Introduction
According to 2018 global cancer statistics, there
are 18.1 million new cancer cases, of which 9.5 million are in men
and 8.6 million are in women. In 2018, ~2.1 million women were
newly diagnosed with breast cancer, accounting for ~25% of all
cancer cases in women, which far exceeded the proportion of other
types of cancer (1). In patients
with breast cancer, 15–30% of patients overexpress the human
epidermal growth factor receptor-2 (HER-2) (2). HER-2+ breast cancer is
considered the most dangerous subtype due to its invasiveness, poor
differentiation, high risk of recurrence, insensitivity to
conventional chemoradiotherapy and poor prognosis (3). Trastuzumab is currently the most
effective targeted drug used to treat HER-2+ breast
cancer, and it has greatly improved the survival rate of these
patients (4). However, clinically,
most patients who receive trastuzumab exhibit primary resistance
during the initial stage of treatment, or, even if they initially
respond to the drug, they will acquire resistance within 1 year of
treatment, which decreases the treatment effectiveness and can
result in disease progression (5).
Therefore, the mechanism of trastuzumab resistance must be
ascertained to increase the efficacy of targeted drug therapy and
improve the prognosis and survival of patients with breast
cancer.
Type II insulin-like growth factor receptor
(IGF-IIR) is a member of the insulin-like growth factor (IGF)
protein family that binds IGF-II. Since the IGF-IIR lacks tyrosine
kinase activity, it can degrade IGF-II and reduce the binding of
IGF-II and IGF-IR to promote anti-proliferative and pro-apoptotic
activities; therefore, it is considered a tumor suppressor gene
(6). On the other hand, IGF-IIR can
bind proteins that contain a mannose 6-phosphate sugar group and
participate in lysosomal transport (7). Overexpression of the IGF-IIR gene
increases the secretion of lysosomal cathepsin D in MCF-7 breast
cancer cells, promotes metastasis of breast cancer cells and
decreases the disease-free survival time of patients (8). However, the effect of IGF-IIR on
HER-2+ breast cancer prognosis remains unclear.
CRISPR/Cas9 technology is based on the short
palindrome repeat sequences with regular intervals of clusters
found in Archaea. Compared with traditional gene editing
technologies, CRISPR/Cas9 can significantly improve the efficiency
of gene integration and significantly shorten the homologous arm
length of mediated homologous recombination, which is conducive to
subsequent detection (9). For
example, Ruan et al (10)
used CRISPR/Cas9 technology to successfully knock in a 9.4-kb
fragment of pig H11 site, with a homology arm of only 800 bp, which
reduces the difficulty of amplification and the rate of mismatches.
Moreover, it is simple to operate, has low cost and a short cycle
time. On the other hand, adeno-associated virus integration site 1
(AAVS1), as a major hotspot for AAV integration, intron 1 of the
protein phosphatase 1, is a regulatory subunit 12C (PPP1R12C) gene
on human chromosome 19. This locus allows for stable, long-term
transgene expression in numerous cell types, including embryonic
stem cells. As disruption of PPP1R12C is not associated with any
known diseases, the AAVS1 locus is often considered a safe harbor
for transgene targeting (11). AAVS1
is an exemplary locus within the PPP1R12C gene that permits robust
expression of CAG promoter-driven transgenes (11). In the present study, CRISPR/Cas9
technology was used to mediate IGF-IIR gene knock-in at the safe
harbor of AAVS1 in order to construct a HER-2 positive breast
cancer cell line SKBR3 that overexpresses IGF-IIR, providing a cell
model to determine the influence of IGF-IIR overexpression and the
potential to explore the resistance of HER-2+ breast
cancer cells to trastuzumab.
Materials and methods
Materials
The SKBR3 cell line was purchased from BeNa Culture
Collection; Beijing Beina Chunglian Biotechnology Research
Institute. Cells were screened periodically for mycoplasma
contamination using the Universal Mycoplasma Detection kit
(American Type Culture Collection), which was used according to the
manufacturer's protocol. The Universal CRISPR Activity (UCA)™
CRISPR/Cas9 rapid construction and activity detection kit contains
three plasmids: Precut pCS, precut pUCA(Luc), and pCS-Positive,
which can be used to construct Cas9/sgRNA, identify and screen
sgRNA. As well as the human (h)AAVS1-KI vector, were purchased from
Beijing Biocytogen Co., Ltd. The FlexiGene® DNA gene
extraction kit was purchased from Qiagen GmbH, and the GeneRuler™ 1
kb plus DNA Ladder, T4 DNA ligase, EcoRI, ScaI,
NcoI, BglII, Bstz17I+KpnI+ScaI
and AsiSI+KpnI were purchased from Thermo Fisher
Scientific, Inc. The TIANprep Mini Plasmid Kit and Top10 competent
cells were purchased from Tiangen Biotech Co., Ltd. McCoy's 5A+10%
FBS+1% L-G+1% NE A+1% SP+1% HEPES was purchased from Gibco; Thermo
Fisher Scientific, Inc.
Cell culture
SKBR3 cells were removed from liquid nitrogen,
rapidly thawed in a water bath at room temperature, and transferred
to a 15-ml tube for centrifugation at 150 × g for 3 min at room
temperature. After removing the supernatant, 1 ml complete medium
(McCoy's 5A medium with 10% FBS, 1% L-G, 1% Non-Essential Amino
Acids, 1% sodium pyruvate and 1% HEPES) was added to the pellet and
mixed by pipetting, after which the cell suspension was inoculated
in 10 ml complete medium. The cells were cultured at 37°C in a 5%
CO2 incubator, and the medium was changed every other
day. When the cells were in the logarithmic growth stage, the
medium was discarded, and cells were washed with PBS once and
digested using 0.25% trypsin at 37°C for 2 min. After gently
pipetting, the medium was transferred to a new 15-ml tube for
centrifugation at 150 × g for 3 min at room temperature, the
supernatant was discarded, and 1 ml complete medium was added.
After gentle pipetting to mix, the cells were counted and
inoculated into 24-well plates at a concentration of
1×106 cells/well for further culture.
Construction of the Cas9/single guide
(sg)RNA plasmid
Using NCBI Primer BLAST (http://www.ncbi.nim.nih.gov/BLAST/), primers were
designed 400 bp upstream or downstream of the AAVS1 (upstream
primer sequence, 5′-GCATCAAGCTTGGTACCGAT-3′ and downstream primer
sequence, 5′-ACTTAATCGTGGAGGATGAT-3′). DNA from the SKBR3
HER-2+ breast cancer cells was extracted using the
FlexiGene DNA gene extraction kit, and the target site sequence was
amplified using this DNA as a template, aforementioned primers and
PrimesSTAR HS DNA Polymerase, named MSD. PCR reaction conditions
were pre-denaturation at 94°C for 5 min; 94°C denaturation for 30
sec, annealing at 67°C for 30 sec and extension at 68°C for 1 min,
for 15 cycles; denaturation 94°C for 30 sec, annealing at 56°C for
30 sec and extension at 68°C for 1 min, for 25 cycles; extension at
68°C for another 10 min maintained at 4°C. Sequencing confirmed
whether the target was consistent with the AAVS1 gene reference
sequence in GenBank (https://www.ncbi.nlm.nih.gov/), and the sequencing was
completed by Invitrogen; Thermo Fisher Scientific, Inc. The
full-length genome sequence of human AAVS1 (Gene ID, 54776) was
obtained from GenBank, and six pairs of complementary sgRNA
sequences were designed at the AAVS1 target site using the sgRNA
Guide Design Resources website (http://crispr.mit.edu/), where the following
parameters were selected: CRISPOR tool, where the AAVS1 sequence
was first copied; human species; and spCAS9 type. After submission,
the sequence with the highest specificity scores were selected as
sgRNAs and named sgRNA1-6, where the italic lowercase letters in
Oligo are the sticky ends produced by annealing (Table I). According to the Universal CRISPR
Activity (UCA)™ CRISPR/Cas9 rapid construction and activity
detection kit manufacturer's protocol, sgRNA oligos were
denatured-annealed to form a double chain and linked to the
linearized pCS plasmid vector according to the manufacturer's
protocol (Biocytogen). After confirming the sequence of the
resulting construct, named pCS-sgRNA1-6, the activities of the
sgRNAs were tested by the Universal CRISPR Activity (UCA)™
CRISPR/Cas9 rapid construction and activity detection kit to detect
the level of luciferase activity. Luciferase activity is positively
correlated with sgRNA activity. The most efficient sgRNA pair was
selected for use in subsequent gene editing experiments.
 | Table I.sgRNA design. |
Table I.
sgRNA design.
| sgRNA | Sequence
(5′-3′) |
|---|
| sgRNA1 | F: cacc
GGAAGGAGGAGGCCTAAGGA |
|
| R: aaac
TCCTTAGGCCTCCTCCTTCC |
| sgRNA2 | F: cacc
GTCACCAATCCTGTCCCTAG |
|
| R: aaac
CTAGGGACAGGATTGGTGAC |
| sgRNA3 | F: cacc
GGGGCCACTAGGGACAGGAT |
|
| R: aaac
ATCCTGTCCCTAGTGGCCCC |
| sgRNA4 | F: cacc
GCACCCCACAGRGGGGCCACT |
|
| R: aaac
AGTGGCCCCACTGTGGGGTGC |
| sgRNA5 | F: cacc
GTCCCCTCCACCCCACAGTG |
|
| R: aaac
CACTGTGGGGTGGAGGGGAC |
| sgRNA6 | F: cacc
GGTTAATGTGGCTCTGGTTC |
|
| R: aaac
GAACCAGAGCCACATTAACC |
Construction of the IGF-IIR target
vector
The IGF-IIR coding sequence (CDS) (https://www.ncbi.nlm.nih.gov/nuccore/NM_000876.3)
fragment was divided into two segments (A1 and A2), and A1 was
digested using Bstz17I, KpnI and ScaI. After
30 min of enzyme digestion at 37°C, 1% agarose gel electrophoresis
was performed, and 3,104-bp bands were recovered. A2 was digested
using AsiSI and KpnI, with 4,386-bp bands recovered
following electrophoresis. AsiSI and Bstz17I were
used for enzyme digestion of the hAAVS1-KI vector (1 kb upstream
homology arm, a strong promoter CAG, WPRE regulatory element, ploy
A, puromycin resistance gene, 1 kb downstream homology arm), and 1%
agarose gel electrophoresis was conducted to recover bands of
~11,232 bp. T4 DNA ligase was used to connect each recovered
fragment. After connecting for 30 min at room temperature, the
product was transformed into top 10 competent cells for 90 sec at
42°C and cultured overnight. Plasmids were extracted using the
TIANprep Mini Plasmid kit, identified using EcoRI and
ScaI, and NcoI and BglII restriction
endonucleases, and subsequently sequenced by Invitrogen Shanghai
Trading Co., Ltd. Sequencing primers for M13 were: Forward, is
5′-GTAAAACGACGGCCAGT-3′ and reverse, 5′-CAGGAAACAGCTATGAC-3′.
Cell transfection and screening
According to the Invitrogen Neon electroporation
instrument and the matching electroporation kit (cat. no. MPK10096;
Invitrogen; Thermo Fisher Scientific, Inc.) manufacturer's
protocol, four sets of electrical parameters were used for cell
transfection: i) 1,050 V, 30 ms and one pulse; ii) 1,000 V, 40 ms
and one pulse; iii) 1,100 V, 30 ms and two pulses; and iv) 1,200 V,
20 ms and two pulses. Next, transfection efficiency was analyzed
using flow cytometry (FACSCalibur; BD Bioscience) and FlowJo 7.6
analysis software, and detected the expression of GFP with a 488-nm
laser. SKBR3 antibiotic screening concentrations were determined by
treating cells with puromycin at concentrations of 0.2, 0.5, 1.0,
1.5 and 2.0 µg/ml at 37°C and observing cell death under an
inverted phase contrast microscope at day 7. The IGF-IIR target
vector and pCS-sgRNA plasmid were transferred to SKBR3 cells with a
mass ratio of 1:2 (2.4 µg/100 µl:4.8 µg/100 µl) under optimal
transfection conditions (1,200 V, 20 ms and 2 pulses), and then
puromycin at the optimal concentration (0.5 µg/ml) was added after
2 days. The cells were cultured at 37°C in a 5% CO2
incubator, and the recovery medium used McCoy′5A+10%FBS+1%L-G+1%
SP+1%NEAA+1%HEPES. After 7 days of drug exposure at 37°C, the cells
were examined via PCR, and monoclonal cell screening was
performed.
Identification of mixed clone
genotypes
DNA was extracted from the mixed cells according to
the FlexiGene® DNA gene extraction kit instructions and
used for PCR. The PCR reaction conditions were as follows:
Pre-denaturation at 94°C for 2 min; 98°C denaturation for 10 sec,
annealing at 67°C for 30 sec, and extension at 68°C for 1 min for
15 cycles; denaturation at 98°C for 10 sec, annealing at 56°C for
30 sec and extension at 68°C for 1 min, for 25 cycles; extension at
68°C for another 10 min and maintained at 4°C. One primer
(P1) targeted the lateral side of the left homologous
arm, and another primer (P2) targeted the CAG promoter,
resulting in an amplification product of 2,307 bp. Primer
P3 targeted the IGF-IIR gene, and another primer
(P4) targeted the poly-A tail. This primer pair was used
to confirm integration of the exogenous gene IGF-IIR with an
amplified product size of 8,555 bp. The P5 primer
targeted the puromycin resistance gene, and primer P6
targeted the lateral side of the right homologous arm. This pair of
primers was used to identify recombination in the right homologous
arm, and produced an amplification product of 2,789 bp. Primer
sequences are shown in Table II.
After identification, single cell cloning was performed.
| Table II.Primer sequences used for PCR
identification. |
Table II.
Primer sequences used for PCR
identification.
| Primer | Sequence
(5′-3′) |
|---|
| P1 |
CTATGCTGACACCCCGTCCCAGTC |
| P2 |
TCGTTGGGCGGTCAGCCAGG |
| P3 |
AGCTGAGAGTAGCACAATCTAGGCGTC |
| P4 |
GAGCCTCTGCTAACCATGTTC |
| P5 |
GCAACCTCCCCTTCTACGAGC |
| P6 |
CTAAGAACTTGGGAACAGCCACAGC |
Single cell cloning
The mixed clones were centrifuged at 150 × g for 3
min at room temperature and added to the culture medium. A
monoclonal preparation was obtained using the semi-solid and finite
dilution methods (12,13). The semi-solid medium was thawed at
4°C overnight, left at room temperature for 10–15 min before the
addition of 400 µl complete medium, subjected to vortex oscillation
for 4–5 sec, and added to 6-well plates. The remaining semi-solid
medium was added to the cell suspension, mixed and cultured in
6-well plates, which was placed in a 37°C incubator for 16 days.
For selective cloning, 50 µl/well trypsin was added to 96-well
plates, which were placed in an incubator at 37°C for digestion for
2 min. Subsequently, 100 µl complete medium was added to each well
to terminate digestion, gently pipetting, and to initiate culture.
The cell suspension was diluted to a concentration of 1 cell/100 µl
with complete medium using the finite dilution method. After
mixing, the cells were divided into 96-well plates with 100 µl
complete medium added to each well for monoclonal culture.
Comparison between the proliferation
of SKBR3 and mixed clone cells
A cell suspension of SKBR3 or mixed cloned cells was
inoculated in a 96-well culture plate (6,000 cells/well) and
cultured at 37°C in a 5% CO2 incubator, and observed the
SKBR3 and mixed clone cells under an inverted phase contrast
microscope. After culturing for 1–5 days, according to a Cell
Counting Kit-8 (CCK-8) manufacturer's instructions (Beyotime
Institute of Biotechnology), 10 µl/well CCK-8 reagent was added.
After mixing, cells were incubated for 2 h at 37°C, and the
absorbance was measured at 450 nm. The growth curves were plotted
with the culture time as the abscissa and the absorbance as the
ordinate.
Statistical analysis
The activity of the Cas9/sgRNA plasmid was
determined using SPSS 22.0 software (IBM Corp.). The data obtained
from three replicates were analysis of variance to determine the
mean ± standard error of each group. The control group was used as
the standard to calculate the relative sgRNA activity of each
group.
Results
Plasmid construction and assessment of
Cas9/sgRNA activity
The SKBR3 genome was used as a template to amplify
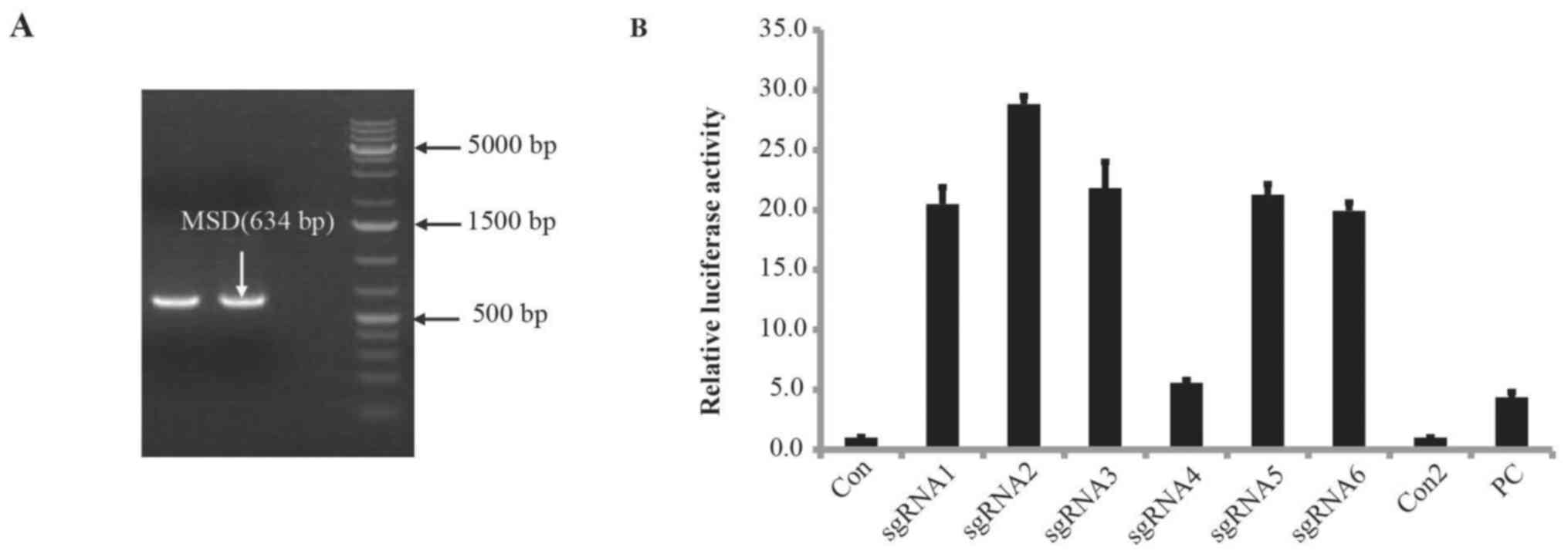
the target sequence AAVS1 with a band size of 634 bp, as shown in
Fig. 1A. Sequencing results
confirmed that the target sequence was consistent with the
reference sequence in GenBank; therefore, sgRNAs were designed
according to this sequence information (data not shown). Next, six
20-bp sgRNAs were designed to target the AAVS1 gene sequence and
the PAM sequence was NGG (Table
III). A Cas9/sgRNA plasmid acting on the AAVS1 gene was
successfully constructed, and a UCA™ CRISPR/Cas9 activity detection
kit was used to assess sgRNA activity. This assay revealed that
sgRNA2 was the most efficient (Fig.
1B).
| Table III.sgRNA sequences. |
Table III.
sgRNA sequences.
| sgRNA sequence
number | Sequence
(5′-3′) | Length |
|---|
| sgRNA1 |
ggaaggaggaggcctaagga tgg | 20 bp + PAM |
| sgRNA2 |
gtcaccaatcctgtccctag tgg | 20 bp + PAM |
| sgRNA3 |
ggggccactagggacaggat tgg | 20 bp + PAM |
| sgRNA4 |
caccccacagtggggccact agg | 20 bp + PAM |
| sgRNA5 |
gtcccctccaccccacagtg ggg | 20 bp + PAM |
| sgRNA6 |
ggttaatgtggctctggttc tgg | 20 bp + PAM |
Enzyme digestion and identification of
IGF-IIR target vectors
The target gene of IGF-IIR was digested using
AsiSI and Bstz17I, and ligated into the hAAVS1-KI
vector to construct the IGF-IIR target vector. The schematic
diagram is displayed in Fig. 2A.
IGF-IIR target vectors were identified by double enzyme digestion
using EcoRI and Scal, and agarose gel electrophoresis
revealed five bands with sizes of 7,280, 4,913, 3,003, 2,124 and
1,402 bp. After Ncol enzyme digestion, five bands with sizes
of 6,537, 5,174, 3,216, 2,388 and 1,407 bp were observed. After
BglII enzyme digestion, three bands of the expected sizes
(10,473, 5,324 and 2,925 bp) were observed following
electrophoresis (Fig. 2B). The
restriction sites are shown in Fig.
2C. The IGF-IIR target vector was confirmed by sequencing
(Fig. 2D).
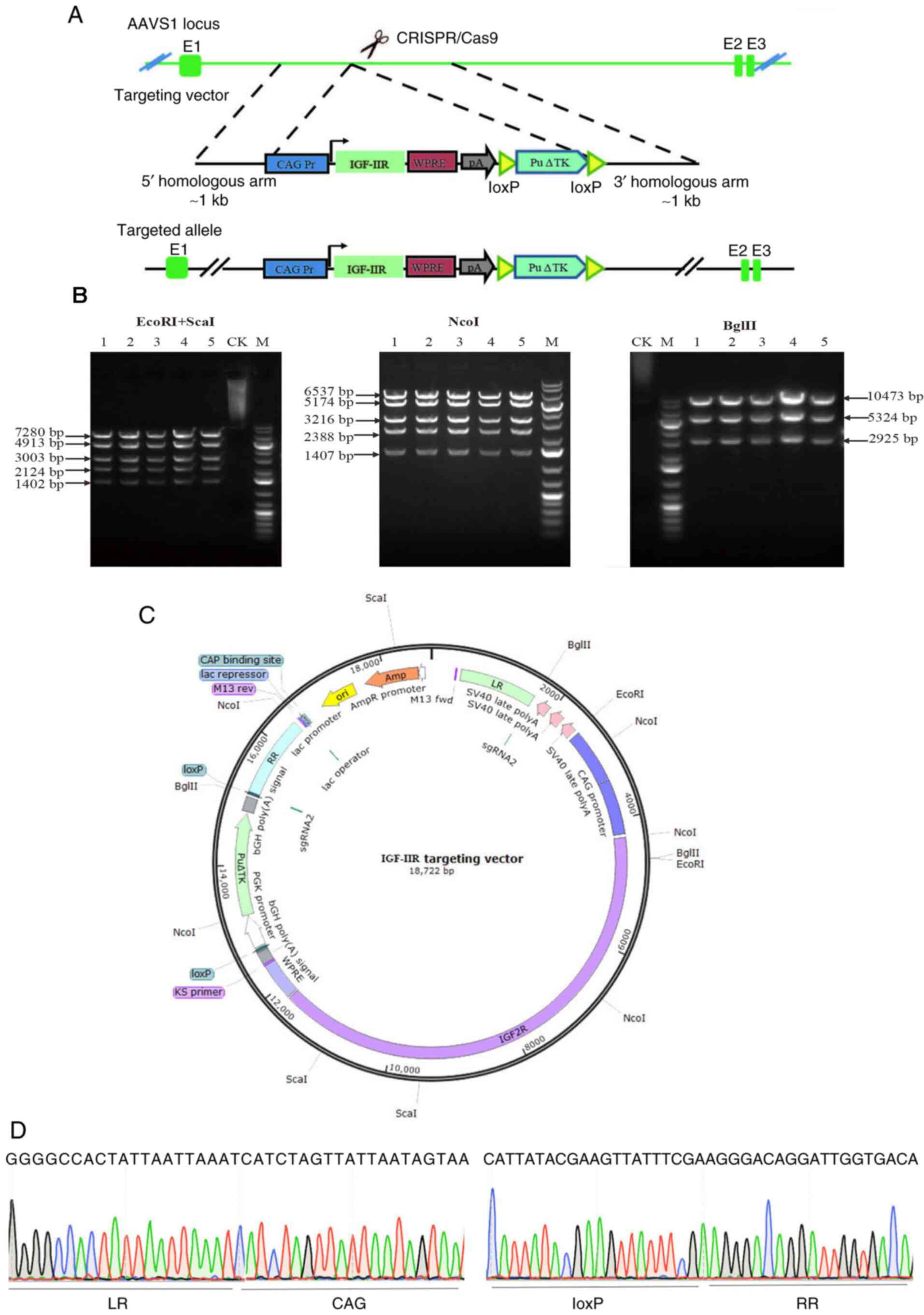 | Figure 2.Schematic diagram of the IGF-IIR
targeting vector, confirmation of the IGF-IIR targeting vector and
target vector sequencing. (A) Schematic diagram of the IGF-IIR
targeting vector, including the homologous arms, CAG promoter,
IGF-IIR coding sequence, Woodchuck hepatitis virus
post-transcriptional regulatory element, poly A and puromycin. (B)
IGF-IIR targeting vector confirmed via enzyme digestion (plasmid
numbers 1–5). (C) Schematic diagram of restriction sites of IGF-IIR
targeting vector. (D) LR provides a partial sequencing diagram
connecting the left homology arm to the CAG promoter; RR is a
partial sequencing diagram connecting loxP to the right homology
arm. CK, undigested control; M, 1-kb DNA ladder; AAVS1,
adeno-associated virus integration site 1; IGF-IIR, type II
insulin-like growth factor receptor; LR, upstream homology arm; RR,
downstream homology arm. |
Cell transfection and mixed clone
genotyping
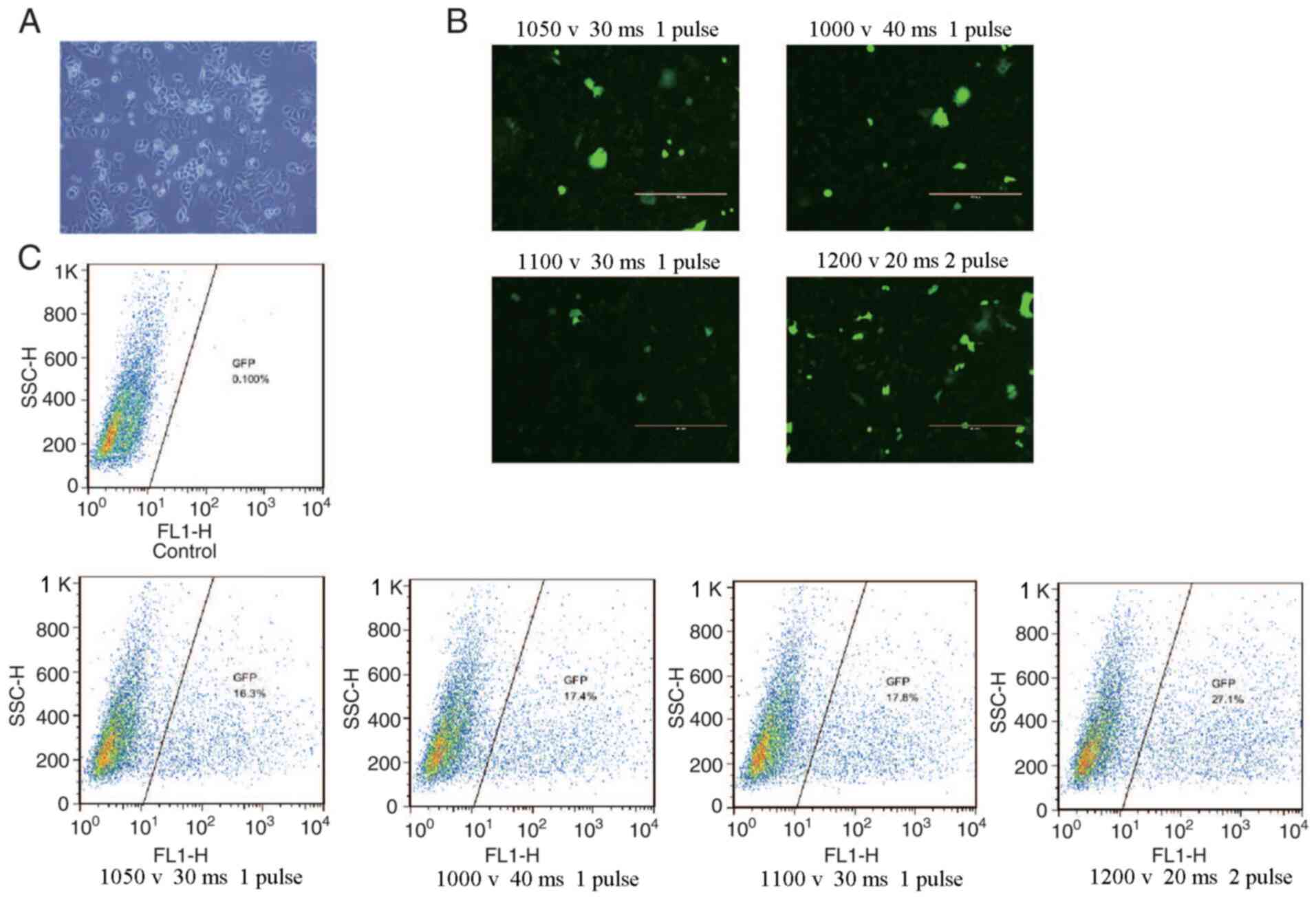
Flow cytometry was used to determine the
transfection efficiency of green fluorescent protein particles
under different electrotransfer parameters, as indicated in
Fig. 3. The results revealed that
the highest transfection efficiency was 27.1% obtained at 1,200 V,
20 ms and 2 pulses at 90% cell viability. The cells were treated
with different concentrations of puromycin for 7 days, and the
optimal concentration of puromycin was determined to be 0.5 µg/ml
(data not shown). The pCS-sgRNA2 plasmid with the highest activity
and IGF-IIR target vector were transferred to SKBR3 cells at 1,200
V, 20 ms and 2 pulses, and the cells were collected after screening
with 0.5 µg/ml puromycin. The genome of the mixed cells was then
extracted and used as a template for PCR identification. The
identification method is shown in Fig.
4A. As expected, the bands of P1 and P2
corresponded to 2,307 bp, those of P3 and P4
to 8,555 bp, and those of P5 and P6 to 2,789
bp (Fig. 4B). The mixed clones were
confirmed by sequencing (Fig. 4C).
The results revealed homologous recombination of the IGF-IIR gene
with the AAVS1 gene target, and single cell cloning was used for
subsequent experiments.
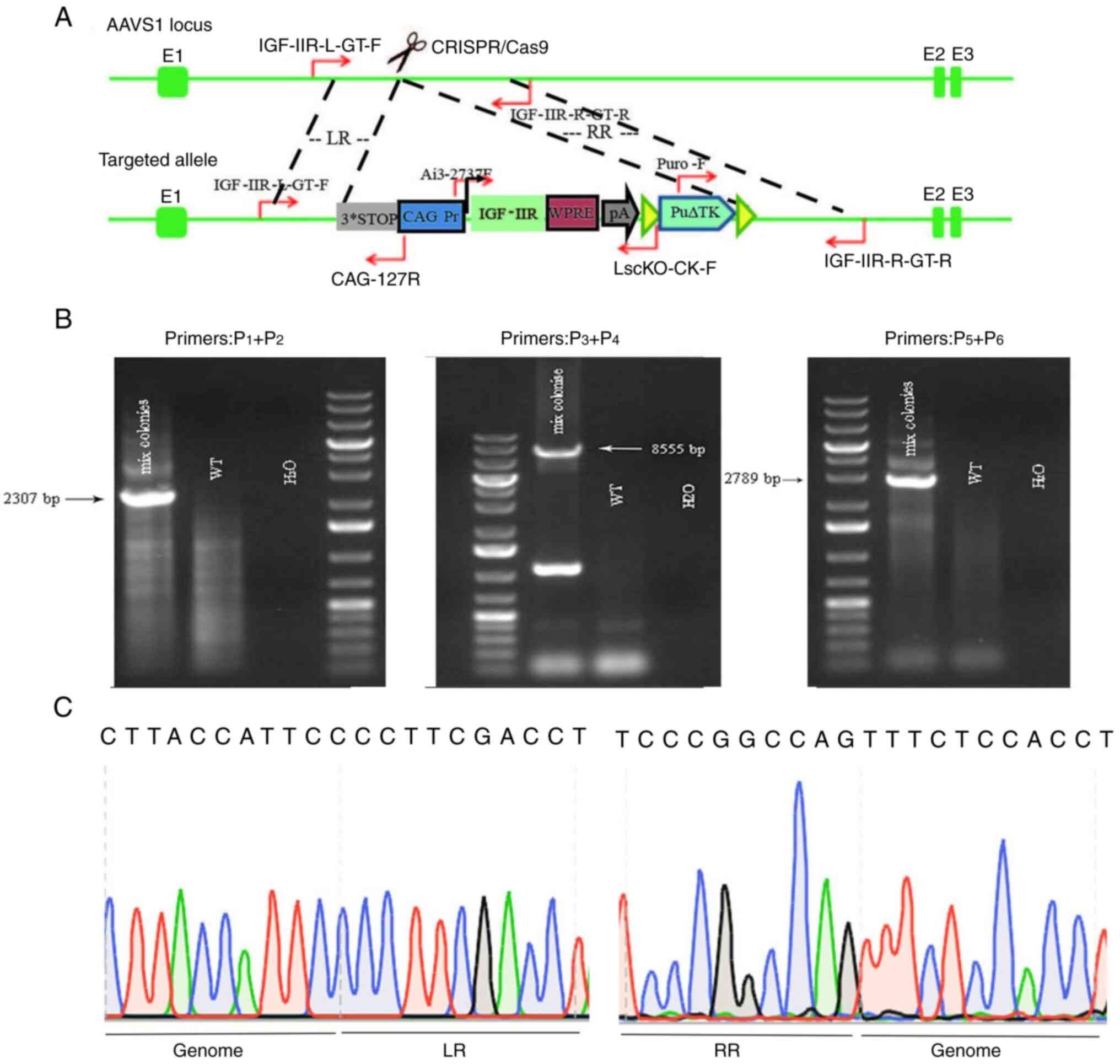 | Figure 4.Mixed clone identification procedure,
determination of mixed clone genotypes and mixed clone fragment
sequencing. (A) Mixed clone map showing primer positions for
identification via PCR. IGF-IIR-L-GT-F represents primer
P1 located upstream of LR. CAG-127R represents primer
P2 located at the CAG promoter. Ai3-2737F represents
primer P3 located at IGF-IIR. LscKO-CK-F represents
primer P4 located at ploy A. Puro-F represents primer
P5 located at the puromycin resistance gene.
IGF-IIR-R-GT-R represents primer P6 located downstream
of the RR. (B) Schematic diagram for PCR identification of three
sets of primer mixed clones (WT cells were used as a negative
control; H2O was used as a blank control). Mixed cloning
primer P1 is located upstream of LR and P2 is
located at the CAG promoter, amplifying a fragment of 2,307 bp;
P3 is located at IGF-IIR and P4 is located at
plot A, amplifying a fragment of 8,555 bp; P5 is located
at the puromycin resistance gene and P6 is located
downstream of the RR, amplifying a fragment of 2,789 bp. WT,
wild-type; IGF-IIR, type II insulin-like growth factor receptor.
(C) Schematic diagram of mixed clone sequencing; LR, upstream
homology arm; RR, downstream homology arm. |
Monoclonal preparation
After 19 days of culture in semi-solid medium, the
cell mass was small, and cell proliferation was slow. After 22 days
of culture, monoclonal cells were prepared using the limited
dilution method. The number of cloned cells was low (~20), their
proliferation was extremely slow, and gradually underwent apoptosis
(data not shown). Therefore, the cells were not cultured
further.
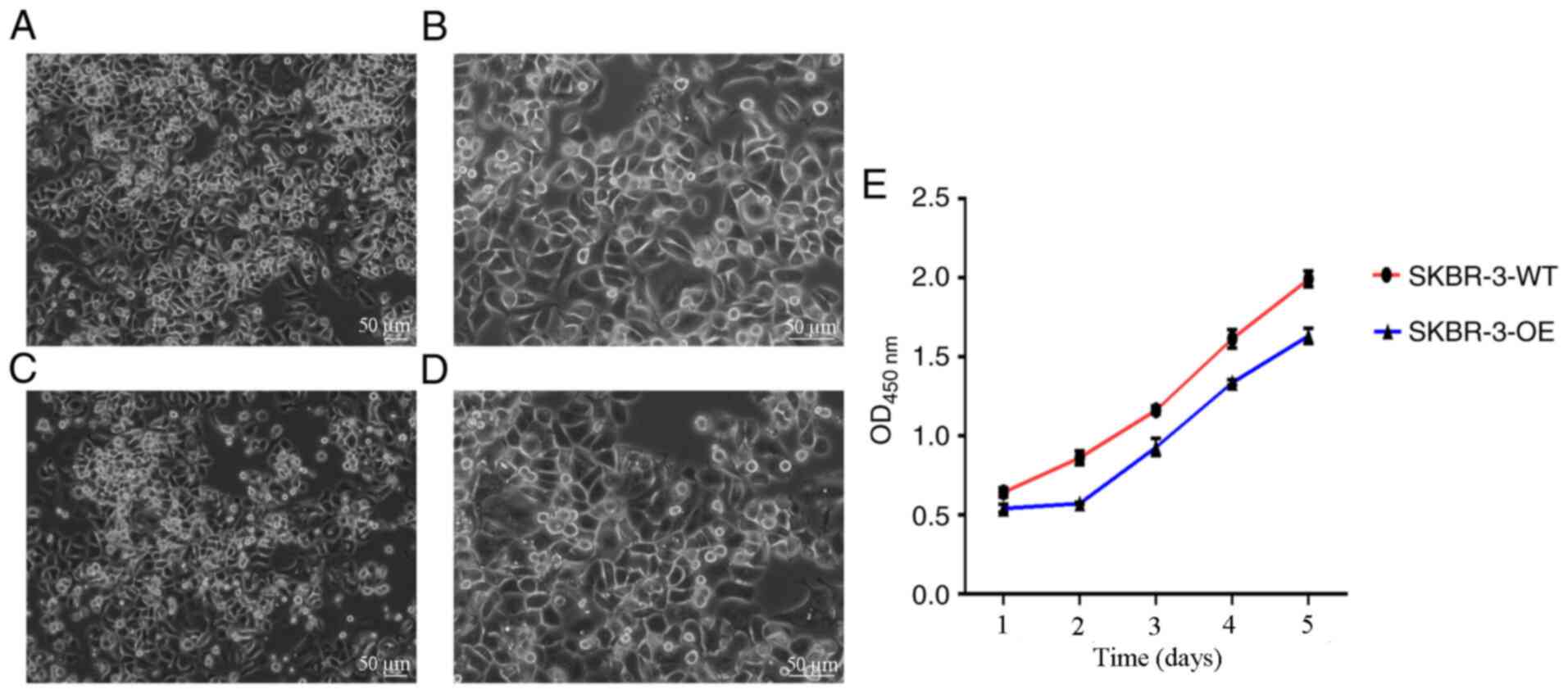
Proliferation of SKBR3 and mixed clone
cells
It was observed under an inverted phase contrast
microscope that SKBR3 and mixed clone cells were full in morphology
and uniform in proliferation. The CCK-8 method was used to detect
cell proliferation of SKBR3 cells (SKBR-3-WT) and mixed clone cells
(SKBR-3-OE). The results revealed that the proliferation of
SKBR-3-OE was markedly lower compared with SKBR-3-WT (Fig. 5).
Discussion
IGF-IIR is a multifunctional receptor capable of
binding to the mitotic peptide IGF-II and inhibiting tumor growth
by degrading IGF-II to regulate its extracellular levels (14). According to previous studies, the
loss of IGF-IIR heterozygosity at the 6q26-27 gene locus can result
in the development of multiple types of tumor, including breast
cancer, hepatocellular carcinoma and lung squamous cell carcinoma
(8,15,16),
confirming that IGF-IIR may be involved in inhibiting tumor growth.
Furthermore, Lee et al (17)
reported that in MDA-MB-231 breast cancer cells, increased IGF-IIR
expression inhibited cell invasion and migration, and inhibited
tumor growth in mouse models. Similarly, in vivo and study
by Souza et al (18) have
revealed that overexpression of IGF-IIR in breast cancer cells
decreases the growth rate of cancer cells and in certain cases
promotes cell death, while IGF-IIR silencing increases cell
proliferation and survival. Devi et al (19) demonstrated that IGF-II can promote
cell proliferation, inhibit apoptosis and enhance local
infiltration and metastasis in breast tumors. Due to the lack of
IGF-IIR tyrosine kinase activity, it cannot transmit intracellular
mitotic signals. IGF-IR is mainly used as a buffer for the
biological activity of IGF-II and its effects, and it can degrade
IGF-II and reduce its interaction with IGF-IR combines to exert
anti-proliferation and pro-apoptotic activity (20). Specifically, IGF-IR can activate the
PI3K/Akt/MAPK/FAK signaling pathways to effectively regulate cell
proliferation, anti-apoptosis mechanisms and drug resistance
(21).
In this study, the molecular typing of SKBR3 cells
is estrogen receptor (ER)−, progesterone receptor
(PR)− and HER-23+, so they do not express ER
or PR, but highly express the HER-2 gene (22). Using this cell type helps to exclude
the impact of ER and PR on the experimental results. Additionally,
the Cancer Cell Line Encyclopedia (https://portals.broadinstitute.org/ccle) and BioGPS
websites (http://biogps.org/#goto=welcome) revealed that the low
expression of IGF-IIR in SKBR3 cells is more conducive to our
construction of over-expressing cells, and it is more conducive to
compare the growth of wild-type SKBR3 and IGF-IIR over-expressed
SKBR3 cells. Therefore, in the present study, CRISPR/Cas9
technology was used to mediate the site-specific knock-in of the
IGF-IIR gene at the safe site of AAVS1 to construct a
HER-2+ breast cancer SKBR3 cell line that overexpresses
IGF-IIR, thereby providing the basis to investigate drug-resistance
in breast cancer later, for example trastuzumab.
The CRISPR/Cas9 technology is based on clusters of
regularly spaced short palindrome repeats found in Archaea. The
CRISPR gene sequence was composed of Cas protein gene, leader
sequence and CRISPR locus, of which the nuclease was Cas9
endonuclease, and DNA double-strand cutting was performed through
identification of PAM. This technology recognizes DNA sequences
with PAM sequences through sgRNA, and brings Cas9 nuclease to
specific targets on the genome to complete the cleavage of DNA
sequences at specific gene sites (23). This technology is simple and fast to
operate, with low associated costs and higher efficiency compared
with traditional gene editing technologies (9). van Diemen et al (24) used CRISPR/Cas9 technology to inhibit
the three herpes viruses EBV, HSV and HCMV to prevent the
replication of herpes viruses in latent infection and lytic
infection models. In 2018, Ophinni et al (25) used CRISPR/Cas9 to target HIV-1
regulatory genes and successfully inhibited the replication of
HIV-1 in human CD4+ T cell lines with persistent and latent
infection. In terms of breast cancer, Feng et al (26) used the CRISPR/Cas9 system to
construct an MDA-MB-231 cell line with PinX1 overexpression and
knockout, which confirmed that PinX1 overexpression inhibits the
proliferation and migration of human basal breast cancer. Low PinX1
expression is associated the malignant behavior of these cells. Mei
et al (27) successfully
designed an efficient multiplexed CRISPR/dCas9 system using
CRISPR/Cas9 technology, and applied it to study the phylogenetic
relationship between breast cancer subtypes driven by cancer stems,
and clarified that Differentiating the treatment strategies of
triple-negative breast cancer and HER-2 positive breast cancer. The
CRISPR/Cas9 system has provided a plethora of ideal methods to
improve cell and animal models for studying the functions of genes
and biological progresses; therefore, it has greatly promoted the
study of various diseases in vitro and in vivo
(28).
In the present study, six sgRNAs at the AAVS1 target
site with a length of ~20 bp were designed, and the PAM sequence
was NGG. Using the CRISPOR tool to determine the specificity of the
sgRNA sequences, the sequences with high specificity, low
off-target effects and high activity were selected to ensure
reliable research results. A strong CAG promoter was used to
construct the IGF-IIR gene overexpression vector and the
transfection conditions were optimized prior to
electrotransfection. Four different sets of electrotransfection
parameters were used to transfect the green fluorescent protein
particles into SKBR3 cells. Flow cytometry analysis of the
transfection efficiency under these different parameters revealed
that the transfection efficiency was the highest using 1,200 V, 20
ms and 2 pulses at 90% cell viability. Therefore, the most
successful transfection conditions were used for subsequent
experiments to ensure the generation of a large number of
transfected cells. After successful transfection, puromycin
screening and PCR identification, a SKBR3 mixed clone cell line
with IGF-IIR site-integrated integration at the AAVS1 site was
successfully constructed.
By using the limited dilution method with monoclonal
cells, there is essentially no damage done to the cells, the
operation is simple, the technology is advanced, the cost is low,
and no special antibody reagents or equipment are required
(29). Furthermore, using a
semi-solid medium for the cell growth medium allows for the
formation of a single independent dispersed clone, which can
prevent the clone from moving, thereby reducing the workload
associated with the cell culture and experimental cycle procedure
(30). Therefore, the present study
used the aforementioned methods for monoclonal cell preparation.
However, when selecting monoclonal cells, the proliferation was
extremely slow and the cells were in poor condition. Moreover,
since the monoclonal preparation method was performed manually, the
damage to the cells was low, but the steps were cumbersome.
Compared with the coordinated growth of mixed clone cells, the
survival efficiency of a single cell is significantly reduced. It
was demonstrated that the apoptosis of monoclonal cells is
associated with the overexpression of IGF-IIR. IGF-IIR inhibits
cell proliferation to a certain extent, leading to cell apoptosis.
Future studies should aim to treat the successfully constructed
IGF-IIR mixed-site cloned cell line with trastuzumab to observe the
effects on cell proliferation, apoptosis, invasion and other
characteristics, as well as to construct trastuzumab-resistant
cells to investigate the effect of differential IGF-IIR expression
on trastuzumab-resistant cells, and explore the role of IGF-IIR in
the development of trastuzumab resistance.
In summary, the present study successfully generated
a mixed clone cell line of the IGF-IIR gene at the AAVS1 using
CRISPR/Cas9 gene editing technology which may be used to explore
the effect of IGF-IIR on trastuzumab resistance in
HER-2+ breast cancer cells.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Science and
technology project of Inner Mongolia autonomous region (grant no.
201802116).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XM conceived and designed the study, guaranteed its
integrity, performed the experiments and wrote the manuscript. RC
and HX participated in the design of the experiments, performed the
literature search, performed experimental studies and analyzed the
data. ZC designed the study, provided financial support, analyzed
and interpreted the data, and revised the manuscript. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Liao N: HER2-positive breast cancer, how
far away from the cure? On the current situation of anti-HER2
therapy in breast cancer treatment and survival of patients. Chin
Clin Oncol. 5:412016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Harbeck N: Advances in targeting
HER2-positive breast cancer. Curr Opin Obstet Gynecol. 30:55–59.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gianni L, Eiermann W, Semiglazov V, Lluch
A, Tjulandin S, Zambetti M, Moliterni A, Vazquez F, Byakhov MJ,
Lichinitser M, et al: Neoadjuvant and adju-vant trastuzumab in
patients with HER2-positive locally advanced breast cancer (NOAH):
Follow-up of a randomised controlled superiority trial with a
parallel HER2-negative cohort. Lancet Oncol. 15:640–647. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhou P, Jiang YZ, Hu X, Sun W, Liu YR, Liu
F, Luo RC and Shao ZM: Clinicopathological characteristics of
patients with HER2-positive breast cancer and the efficacy of
trastuzumab in the People's Republic of China. Onco Targets Ther.
9:2287–2295. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wong H, Leung R, Kwong A, Chiu J, Liang R,
Swanton C and Yau T: Integrating molecular mechanisms and clinical
evidence in the management of trastuzumab resistant or refractory
Her-2(+) metastatic breast cancer. Oncologist. 16:1535–1546. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pollak M: The insulin and insulin-like
growth factor receptor family in neoplasia: An update. Nat Rev
Cancer. 12:159–169. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hankins GR, De Souza AT, Bentley RC, Patel
MR, Marks JR, Iglehart JD and Jirtle RL: M6P/IGF2 receptor: A
candidate breast tumor suppressor gene. Oncogene. 12:2003–2009.
1996.PubMed/NCBI
|
|
9
|
Joung JK and Sander JD: TALENs: A widely
applicable technology for targeted genome editing. Nat Rev Mol Cell
Biol. 14:49–55. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ruan J, Li H, Xu K, Wu T, Wei J, Zhou R,
Liu Z, Mu Y, Yang S, Ouyang H, et al: Highly efficient
CRISPR/Cas9-mediated transgene knockin at the H11 locus in pigs.
Sci Rep. 5:142532015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Oceguera-Yanez F, Kim SI, Matsumoto T, Tan
GW, Xiang L, Hatani T, Kondo T, Ikeya M, Yoshida Y, Inoue H and
Woltjen K: Engineering the AAVS1 locus for consistent and scalable
transgene expression in human iPSCs and their differentiated
derivatives. Methods. 101:43–55. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Davis JM, Pennington JE, Kubler AM and
Conscience JF: A simple, single-step technique for selecting and
cloning hybridomas for the production of monoclonal antibodies.
Immunol Methods. 50:161–171. 1982. View Article : Google Scholar
|
|
13
|
Mao SJ and France DS: Enhancement of
limiting dilution in cloning mouse myeloma-spleen hybridomas by
human low density lipoproteins. J Immunol Methods. 75:309–316.
1984. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Iwamoto KS and Barber CL:
Radiation-induced posttranscriptional control of M6P/IGF2r
expression in breast cancer cell lines. Mol Carcinog. 46:497–502.
2007. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
O'Gorman DB, Weiss J, Hettiaratchi A,
Firth SM and Scott CD: Insulin-like growth factor-II/mannose
6-phosphate receptor overexpression reduces growth of
choriocarcinoma cells in vitro and in vivo. Endocrinology.
143:4287–4294. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
O'Gorman DB, Costello M, Weiss J, Firth SM
and Scott CD: Decreased insulin-like growth factor-II/mannose
6-phosphate receptor expression enhances tumorigenicity in JEG-3
cells. Cancer Res. 59:5692–5694. 1999.PubMed/NCBI
|
|
17
|
Lee JS, Weiss J, Martin JL and Scott CD:
Increased expression of the mannose 6-phosphate/insulin-like growth
factor-II receptor in breast cancer cells alters tumorigenic
properties in vitro and in vivo. Int J Cancer. 107:564–570. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Souza RF, Wang S, Thakar M, Smolinski KN,
Yin J, Zou TT, Kong D, Abraham JM, Toretsky JA and Meltzer SJ:
Expression of the wild-type insulin-like growth factor II receptor
gene suppresses growth and causes death in colorectal carcinoma
cells. Oncogene. 18:4063–4068. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Devi GR, De Souza AT, Byrd JC, Jirtle RL
and Macdonald RG: Altered ligand binding insulin-like growth
facter2/mannose 6-phosphate receptor bearing missense mutations in
human cancers. Cancer Res. 59:4314–4319. 1999.PubMed/NCBI
|
|
20
|
Haisa M: The type 1 insulin-like growth
factor receptor signalling system and targeted tyrosine kinase
inhibition in cancer. J Int Med Res. 41:253–264. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Pian L, Wen X, Kang L, Li Z, Nie Y, Du Z,
Yu D, Zhou L, Jia L, Chen N, et al: Targeting the IGF 1R pathway in
breast cancer using antisense inc RNA-mediated promoter cis
competition. Mol Ther Nucleic Acids. 12:105–117. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Holliday DL and Speirs V: Choosing the
right cell line for breast cancer research. Breast Cancer Res.
13:2152011. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hussain W, Mahmood T, Hussain J, Ali N,
Shah T, Qayyum S and Khan I: CRISPR/Cas system: A game changing
genome editing technology, to treat human genetic diseases. Gene.
685:70–75. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
van Diemen FR, Kruse EM, Hooykaas MJ,
Bruggeling CE, Schürch AC, van Ham PM, Imhof SM, Nijhuis M, Wiertz
EJ and Lebbink RJ: CRISPR/Cas9-mediated genome editing of
herpesviruses limits productive and latent infections. PLoS Pathog.
12:e10057012016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ophinni Y, Inoue M, Kotaki T and Kameoka
M: CRISPR/Cas9 system targeting regulatory genes of HIV-1 inhibits
viral replication in infected T-cell cultures. Sci Rep. 8:77842018.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Feng YZ, Zhang QY, Fu MT, Zhang ZF, Wei M,
Zhou JY and Shi R: Low expression of PinX1 is associated with
malignant behavior in basal-like breast cancer. Oncol Rep.
38:109–119. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Mei Y, Cai D and Dai X: Modulating cancer
stemness provides luminal a breast cancer cells with HER2
positive-like features. J Cancer. 11:1162–1169. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mali P, Yang L, Esvelt KM, Aach J, Guell
M, DiCarlo JE, Norville JE and Church GM: RNA-guided human genome
engineering via Cas9. Science. 339:823–826. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yoon DS, Kim YH, Jung HS, Paik S and Lee
JW: Importance of Sox2 in maintenance of cell proliferation and
multipotency of mesenchymal stem cells in low-density culture. Cell
Prolif. 44:428–440. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Teh CZ, Wong E and Lee CY: Generation of
monoclonal antibodies to human chorionic gonadotropin by a facile
cloning procedure. J Appl Biochem. 6:48–55. 1984.PubMed/NCBI
|