon A, Birger C, Thorvaldsdottir H,
Ghandi M, Mesirov JP and Tamayo P: The molecular signatures
database (MSigDB) hallmark gene set collection. Cell Syst.
1:417–425. 2015.PubMed/NCBI
|
|
47
|
Faulstich H, Zobeley S, Rinnerthaler G and
Small JV: Fluorescent phallotoxins as probes for filamentous actin.
J Muscle Res Cell Motil. 9:370–383. 1988.PubMed/NCBI
|
|
48
|
Deckers M, van Dinther M, Buijs J, Que I,
Lowik C, van der Pluijm G and ten Dijke P: The tumor suppressor
Smad4 is required for transforming growth factor beta-induced
epithelial to mesenchymal transition and bone metastasis of breast
cancer cells. Cancer Res. 66:2202–2209. 2006.PubMed/NCBI
|
|
49
|
Huang G, Du MY, Zhu H, Zhang N, Lu ZW,
Qian LX, Zhang W, Tian X, He X and Yin L: MiRNA-34a reversed
TGF-β-induced epithelial-mesenchymal transition via suppression of
SMAD4 in NPC cells. Biomed Pharmacother. 106:217–224.
2018.PubMed/NCBI
|
|
50
|
Yu C, Liu Y, Huang D, Dai Y, Cai G, Sun J,
Xu T, Tian Y and Zhang X: TGF-β1 mediates epithelial to mesenchymal
transition via the TGF-β/Smad pathway in squamous cell carcinoma of
the head and neck. Oncol Rep. 25:1581–1587. 2011.PubMed/NCBI
|
|
51
|
Levy L and Hill CS: Smad4 dependency
defines two classes of transforming growth factor β (TGF-β) target
genes and distinguishes TGF-β-induced epithelial-mesenchymal
transition from its antiproliferative and migratory responses. Mol
Cell Biol. 25:8108–8125. 2005.PubMed/NCBI
|
|
52
|
Bray F, Colombet M, Mery L, Piñeros M,
Znaor A, Zanetti R and Ferlay J: Cancer Incidence in Five
Continents. XI. International Agency for Research on Cancer; Lyon:
2017, http://ci5.iarc.frJuly
20–2020
|
|
53
|
Curado MP, Edwards B, Shin HR, Storm H,
Ferlay J, Heanue M and Boyle P: Cancer Incidence in Five
Continents. IX. International Agency for Research on Cancer; Lyon:
2007, http://ci5.iarc.frJuly
20–2020
|
|
54
|
Heit C, Jackson BC, McAndrews M, Wright
MW, Thompson DC, Silverman GA, Nebert DW and Vasiliou V: Update of
the human and mouse SERPIN gene superfamily. Hum Genomics.
7:222013.PubMed/NCBI
|
|
55
|
El-Akawi ZJ, Al-Hindawi FK and Bashir NA:
Alpha-1 antitrypsin (alpha1-AT) plasma levels in lung, prostate and
breast cancer patients. Neuro Endocrinol Lett. 29:482–484.
2008.PubMed/NCBI
|
|
56
|
Yang J, Xiong X, Wang X, Guo B, He K and
Huang C: Identification of peptide regions of SERPINA1 and ENOSF1
and their protein expression as potential serum biomarkers for
gastric cancer. Tumour Biol. 36:5109–5118. 2015.PubMed/NCBI
|
|
57
|
Kwon CH, Park HJ, Lee JR, Kim HK, Jeon TY,
Jo HJ, Kim DH, Kim GH and Park DY: Serpin peptidase inhibitor clade
A member 1 is a biomarker of poor prognosis in gastric cancer. Br J
Cancer. 111:1993–2002. 2014.PubMed/NCBI
|
|
58
|
Pollard TD: The cytoskeleton, cellular
motility and the reductionist agenda. Nature. 422:741–745.
2003.PubMed/NCBI
|
|
59
|
Byon CH, Hardy RW, Ren C, Ponnazhagan S,
Welch DR, McDonald JM and Chen Y: Free fatty acids enhance breast
cancer cell migration through plasminogen activator inhibitor-1 and
SMAD4. Lab Invest. 89:1221–1228. 2009.PubMed/NCBI
|
|
60
|
Hernanda PY, Chen K, Das AM, Sideras K,
Wang W, Li J, Cao W, Bots SJ, Kodach LL, de Man RA, et al: SMAD4
exerts a tumor-promoting role in hepatocellular carcinoma.
Oncogene. 34:5055–5068. 2015.PubMed/NCBI
|
|
61
|
Heldin CH, Miyazono K and Ten Dijke P:
TGF-beta signalling from cell membrane to nucleus through SMAD
proteins. Nature. 390:465–471. 1997.PubMed/NCBI
|
|
62
|
Komiyama T, Gron H, Pemberton PA and
Salvesen GS: Interaction of subtilisins with serpins. Protein Sci.
5:874–882. 1996.PubMed/NCBI
|
|
63
|
Christensson A, Laurell CB and Lilja H:
Enzymatic activity of prostate-specific antigen and its reactions
with extracellular serine proteinase inhibitors. Eur J Biochem.
194:755–763. 1990.PubMed/NCBI
|
|
64
|
Korkmaz B, Attucci S, Hazouard E,
Ferrandiere M, Jourdan ML, Brillard-Bourdet M, Juliano L and
Gauthier F: Discriminating between the activities of human
neutrophil elastase and proteinase 3 using serpin-derived
fluorogenic substrates. J Biol Chem. 277:39074–39081.
2002.PubMed/NCBI
|
|
65
|
Taggart C, Cervantes-Laurean D, Kim G,
McElvaney NG, Wehr N, Moss J and Levine RL: Oxidation of either
methionine 351 or methionine 358 in alpha 1-antitrypsin causes loss
of anti-neutrophil elastase activity. J Biol Chem. 275:27258–27265.
2000.PubMed/NCBI
|
|
66
|
Greenblatt EJ, Olzmann JA and Kopito RR:
Derlin-1 is a rhomboid pseudoprotease required for the dislocation
of mutant α-1 antitrypsin from the endoplasmic reticulum. Nat
Struct Mol Biol. 18:1147–1152. 2011.PubMed/NCBI
|
|
67
|
Hutchins JR, Toyoda Y, Hegemann B, Poser
I, Heriche JK, Sykora MM, Augsburg M, Hudecz O, Buschhorn BA,
Bulkescher J, et al: Systematic analysis of human protein complexes
identifies chromosome segregation proteins. Science. 328:593–599.
2010.PubMed/NCBI
|
|
68
|
Wang J, Yuan Y, Zhou Y, Guo L, Zhang L,
Kuai X, Deng B, Pan Z, Li D and He F: Protein interaction data set
highlighted with human Ras-MAPK/PI3K signaling pathways. J Proteome
Res. 7:3879–3889. 2008.PubMed/NCBI
|
|
69
|
LaLonde DP and Bretscher A: The UBX
protein SAKS1 negatively regulates endoplasmic reticulum-associated
degradation and p97-dependent degradation. J Biol Chem.
286:4892–4901. 2011.PubMed/NCBI
|
|
70
|
Satoh T, Chen Y, Hu D, Hanashima S,
Yamamoto K and Yamaguchi Y: Structural basis for oligosaccharide
recognition of misfolded glycoproteins by OS-9 in ER-associated
degradation. Mol Cell. 40:905–916. 2010.PubMed/NCBI
|
|
71
|
Christianson JC, Shaler TA, Tyler RE and
Kopito RR: OS-9 and GRP94 deliver mutant alpha1-antitrypsin to the
Hrd1-SEL1L ubiquitin ligase complex for ERAD. Nat Cell Biol.
10:272–282. 2008.PubMed/NCBI
|
|
72
|
Fujimori T, Kamiya Y, Nagata K, Kato K and
Hosokawa N: Endoplasmic reticulum lectin XTP3-B inhibits
endoplasmic reticulum-associated degradation of a misfolded
α1-antitrypsin variant. FEBS J. 280:1563–1575. 2013.PubMed/NCBI
|
|
73
|
Schaafhausen A, Rost S, Oldenburg J and
Muller CR: Identification of VKORC1 interaction partners by
split-ubiquitin system and coimmunoprecipitation. Thromb Haemost.
105:285–294. 2011.PubMed/NCBI
|
|
74
|
Mikami K, Yamaguchi D, Tateno H, Hu D, Qin
SY, Kawasaki N, Yamada M, Matsumoto N, Hirabayashi J, Ito Y and
Yamamoto K: The sugar-binding ability of human OS-9 and its
involvement in ER-associated degradation. Glycobiology. 20:310–321.
2010.PubMed/NCBI
|
|
75
|
Yamaguchi D, Hu D, Matsumoto N and
Yamamoto K: Human XTP3-B binds to alpha1-antitrypsin variant null
(Hong Kong) via the C-terminal MRH domain in a glycan-dependent
manner. Glycobiology. 20:348–355. 2010.PubMed/NCBI
|
|
76
|
Nagasawa K, Higashi T, Hosokawa N, Kaufman
RJ and Nagata K: Simultaneous induction of the four subunits of the
TRAP complex by ER stress accelerates ER degradation. EMBO Rep.
8:483–489. 2007.PubMed/NCBI
|
|
77
|
Moussavi-Harami SF, Annis DS, Ma W, Berry
SM, Coughlin EE, Strotman LN, Maurer LM, Westphall MS, Coon JJ,
Mosher DF and Beebe DJ: Characterization of molecules binding to
the 70 K N-terminal region of fibronectin by IFAST purification
coupled with mass spectrometry. J Proteome Res. 12:3393–3404.
2013.PubMed/NCBI
|
|
78
|
Huttlin EL, Ting L, Bruckner RJ, Gebreab
F, Gygi MP, Szpyt J, Tam S, Zarraga G, Colby G, Baltier K, et al:
The BioPlex network: A systematic exploration of the human
interactome. Cell. 162:425–440. 2015.PubMed/NCBI
|
|
79
|
Jung CH, Kim YH, Lee K and Im H: Retarded
protein folding of the human Z-type α1-antitrypsin
variant is suppressed by Cpr2p. Biochem Biophys Res Commun.
445:191–195. 2014.PubMed/NCBI
|
|
80
|
Huang CH, Hsiao HT, Chu YR, Ye Y and Chen
X: Derlin2 protein facilitates HRD1-mediated retro-translocation of
sonic hedgehog at the endoplasmic reticulum. J Biol Chem.
288:25330–25339. 2013.PubMed/NCBI
|
|
81
|
Fan W, Cheng J, Zhang S and Liu X: Cloning
and functions of the HBxAg-binding protein XBP1. Mol Med Rep.
7:618–622. 2013.PubMed/NCBI
|
|
82
|
Wang J, Huo K, Ma L, Tang L, Li D, Huang
X, Yuan Y, Li C, Wang W, Guan W, et al: Toward an understanding of
the protein interaction network of the human liver. Mol Syst Biol.
7:5362011.PubMed/NCBI
|
|
83
|
Feng L, Zhang J, Zhu N, Ding Q, Zhang X,
Yu J, Qiang W, Zhang Z, Ma Y, Huang D, et al: Ubiquitin ligase
SYVN1/HRD1 facilitates degradation of the SERPINA1 Z
variant/α-1-antitrypsin Z variant via SQSTM1/p62-dependent
selective autophagy. Autophagy. 13:686–702. 2017.PubMed/NCBI
|
|
84
|
Huttlin EL, Bruckner RJ, Paulo JA, Cannon
JR, Ting L, Baltier K, Colby G, Gebreab F, Gygi MP, Parzen H, et
al: Architecture of the human interactome defines protein
communities and disease networks. Nature. 545:505–509.
2017.PubMed/NCBI
|
|
85
|
Wen JH, Wen H, Gibson-Corley KN and Glenn
KA: FBG1 is the final arbitrator of A1AT-Z degradation. PLoS One.
10:e1355912015.
|
|
86
|
Dersh D, Jones SM, Eletto D, Christianson
JC and Argon Y: OS-9 facilitates turnover of nonnative GRP94 marked
by hyperglycosylation. Mol Biol Cell. 25:2220–2234. 2014.PubMed/NCBI
|
|
87
|
Khodayari N, Wang RL, Marek G, Krotova K,
Kirst M, Liu C, Rouhani F and Brantly M: SVIP regulates Z variant
alpha-1 antitrypsin retro-translocation by inhibiting ubiquitin
ligase gp78. PLoS One. 12:e1729832017.
|
|
88
|
Xiao Y, Han J, Wang Q, Mao Y, Wei M, Jia W
and Wei L: A novel interacting protein SERP1 regulates the N-Linked
glycosylation and function of GLP-1 receptor in the liver. J Cell
Biochem. 118:3616–3626. 2017.PubMed/NCBI
|
|
89
|
Kadowaki H, Nagai A, Maruyama T, Takami Y,
Satrimafitrah P, Kato H, Honda A, Hatta T, Natsume T, Sato T, et
al: Pre-emptive quality control protects the ER from protein
overload via the proximity of ERAD components and SRP. Cell Rep.
13:944–956. 2015.PubMed/NCBI
|
|
90
|
Pankow S, Bamberger C, Calzolari D,
Martinez-Bartolome S, Lavallee-Adam M, Balch WE and Yates JR III:
∆F508 CFTR interactome remodelling promotes rescue of cystic
fibrosis. Nature. 528:510–516. 2015.PubMed/NCBI
|
|
91
|
Kadowaki H, Satrimafitrah P, Takami Y and
Nishitoh H: Molecular mechanism of ER stress-induced pre-emptive
quality control involving association of the translocon, Derlin-1,
and HRD1. Sci Rep. 8:73172018.PubMed/NCBI
|
|
92
|
Giurato G, Nassa G, Salvati A, Alexandrova
E, Rizzo F, Nyman TA, Weisz A and Tarallo R: Quantitative mapping
of RNA-mediated nuclear estrogen receptor β interactome in human
breast cancer cells. Sci Data. 5:1800312018.PubMed/NCBI
|
|
93
|
Yu S, Ito S, Wada I and Hosokawa N:
ER-resident protein 46 (ERp46) triggers the mannose-trimming
activity of ER degradation-enhancing α-mannosidase-like protein 3
(EDEM3). J Biol Chem. 293:10663–10674. 2018.PubMed/NCBI
|
|
94
|
Lamriben L, Oster ME, Tamura T, Tian W,
Yang Z, Clausen H and Hebert DN: EDEM1's mannosidase-like domain
binds ERAD client proteins in a redox-sensitive manner and
possesses catalytic activity. J Biol Chem. 293:13932–13945.
2018.PubMed/NCBI
|