Introduction
Mitochondria are the main producer of energy
(1) and an essential component of
redox control in cancer cells (2).
Since high levels of reactive oxygen species (ROS) are toxic to
cancer cells, mitochondria are a promising target for novel
anti-cancer therapeutics. Elesclomol is a first-in-class
investigational mitochondria-targeting agent for treating solid
tumors (3). Although elesclomol has
been reported to induce oxidative stress (4,5), several
factors may limit its efficacy for eradicating tumors when used as
a single agent. For instance, cancer cells shift energy metabolism
from oxidative phosphorylation to glycolysis (6), which helps to prevent excessive ROS
accumulation. Cancer cells can also increase tolerance to
mitochondria-targeting agents through antioxidant mechanisms
(7).
Another potentially major protective mechanism
against excessive ROS generation is mitochondrial uncoupling
proteins (UCPs), specialized mitochondrial anion transporters that
mediate leakage of protons at the inner mitochondrial membrane
(8). Among the five types of UCPs in
mammalian cells, UCP2, which has been previously linked to obesity
and diabetes (9), has gained
increasing interest; studies have demonstrated high UCP2 expression
in various cancer cells, including those of the breast, ovary,
bladder, esophagus, liver, colon, lung and pancreas (10,11). By
reducing electron transport chain efficiency, UCP2 increases energy
expenditure (9) and reduces ROS
generation (8). UCP2 is thus
involved in various metabolic disorders (12) and cellular ROS regulation (8). In particular, UCP2-mediated
mitochondrial uncoupling exerts protective effects under various
stressful conditions, such as excessive production of mitochondrial
superoxide ions and aerobic glycolysis stimulation (13). Previous studies have also reported
that UCP2 serves a role in tumor aggressiveness and anticancer
resistance (14,15), which is considered to be associated
with its ability to attenuate mitochondrial ROS production
(13,16). Thus, blocking UCP2 function may be a
viable strategy for eliminating chemotherapy-resistant cancer cells
(17).
A promising drug with highly selective inhibitory
action against UCP2 activity is a natural compound termed genipin,
an aglycone of geniposide present in an extract from Gardenia
jasminoides, which has been demonstrated to block UCP2-mediated
proton leak in several cell types, such as pancreatic islet cells
(9). Previous studies have reported
that genipin stimulates ROS generation (18) and induces cytotoxic effects on
leukemia and liver cancer cells (19,20).
Although combination chemotherapy for cancer has been extensively
studied (21,22), the present study focused on
combination therapy for cancer cells with poor response to the
promising ROS inducer elesclomol and hypothesized the lack of
effects of elesclomol on A549 lung cancer cells to UCP2 activity
for the first time.
We hypothesized that UCP2 may protect cancer cells
from elesclomol cytotoxicity, and that blocking UCP2 function with
genipin may enhance the antitumor efficacy of elesclomol. The aim
of the present study was to investigate the combined therapeutic
effect of genipin and elesclomol, indicating the potential of a
cancer inhibition strategy using co-treatment with the UCP2
inhibitor genipin in A549 lung cancer cells, where there are
currently limited effects on ROS induction due to the high
expression levels of UCP2.
Materials and methods
Cell culture and reagents
A549 human lung cancer cells from the American Type
Culture Collection were maintained in RPMI-1640 medium (Lonza
Group, Ltd.) supplemented with 10% FBS (Serana Europe GmbH) and 1%
penicillin/streptomycin (Gibco; Thermo Fisher Scientific, Inc.) at
37°C with 5% CO2 in a humidified atmosphere. Cells were
sub-cultured twice a week and used at passages <15. The cells
were confirmed to be mycoplasma free and authenticated by the
Samsung Medical Center institutional research support center. All
experiments were performed at cell confluence of 70–80%.
Genipin, N-acetylcysteine (NAC) and
tert-butylhydroperoxide (TBHP) were obtained from Sigma-Aldrich;
Merck KGaA. Treatment concentrations of NAC and TBHP were 10 mM and
200 µM, respectively, and were co-treated with elesclomol and
genipin at 37°C for 24 or 48 h. Elesclomol was obtained from
Biorbyt. Elesclomol and genipin stocks were dissolved in DMSO at 20
and 250 mM, respectively, and further diluted with DMSO.
5-(And-6)-Chloromethyl-2′, 7′-dichlorodihydrofluoresscein diacetate
acetyl ester (CM-H2DCFDA), MitoSOX Red and
Lipofectamine® LTX were obtained from Invitrogen; Thermo
Fisher Scientific, Inc. Non-targeting small interfering RNA
(siRNA), siRNA targeting UCP2 and the anti-β-actin antibody (cat.
no. 18470) were obtained from Santa Cruz Biotechnology, Inc.
Primary anti-UCP2 (cat. no. 89326), anti-cleaved caspase-3 (cat.
no. 9661) and anti-PARP (cat. no. 9542), and secondary anti-rabbit
(cat. no. 7074) and anti-mouse IgG (cat. no. 7076) antibodies were
obtained from Cell Signaling Technology, Inc.
Silencing of UCP2 by siRNA
transfection
After the culture medium was changed to that
containing 5% FBS without antibiotics, 80% confluent A549 cells
were transfected with 40 nM UCP2-specific or non-targeted siRNA
mixed with Lipofectamine® RNAiMAX (Invitrogen; Thermo
Fisher Scientific, Inc.) in opti-MEM (Gibco; Thermo Fisher
Scientific, Inc.). At 24 h, the medium was replaced with fresh
medium containing 10% FBS and antibiotics, and the cells were
incubated for another 24 h prior to subsequent experiments.
Clonogenic assay
A549 cells treated with elesclomol and genipin on
100 mm plates were washed twice with 4°C cold PBS, harvested by
trypsinization and seeded in 6-well plates at 500 cells/well. After
9 days, the cells were fixed with 100% methanol at room temperature
(RT) for 2 min and washed twice with cold PBS. The cells were then
stained with crystal violet solution (Sigma-Aldrich; Merck KGaA) at
RT for 2 min and washed twice with cold PBS. The number of colonies
containing ≥50 cells were finally manually counted in each well
using an inverted phase contrast microscopy (magnification, ×4;
Olympus Corporation).
Sulforhodamine B (SRB) assay
Cell survival was evaluated by SRB assay in
1×104 cells/well seeded overnight in a 96-well plate.
Following treatment for 24 and 48 h, A549 cells were fixed in 10%
(wt/vol) trichloroacetic acid at 4°C for 2 h. After the cells were
stained with SRB dye at RT for 30 min, excess dye was removed by
repeated washing with 1% (v/v) acetic acid. The protein-bound dye
was dissolved in 10 mM Tris base solution, and absorbance at 510 nm
was measured using a VERSA max microplate reader (Molecular
Devices, LLC).
Lactate dehydrogenase (LDH) activity
assay
A549 cells were seeded in a 12-well plate at a
density of 2×105 cells/well and genipin and elelsclomol
treated the following day for 24 h. LDH activity was measured in
500 µl cell culture medium for released LDH and 20 µg cell lysate
for intracellular LDH using a Cobas c501 assay kit (cat. no. c501;
Roche Diagnostics) according to the manufacturer's instructions.
Tumor tissues were homogenized with T-PER™ tissue protein
extraction reagent (Thermo Fisher Scientific, Inc.) on ice and
centrifuged for 10 min at 10,000 × g at 4°C. Subsequently, 20 µg of
supernatants were assayed for LDH activity as above.
Mitochondrial membrane potential (MMP)
measurement
A549 cells were seeded in a black 96-well plate at a
density of 4×104 cells/well and genipin and elesclomol
treated the following day for 24 h. The culture medium was replaced
with 100 µl phenol red-free RPMI-1640 medium containing 2% FBS and
500 nM MitoTracker™ Red FM (Invitrogen; Thermo Fisher Scientific,
Inc.), a fluorescent dye that stains the mitochondria of live cells
as a function of MMP The cells were incubated for 30 min at 37°C in
5% CO2 and washed with 100 µl cold PBS per well. The
fluorescence in each well was measured using a microplate reader
(Mithras LB 940; Titertek-Berthold) using 594 nm excitation and 642
nm emission wavelengths.
Annexin V and propidium iodide (PI)
flow cytometric analysis
A549 cells at density of 5×105 cells/ml
were genipin and elesclomol treated for 24 h, and apoptotic rates
were determined using an Annexin V-FITC Apoptosis kit (BD
Biosciences) according to the manufacturer's instructions. Annexin
V for early apoptotic cells and propidium iodide (PI) for late
apoptotic cells were used. A total of 10,000 cells per sample
underwent FACS analysis on a Calibur flow-cytometer (BD
Biosciences) using CellQuest software version 5.1 (Becton-Dickinson
and Company).
Intracellular ROS measurement
Intracellular ROS levels were quantified using the
cell permeant indicator CM-H2DCFDA. Briefly, A549 cells
were seeded at a density of 4×104 cells/well in a black 96-well
plate. After 24 h, the culture medium was changed to 10%
FBS-containing phenol red-free RPMI-1-1640 medium (Gibco; Thermo
Fisher Scientific, Inc.) and cells were treated with genipin and
elesclomol or transfected with UCP2 siRNA. Following incubation
with 10 µM CM-H2DCFDA at 37°C for 30 min, fluorescence
was measured using a GloMax®-Multi Detection System
microplate reader (Promega Corporation) at 490 nm excitation and
510–570 nm emission wavelengths.
Mitochondrial ROS measurement
Mitochondrial ROS was measured using MitoSOX Red
with dihydroethidium conjugated to a mitochondria-localization tag.
A549 cells were seeded at a density of 4×104 cells/well
in a black 96-well plate, and the culture medium was changed the
next day to 10% FBS-containing phenol red-free RPMI-1640 medium.
Following genipin and elesclomol treatment, the culture medium was
removed, and MitoSOX dye in HBSS buffer (Lonza Group, Ltd.)
containing 2% bovine serum albumin (Bovogen Biologicals Pvt. Ltd.)
was added. Following incubation at 37°C for 10 min, the buffer was
changed to HBSS without dye, and fluorescence was measured on a
Mithras LB 940 microplate reader at 510 nm excitation and 580 nm
emission wavelengths.
Glucose uptake measurement
A549 cells in 24-well plates at a densities of
5×104 cells/well were incubated for 40 min with 100–250
kBq 18F-fluorodeoxyglucose (FDG) (provided by Samsung
Medical Centre) added to the culture medium at 37°C in 5%
CO2. The cells were rapidly washed twice with cold PBS
and lysed with 0.05 N NaOH, and cell-associated radioactivity was
measured on a Wallac high-energy γ-counter (PerkinElmer, Inc.).
Glucose uptake levels were corrected for cell protein content as
assessed by Bradford assay.
Immunoblotting
Immunoblotting was performed in A549 cells and tumor
tissues lysed with cold PRO-PREP™ protein extraction solution
(iNtRON Biotechnology, Inc.) and T-PER™ tissue protein extraction
reagent (Thermo Fisher Scientific, Inc.), respectively, with
protease inhibitor cocktail (Sigma-Aldrich; Merck KGaA). Cell
experiments were performed by obtaining cell lysates following 24-h
incubation with 250 µM genpin and 0.1 µM elesclomol Following
Bradford assays, 30 µg of proteins were separated by 10% sodium
dodecyl sulfate polyacrylamide gel electrophoresis, followed by
transfer to a polyvinylidene difluoride membrane. The membrane was
blocked with 5% skim milk in Tris-buffered saline and 0.1% Tween-20
for 1 h at room temperature and incubated overnight at 4°C with
primary antibodies (1:1,000 dilution), followed by 1 h incubation
at room temperature with secondary antibodies (1:2,000 dilution).
Immunoreactive protein was detected by chemiluminescence (Pierce
ECL plus western blotting substrate; Pierce; Thermo Fisher
Scientific, Inc.), and band intensities were quantified using a
GS-800 densitometer with Quantity One software version 4.6.6
(Bio-Rad Laboratories, Inc.).
Mouse tumor model and drug
treatment
Animal experiments were performed in accordance with
the National Institutes of Health Guide for Care and Use of
Laboratory Animals and approved by the Institutional Animal Care
and Use Committee of Samsung Biomedical Research Institute at
Samsung Medical Center (approval no. 20200413001). The tumor model
was established in 5-week-old male BALB/c-nude mice weighing 20–25
g (Orient Bio, Inc.) by subcutaneous injection of 5×106
A549 cells into the right shoulder. After two weeks, when tumors
reached a diameter of ~0.5 cm, the animals were treated with
intraperitoneal injections of DMSO control (n=6), 60 mg/kg genipin
(n=6), 30 mg/kg elesclomol (n=7) or 60 mg/kg genipin plus 30 mg/kg
elesclomol (n=7) three times per week for a total of four weeks.
The dosages of genipin and elesclomol were based on previous
studies (23,24). The mice were weighed and the tumor
size was measured with a caliper before each injection. The
experiment was repeated twice for a final total of 6–7 animals per
treatment group.
In vivo imaging of tumor FDG
uptake
Positron emission tomography (PET) imaging was
performed on living tumor-bearing mice on the day following the
final drug injection. At 1 h post-injection with 7.4 MBq of FDG via
the tail vein, the animals were anesthetized by isoflurane
inhalation, and PET images were acquired on an Inveon micro-PET
scanner (Siemens Healthineers). Non-attenuation-corrected
tomographic images were analyzed on an Inveon Research Workplace
workstation version 4.2 (Siemens Healthineers). Polygonal regions
of interest were manually drawn with care to include all tumor mass
while excluding adjacent tissue. After PET imaging, the
anesthetized mice were sacrificed by cervical dislocation, and the
tumor tissues were excised.
Statistical analysis
Data are presented as mean ± standard deviation or
the mean ± standard error of the mean. Significant differences
between groups were analyzed by paired Student's t-test for two
groups and one-way ANOVA with Bonferroni post hoc test for multiple
groups. P<0.05 was considered to indicate a statistically
significant difference.
Results
Effects of UCP2 knockdown on A549 cell
FDG uptake, ROS production and survival
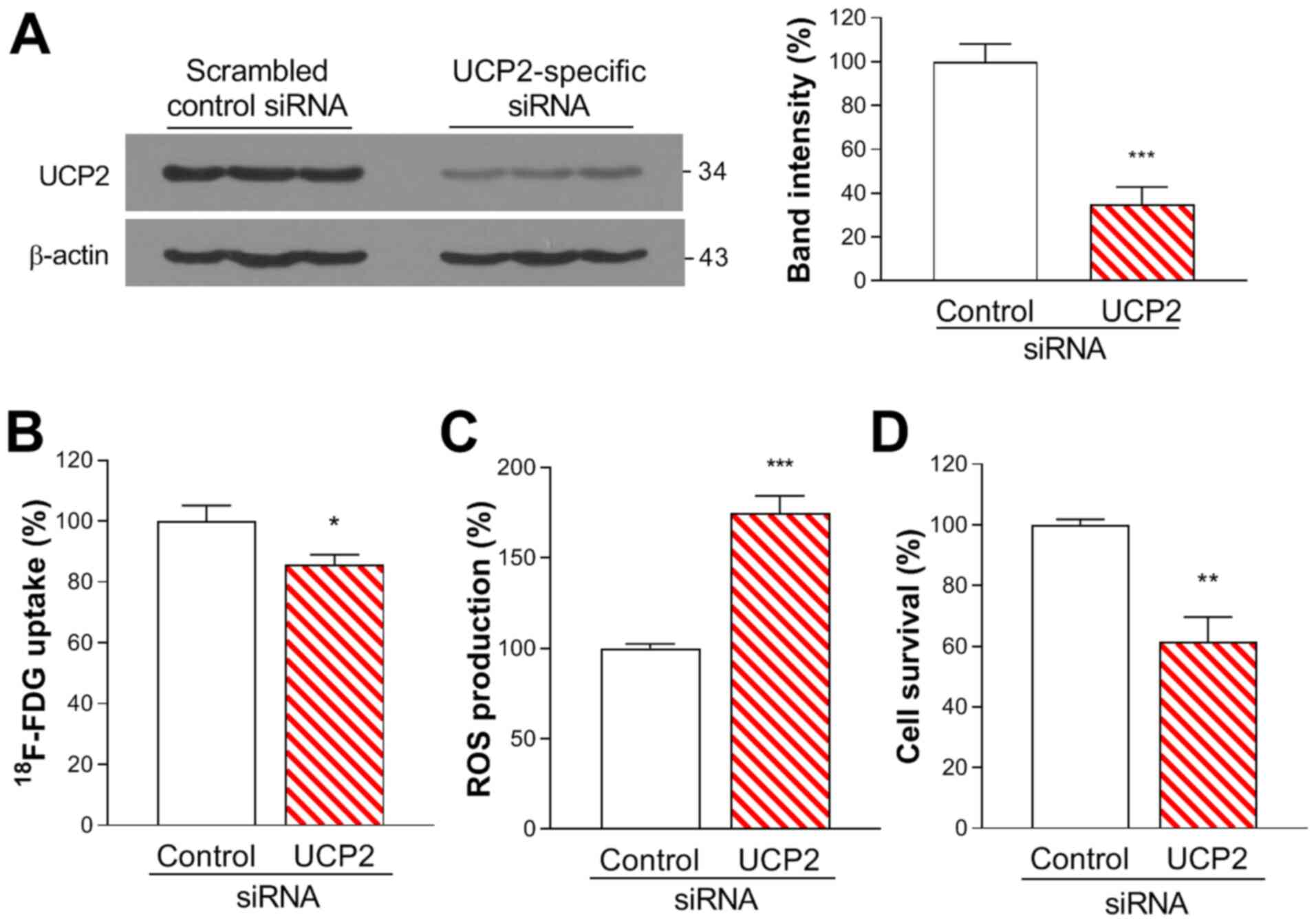
Western blot analysis demonstrated that A549 cells
exhibited high protein expression levels of UCP2. Transfection with
UCP2-specific siRNA effectively knocked down UCP2 expression to
34.8±8.0% of that in the non-targeted siRNA-transfected control
cells (Fig. 1A).
Silencing of UCP2 expression with siRNA slightly
suppressed FDG uptake to 88.5±3.3% of that in control cells
(Fig. 1B. ROS production was
significantly enhanced by UCP2 knockdown to 174.7±9.6% of that in
controls (Fig. 1C). SRB assays
results revealed that UCP2 silencing significantly decreased A549
cell viability to 60.8±7.9% of that in the control group (Fig. 1D).
Effects of genipin and elesclomol on
FDG uptake and ROS production
Treatment of A549 cells for 24 h with 250 µM
genipin, which blocks UCP2 function without reducing its expression
(16), did not affect the protein
expression levels of UCP2 (Fig. 2A).
FDG uptake was unaffected by 24-h treatment with 0.1 µM elesclomol
alone, but was significantly suppressed by 250 µM genipin alone or
by 250 µM genipin plus 0.1 µM elesclomol to 72.0±7.1 and 65.8±4.6%
of that in the control group, respectively (Fig. 2B).
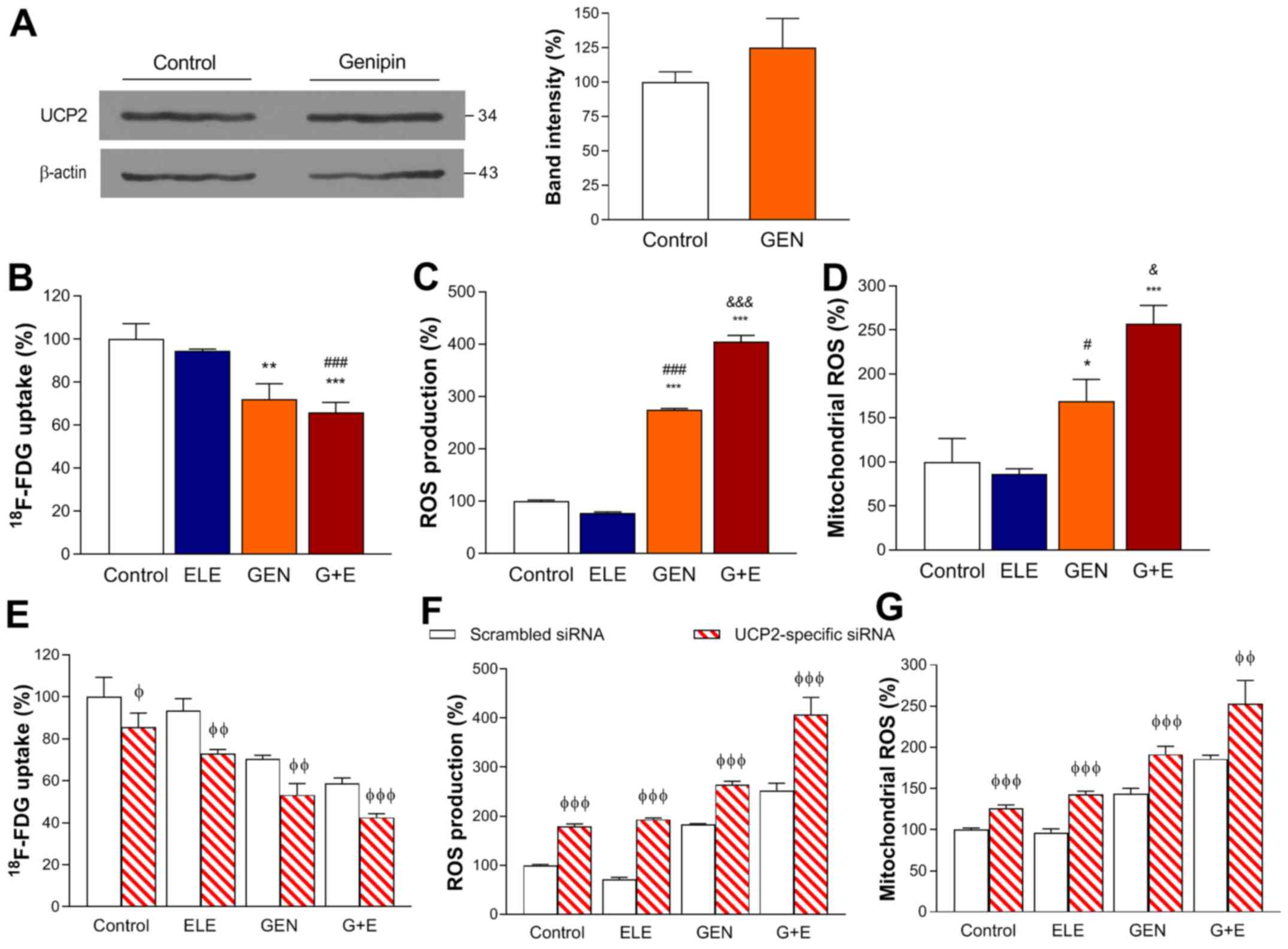 | Figure 2.Effects of GEN and ELE treatment on
A549 lung cancer cells with or without UCP2 knockdown. (A) Western
blots of UCP2 expression in A549 cells following 24-h treatment
with 250 µM GEN or vehicle (control). Quantified band intensities
were normalized to those of β-actin. (B) FDG uptake, (C)
intracellular and (D) mitochondrial ROS production following 24-h
treatment with 0.1 µM ELE, 250 µM GEN, G + E or vehicle (control).
(E) FDG uptake, (F) intracellular and (G) mitochondrial ROS
production following 48-h ELE, GEN and G + E treatment in cells
transfected with scrambled control or UCP2-specific siRNA. Data are
presented as the mean ± SD of values obtained from triplicate
samples of a single representative experiment expressed as
percentage values relative to the control group. *P<0.05,
**P<0.01 and ***P<0.001 vs. control; #P<0.05,
###P<0.001 vs. ELE; &P<0.05,
&&&P<0.001 vs. GEN;
φP<0.05, φφP<0.01 and
φφφP<0.001 vs. scrambled siRNA. GEN, genipin; ELE,
elesclomol; G + E, genipin and elesclomol; UCP2, uncoupling
protein-2; siRNA, small interfering RNA; FDG,
18F-fluorodeoxyglucose; ROS, reactive oxygen
species. |
Intracellular ROS production was also unaffected by
0.1 µM elesclomol alone. However, it was increased to 274.9±1.8% of
that in the control group following 6-h treatment with 250 µM
genipin, and was further increased to 404.3±21.4% of that in the
control group when elesclomol was also present (Fig. 2C). Similarly, mitochondrial ROS
production, which was unaffected by elesclomol alone, increased to
169.3±24.5% of that in the control group in the cells treated with
250 µM genipin and further increased to 256.9±36% of that in the
control group following co-treatment with genipin and elesclomol
(Fig. 2D).
Effects of genipin and elesclomol
following UCP2 knockdown
Following UCP2 knockdown, the effects of genipin and
elesclomol were more pronounced compared with those in cells
transfected with the scrambled control. FDG uptake was decreased to
58.6±2.6% of that in the control group by genipin plus elesclomol
in the scrambled siRNA-transfected group, and it was further
reduced to 42.4±1.8% when UCP2 was knocked down (Fig. 2E). Similarly, cytosolic and
mitochondrial ROS production levels, which were increased to
252.3±14.8 and 186.0±4.5% of those in controls by genipin plus
elesclomol, respectively, were further increased to 406.8±35.0 and
253.2±27.9%, respectively, when UCP2 was knocked down (Fig. 2F and G).
Effects of genipin and elesclomol on
cell clonogenic capacity and survival
The anticancer effects of co-treatment with genipin
and elesclomol were analyzed in vitro. Clonogenic assays
demonstrated that, when used alone, 0.1 µM elesclomol and 250 µM
genipin exerted limited effects on the colony formation capacity of
A549 cells. However, combining the two drugs reduced cell colony
numbers to 50.6±7.4% of those in the control group following 48-h
treatment (Fig. 3A).
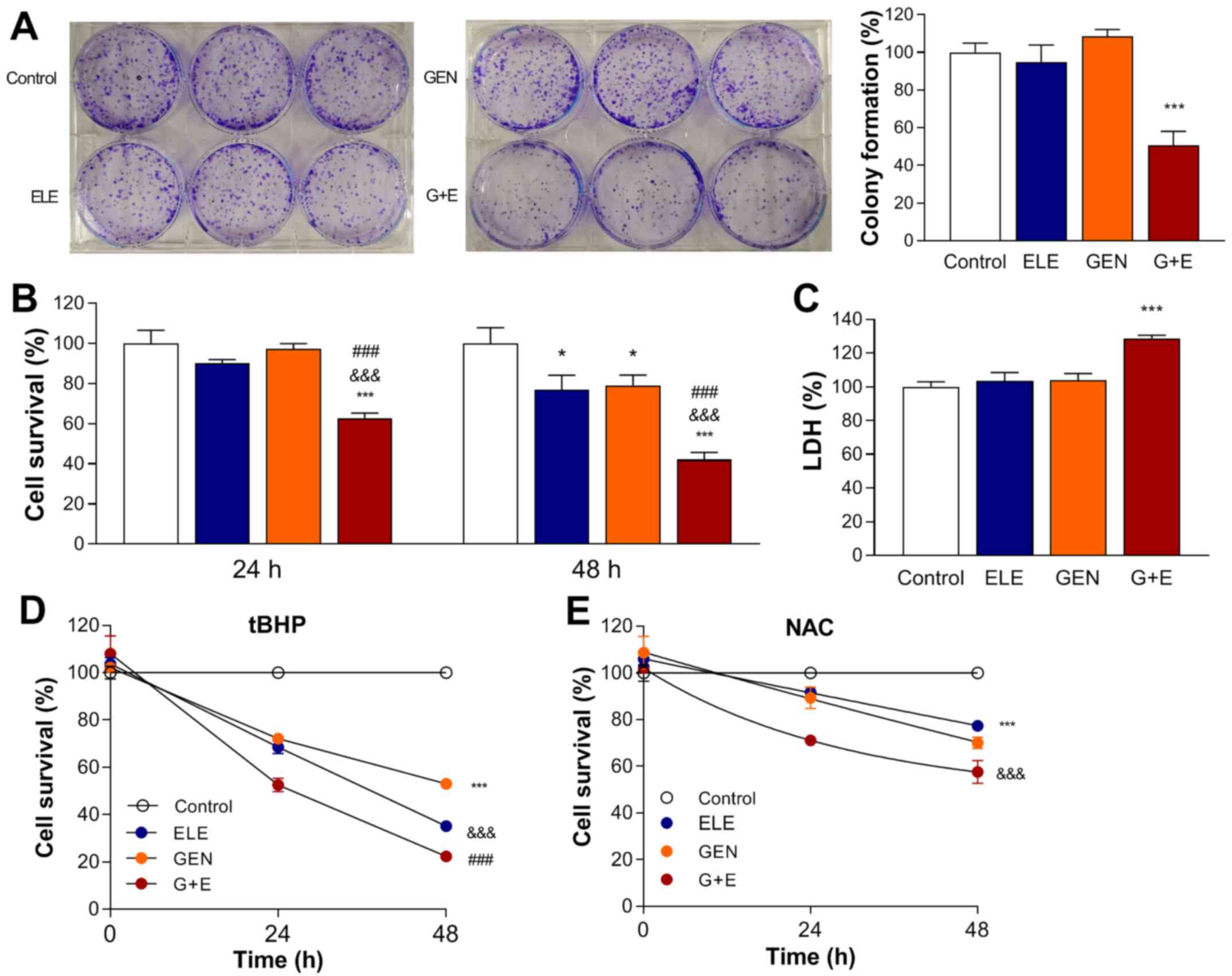 | Figure 3.Effects of GEN and ELE on A549 cancer
cell viability in vitro. (A) Colony forming capacity of A549
cells following 48-h treatment with 0.1 µM ELE, 250 µM GEN, G + E
or vehicle (control). (B) Cell survival following 24- or 48-h ELE,
GEN and G + E treatment and (C) LDH release after 24-h treatment.
(D) Effects of 250 µM GEN and 0.1 µM ELE on A549 cell survival
under co-treatment with 200 µM ROS inducer TBHP or (E) 10 mM ROS
scavenger NAC. Data are presented as the mean ± SD of the
percentage of the control values obtained from triplicate samples
of a single representative experiment. *P<0.05 and ***P<0.001
vs. control; ###P<0.001 vs. ELE;
&&&P<0.001 vs. GEN. GEN, genipin; ELE,
elesclomol; G + E, genipin and elesclomol; LDH, lactate
dehydrogenase; NAC, N-acetylcysteine; TBHP,
tert-butylhydroperoxide; ROS, reactive oxygen species. |
Similarly, the results of the SRB assay demonstrated
that 48-h treatment with 0.1 µM elesclomol or 250 µM genipin alone
suppressed cell survival to 78.9±5.2 and 76.9±7.1% of that in the
control group, respectively. However, combining the two drugs
significantly reduced cell survival to 42.0±3.4% of controls
(Fig. 3B, middle). Furthermore, LDH
release, assessed as a measure of cytotoxicity, was unaffected by
24-h treatment with 0.1 µM elesclomol or 250 µM genipin alone, but
was increased to 128.8±1.6% of that in the control group by
co-treatment (Fig. 3C), accompanied
by decreased intracellular LDH to 63.8±1.3% of that in the control
group (data not shown).
The role of ROS on the cytotoxic effects of the two
drugs was assessed in the presence of the exogenous ROS inducer
TBHP and ROS scavenger NAC. The results demonstrated that the
cytotoxic effects of genipin, elesclomol and their combination were
enhanced by TBHP and reduced by NAC; the cell survival in the
elesclomol and genipin co-treated cells at 48 h was decreased to
22.3±1.9% of that in the control group by TBHP (Fig. 3D) and was partially recovered to
57.5±4.8% of that in the control group by NAC (Fig. 3E).
Effects of genipin and elesclomol on
A549 cell apoptosis
The present study next evaluated whether the
antitumor effect of combining elesclomol with genipin involved the
induction of apoptosis. The results of the MitoTracker Red assay
demonstrated that co-treatment with elesclomol and genipin for 24 h
significantly reduced A549 cell MMP to 70.4±2.5% of that in the
control group (Fig. 4A). This was
accompanied by a 6-fold increase in the percentage of Annexin
V-positive apoptotic cells (Fig.
4B). In addition, the co-treatment induced a marked increase in
the levels of cleaved PARP to 837.6±221.9% and of cleaved caspase-3
to 495.9±68.6% of those in untreated controls (Fig. 4C and D).
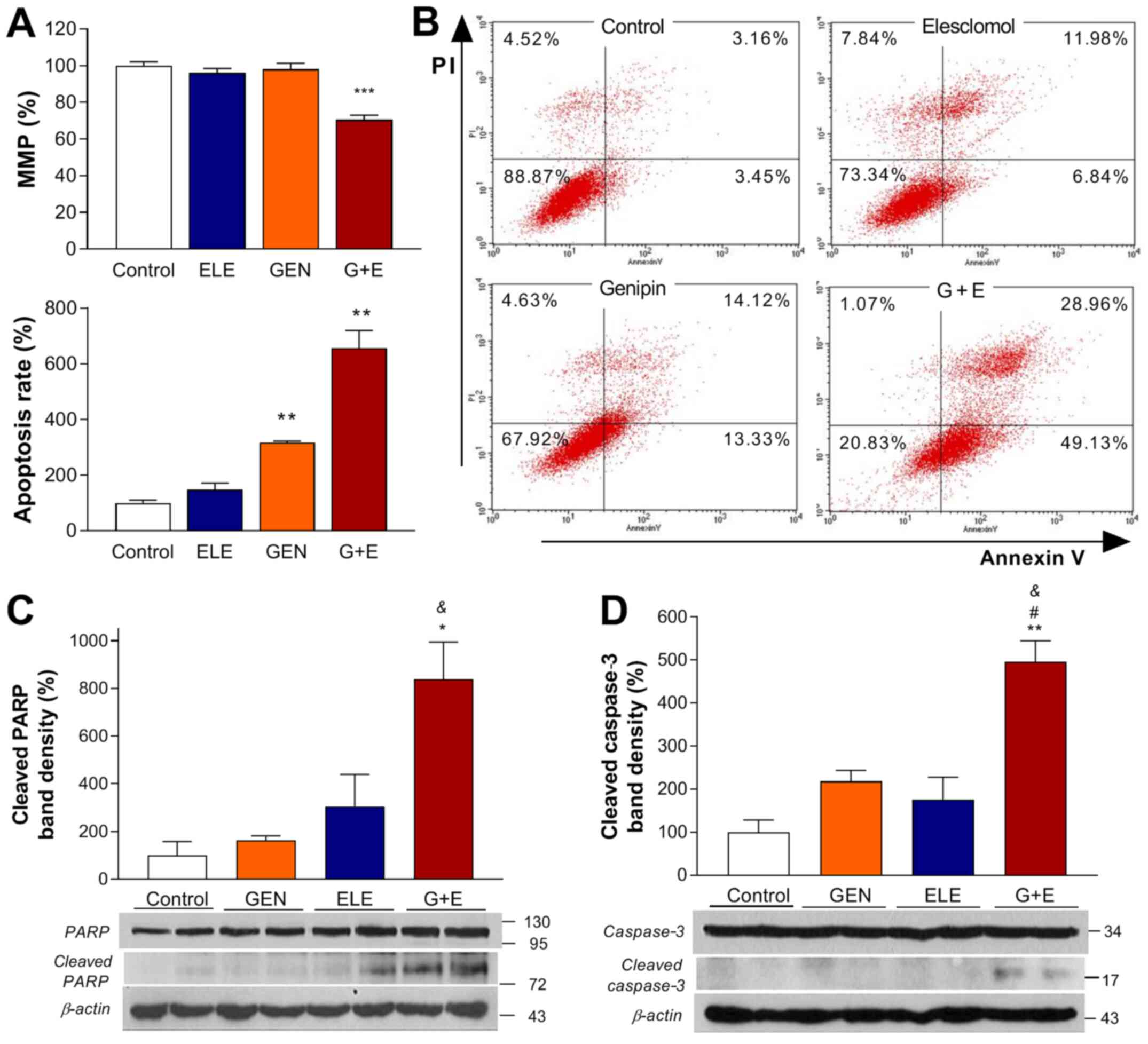 | Figure 4.Effects of GEN and ELE on apoptosis
in A549 cells. (A) MMP was measured by MitoTracker Red FM, and (B)
apoptosis was analyzed using FITC-Annexin V and PI staining
following 24-h treatment with 0.1 µM ELE, 250 µM GEN, G + E or
vehicle (control). (C) Western blots of PARP, cleaved PARP, (D)
caspase-3 and cleaved caspase-3 following 24-h treatment with ELE,
GEN or G + E. Quantified band intensities were normalized to those
of β-actin. Data are presented as the mean ± SD of the percentage
of the control values obtained from (A and B) triplicate or (C and
D) duplicate samples of a single representative experiment.
*P<0.05, **P<0.01, ***P<0.001 vs. control;
#P<0.05 vs. ELE; &P<0.05 vs. GEN.
GEN, genipin; ELE, elesclomol; G + E, genipin and elesclomol; MMP,
mitochondrial membrane potential; PARP, poly(ADP-ribose)
polymerase. |
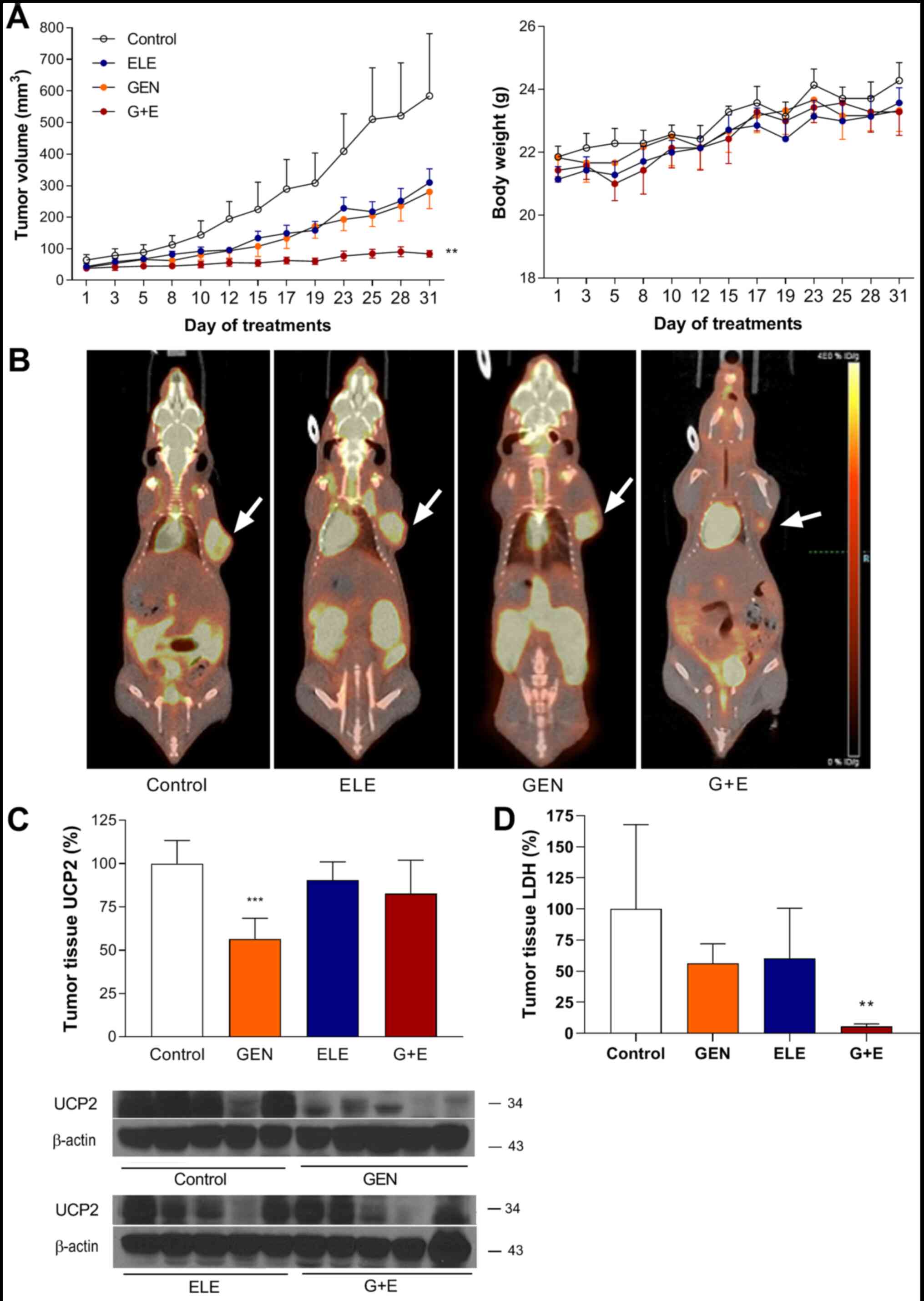
In vivo antitumor effects of genipin
and elesclomol on A549 ×enografts in mice
Treatment of A549 tumor xenografts in mice with
genipin or elesclomol led to similar magnitudes of modest
retardation in tumor growth (280.1±130.8 and 310.1±114.6
mm3 on day 31, respectively) compared with that in the
vehicle-treated control mice (584.2±482.5 mm3 on day 31;
Fig. 5A). By contrast, combination
therapy with elesclomol plus genipin resulted in near complete
suppression of tumor growth (Fig.
5A); the tumor volume in the co-treatment group (84.0±27.8
mm3 on day 31) was significantly lower compared with
that in the control group No significant differences were observed
in animal weight during treatment (Fig.
5A).
PET/CT performed after the final treatment revealed
significantly smaller tumors with lower FDG uptake in animals
co-treated with elesclomol and genipin compared with those in
animals treated with control or with elesclomol or genipin alone
(Fig. 5B).
Western blots analysis of UCP2 protein expression in
the xenograft tumor tissues demonstrated that UCP2 expression
levels in the tumors isolated from genipin-treated mice was reduced
to 56.4±12.0% of those in the tissues from the untreated control
group (Fig. 5C). In addition, the
LDH levels were lower in the tumor tissue of mice treated with
genipin plus elesclomol compared with the control groups (Fig. 5D).
Discussion
Elesclomol enters cells in a complex with copper,
which it transports into the mitochondria to stimulate ROS
generation; dissociated elesclomol then effluxes out of the cells
to repeat mitochondrial copper shuttling and accumulation (25). However, despite its high potential
for treating various malignancies including lung cancer (3–5),
elesclomol has been limited in increasing ROS and eliminating small
and non-small lung cancer cells (26). Although a number of studies have
examined the beneficial effects of combining chemotherapeutic drugs
for cancer therapy, the present study tested a unique hypothesis
that the natural compound genipin may enhance the efficacy of
elesclomol to induce ROS generation and promote cell death through
the suppression of UCP2 function.
The present study first confirmed that A549 cancer
cells exhibited a high level of UCP2 protein expression. In these
cells, UCP2 knockdown with a specific siRNA mildly decreased FDG
uptake, significantly elevated ROS production and suppressed cell
survival compared with those in the control group. When the
biological effects of elesclomol and genipin were compared, it was
observed that elesclomol monotherapy was unable to significantly
affect FDG uptake or ROS production. By contrast, genipin mildly
decreased FDG uptake and significantly elevated cellular and
mitochondrial ROS production in a manner similar to that obtained
by UCP2 knockdown. Furthermore, the effects were substantially
enhanced in cells treated with a combination of elesclomol and
genipin. In these experiments, elesclomol and genipin were used at
the concentrations of 0.1 and 250 µM, respectively, which were
slightly lower compared with the 0.5 µM (4) and 400 µM (27) used in previous studies, respectively,
to determine their anticancer effects.
Genipin is an inhibitor of UCP2 function rather than
expression (16,20,28–30),
which is consistent with the result of the present study that the
protein levels of UCP2 were not reduced by the genipin treatment.
The major role of UCP2 is to inhibit ROS generation (15–19).
Inhibition of UCP2 function by genipin may therefore be
demonstrated by an increase of ROS production or a reduction of
proton leak on oxygen consumption rate analysis. This has been
confirmed in various cancer cell types, such as pancreatic cancer
cells, breast cancer cells by our group and others (11,19,20,29–31).
Thus, the results of the present study that the significantly
increased ROS production induced by UCP2 knockdown or genipin may
represent the blocking of UCP2 activity-mediated protection against
oxidative stress. In addition, the mild reduction of glucose uptake
by UCP2 knockdown or by genipin may be explained by the inhibition
of the UCP2-mediated glucose expenditure that shifts glucose
metabolism toward glycolytic flux (31,32).
These effects were more pronounced when genipin treatment was
performed in UCP2-knockdown cells, which may indicate a more
complete blockage of mitochondrial proton leakage.
When in vitro cytotoxic effects were assessed
in the present study, elesclomol alone exhibited limited efficacy
in suppressing the colony forming capacity and survival of A549
cells. High UCP2 expression has been previously indicated to
attenuate the ROS-stimulatory and cytotoxic effects of paclitaxel
(33). Therefore, the limited
cytotoxicity of elesclomol observed in the present study may be
attributed to the protective action of UCP2 against mitochondrial
ROS buildup. This was supported by substantial enhancement by
genipin of the ability of elesclomol to suppress A549 cell colony
formation and survival, as well as by enhanced cell death in the
presence of an ROS donor and suppressed cell death in the presence
of a ROS scavenger.
The results of the present study also demonstrated
that co-treatment with elesclomol and genipin significantly reduced
A549 cell MMP and induced apoptosis, as assessed by Annexin V FACS
analysis. This was accompanied by markedly increased levels of the
cleaved forms of caspase-3 and PARP proteins. These results were
consistent with the notion that UCP2 protects cancer cells from ROS
stress-induced apoptosis (34–36),
which occurs through the intrinsic apoptotic pathway (37). Taken together, these results
indicated that combining the UCP2 inhibitor genipin enhanced the
capacity of elesclomol to induce cancer cell death by activating
the intrinsic apoptotic pathway through ROS stimulation and MMP
suppression.
Notably, in the present study, the in vivo
antitumor effect was also augmented when genipin and elesclomol
were used in combination. For these experiments, A549 tumor-bearing
mice were treated with 60 mg/kg genipin, which was within the range
of 10–100 mg/kg previously administered to mice in combination with
other drugs (23,38). Elesclomol was administered at a dose
of 30 mg/kg, which was in the lower range of the doses previously
used for treating tumor-bearing mice (24). These doses were selected to
demonstrate that the natural compound genipin may be safely added
to enhance the anticancer effects of low-dose elesclomol. The
results revealed that elesclomol and genipin therapy alone
exhibited modest antitumor efficacies that led to similar
retardations of tumor growth. By contrast, combining genipin with
elesclomol nearly completely suppressed tumor growth.
Further analysis of the xenograft tumor tissues
revealed that the protein levels of UCP2 in tumors isolated from
the genipin-treated mice were reduced compared with the control
group. This result may indicate the preferential killing of tumor
cells with greater UCP2 expression by genipin, which led to the
remaining tumor tissue containing cells with lower UCP2 levels.
Tumor tissue LDH level was reduced in mice treated with genipin
plus elesclomol compared with the control group. This finding may
indicate that fewer dying cancer cells exist in the reduced tumor
tissues after long-term treatment with genipin and elesclomol.
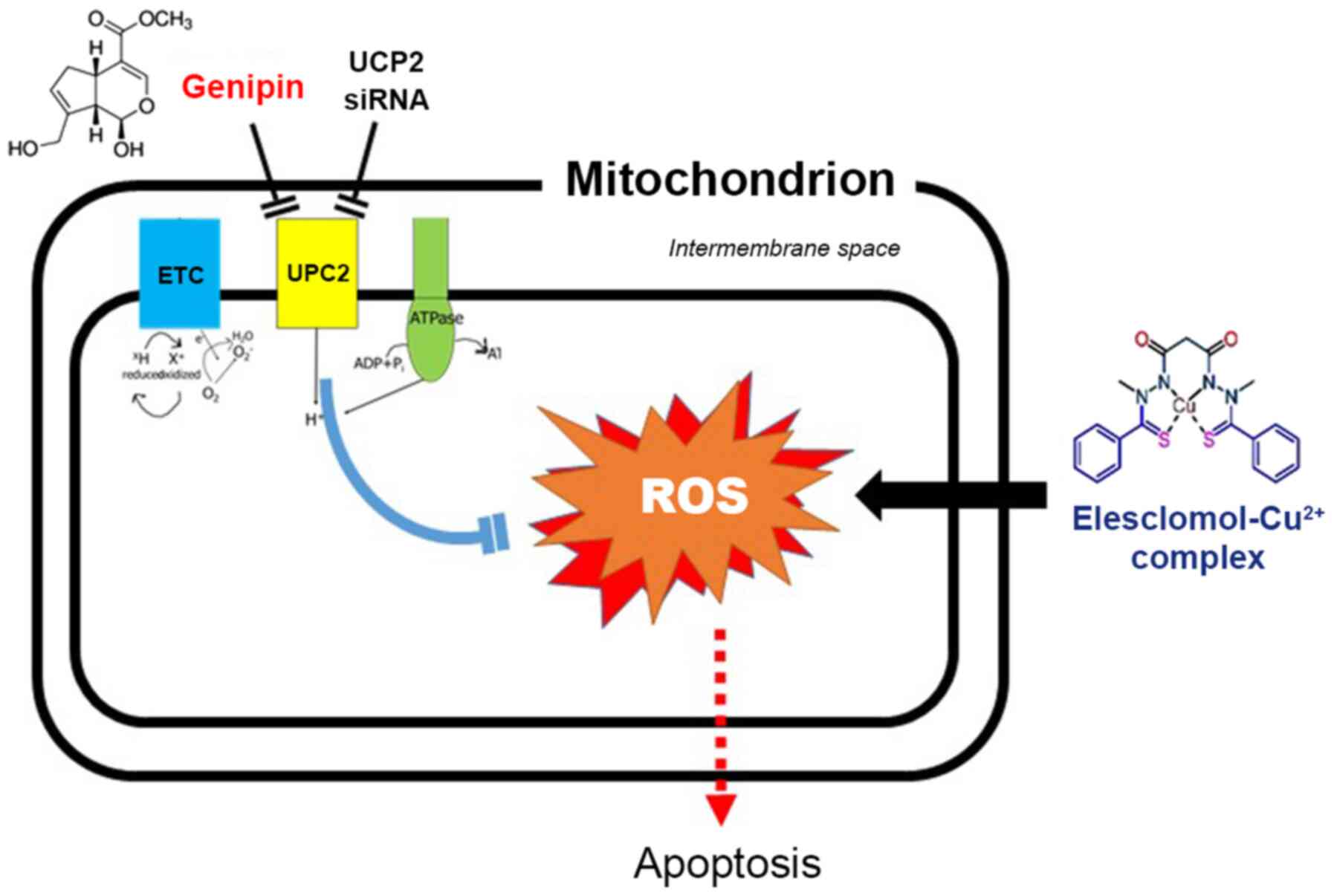
Fig. 6 illustrates
our proposed model for the synergistic anticancer effects of
elesclomol and genipin. In this model, elesclomol enters cancer
cells to induce an increase of mitochondrial oxidative stress, but
this effect can be alleviated by UCP2-mediated oxygen consumption
uncoupling. UCP2 inhibition with genipin or UCP2 siRNA blocks this
protective response and augments the stimulation of ROS production
by elesclomol, which in turn triggers apoptosis of the cancer
cells.
A limitation of the present study was that the
results were restricted to A549 lung cancer cells, which exhibit an
efficient UCP2 function, and therefore may not apply to other
cancer cell types. It is possible that cancer cells with low UCP2
expression may respond to elesclomol monotherapy without the need
for UCP2 inhibition. By contrast, cancer cells with greater UCP2
activity may benefit even more from the combination of UCP2
inhibitors and elesclomol. In addition, the results of the UCP2
western blots in tumor tissue were limited as only some of the
tumor samples were included for analysis. In the animal experiments
conducted in the present study, some tumor samples included in the
tumor growth data were not included in the UCP2 western data. These
issues require validation by further studies.
In conclusion, high UCP2 expression may limit the
antitumor effect of elesclomol by attenuating ROS response. This
can be overcome by blocking UCP2 function using the natural product
genipin, which suppresses glucose uptake, augments mitochondrial
ROS production and enhances cytotoxic effects in cancer cells.
Therefore, a combination of elesclomol with genipin may be an
effective strategy for the treatment of tumors with high UCP
expression.
Acknowledgements
Not applicable.
Funding
This work was supported by the Basic Science
Research Program through the National Research Foundation of Korea
funded by the Ministry of Science, ICT and Future Planning (grant
nos. NRF-2015R1C1A1A01053454 and NRF-2019R1H1A1035585).
Availability of data and materials
The datasets used and analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
KHL, JHL and YSC designed the experiments, analyzed
and interpreted the data, and wrote the manuscript. JHL, YSC, KHJ
and JWP performed the experiments. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
This study was reviewed and approved by the
Institutional Animal Care and Use Committee of Samsung Biomedical
Research Institute (SBRI) at Samsung Medical Center (Seoul, Korea).
The SBRI is accredited by the Association for Assessment and
Accreditation of Laboratory Animal Care International and abides
the Institute of Laboratory Animal Resources guide.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
UCP
|
uncoupling protein
|
|
ROS
|
reactive oxygen species
|
|
NAC
|
N-acetylcysteine
|
|
TBHP
|
tert-butylhydroperoxide
|
|
FDG
|
18F-fluorodeoxyglucose
|
|
PET
|
positron emission tomography.
|
References
|
1
|
Chandel NS: Mitochondria as signaling
organelles. BMC Biol. 12:342014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Vyas S, Zaganjor E and Haigis MC:
Mitochondria and cancer. Cell. 166:555–566. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Berkenblit A, Eder JP Jr, Ryan DP, Seiden
MV, Tatsuta N, Sherman ML, Dahl TA, Dezube BJ and Supko JG: Phase I
clinical trial of STA-4783 in combination with paclitaxel in
patients with refractory solid tumors. Clin Cancer Res. 13:584–590.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kirshner JR, He S, Balasubramanyam V,
Kepros J, Yang CY, Zhang M, Du Z, Barsoum J and Bertin J:
Elesclomol induces cancer cell apoptosis through oxidative stress.
Mol Cancer Ther. 7:2319–2327. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Blackman RK, Cheung-Ong K, Gebbia M, Proia
DA, He S, Kepros J, Jonneaux A, Marchetti P, Kluza J, Rao PE, et
al: Mitochondrial electron transport is the cellular target of the
oncology drug elesclomol. PLoS One. 7:e297982012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Vander Heiden MG, Cantley LC and Thompson
CB: Understanding the Warburg effect: The metabolic requirements of
cell proliferation. Science. 324:1029–1033. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wangpaichitr M, Sullivan EJ,
Theodoropoulos G, Wu C, You M, Feun LG, Lampidis TJ, Kuo MT and
Savaraj N: The relationship of thioredoxin-1 and cisplatin
resistance: Its impact on ROS and oxidative metabolism in lung
cancer cells. Mol Cancer Ther. 11:604–615. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Arsenijevic D, Onuma H, Pecqueur C,
Raimbault S, Manning BS, Miroux B, Couplan E, Alves-Guerra MC,
Goubern M, Surwit R, et al: Disruption of the uncoupling protein-2
gene in mice reveals a role in immunity and reactive oxygen species
production. Nat Genet. 26:435–439. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang CY, Parton LE, Ye CP, Krauss S, Shen
R, Lin CT, Porco JA Jr and Lowell BB: Genipin inhibits
UCP2-mediated proton leak and acutely reverses obesity- and high
glucose-induced beta cell dysfunction in isolated pancreatic
islets. Cell Metab. 3:417–427. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ayyasamy V, Owens KM, Desouki MM, Liang P,
Bakin A, Thangaraj K, Buchsbaum DJ, LoBuglio AF and Singh KK:
Cellular model of Warburg effect identifies tumor promoting
function of UCP2 in breast cancer and its suppression by genipin.
PLoS One. 6:e247922011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kuai XY, Ji ZY and Zhang HJ: Mitochondrial
uncoupling protein 2 expression in colon cancer and its clinical
significance. World J Gastroenterol. 16:5773–5778. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Diano S and Horvath TL: Mitochondrial
uncoupling protein 2 (UCP2) in glucose and lipid metabolism. Trends
Mol Med. 18:52–58. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Donadelli M, Dando I, Dalla Pozza E and
Palmieri M: Mitochondrial uncoupling protein 2 and pancreatic
cancer: A new potential target therapy. World J Gastroenterol.
21:3232–3238. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Michael A: Pitt: Overexpression of
uncoupling protein-2 in cancer: metabolic and heat changes,
inhibition and effects on drug resistance. Inflammopharmacol.
23:365–369. 2015. View Article : Google Scholar
|
|
15
|
Derdak Z, Mark NM, Beldi G, Robson SC,
Wands JR and Baffy G: The mitochondrial uncoupling protein-2
promotes chemoresistance in cancer cells. Cancer Res. 68:2813–2819.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Derdák Z, Fülöp P, Sabo E, Tavares R,
Berthiaume EP, Resnick MB, Paragh G, Wands JR and Baffy G: Enhanced
colon tumor induction in uncoupling protein-2 deficient mice is
associated with NF-kappaB activation and oxidative stress.
Carcinogenesis. 27:956–961. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Santandreu FM, Roca P and Oliver J:
Uncoupling protein-2 knockdown mediates the cytotoxic effects of
cisplatin. Free Radic Biol Med. 49:658–666. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kim BC, Kim HG, Lee SA, Lim S, Park EH,
Kim S-J and Lim CJ: Genipin-induced apoptosis in hepatoma cells is
mediated by reactive oxygen species/c-Jun NH2-terminal
kinase-dependent activation of mitochondrial pathway. Biochem
Pharmacol. 70:1398–1407. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mailloux RJ, Adjeitey CN and Harper ME:
Genipin-induced inhibition of uncoupling protein-2 sensitizes
drug-resistant cancer cells to cytotoxic agents. PLoS One.
5:e132892010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Khanal T, Kim HG, Choi JH, Do MT, Kong MJ,
Kang MJ, Noh K, Yeo HK, Ahn YT, Kang W, et al: Biotransformation of
geniposide by human intestinal microflora on cytotoxicity against
HepG2 cells. Toxicol Lett. 209:246–254. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kalemkerian GP: Combination chemotherapy
for relapsed small-cell lung cancer. Lancet Oncol. 17:1033–1035.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li Y, Atkinson K and Zhang T: Combination
of chemotherapy and cancer stem cell targeting agents: Preclinical
and clinical studies. Cancer Lett. 396:103–109. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang N, Zhu M, Tsao SW, Man K, Zhang Z and
Feng Y: Up-regulation of TIMP-1 by genipin inhibits MMP-2
activities and suppresses the metastatic potential of human
hepatocellular carcinoma. PLoS One. 7:e463182012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gehrmann M: Drug evaluation: STA-4783 -
enhancing taxane efficacy by induction of Hsp70. Curr Opin Investig
Drugs. 7:574–580. 2006.PubMed/NCBI
|
|
25
|
Nagai M, Vo NH, Shin Ogawa L, Chimmanamada
D, Inoue T, Chu J, Beaudette-Zlatanova BC, Lu R, Blackman RK,
Barsoum J, et al: The oncology drug elesclomol selectively
transports copper to the mitochondria to induce oxidative stress in
cancer cells. Free Radic Biol Med. 52:2142–2150. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wangpaichitr M, Wu C, You M, Maher JC,
Dinh V, Feun LG and Savaraj N:
N,N-Dimethyl-N′,N-bis(phenylcarbonothioyl) propanedihydrazide
(elesclomol) selectively kills cisplatin resistant lung cancer
cells through reactive oxygen species (ROS). Cancers (Basel).
1:23–38. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Feng Q, Cao HL, Xu W, Li XR, Ren YQ and Du
LF: Apoptosis induced by genipin in human leukemia K562 cells:
Involvement of c-Jun N-terminal kinase in G2/M arrest.
Acta Pharmacol Sin. 32:519–527. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Valle A, Oliver J and Roca P: Role of
uncoupling proteins in cancer. Cancers (Basel). 2:567–591. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhou H, Zhao J and Zhang X: Inhibition of
uncoupling protein 2 by genipin reduces insulin-stimulated glucose
uptake in 3T3-L1 adipocytes. Arch Biochem Biophys. 486:88–93. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Dando I, Fiorini C, Pozza ED, Padroni C,
Costanzo C, Palmieri M and Donadelli M: UCP2 inhibition triggers
ROS-dependent nuclear translocation of GAPDH and autophagic cell
death in pancreatic adenocarcinoma cells. Biochim Biophys Acta.
1833:672–679. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cho YS, Lee JH, Jung K-H, Park JW, Moon
SH, Choe YS and Lee KH: Molecular mechanism of (18)F-FDG uptake
reduction induced by genipin in T47D cancer cell and role of
uncoupling protein-2 in cancer cell glucose metabolism. Nucl Med
Biol. 43:587–592. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Brandi J, Cecconi D, Cordani M,
Torrens-Mas M, Pacchiana R, Dalla Pozza E, Butera G, Manfredi M,
Marengo E, Oliver J, et al: The antioxidant uncoupling protein 2
stimulates hnRNPA2/B1, GLUT1 and PKM2 expression and sensitizes
pancreas cancer cells to glycolysis inhibition. Free Radic Biol
Med. 101:305–316. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Su WP, Lo YC, Yan JJ, Liao IC, Tsai PJ,
Wang HC, Yeh HH, Lin CC, Chen HH, Lai WW, et al: Mitochondrial
uncoupling protein 2 regulates the effects of paclitaxel on Stat3
activation and cellular survival in lung cancer cells.
Carcinogenesis. 33:2065–2075. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Horimoto M, Resnick MB, Konkin TA,
Routhier J, Wands JR and Baffy G: Expression of uncoupling
protein-2 in human colon cancer. Clin Cancer Res. 10:6203–6207.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Harper ME, Antoniou A, Villalobos-Menuey
E, Russo A, Trauger R, Vendemelio M, George A, Bartholomew R, Carlo
D, Shaikh A, et al: Characterization of a novel metabolic strategy
used by drug-resistant tumor cells. FASEB J. 16:1550–1557. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Collins P, Jones C, Choudhury S, Damelin L
and Hodgson H: Increased expression of uncoupling protein 2 in
HepG2 cells attenuates oxidative damage and apoptosis. Liver Int.
25:880–887. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wasim L and Chopra M: Synergistic
anticancer effect of panobinostat and topoisomerase inhibitors
through ROS generation and intrinsic apoptotic pathway induction in
cervical cancer cells. Cell Oncol (Dordr). 41:201–212. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Li CC, Hsiang CY, Lo HY, Pai FT, Wu SL and
Ho TY: Genipin inhibits lipopolysaccharide-induced acute systemic
inflammation in mice as evidenced by nuclear factor-κB
bioluminescent imaging-guided transcriptomic analysis. Food Chem
Toxicol. 50:2978–2986. 2012. View Article : Google Scholar : PubMed/NCBI
|