Introduction
Recent advances in the areas of medical genetics,
molecular diagnostics and molecular pathology have significantly
helped provide a mechanistic understanding of various diseases
(1). Personalized medicine is a new
and innovative field of healthcare, especially in cancer (2). This field of medicine focuses on the
detection of genomic signatures in an individual, and helps
accurately forecast a person's susceptibility to a disease and
their prognosis, as well as develop a treatment strategy, that
became known as targeted therapy (3).
Gliomas are very aggressive forms of brain cancer
and the most common form of primary brain tumors; despite many
advances in the molecular biology and genetics of gliomas, these
tumors remain incurable (4,5). World Health Organization (WHO) grade-IV
gliomas, also known as glioblastomas multiforme (GBMs), account for
>50% of gliomas (6). Among all
central nervous system (CNS) malignancies, GBM has a very poor
prognosis (7). According to a
published report by the Central Brain Tumor Registry of the United
States in a cohort analysis of glioblastoma tumors from the
National Program of Cancer Registries between 2001 and 2015, the
1-year survival rates reported were 62.5, 71.8, 58.6, 47.4 and
31.2%, and the 5-year survival rates were 20.8, 21.9, 9.3, 5.9 and
3.9% in age groups ranging 0–19, 20–44, 45–54, 55–64 and 65–74
years, respectively (8).
The isocitrate dehydrogenase (IDH) mutation is an
early event in brain tumorigenesis; >70% of grade-II and -III
gliomas have an IDH gene mutation (9,10). IDH
mutations have been found to be strongly associated with 1p/19q
codeletion and O(6)-methyl guanine methyl transferase (MGMT)
promoter methylation, but are mutually exclusive with epidermal
growth factor receptor (EGFR) amplification and loss of chromosome
10 (11,12). Other common mutations present in GBM
include methylation of promoters in the p14ARF, CDKN2A and RB1
genes, and overexpression and mutations in the platelet-derived
growth factor receptor, amplification of MDM2, CDKN2A gene
mutation, overexpression of or mutations in the oncogene EGFR, as
well as mutations and deletions in tumor suppressor genes
phosphatase and tensin homologue (PTEN) and tumor protein p53
(13–15). Among all these genes altered in GBM,
the EGFR gene was recognized as a convincing target for treatment
development, since it is overexpressed, amplified and/or mutated in
≥40-50% of GBMs (16,17). In addition, out of the EGFR
rearrangements that result in truncated isoforms, EGFR
transcriptional variant III (EGFRvIII) is the most common
mutational variant, expressed in 25–64% of GBMs (18). This mutant product contains an
in-frame deletion of 801 base pairs of the coding sequence due to
the deletion of exons 2–7 (19).
This rearrangement gives rise to a ligand-independent kinase
activation that persists downstream of the Ras-MAP kinase and PI3K
pathways, promoting cell proliferation (20).
IDH1 and IDH2 are homologous metabolic isozymes, and
the mutation of these genes generates a neomorphic enzyme, which
can lead to the abnormal accumulation of 2-hydroxyglutarate (2-HG)
and promote tumorigenesis. The IDH1 protein is localized in the
cytoplasm and the peroxisomes (21).
In the cytoplasm, the role of the IDH1 protein is to provide NADPH,
when the pentose phosphate pathway is impaired, whereas IDH2 is
localized in the mitochondria and catalyzes the same reaction as
IDH1. The mutation of the IDH1 and IDH2 genes play an important
role in gliomas by converting α-ketoglutarate (α-KG), which is
produced in tricarboxylic acid (TCA) cycle, to an oncometabolite,
2-HG (22). Glioma tissue with an
IDH1 mutation produces less α-KG and is known to contain high
levels of 2-HG (23). This
metabolite stimulates cellular proliferation through the
degradation of hypoxia-inducible factor (HIF)-α, and inhibits
α-KG-dependent dioxygenases (24).
The conversion of α-KG to 2-HG decreases intracellular NADPH
levels, which contributes to oncogenesis by creating a prooxidant
state that benefits the development of glioma.
There is an abundance of clinical data associated
with mutation detection, genetic profiling and its use in targeted
therapy, prognosis prediction and survival of various types of
cancer, including gliomas in the literature; however, such data
from the Kingdom of Saudi Arabia is sporadic. Commercial molecular
diagnostics laboratories are not ready to provide several cancer
genetic tests in Saudi Arabia. Several drugs aimed at inhibiting
EGFR signaling have already yielded good results in lung and
colorectal cancer; however, for gliomas, clinical trials are still
on-going, and considerable efforts are currently being made to
develop immunotherapy targeting EGFRvIII for glioma (25,26).
Reverse transcription-quantitative (RT-q) PCR for EGFRvIII became a
basic requirement for monitoring the efficacy of this treatment,
and for selecting the patients for the clinical trials. The
molecular detection of IDH1 and IDH2 mutations has become standard
practice in several institutions, and has been proven useful for
the clinical management of gliomas. The aim of the present study
was to determine the prevalence of IDH1 and IDH2 mutations and
EGFRvIII transcript expression status in Saudi Arabian patients
with glioma. Sequencing for IDH1 and IDH2 mutations in brain tumor
samples was performed using the next-generation DNA sequencing
(NGS) and capillary methods. In addition, the EGFRvIII expression
was analyzed in brain tumor samples, by an in-house RT-qPCR assay
developed for the first time in the Kingdom of Saudi Arabia. This
type of investigations will help transfer the clinical assays to
the diagnostics laboratories in the Kingdom, so that this approach
can be utilized to become independent in molecular diagnostics and
established oncology testing for clinical use.
Materials and methods
Chemicals, reagents and equipment
All NGS reagents for Ion Proton sequencing were
purchased from Thermo Fisher Scientific, Inc. Sequencing reagents
POP-7 (cat. no. 4393708) and BigDye v3.1 (cat. no. 4336923), TaqMan
Fast universal PCR master mix (cat. no. 4352042), EGFRvIII-specific
TaqMan MGB probe, GAPDH-specific TaqMan MGB probe and formamide
were obtained from Applied Biosystems (Thermo Fisher Scientific,
Inc.). A QuantiTect reverse transcription kit was purchased from
Qiagen AB (cat. no. 204443). HPLC-purified EGFRvIII-specific
primers, HPLC-purified GAPDH-specific primers, and IDH1 and IDH2
primers were from Integrated DNA Technologies, Inc. Platinum Taq
DNA polymerase (cat. no. 10966-018). PCRx enhancer system (cat. no.
11495-017) was from Invitrogen (Thermo Fisher Scientific, Inc.).
Real-time PCR instrument (ABI 7500 Fast; Thermo Fisher Scientific,
Inc.), capillary sequencing instrument (3500 Genetic analyzer),
speed vac (Eppendorf), NanoDrop 2000C, microAmp optic adhesive film
for PCR plate (7500 Fast) and fast optical 96 well reaction plate
(7500 Fast; cat. no. 4346907) were also purchased from Thermo
Fisher Scientific, Inc.
Clinical specimens and ethics
statement
This study was performed in accordance with the
principles of the Declaration of Helsinki, and was approved by the
Institutional review board bioethics committee of King Abdullah
Medical City (KAMC), Makkah, Kingdom of Saudi Arabia (approval no.
14-140). Before starting the study, a written informed consent was
obtained from all patients or the parent or guardian, if the
patient was a minor. Formalin-fixed, paraffin-embedded (FFPE) tumor
samples were collected for analysis from the Al-Noor Specialty
Hospital Makkah, KAMC Makkah, and King Khalid University Hospital,
King Saud University, Riyadh. In the present study, 165 CNS tumors
were included; the majority of these (146/165) were gliomas and
some were non-gliomas. Non-glioma tumors included one atypical
choroid plexus papilloma, one primitive neuroectodermal tumor
(PNET), one synovial sarcoma, two atypical teratoid rhabdoid tumors
(ATRTs; 10 h and 2 years of age at the time of diagnosis), two
adamantinomatous craniopharyngiomas, three medulloblastomas, four
meningiomas (including one spinal meningioma Psammomatous type) and
five hemangioblastomas. The mean age of the patients was 39 years
(age range, 10 h to 83 years). The patients included 92 males and
73 females; the mean age was 42 and 36 years for males and females,
respectively. The histological classification of tumors was based
on the criteria set by the WHO (27). The computed tomography or magnetic
resonance imaging data of all patients were reviewed by the
consultant radiologist to confirm the diagnosis. One limitation
that needs to be acknowledged in the present analysis of tumor
tissue is obtaining a brain biopsy or FFPE tissue from the healthy
individuals to use as a control; this was difficult, unless there
is a pathological condition the patients cannot be operated for the
brain tissue. For this reason, RNA and DNA isolated from cell lines
were used as the controls.
RNA isolation
The FFPE samples (5–10 sections of 5-µm thickness)
were collected in Eppendorf tubes, and the paraffin was dissolved
using 1.0 ml xylene (2X) at 55°C for 5 min. The supernatant was
discarded by pipetting, and 1.0 ml ethanol (2X) at room temperature
was added to neutralize the residual xylene. The supernatant was
then removed by pipetting, and the pellets were dried at 65°C for 3
min; if any remaining ethanol was present, the heating continued
until all residual ethanol had evaporated. The pellet was
resuspended in 240 µl proteinase K digestion buffer and treated
with 10–20 µl proteinase K. This incubation was carried out at 55°C
for 2–3 h or overnight, and then the incubations were carried out
at 80°C for 15 min, to reverse the RNA crosslinking. Further steps
were carried out as described in the QIAamp FFPE RNA kit (cat. no.
73504). Finally, RNA was eluted in 50–100 µl of RNAase-free water
and stored at −80°C in aliquots for future use.
Primers and probe design for EGFRvIII
and GAPDH
Junction-specific (exon 1 and 8 junctions) RT-qPCR
primers for EGFRvIII were previously published (Table SI) (28). As this sequence is not present in
genomic DNA, only the cDNA target was amplified. Sequencing PCR
primers for EGFRvIII were designed using NCBI-Primer Blast. These
primers were used to validate the positive cases found by RT-qPCR
for EGFRvIII. To monitor the genomic DNA contamination, for GAPDH
mRNA, intron spanning primers were used (Table SI). The forward primer was in exon
2, and reverse primer in exon 4, and the probe is in exon 3. The
probe was a sense probe, which binds to the reverse strand.
RT-qPCR
The final concentration of the reverse and forward
primers was 25 µM. The primers used were as follows: EGFRvIII
forward, 5′-GGCTCTGGAGGAAAAGAAAGGTAATT-3′ and reverse,
5′-CCGTCTTCCTCCATCTCATAGC-3′; GAPDH forward,
5′-GAAGGTGAAGGTCGGAGTC-3′ and reverse, 5′-GATGGGATTTCCATTGATGAC-3′.
MGB probes were prepared at a final concentration of 10 µM, and
stored in aliquots at −20°C or lower, protected from light. The
sequences of TaqMan-MGB probes used were:
6-FAM-5′-TGACAGATCACGGCTC-NFQ-3′ for EGFRvIII; and
VIC-5′-TCACCAGGGCTGCTT-NFQ-3′ for GAPDH. Total RNA was used in a
two-step RT-qPCR system. In this system, each sample was evaluated
for the EGFRvIII and GAPDH genes using MGB probes. First the
reverse transcription reaction was performed in 20 µl with 1 µg of
RNA at 42°C for 30 mins, as described in the QuantiTect reverse
transcription kit manual. The reverse transcription reaction was
stopped by incubation for 3 mins at 95°C. This c-DNA was stored at
−20°C for future use. qPCR was then performed using MGB probes and
TaqMan Fast universal PCR master mix on a 7500 Fast RT-qPCR machine
(ABI 7500; Applied Biosystems; Thermo Fisher Scientific, Inc.). A
total of 2 µl cDNA was used for qPCR in 20 µl of TaqMan Fast
universal PCR master mix containing 1 µl of 25 µM forward and
reverse primers and 1 µl of 10 µM specific probe. The amplification
protocol was as follows: 95°C for 10 min, followed by 40 cycles at
95°C for 15 sec and 60°C for 1 min. Mean Ct values of duplicate
wells were used for data analysis. The relative expression of
EGFRvIII compared with the reference gene (GAPDH) was calculated as
∆Ct (∆-threshold cycle), by subtracting the Ct value of GAPDH from
that of EGFRvIII as previously described (29).
Cell lines used
The human U-87 MG ATCC® HTB-14
glioblastoma (origin unknown) cell line was originally obtained
from the American Type Culture Collection (ATCC). U-87 MG cells
harbouring the DR-GFP DNA repair reporter (U87 MG/DR-GFP) served as
EGFRvIII-negative control cell line (30). As EGFRvIII-positive cells, U87
MG/DR-GFP were infected with EGFRvIII retrovirus WZL-hygro-EGFRvIII
construct and selected for hygromycin resistance. The generation of
these clones has been previously described (30–32). The
DR-GFP plasmid was a gift from Dr Maria Jasin (Memorial Sloan
Kettering Cancer Center, New York, NY, USA; cat. no. 26475;
Addgene, Inc.) and WZL-hygro plasmid was a gift from Dr Scott Lowe
(Memorial Sloan-Kettering Cancer Center, New York, NY, USA; cat.
no. 18750; Addgene, Inc.). RNA and DNA isolated from these cell
lines were used as the positive and negative control in RT-qPCR.
Additionally, RNA isolated from immortalized human myelogenous
leukaemia cell line K562 (cat. no. 04379012001) part of c-DNA
synthesis kit from Roche was used in RT-qPCR as a negative
control.
Validation of RT-qPCR results by
sequencing the PCR product
The RT-qPCR test was validated with Sanger
sequencing for EGFRvIII-positive samples using the Platinum Taq DNA
polymerase (Invitrogen; Thermo Fisher Scientific, Inc.) and GC
enhancer system (Promega Corporation). The following thermocycling
conditions were used: 95°C for 3 min, followed by 40 cycles at 95°C
for 30 sec, 54°C for 30 sec and 68°C for 30 sec, and a final
extension step at 68°C for 5 min. The products were purified and
sequenced in both directions for confirmation. A separate set of
primers flanking the deletion site resulting in a 238-bp PCR
product (113 bp upstream and 29 bp in downstream of 96 bp RT-qPCR
product) was used for this sequencing. The sequencing primers are
shown in Table SI.
Sanger sequencing for IDH1 and
IDH2
The FFPE samples (5–10 sections of 5-µm thickness),
were collected in a 1.5- or 1.7-ml Eppendorf tube. DNA extraction
was performed using QIAamp DNA FFPE tissue kit (Qiagen) following
the protocol described by the manufacturer (cat. no. 56404).
Investigation of mutations in exon 4 of IDH1 and IDH2 were
performed as previously reported, using primers to flank variants
in the IDH1 (GenBank sequence NM_005896) and IDH2 (GenBank sequence
NM_002168) genes (Table SI)
(33). For IDH1, a ~196-bp partial
exon 4 sequence containing hot spots was amplified using a forward
primer within exon 4 and a reverse primer within intron 4, this
product size was 213 bp. For IDH2, a 288-bp sequence with the
entire exon 4 containing R140 and R172 hot spots was amplified
using forward primer within intron 3 and a reverse primer within
intron 4. For PCR 30–50 ng genomic DNA was amplified. The quality
of the PCR product was checked by electrophoresis on agarose (1.2%)
gel, and images were captured on the Bio-Rad's gel documentation
system. PCR products were purified using Agencourt AM-Pure XP kit
(Beckman Coulter, Inc.). All primers were tagged with M13
sequences, and M13 primers were used in sequencing PCR (BigDye
reaction). BigDye reaction products were purified using the BigDye
xTerminator kit (cat. no. 4376484) containing BigDye xTerminator
and SAM solutions (Life Technologies; Thermo Fisher Scientific,
Inc.). The sequencing was then performed on an ABI 3500 Genetic
analyzer (8-capillary sequencer).
NGS analysis on Ion Proton
DNA (10 ng) was used for NGS analysis. DNA was
sequenced using Ion PI v3 chip on Ion Proton instrument. Libraries
were prepared using Ion AmpliSeq primer pools by Ion AmpliSeq™
cancer panel v.1 (cat. no. 4471262), Ion AmpliSeq™ Cancer Hotspot
Panel v2 (cat. no. 4475346), or Comprehensive cancer panel genes
(cat. no. 4477685) (34–37). Ion AmpliSeq 2.0 library and Ion PI
Hi-Q OT2 200 kits were used for library and template preparation,
respectively. Sequencing was performed using an Ion PI Hi-Q
sequencing 200 kit, and the library was tagged with Ion Xpress™
Barcode Adapters. Sixteen tumour samples were pooled per chip run
when libraries were prepared using Ion AmpliSeq™ cancer panel v.1
and Ion AmpliSeq™ Cancer Hotspot Panel v2, and 8 samples were
pooled when comprehensive cancer panel primers were used. Following
sequencing, amplicon sequences were aligned to the human reference
genome GRCh37 (hg19) in the target regions of the cancer panel
using the Torrent Suite v.5.0.2 software (Life Technologies; Thermo
Fisher Scientific, Inc.). Variants call format (vcf) file was
generated by running the Ampliseq™ Variant Caller plug-in v5.2
within the Ion Torrent Suite software. The vcf file data were
analysed using Ion Reporter v5.6 (Thermo Fisher Scientific, Inc.).
True mutations were considered based on a Phred score of >100,
and mutation calling was considered significant by Ion Reporter
software at P<0.05 (36,37).
Results
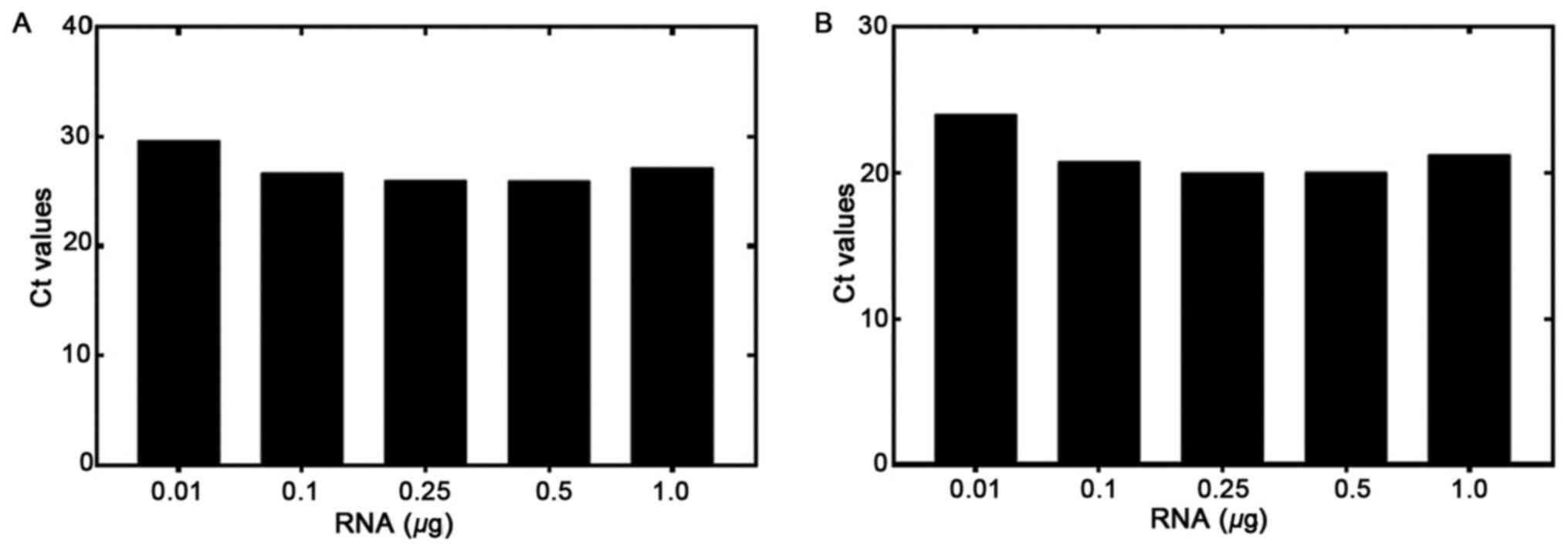
The RT-qPCR sensitivity was tested using various
concentrations of RNA isolated from the EGFRvIII-positive U87
MG/DR-GFP cells. RNA was diluted from 1.0 to 0.01 µg and used for
the RT reaction. RT-qPCR was then performed to detect the levels of
EGFRvIII and GAPDH (Fig. 1A and B,
respectively). The GAPDH Ct at a starting amount of 0.01 µg RNA was
<30 cycles, demonstrating that RNA quality and not quantity was
a more important determinant of assay success. The amplification
plots for both genes are shown in Figs.
S1–S3. The Ct values ranges
from 29.69 to 25.97 for EGFRvIII, and 24.5 to 20.0 for GAPDH, from
0.01 to 1.0 µg RNA, respectively. The difference between the Cts of
the two genes (EGFRvIII and GAPDH) was <7. This showed that the
amount of starting RNA, over a 100-fold range, was not a
significant factor determining assay results.
Amplification plots for RNA from K-562 cells, used
as a negative control, are shown in Fig. S3B. The RT reaction was performed
with 1.0 µg RNA, which was followed by the qPCR reaction for
EGFRvIII and GAPDH respectively. There was no amplification of the
EGFRvIII transcript, whereas GAPDH was amplified (Ct value 22.78).
Similarly, in Fig. S4 demonstrates
the EGFRvIII and GAPDH amplification plots of DNA isolated from
positive control cells (panel A) and negative control cells (panel
B). qPCR results confirmed that only DNA from the positive control
cells amplified both the EGFRvIII and GAPDH genes (Ct values 21.23
and 16.85, respectively), whereas in negative control cells
EGFRvIII was not amplified, but only GAPDH amplification was
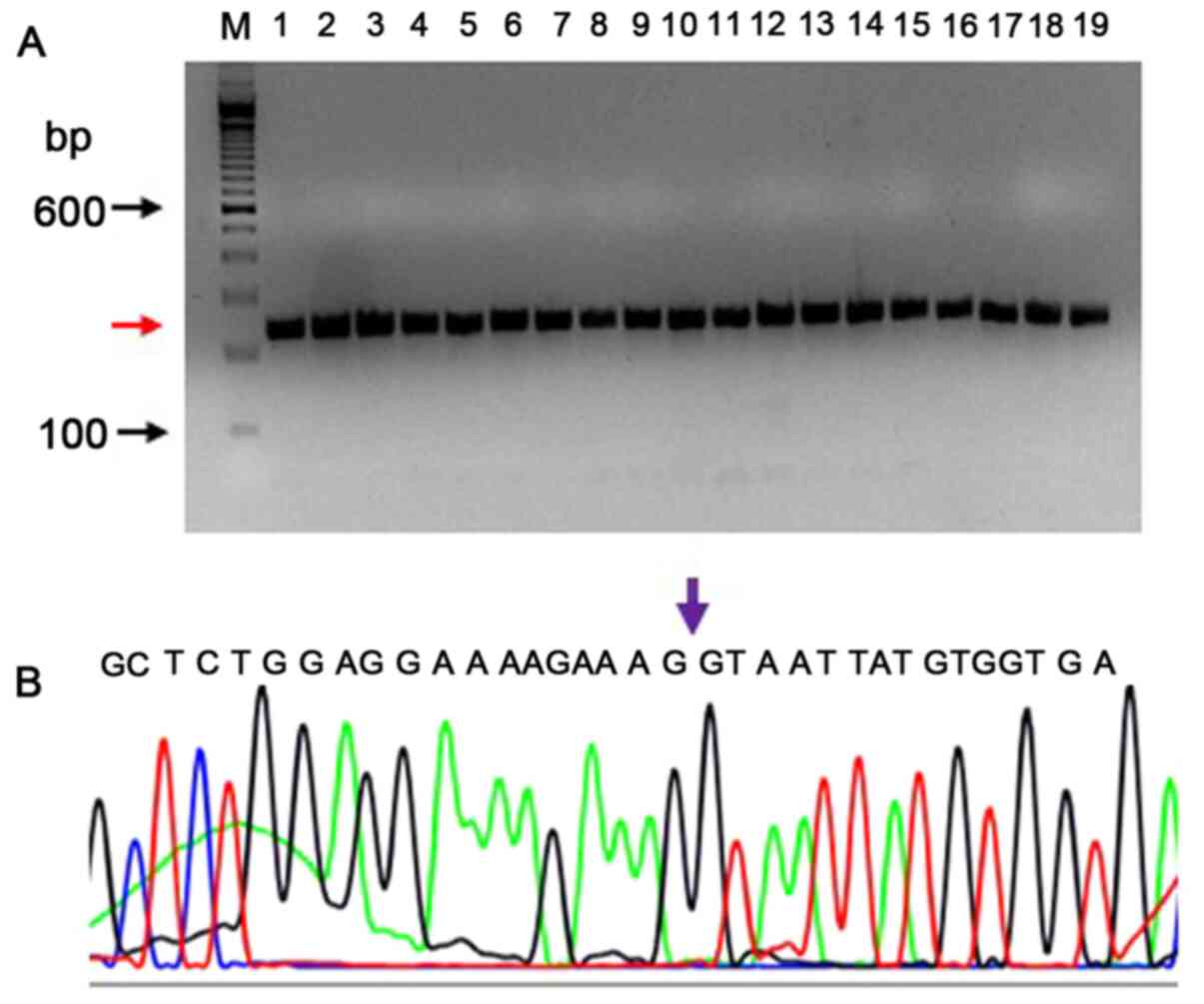
detected (Ct value 19.43). In Fig.
S5 the verification of the EGFRvIII-specific product by PCR is
shown using known positive and negative controls on agarose gel.
PCR products of 238 bp (red arrow) are formed by DNA from positive
cells (lanes 2–10) and no EGFRvIII-specific PCR products (lanes
11–19) were formed in the negative controls. The forward primer is
a junctional primer spanning exons 1–8 (5′-CAGTATTGATCGGGAGAGCC-3′)
and amplifies only the exon 2–7 deletion mutant of EGFR (EGFRvIII);
lane denoted as ‘M’ is a 100 bp DNA ladder.
As shown in Table I,
out of a total of 117 brain tumors tested for EGFRvIII by RT-qPCR,
41 (35%) tumors were positive (Table
I). Out of 55 glioblastoma tumors, which included low-grade
gliomas and GBMs, 28 tumors (50.9%) were positive for EGFRvIII
expression (Table SII). In
addition, 22.7% (5/22) of astrocytoma tumors, which included
pilocytic, anaplastic and diffuse types, 30.8% (4/13) of
oligodendrogliomas and anaplastic oligodendrogliomas, and 14.8%
(4/27) of other brain tumors, including meningiomas,
craniopharyngiomas, medulloblastomas, hemangioblastomas,
gliosarcomas, ependymomas and gangliogliomas were also positive
(Table SIII–SV).
 | Table I.Epidermal growth factor receptor
variant III expression in different brain tumors in Saudi Arabian
population. |
Table I.
Epidermal growth factor receptor
variant III expression in different brain tumors in Saudi Arabian
population.
| Tumor type | Total tested | Positive (n) | Percentage |
|---|
| Glioblastoma | 55 | 28 | 50.9 |
|
Oligodendroglioma | 13 | 4 | 30.8 |
| Astrocytoma | 22 | 5 | 22.7 |
| Others | 27 | 4 | 14.8 |
| Total | 117 | 41 | 35.0 |
The EGFRvIII transcript was sequenced by capillary
electrophoresis to demonstrate the presence of an EGFRvIII-specific
junction where exons 2–7 were deleted. The EGFRvIII expression was
confirmed by sequencing the PCR product on an Applied Biosystem's
sequencing instrument (3500 Genetic analyzer). The sequencing PCR
primers were different from those used in RT-qPCR, in order to
confirm a larger product which extends from downstream to upstream
of the deletion. The RT-qPCR product spanned 20 bp upstream and 76
bp downstream of the deletion. The sequencing PCR product was
238-bp long (113 bp upstream and 29 bp downstream of the 96 bp
RT-qPCR product). A total of 26 DNA samples that were found to be
positive by RT-qPCR were tested to confirm the deletion sequence by
Sanger sequencing. The RNA samples were reverse transcribed, and as
this PCR product had 64.28% of GC content, a PCRx enhancer system
was used with Platinum Taq DNA polymerase. All (26/26) samples
exhibited the correctly-sized product (238 bp; a representative PCR
gel picture is shown in Fig. 2A).
Using the BigDye v3.1 method, the sequences were verified for the
deletion of exons 2–7. A representative electropherogram confirming
the presence of an exon 1 and exon 8 at the junction is shown in
Fig. 2B.
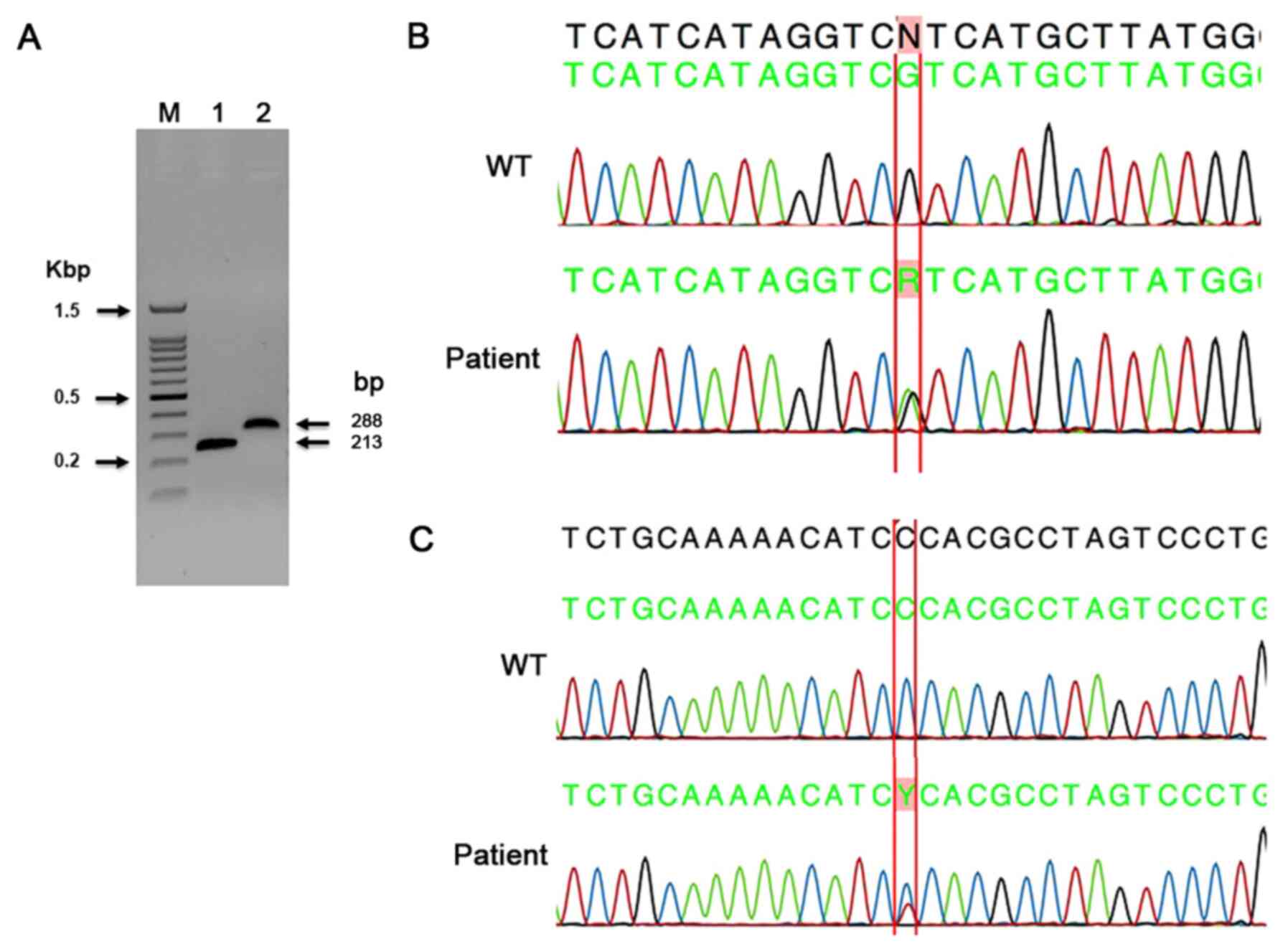
In the present study, 106 tumors were sequenced for
IDH1 exon-4 mutations using the capillary sequencing method. A
summary of all mutations found by capillary sequencing is shown in
Table II. The primers used in the
present study allowed for an interrogation of all three active-site
arginine residues for both IDH1 (R100, R109 and R132) and IDH2
(R140, R149 and R172), as previously reported (38). In the present study, the substitution
missense mutation in c.395G>A; p. (Arg132His) was found in the
majority of the tumors. This mutation was identified in 16 tumor
samples, including 4 oligodendrogliomas, 3 anaplastic
oligodendrogliomas and 1 pilocytic astrocytoma. In addition, 2
astrocytomas, 2 GBMs, 1 diffuse astrocytoma and 1 grade-II
ependymoma were also positive for this mutation (Table II). A representative image of the
PCR gel of IDH1 and IDH2 PCR product sizes (Fig. 3A), and a representative
electropherogram of a known missense mutation in IDH1 c.395G>A;
p. (Arg132His) are shown in Fig. 3B.
In the present study, 45 tumors were sequenced for IDH2 exon-4
mutations using the capillary sequencing method. The most common
IDH2 mutation (Arg172Lys) was not found in any of the present
cases. In a case of adamantinomatous craniopharyngioma, a novel
missense mutation in IDH2 c.472C>T; p. (Pro158Ser) was detected
(Fig. 3C). This novel mutation had
not been reported in CNS tumors so far.
| Table II.Summary of IDH1 and IDH2 mutations
detected by capillary sequencing. |
Table II.
Summary of IDH1 and IDH2 mutations
detected by capillary sequencing.
| Serial number | Lab code | Sex | Age, years | Diagnosis | IDH1 | IDH2 |
|---|
| 1 | 15-N | F | 47 | Glioblastoma
multiforme | c.395G>A; p.
(Arg132His) | ND |
| 2 | HALK-27 | M | 59 | Glioblastoma | c.395G>A; p.
(Arg132His) | ND |
| 3 | 50-N | F | 6 | Oligodendroglioma
(WHO grade-II) | c.395G>A; p.
(Arg132His) | NEG |
| 4 | HALK-10 | F | 17 | Anaplastic
oligodendroglioma | c.395G>A; p.
(Arg132His) | NEG |
| 5 | HALK-25 | F | 25 |
Oligodendroglioma | c.395G>A; p.
(Arg132His) | NEG |
| 6 | HALK-30 | F | 51 |
Oligodendroglioma | c.395G>A; p.
(Arg132His) | NEG |
| 7 | HALK-35 | M | 33 | Anaplastic
oligodendroglioma | c.395G>A; p.
(Arg132His) | ND |
| 8 | HALK-44 | F | 27 | Anaplastic
oligodendroglioma | c.395G>A; p.
(Arg132His) | NEG |
| 9 | KAMC-41 | M | 42 |
Oligodendroglioma | c.395G>A; p.
(Arg132His) | ND |
| 10 | HALK-11 | M | 1 | Diffuse
astrocytoma | c.395G>A; p.
(Arg132His) | NEG |
| 11 | KAMC-33 | M | 36 | Pilocytic
astrocytoma | c.395G>A; p.
(Arg132His) | ND |
| 12 | KAMC-37 | M | 58 | Pilocytic
astrocytoma | c.395G>A; p.
(Arg132His) | ND |
| 13 | KAMC-50 | F | 9 | Astrocytoma | c.395G>A; p.
(Arg132His) | ND |
| 14 | KAMC-51 | M | 6 | Astrocytoma | c.395G>A; p.
(Arg132His) | ND |
| 15 | 13-N | M | 19 | Pilocytic
astrocytoma (WHO grade-I) | c.395G>A; p.
(Arg132His) | ND |
| 16 | 49-N | M | 35 |
Craniopharyngioma | NEG | c.472C>T; p.
(Pro158Ser) |
| 17 | KAMC-28 | F | 39 | Ependymoma (WHO
grade-II) | c.395G>A; p.
(Arg132His) | ND |
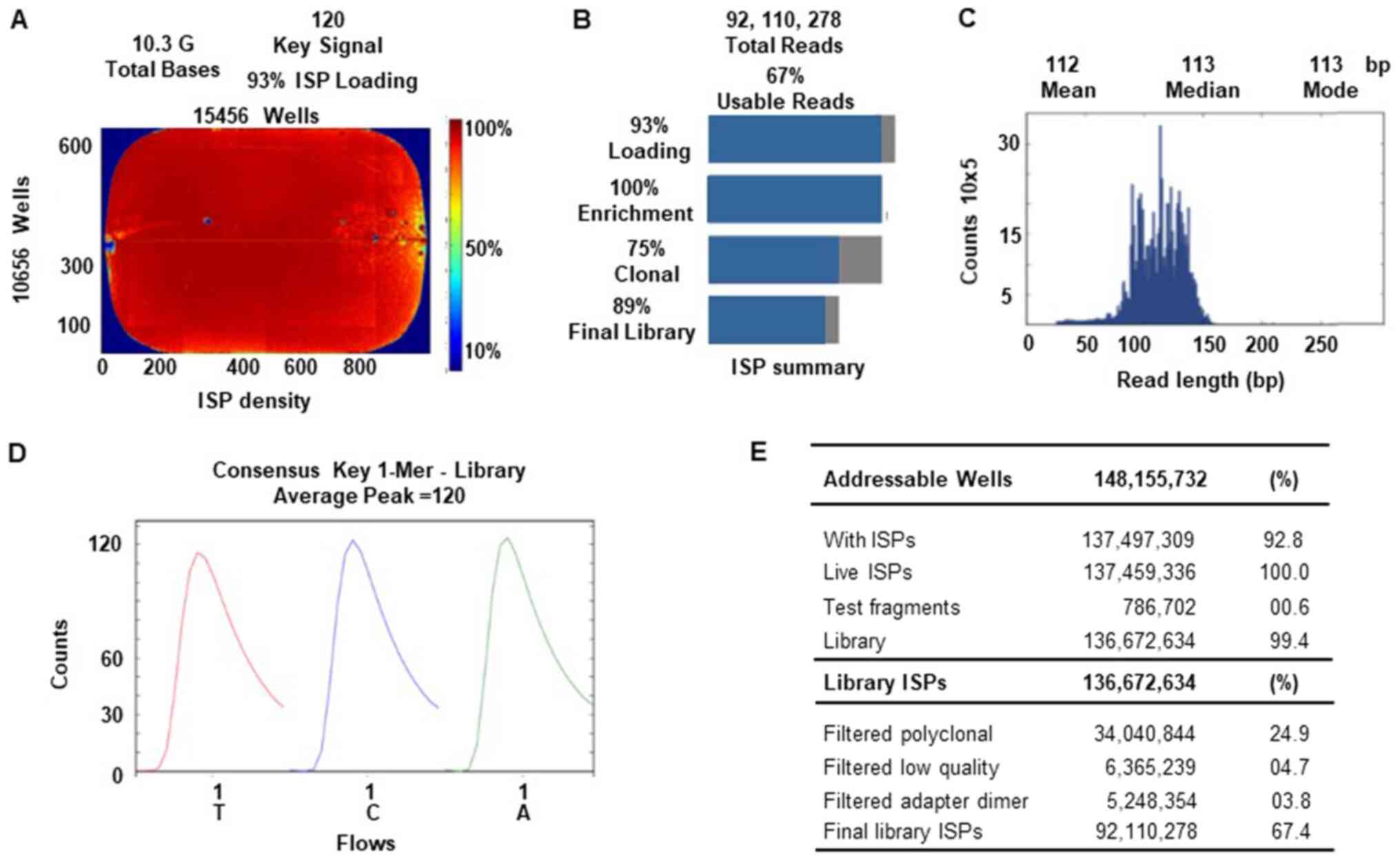
An Ion Torrent Suite v.5.0.2-generated Ion PI chip
run report metrics of NGS sequencing is shown in Fig. 4. The Ion sphere particle (ISP)
density image shows the semi-conductor chip loading across the
wells that contain live ISPs; the total number of bases reported in
the output file were 10.3 G (Fig.
4A). An ISP summary showing loading, enrichment, clonality and
final library quality is shown in Fig.
4B. In the histogram, the first row shows 93% of loaded wells,
of which 7% were empty, 100% were wells with a predicted number of
live library ISPs (enrichment), 75% were clonal ISPs and 25% were
polyclonal ISPs (ISPs carrying clones from >2 templates). The
mean sequencing read length (trimmed lengths) of all reads was 112
bp (Fig. 4C). In Fig. 4D and E, the key signal (i.e.
percentage of live ISPs readings for flows of the bases T, C and A
in the library key), and the numerical values of addressable wells
and library details are shown respectively.
A summary of IDH1 and IDH2 mutations identified by
the NGS method is shown in Tables
II–V. A total of 74 tumors were
sequenced for the IDH1 gene by NGS, and 8 missense variants were
found in 36 tumors in a Saudi Arabian population (Tables III–V). A codon 132 mutation (c.395G>A) was
detected in 29/36 (80.5%) tumor samples by the NGS method. Other
missense mutations in codons c.352C>T, c.368G>A c.369A>G,
c.380C>T, c.394C>A and c.709T>C, each were found in 1
tumor, and 2 tumors were found to contain another missense mutation
in c.356G>A. A novel intronic mutation (c.414+9T>A) was found
in 13 tumors in the IDH1 gene by the NGS method (Tables III and IV). As shown in Tables III and IV, 8 tumors were found to have synonymous
mutations (rs11554137) in c.315C>T; p. (Gly105Gly), and 1 to
have a novel synonymous mutation in c.369A>G; p. (Gly123Gly). A
total of 11 tumors were found to have compound mutations in the
IDH1 gene (Table IV). One case of
astrocytoma and a case of GBM tumor showed a compound, an intronic
and a synonymous mutation (c.414+9T>A; and c.395G>A). Also, a
PNET case contains a compound mutation in exons 2 and 5
respectively (Table IV). Compound
variants were also found in each case of pilocytic astrocytoma,
gliosarcoma and grade-II ependymoma tumors. The allele frequency,
Phred score and P-values for all variants are shown in Tables III–V. One case of ATRT contained a synonymous
mutation p. (Ser332Ser), and an intronic variant in IDH2 gene
(Table V). A total of 6/45 of the
screened cases had 16 mutations in the IDH2 gene (Table V). Nine of these variants were
missense mutations, 5 were synonymous and 1 was intronic. However,
all these IDH2 mutations were present as compound mutations with
IDH1 gene mutations. One case of glioma had a mutation in exon 4 of
the IDH1 gene and 6 compound mutations in exon 4 of the IDH2 gene
also (Table V). However, the allele
frequencies were low for these mutations.
| Table V.Summary of the IDH1 and IDH2 compound
mutations detected by next-generation sequencing. |
Table V.
Summary of the IDH1 and IDH2 compound
mutations detected by next-generation sequencing.
| Serial number | Lab code | Sex | Age, years | Diagnosis | Variant type | Gene name | Frequency,% | Phred score | Coverage | P-value |
|---|
| 1 | KAMC-15 | F | 62 | GBM | c.395G>A; p.
(Arg132His) | IDH1 | 19.41 | 2940.93 | 1999 | 0.00001 |
| 2 | KAMC-15 | F | 62 | GBM | c.368G>A, p.
(Gly123Glu) | IDH1 | 3.3 | 85.9895 | 2000 | 0.00001 |
| 3 | KAMC-15 | F | 62 | GBM | c.453C>T; p.
(Pro151Pro)a | IDH2 |
7.96 |
359.68 | 1986 | 0.00001 |
| 4 | KAMC-15 | F | 62 | GBM | c.420G>A; p.
(Arg140Arg)a | IDH2 |
6.01 | 176.401 | 1997 | 0.00001 |
| 5 | KAMC-15 | F | 62 | GBM | c.410G>A; p.
(Gly137Glu) | IDH2 |
9.59 | 548.867 | 1991 | 0.00001 |
| 6 | KAMC-20 | M | 73 | Glioma | c.395G>A; p.
(Arg132His) | IDH1 | 12.81 |
1488.5 | 1999 | 0.00001 |
| 7 | KAMC-20 | M | 73 | Glioma | c.500C>T; p.
(Pro167Leu)a | IDH2 |
3.45 | 24.9168 | 1999 | 0.00322 |
| 8 | KAMC-20 | M | 73 | Glioma | c.487G>A; p.
(Gly163Ser)a | IDH2 |
5.65 | 148.411 | 2000 | 0.00001 |
| 9 | KAMC-20 | M | 73 | Glioma | c.483C>T; p.
(Gly163Gly)a | IDH2 |
4.05 | 49.1264 | 2000 | 0.00001 |
| 10 | KAMC-20 | M | 73 | Glioma | c.476G>A; p.
(Arg159His) | IDH2 |
5.81 | 160.295 | 1998 | 0.00001 |
| 11 | KAMC-20 | M | 73 | Glioma | c.475C>T; p.
(Arg159Cys) | IDH2 | 4.4 | 67.3793 | 1999 | 0.00001 |
| 12 | KAMC-20 | M | 73 | Glioma | c.404C>T; p.
(Pro135Leu)a | IDH2 |
5.25 | 119.363 | 2000 | 0.00001 |
| 13 | KAMC-44 | M | 53 | GBM | c.395G>A; p.
(Arg132His) | IDH1 | 26.03 | 4639.97 | 1998 | 0.00001 |
| 14 | KAMC-44 | M | 53 | GBM | c.368G>A; p.
(Gly123Glu) | IDH1 |
5.05 | 246.285 | 2000 | 0.00001 |
| 15 | KAMC-44 | M | 53 | GBM | c.519C>T; p.
(His173His)a | IDH2 | 13.44 | 1063.23 | 1942 | 0.00001 |
| 16 | KAMC-44 | M | 53 | GBM | c.459C>G; p.
(Ile153Met)a | IDH2 | 12.71 |
954.66 | 1944 | 0.00001 |
| 17 | 71-N | M | 35 |
Astrocytoma-(grade-II) | c.395G>A; p.
(Arg132His) | IDH1 | 37.84 | 8160.23 | 1998 | 0.00001 |
| 18 | 71-N | M | 35 |
Astrocytoma-(grade-II) | c.448G>A; p.
(Glu150Lys)a | IDH2 |
5.15 | 112.741 | 2000 | 0.00001 |
| 19 | 71-N | M | 35 | Astrocytoma
(grade-II) | c.409G>A; p.
(Gly137Arg) | IDH2 |
3.75 | 36.0244 | 2000 | 0.00025 |
| 20 | 62-N | M | 63 | GBM | c.512G>A; p.
(Gly171Asp) | IDH2 | 2.2 | 22.7072 | 2000 | 0.00536 |
| 21 | 62-N | M | 63 | GBM | c.395G>A; p.
(Arg132His) | IDH1 |
6.46 | 416.312 | 1997 | 0.00001 |
| 23 | 91-N | M | 10b | ATRT grade-IV | c.996C>T; p.
(Ser332Ser)a | IDH2 | 51.34 | 2138.71 | 411 | 0.00001 |
| 24 | 91-N | M | 10b | ATRT grade-IV |
c.116-65T>Ca | IDH2 | 44.07 | 2328.36 | 565 | 0.00001 |
| Table III.IDH1 missense and intronic mutation
status in brain tumors of Saudi Arabian population analyzed by
next-generation sequencing. |
Table III.
IDH1 missense and intronic mutation
status in brain tumors of Saudi Arabian population analyzed by
next-generation sequencing.
| Serial number | Lab code | Sex | Age, years | Diagnosis | IDH1 | Exon | Allele frequency,
% | Phred score | Coverage | P-value | IDH2 |
|---|
| 1 | KAMC-12 | F | 51 | GBM | c.395G>A; p.
(Arg132His) | 4 |
9.66 |
901.25 | 1988 | 0.00001 | NEG |
| 2 | 83-N | M | 25 | Ependymoma (WHO
grade-II) | c.395G>A; p.
(Arg132His) | 4 | 17.72 | 2519.02 | 1981 | 0.00001 | NEG |
| 3 | HALK-36 | M | 13 | Pilocytic
astrocytoma | c.394C>A; p.
(Arg132Ser) | 4 | 35.72 | 4976.78 | 1996 | 0.00001 | ND |
| 4 | HALK-19 | F | 40 | Gliosarcoma | c.395G>A; p.
(Arg132His) | 4 | 10.16 | 986.822 | 1998 | 0.00001 | NEG |
| 5 | 61-N | M | 7 |
Medulloblastoma | c.315C>T; p.
(Gly105Gly) | 4 | 9.6 | 350.583 | 2000 | 0.00001 | NEG |
| 6 | 63-N | M | 63 | GBM | c.352C>T; p.
(Pro118Ser) | 4 |
6.26 | 388.295 | 1934 | 0.00001 | NEG |
| 7 | 65-N | M | 13 | GBM-grade-IV | c.315C>T; p.
(Gly105Gly) | 4 | 59.53 | 11659.2 | 1999 | 0.00001 | NEG |
| 8 | 46-N | F | 38 | GBM-grade-IV | c.356G>A, p.
(Arg119Gln) | 4 | CC=0.82 | 49.4243 | 1941 | 0.00001 | NEG |
| 9 | 47-N | M | 62 | GBM-grade-IV | c.356G>A, p.
(Arg119Gln) | 4 | CC=0.51 | 69.8561 | 1967 | 0.00001 | NEG |
| 10 | 51-N | M | 73 | GBM-grade-IV | c.315C>T; p.
(Gly105Gly) | 4 | 90.91 | 328.334 | 44 | 0.00001 | ND |
| 11 | HALK-13 | M | 7 | Pilocytic
astrocytoma | c.395G>A; p.
(Arg132His) | 4 |
5.50 | 298.817 | 2000 | 0.00001 | NEG |
| 12 | HALK-37 | M | 5 | Pilocytic
astrocytoma | c.395G>A; p.
(Arg132His) | 4 |
3.71 | 116.617 | 1997 | 0.00001 | NEG |
| 13 | HALK-42 | F | 4 | Pilocytic
astrocytoma | c.395G>A; p.
(Arg132His) | 4 |
3.05 | 67.7037 | 1999 | 0.00001 | NEG |
| 14 | HALK-46 | M | 32 | Ganglioglioma | c.395G>A; p.
(Arg132His) | 4 |
3.85 | 129.056 | 1999 | 0.00001 | NEG |
| 15 | KAMC-2 | M | 67 |
Oligodendroglioma | c.395G>A; p.
(Arg132His) | 4 | 19.35 | 2918.86 | 1995 | 0.00001 | NEG |
| 16 | KAMC-47 | F | 75 | GBM | c.395G>A; p.
(Arg132His) | 4 |
9.00 | 792.927 | 1988 | 0.00001 | NEG |
| 17 | HALK-25 | F | 25 |
Oligodendroglioma | c.395G>A; p.
(Arg132His) | 4 | 43.06 |
9872.2 | 1995 | 0.00001 | NEG |
| 18 | HALK-30 | F | 51 |
Oligodendroglioma | c.395G>A; p.
(Arg132His) | 4 | 40.07 |
8890.1 | 1999 | 0.00001 | NEG |
| 19 | HALK-35 | M | 33 | Anaplastic
oligodendroglioma | c.395G>A; p.
(Arg132His) | 4 | 35.97 | 7562.31 | 1996 | 0.00001 | NEG |
| 20 | 40-N | F | 59 | Meningioma-(WHO
grade-I) |
c.414+9T>Aa | Intronic | 75 | 1001.64 | 200 | 0.00001 | ND |
| 21 | 73-N | F | 65 | Meningioma-(WHO
grade-I) |
c.414+9T>Aa | Intronic | 72.28 | 469.808 | 101 | 0.00001 | ND |
| 22 | 75-N | M | 65 | Meningioma-(WHO
grade-I) |
c.414+9T>Aa | Intronic | 80.9 | 526.674 | 89 | 0.00001 | ND |
| 23 | 72-N | M | 14 | GBM |
c.414+9T>Aa | Intronic | 77.07 | 829.327 | 157 | 0.00001 | ND |
| 24 | 78-N | M | 83 | GBM- (WHO
grade-VI) |
c.414+9T>Aa | Intronic | 84.92 | 816.555 | 126 | 0.00001 | ND |
| 25 | 79-N | M | 3 | Meningioma-(WHO
grade-I) |
c.414+9T>Aa | Intronic | 84.62 | 333.176 | 52 | 0.00001 | ND |
| 26 | 80-N | F | 31 |
Hemangioblastoma |
c.414+9T>Aa | Intronic | 79.1 | 376.734 | 67 | 0.00001 | ND |
| 27 | KAMC-52 | M | 55 | GBM-(WHO
grade-VI) |
c.414+9T>Aa | Intronic | 68.18 | 273.568 | 66 | 0.00001 | ND |
| 28 | 76-N | F | 72 | GBM- (WHO
grade-VI) |
c.414+9T>Aa | Intronic | 66.67 | 160.187 | 39 | 0.00001 | ND |
| 29 | HALK-2 | M | 45 | GBM |
c.414+9T>Aa | Intronic | 78.85 | 576.863 | 104 | 0.00001 | ND |
| 30 | HALK-3 | M | 56 | GBM |
c.414+9T>Aa | Intronic | 79.05 | 583.85 | 105 | 0.00001 | ND |
| 31 | KAMC-10 | F | 60 | Astrocytoma | c.395G>A; p.
(Arg132His) | 4 |
8.68 | 742.359 | 1994 | 0.00001 | NEG |
| 32 | KAMC-31 | M | 28 | Astrocytoma | c.395G>A; p.
(Arg132His) | 4 |
7.67 | 588.959 | 1995 | 0.00001 | NEG |
| 33 | KAMC-50 | F | 9 | Astrocytoma | c.395G>A; p.
(Arg132His) | 4 | 43.54 |
6950 | 1998 | 0.00001 | NEG |
| 34 | HALK-41 | M | 51 | GBM | c.395G>A; p.
(Arg132His) | 4 | 10.19 | 995.207 | 1993 | 0.00001 | NEG |
| Table IV.IDH1 compound mutation status in
brain tumors of Saudi Arabian population analyzed by
next-generation sequencing. |
Table IV.
IDH1 compound mutation status in
brain tumors of Saudi Arabian population analyzed by
next-generation sequencing.
| Serial number | Lab code | Sex | Age, years | Diagnosis | IDH1 | Exon | Allele frequency,
% | Phred score | Coverage | P-value | IDH2 |
|---|
| 1 | HALK-45 | F | 38 | Gliosarcoma | c.395G>A; p.
(Arg132His) | 4 |
5.01 | 242.779 | 1996 | 0.00001 | NEG |
| 2 | HALK-45 | F | 38 | Gliosarcoma | c.315C>T; p.
(Gly105Gly) | 4 | 30.18 | 5818.57 | 1998 | 0.00001 | NEG |
| 3 | HALK-48 | M | 68 | GBM | c.395G>A; p.
(Arg132His) | 4 |
3.66 | 113.809 | 1995 | 0.00001 | NEG |
| 4 | HALK-48 | M | 68 | GBM | c.315C>T; p.
(Gly105Gly) | 4 | 33.35 | 6742.06 | 1994 | 0.00001 | NEG |
| 5 | HALK-39 | M | 55 | Low grade
glioma | c.369A>G; p.
(Gly123Gly)a | 4 |
5.26 | 119.741 | 1998 | 0.00001 | NEG |
| 6 | HALK-39 | M | 55 | Low grade
glioma | c.315C>T; p.
(Gly105Gly) | 4 | 4 | 143.241 | 2000 | 0.00001 | NEG |
| 7 | 84-N | M | 9 | Pilocytic
astrocytoma | c.395G>A; p.
(Arg132His) | 4 | 18.82 | 2798.99 | 1998 | 0.00001 | NEG |
| 8 | 84-N | M | 9 | Pilocytic
astrocytoma | c.368G>A; p.
Gly123Glu) | 4 | 3 | 65.2141 | 2000 | 0.00001 | NEG |
| 9 | KAMC-27 | M | 42 | GBM | c.395G>A; p.
(Arg132His) | 4 | 20.92 | 2191.56 | 1319 | 0.00001 | NEG |
| 10 | KAMC-27 | M | 42 | GBM | c.394G>A; p.
(Arg132Ser) | 4 | 33.75 | 6868.22 | 1994 | 0.00001 | NEG |
| 11 | HALK-44 | F | 27 | Anaplastic
oligodendroglioma | c.315C>T; p.
(Gly105Gly) | 4 | 43.57 |
10052 | 1997 | 0.00001 | NEG |
| 12 | HALK-44 | F | 27 | Anaplastic
Oligodendroglioma | c.395G>A; p.
(Arg132His) | 4 | 36.95 | 7849.08 | 1992 | 0.00001 | NEG |
| 13 | 92-N | F | 8 | PNET | c.709T>C; p.
(Ser237Pro)a | 5 | 47.22 | 472.971 | 108 | 0.00001 | NEG |
| 14 | 92-N | F | 8 | PNET | c.395G>A; p.
(Arg132His) | 4 |
0.79 |
21.675 | 252 | 0.0068 | NEG |
| 15 | HALK-27 | M | 59 | GBM |
c.414+9T>Aa | Intronic | 70.93 | 384.357 | 86 | 0.00001 | ND |
| 16 | HALK-27 | M | 59 | GBM | c.395G>A; p.
(Arg132His) | 4 | 49 | 809.429 | 400 | 0.00001 | ND |
| 17 | 77-N | M | 14 | Astrocytoma |
c.414+9T>Aa | Intronic | 85.29 | 221.041 | 34 | 0.00001 | ND |
| 18 | 77-N | M | 14 | Astrocytoma | c.395G>A; p.
(Arg132His) | 4 | 42.5 | 69.0889 | 40 | 0.00001 | ND |
| 19 | KAMC-39 | F | 39 |
Oligoastrocytoma | c.395G>A; p.
(Arg132His) | 4 |
6.41 | 411.865 | 1996 | 0.00001 | NEG |
| 20 | KAMC-39 | F | 39 |
Oligoastrocytoma | c.315C>T p.
(Gly105Gly) | 4 |
6.21 | 384.588 | 1998 | 0.00001 | NEG |
| 21 | 54-N | F | 14 |
Medulloblastoma | c.380C>T; p.
(Pro127Leu) | 4 |
3.97 | 45.0045 | 1992 | 0.00003 | NEG |
| 22 | 54-N | F | 14 |
Medulloblastoma | c.315C>T p.
(Gly105Gly) | 4 | 34.1 | 4605.54 | 2000 | 0.00001 | NEG |
The EGFRvIII expression status in the IDH1 and IDH2
mutated cases is shown in the Table
SVI. Five of the tumors contain both IDH1 and IDH2 mutations
along with EGFRvIII expression also. Out of 36 tumors with IDH1
mutations, 14 case showed EGFRvIII expression, and 22 cases were
negative for EGFRvIII. In GBM tumor category 5/14 tumors were
positive and 9 were negative. Furthermore, in oligodendrogliomas
including anaplastic type 2/8, and in astrocytoma's including
pilocytic and diffuse type, 5/6 were positive for EGFRvIII
respectively. In medulloblastoma 1/2 tumor was positive; and each
case of adamantinomatous craniopharyngioma, ganglioglioma, and two
case of gliosarcoma tumors have IDH1 mutations but they were
negative for EGFRvIII.
Discussion
Studies describing the prevalence of IDH mutations
and its associations with overall survival and tumor progression in
the Saudi Arabian population are limited. The fields of genomic
medicine and targeted therapy are new in the Kingdom of Saudi
Arabia. Mutations in the IDH1 and IDH2 genes have been shown to be
predictive markers for favorable clinical outcomes in gliomas; it
has also been shown that PARP inhibitors enhance the radio
sensitization of glioma cells with an IDH mutation (38,39). The
clinical trials on IDH1 inhibitors, such as Ivosidenib
(TIBSOVO®), have recently yielded successful results for
acute myeloid leukemia (AML) cases with an IDH1 gene mutation;
however, this approach remains unclear for gliomas. As IDH-mutant
astrocytomas have a more favorable survival, they may require a
less aggressive treatment approach. Tumors without IDH mutations
may have other genetic abnormalities characteristic of GBM that
predict an aggressive clinical course and require an intensified
treatment protocol. Several studies have shown that the IDH1 exon 4
mutations in grade-II and -III gliomas and secondary glioblastomas
are in fact common (70%), and are less frequent in primary
glioblastomas (29.4%) (9,35,40,41). In
the present study, 71% (5/7) of anaplastic oligodendrogliomas, 75%
(6/8) of astrocytomas and 31% (5/16) of pilocytic astrocytomas
contained this exon 4 missense mutation, which was in agreement
with previous reports (35,41,42).
However, in our glioblastoma tumors, which included grade-IV GBM,
the mutation detection rate was 43% (14/32) which is higher than
that found in previous studies (15,42,43). The
most common Arg132His mutation accounted for 92.7% of all exon 4
mutations in the IDH1 gene, with other mutations in this codon such
as Arg132Cys, Ser, Gly, Leu and Val being very rare (9). In the present study, medulloblastoma
(1/2) and grade-II ependymoma (1/1) tumors were found to have the
missense mutation, and 2 of these medulloblastoma tumors also
contained the synonymous mutations. Previous studies have reported
that in meningiomas, medulloblastomas and ependymal tumors, IDH
mutations were absent (42,44). Previously, a case of anaplastic
grade-III ependymoma with a c.395G>A mutation was reported
(37). Few studies have reported
IDH1 codon 132 (Arg132His) mutations in PNET cases (42). In the present study, a novel mutation
in c.709T>C; p. (Ser237Pro) was detected in exon 5 of 1 PNET
case, which had not been reported in the COSMIC or ExAC databases;
a codon 132 mutation was also present in this tumor. In the
majority of cases, the arginine in position 132 was replaced by
histidine (Arg132His) in IDH1, and in IDH2 the arginine residue at
amino acid codon 172 or codon 140 was mutated (45). Mutations affected in these codons,
which belongs to an evolutionary conserved region of the isocitrate
binding site. Using the capillary and NGS methods, it was also
found in the present study that c.395G>A was the major mutation
in this Saudi Arabian population. This mutation in IDH1 (COSM28746)
had a FATHMM score of 0.94 and Polyphen score of 0.127, suggesting
it is a deleterious mutation; this mutation is also listed in the
ClinVar database as pathogenic. Other IDH1 missense mutations found
in the present studied cases, such as c.394G>A, c.368G>A, and
c.356G>A, were also described in the COSMIC database as
pathogenic. The novel IDH1 synonymous mutation c.369 A>G, found
in low-grade glioma, had not previously been reported in the SNP
and COSMIC databases; however, another synonymous variant,
c.315C>T, was reported in the ClinVar database as a benign one.
Recently, a study describing the prevalence of IDH mutations
reported from Saudi Arabian patients with glioma; Alassiri et
al (46) have shown that the
MGMT promoter methylation, and IDH1 mutations and their
associations with survival. However, in their study, the IDH1 was
analyzed by IHC/qPCR targeting limited codons, and MGMT methylation
was performed at the Mayo clinic. Alissiri et al, have
reported that out of 65 cases screened for the IDH mutation, 6
(9.2%) tested positive. In another study this mutation rate was
33.87% by droplet digital PCR, 27.42% RT-qPCR, and 30.65% by Sanger
direct sequencing (47). By NGS
method this mutation rate was found to be ~47%, and by Sanger
sequencing this was 15%. Recent advances are transferring many of
the molecular tests to the NGS platforms for its sensitivity and
accuracy in mutation detection (48). The synonymous IDH1 SNP (rs11554137;
c.315C>T) mutations found in the present study were also shown
to have an adverse prognosis in Egyptian adult patients with AML
(49). This synonymous mutation, p.
(Gly105Gly), and the intronic c.414+9T>A variants were found
mostly is meningioma and glioblastoma tumors, but few cases of
medulloblastoma, gliosarcoma, hemangioblastoma and anaplastic
oligodendroglioma also contained these variants.
It has been shown that mutations in the IDH1 and
IDH2 genes are mutually exclusive in gliomas, and mutations in IDH2
are mutually exclusive with PTEN, P53 and ATRX mutations (10,41,50). As
compared with the IDH1 gene, mutations in the IDH2 gene are less
prevalent in gliomas (40,45). The IDH2 missense mutation in
c.410G>A; and c.476G>A are pathogenic mutations; these two
variants were reported only twice each in the COSMIC database, in
cutaneous melanoma, and in bladder urothelial carcinoma, and
endometrial carcinomas respectively. Other IDH2 missense mutations
in c.409G>A and c.475C>T were also reported in 1 sebaceous
neoplasm and 1 bladder carcinoma, respectively, and they are
pathogenic. This is the first report describing these 4 variants in
brain tissue. Four of the IDH2 missense mutations detected in the
present study (in c.404C>T, c.448G>A, c.459C>G,
c.487G>A and c.512C>T) and the 5 synonymous variants (in
c.420G>A, c.453C>T, c.483C>T and c.519C>T), as well as
a synonymous mutation in c.996C>T and an intronic variant in
c.116-65T>C were not reported in the COSMIC database; these
novel mutations in the IDH2 gene were identified in the present
study. In 1 GBM, the IDH2 synonymous mutation detected in this
study in c.519C>T; p. (His173His) had also been reported in an
atypical choroid plexus papilloma tumor (36). In the present study, no missense
mutations were detected in IDH2 amino acid codons 172 and 140;
however, all IDH2 mutations found were in exon 4, with the
exception of c.996C>T; p. (Ser332Ser), which was present in exon
8 of an ATRT case. The c.116-65T>C variant detected in this case
was present in intron 1. Only 1 synonymous mutation was present in
codon 140 [c.420G>A; p. (Arg140Arg)] in exon 4. The novel exon 4
mutation in c.472C>T; p. (Pro158Ser) detected in the IDH2 gene
by capillary sequencing was not reported in CNS tumors, and has
only been reported in a case of pancreatic ductal adenocarcinoma so
far (51).
Previous investigation revealed that EGFRvIII
expression predominated in primary glioblastomas and in high grade
GBMs but was rare in secondary glioblastomas. Furthermore, IDH1
mutation are rare in primary glioblastomas and they are common in
secondary glioblastomas (52,53). By
NGS analysis it was observed that 32% (12/37) of GBMs that includes
glioblastomas also, and in 69% (11/16) of astrocytoma's tumors
including pilocytic, anaplastic, and diffuse type, and 63% (5/8) of
oligodendrogliomas (all grades) showed IDH1 c.395G>A mutations.
One patient had right frontal lobe tumor with a history of GBM
grade-IV, who was operated for decompression by excision biopsy.
This tumor was tested negative for EGFRvIII, nine months post
surgically this case had a recurrence and the tumor became positive
for EGFRvIII. Immunohistochemistry results also positive for EGFR,
FISH results showed EGFR amplification, and Ki 67 proliferation
index was high in this tumor, suggesting EGFRvIII expression may
change in a subset of patients at recurrence (54). This case had IDH1 mutation in
c.352C>T preoperatively, and post-operatively showed mutations
in both IDH1 in c.395G>A and in c.512G>A of IDH2 gene.
Patients with meningiomas generally have a good prognosis, the
present study identified that one grade-I secretary meningioma case
doesn't contain IDH1 mutation and it is EGFRvIII-negative. In a
total of 22 astrocytomas, two anaplastic tumors (grade-III)
including one recurrent, two pilocytic (grade-I), and one diffuse
type (grade-II) were positive for EGFRvIII. All these tumors were
also positive for IDH1 mutation. One case of astrocytoma grade-II
was negative for EGFRvIII, but this tumor had IDH1 c.395G>A, and
IDH2 mutations in c.448G>A, and in c.409G>A which are
pathogenic. Of a total of 13 oligodendrogliomas, four tumors that
includes two anaplastic (grade-III) types are positive for both
EGFRvIII and IDH1. In an adamantinomatous craniopharyngioma grade-I
case which is negative for IDH1 mutation and EGFRvIII and no
evidence of tumor recurrence after craniotomy. A case of ependymoma
(grade-II) had an IDH1 exon-4 mutation and positive for EGFRvIII,
another medulloblastoma (grade-IV) case had an IDH1 different
missense mutation in c.380C>T and positive for EGFRvIII.
It has been postulated that the EGFRvIII detection
may help distinguish patients with glioma who will respond to TKIs
therapy, which makes this mutant an interesting target for
immunotherapy too (55). These
clinical trials showed promise in early phase II clinical trials;
however, at later stages, the results were not promising (56,57).
Such molecular targeted approaches for clinical trials have made
the detection of EGFRvIII a priority, at least for the time being
(58). Thus, few molecular
laboratories are providing RT-qPCR for EGFRvIII; however, the
utility of this assay is limited to clinical trials, since the
targeted therapy for this molecule is at an early stage.
Immunochemical detection and southern blotting are not very
sensitive and accurate compared to molecular techniques like PCR
and RT-qPCR. Frozen tissue is not routinely available due to
transportation delays, and the instability of RNA causes
difficulties; therefore, the development of molecular tests for
EGFRvIII detection in the FFPE tissue is a convenient approach
(28,29).
In The Cancer Genome Atlas (TCGA) samples, this
mutated form of EGFRvIII was reported to be present in 24% of GBMs,
and this was reported in 50–60% of EGFR-overexpressing GBMs
(45,59). In the present study this deletion was
found in 41 (35%; n=117) tumors that includes all brain tumors.
Only 51% of GBMs, 23% of astrocytomas, including diffuse and
pilocytic astrocytomas, and 31% of anaplastic oligodendrogliomas
were positive for EGFRvIII. In addition, 1 case of each from
medulloblastoma, atypical ganglioglioma, reactive gliosis and
ependymoma were positive. Previous studies found that nearly 34.5%
of tumors, all grades of glioma, including astrocytoma and
pilocytic astrocytoma tumors, express EGFRvIII, a percentage high
enough for therapeutic targeting (60). Previous studies have also reported
the EGFRvIII expression in ependymomas, medulloblastomas, ATRT
grade-IV, oligoastrocytomas grade-III and anaplastic astrocytoma's
(61,62). The present data from a Saudi
population is in concordance to these published reports.
NGS screening for the diagnosis of glioma is already
provided in many genetics' laboratories. The NGS method detected
more mutations than the capillary method in this study, suggesting
the advantage of the NGS screening technique on all exons of IDH
genes. That may have been the reason compound mutations were not
detected in previous studies that used capillary sequencing. The
IDH2 primers were not included in Ion AmpliSeq™ cancer panel v.1,
but in Ion AmpliSeq™ Cancer Hotspot Panel v2, and Comprehensive
cancer primer panels IDH2 gene was included (34–37). The
samples screened with Ion AmpliSeq™ cancer panel v.1 primers are
marked as ‘ND’ in the Tables for the IDH2 gene. The NGS method
detected mutations with a high accuracy, as evidenced by the
P-values (0.00001) and the high Phred quality score of all
variants, indicating high confidence in the variants found in this
tumor. Many of the IDH mutations found in this study had low
frequencies, which indicates they are somatic variants; however,
they all had a Phred score of ~100 and high suggest they are true
variants. All mutations were verified in various databases (COSMIC,
ExAc and dbSNP) to confirm whether the variants were novel in the
IDH1 and IDH2 genes.
Molecular genetic testing for the detection of
mutations as cancer markers have not been established in the
diagnostics laboratories of Saudi Arabia. Considering the lack of
molecular services to test cancer markers in the Kingdom, EGFRvIII
RT-qPCR tests in brain tumors were developed. This is the first
report from Saudi Arabian laboratories analyzing gliomas for
EGFRvIII expression, and the second report in a Saudi Arabian
population showing the prevalence of IDH mutations in gliomas. It
is also the first study from Saudi Arabia to analyze IDH mutations
in glioma cases using the capillary and NGS methods (46). In certain circumstances, the
differential diagnosis of CNS lesions based on histology alone can
be difficult. For example, distinguishing between reactive gliosis
and diffuse glioma, or between oligodendroglioma and other similar
entities, can be challenging. In some of these cases, the IDH
status was proven to be an extremely useful diagnostic biomarker,
and immunochemical detection methods are not very sensitive and
accurate compared with molecular techniques, such as Sanger
sequencing, a gold standard in molecular pathology for mutation
detection (63,64).
The aim of the present study was to determine the
prevalence of IDH1 and IDH2 mutations and EGFRvIII transcript
expression in Saudi Arabian patients with glioma. As these methods
are not available in this region, the aim of the present study was
to transfer these tests to specialty hospitals for patient's use.
This type of research will help transfer the clinical assays to the
diagnostics laboratories in the Kingdom, so that this approach can
be utilized to become independent in molecular diagnostics and
established oncology testing for clinical use. Finally, one
limitation that needs to be acknowledged in the present study is
the lack of functional studies. Retrospective, comparative survival
analysis correlating with glioma characteristics and types, surgery
type, the specifics of radiotherapy and the adjuvant therapies were
not performed in the present study. In the International Cancer
Genome Consortium (https://dcc.icgc.org) data portal IDH1 and EGFR genes
are in top 20 mutated cancer genes with high functional impact
somatic mutations. IDH1 and EGFR are most recurrently mutated
cancer driver genes in GBM_TCGA dataset (https://www.intogen.org). Poor prognostic markers
included genetic changes in the EGFR mutations in this group
(65), and among most driver genes,
IDH mutations are good prognostic factor in diffuse gliomas
(66,67).
In conclusion, the present study analyzed the IDH
mutations by NGS and capillary methods. The p. (Arg132His) mutation
in IDH1 was the predominant mutation in the Saudi Arabian
population. Several novel IDH1 and IDH2 mutations were reported in
this study. The novel IDH1 missense mutation in c.709T>C, a
novel synonymous mutation c.369 A>G, and a novel intronic
mutation (c.414+9T>A) was not reported previously. Also, in IDH2
gene four missense mutations, six synonymous variants, and an
intronic variant were novel variants found. In the present study,
missense mutations in IDH2 amino acid codons 172 and 140 were not
detected. The exon 4 mutation in IDH2 (c.472C>T) was not
reported previously in CNS tumors. In this study, 35% brain tumors
were positive for EGFRvIII expression. In the GBM category, 51%
were positive, and in astrocytoma and in anaplastic
oligodendroglioma 23 and 31% were positive, respectively. The
RT-qPCR test was validated by Sanger sequencing in
EGFRvIII-positive samples.
Development of molecular testing helps the clinician
to prescribe the correct medication, as this approach is giving
good results to treat different cancer types effectively, saves
money, avoids purchasing the diagnostic kits, and this will prevent
sending the samples to companies outside the Kingdom. The
development and application of a genetic test for brain tumors,
such as IDH1 and IDH2 mutations, is significant in this regard
because these tests have prognostic and therapeutic values.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank Mrs. Rowa Abbas
Bakhsh and Mr. Mohammed Bader Al-Hamad (Histopathology Division,
Al-Noor Specialty Hospital, Makkah, Saudi Arabia) for their
technical help. The authors would also like to thank Dr Emad A.
Felemban and Dr Ahmed Shawky (Science and Technology Unit, Umm
Al-Qura University, Makkah, Saudi Arabia) for their continuous
support.
Funding
The present study was supported by the National Plan
for Science, Technology and Innovation (MAARIFAH), King Abdul Aziz
City for Science and Technology (grant no. 12-MED 2961-10), to Dr
MM. Taher.
Availability of data and materials
The IDH2 novel variant [(c.472C>T; p.
(Pro158Ser)] sequence file was deposited in SRA (Sequence Read
Archive) database with access numbers SRA: PRJNA644191; BioSample
access number: SAMN15452876.
Authors' contributions
KV, FAA, and MMT conceived and designed the study.
RAJ, HK and EMB performed the pathologic diagnoses. MMT, GD, NMB
and MA performed the mutational analyses and acquired and
interpreted the data. KQ, EMB and THN drafted the manuscript and
analyzed the clinical data. KV, MMT, FAA and NMB revised it
critically and gave final approval of the version to be published.
All authors read and approved the final version of the
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Institutional
review board bioethics committee of King Abdullah Medical City,
Makkah, Kingdom of Saudi Arabia (approval no. 14-140). Written
informed consent was obtained from all patients or the parent or
guardian, if the patient was a minor.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
AML
|
acute myeloid leukemia
|
|
ATRT
|
atypical teratoid rhabdoid tumor
|
|
ClinVar
|
clinical variant
|
|
CNS
|
central nervous system
|
|
COSMIC
|
Catalogue Of Somatic Mutations In
Cancer
|
|
Ct
|
cycle threshold
|
|
ddPCR
|
droplet digital PCR
|
|
EGFRvIII
|
epidermal growth factor receptor
variant III
|
|
ExAc
|
The Exome Aggregation Consortium
|
|
FDA
|
The Food and Drug Administration
|
|
HG
|
hydroxyglutarate
|
|
HIF
|
hypoxia-inducible factor
|
|
IDH
|
isocitrate dehydrogenase
|
|
ISP
|
Ion sphere particle
|
|
KAMC
|
King Abdullah Medical City
|
|
KG
|
ketoglutarate
|
|
KSA
|
Kingdom of Saudi Arabia
|
|
MGMT
|
O(6)-methyl guanine methyl
transferase
|
|
NGS
|
next-generation DNA sequencing
|
|
PNET
|
primitive neuroectodermal tumor
|
|
RT-qPCR
|
reverse transcription-quantitative
PCR
|
|
TCGA
|
The Cancer Genome Atlas
|
|
TKI
|
tyrosine kinase inhibitor
|
References
|
1
|
Claussnitzer M, Cho JH, Collins R, Cox NJ,
Dermitzakis ET, Hurles ME, Kathiresan S, Kenny EE, Lindgren CM,
MacArthur DG, et al: A brief history of human disease genetics.
Nature. 577:179–189. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sobel ME, Bagg A, Caliendo AM, Ladanyi M
and Zehnbauer B: The evolution of molecular genetic pathology:
Advancing 20th-Century diagnostic methods into potent tools for the
new millennium. J Mol Diagn. 10:480–483. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Shendure J, Findlay GM and Snyder MW:
Genomic medicine-progress, pitfalls, and promise. Cell. 177:45–57.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Weller M, Wick W, Aldape K, Brada M,
Berger M, Pfister SM, Nishikawa R, Rosenthal M, Wen PY, Stupp R and
Reifenberger G: Glioma. Nat Rev Dis Primers. 1:150172015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ohgaki H, Dessen P, Jourde B, Horstmann S,
Nishikawa T, Di Patre PL, Burkhard C, Schüler D, Probst-Hensch NM,
Maiorka PC, et al: Genetic pathways to glioblastoma: A
population-based study. Cancer Res. 64:6892–6899. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Adamson C, Kanu OO, Mehta AI, Di C, Lin N,
Mattox AK and Bigner DD: Glioblastoma multiforme: A review of where
we have been and where we are going. Expert Opin Investig Drugs.
18:1061–1083. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Delgado-Lopez PD and Corrales-Garcıa EM:
Survival in glioblastoma: A review on the impact of treatment
modalities. Clin Transl Oncol. 18:1062–1071. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ostrom QT, Cioffi G, Gittleman H, Patil N,
Waite K, Kruchko C and Barnholtz-Sloan JS: CBTRUS statistical
report: Primary brain and other central nervous system tumors
diagnosed in the United States in 2012–2016. Neuro Oncol. 21 (Suppl
5):v1–v100. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Balss J, Meyer J, Mueller W, Korshunov A,
Hartmann C and von Deimling A: Analysis of the IDH1 codon 132
mutation in brain tumors. Acta Neuropathol. 116:597–602. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yan H, Parsons DW, Jin G, McLendon R,
Rasheed BA, Yuan W, Kos I, Batinic-Haberle I, Jones S, Riggins GJ,
et al: IDH1 and IDH2 mutations in gliomas. N Engl J Med.
360:765–773. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Christensen BC, Smith AA, Zheng S,
Koestler DC, Houseman EA, Marsit CJ, Wiemels JL, Nelson HH, Karagas
MR, Wrensch MR, et al: DNA methylation, isocitrate dehydrogenase
mutation, and survival in glioma. J Natl Cancer Inst. 103:143–153.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
van den Bent MJ, Dubbink HJ, Marie Y,
Brandes AA, Taphoorn MJ, Wesseling P, Frenay M, Tijssen CC, Lacombe
D, Idbaih A, et al: IDH1 and IDH2 mutations are prognostic but not
predictive for outcome in anaplastic oligodendroglial tumors: A
report of the European organization for research and treatment of
cancer brain tumor group. Clin Cancer Res. 16:1597–1604. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hanif F, Muzaffar K, Perveen K, Malhi SM
and Simjee ShU: Glioblastoma multiforme: A review of its
epidemiology and pathogenesis through clinical presentation and
treatment. Asian Pac J Cancer Prev. 18:3–9. 2017.PubMed/NCBI
|
|
14
|
Verhaak RG, Hoadley KA, Purdom E, Wang V,
Qi Y, Wilkerson MD, Miller CR, Ding L, Golub T, Mesirov JP, et al:
L Integrated genomic analysis identifies clinically relevant
subtypes of glioblastoma characterized by abnormalities in PDGFRA,
IDH1, EGFR, and NF1. Cancer Cell. 17:98–110. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Parsons DW, Jones S, Zhang X, Lin JC,
Leary RJ, Angenendt P, Mankoo P, Carter H, Siu IM, Gallia GL, et
al: An integrated genomic analysis of human glioblastoma
multiforme. Science. 321:1807–1812. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Frederick L, Wang XY, Eley G and James CD:
Diversity and frequency of epidermal growth factor receptor
mutations in human glioblastomas. Cancer Res. 60:1383–1387.
2000.PubMed/NCBI
|
|
17
|
Herbst RS and Shin DM: Monoclonal
antibodies to target epidermal growth factor receptor-positive
tumors. A new paradigm for cancer therapy. Cancer. 94:1593–1611.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pelloski CE, Ballman KV, Furth AF, Zhang
L, Lin E, Sulman EP, Bhat K, McDonald JM, Yung WK, Colman H, et al:
Epidermal growth factor receptor variant III status defines
clinically distinct subtypes of glioblastoma. J Clin Oncol.
25:2288–2294. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wong AJ, Ruppert JM, Bigner SH, Grzeschik
CH, Humphrey PA, Bigner DS and Vogelstein B: Structural alterations
of the epidermal growth factor receptor gene in human gliomas. Proc
Natl Acad Sci USA. 89:2965–2969. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Nishikawa R, Ji XD, Harmon RC, Lazar CS,
Gill GN, Cavenee WK and Huang HJ: A mutant epidermal growth factor
receptor common in human glioma confers enhanced tumorigenicity.
Proc Natl Acad Sci USA. 91:7727–7731. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Geisbrecht BV and Gould SJ: The human PICD
gene encodes a cytoplasmic and peroxisomal NADP (+)-dependent
isocitrate dehydrogenase. J Biol Chem. 274:30527–30533. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cairns RA and Mak TW: Oncogenic isocitrate
dehydrogenase mutations: Mechanisms models, and clinical
opportunities. Cancer Discov. 3:730–741. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dang L, White DW, Gross S, Bennett BD,
Bittinger MA, Driggers EM, Fantin VR, Jang HG, Jin S, Keenan MC, et
al: Cancer-associated IDH1 mutations produce 2-hydroxyglutarate.
Nature. 462:739–744. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhao S, Lin Y, Xu W, Jiang W, Zha Z, Wang
P, Yu W, Li Z, Gong L, Peng Y, et al: Glioma-derived mutations in
IDH1 dominantly inhibit IDH1 catalytic activity and induce
HIF-1alpha. Science. 324:261–265. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Choi BD, Maus MV, June CH and Sampson JH:
Immunotherapy for glioblastoma: Adoptive T-cell strategies. Clin
Cancer Res. 25:2042–2048. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
An Z, Aksoy O, Zheng T, Fan QW and Weiss
WA: Epidermal growth factor receptor and EGFRvIII in glioblastoma:
Signaling pathways and targeted therapies. Oncogene. 37:1561–1575.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Louis DN, Perry A, Reifenberger G, von
Deimling A, Figarella-Branger D, Cavenee WK, Ohgaki H, Wiestler OD,
Kleihues P and Ellison DW: The 2016 world health organization
classification of tumors of the central nervous system: A summary.
Acta Neuropathol. 131:803–820. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Steffensen KD, Waldstrøm M, Olsen DA,
Corydon T, Lorentzen KA, Knudsen HJ, Jeppesen U, Brandslund I and
Jakobsen A: Mutant epidermal growth factor receptor in benign,
borderline, and malignant ovarian tumors. Clin Cancer Res.
14:3278–3282. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yoshimoto K, Dang J, Zhu S, Nathanson D,
Huang T, Dumont R, Seligson DB, Yong WH, Xiong Z, Rao N, et al:
Development of a real-time RT-PCR assay for detecting EGFRvIII in
glioblastoma samples. Clin Cancer Res. 14:488–493. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Golding SE, Rosenberg E, Khalil A, McEwen
A, Holmes M, Neill S, Povirk LF and Valerie K: Double strand break
repair by homologous recombination is regulated by cell
cycle-independent signaling via ATM in human glioma cells. J Biol
Chem. 279:15402–15410. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Golding SE, Morgan RN, Adams BR, Hawkins
AJ, Povirk LF and Valerie K: Pro-survival AKT and ERK signaling
from EGFR and mutant EGFRvIII enhances DNA double-strand break
repair in human glioma cells. Cancer Biol Ther. 8:730–738. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lammering G, Hewit TH, Holmes M, Valerie
K, Hawkins W, Lin PS, Mikkelsen RB and Schmidt-Ullrich RK:
Inhibition of the type III epidermal growth factor receptor variant
mutant receptor by dominant-negative EGFR-CD533 enhances malignant
glioma cell radiosensitivity. Clin Cancer Res. 10:6732–6743. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Patel KP, Barkoh BA, Chen Z, Ma D, Reddy
N, Medeiros LJ and Luthra R: Diagnostic testing for IDH1 and IDH2
variants in acute myeloid leukemia an algorithmic approach using
high-resolution melting curve analysis. J Mol Diagn. 13:678–686.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mehrotra M, Duose DY, Singh RR, Barkoh BA,
Manekia J, Harmon MA, Patel KP, Routbort MJ, Medeiros LJ, Wistuba
II and Luthra R: Versatile ion S5XL sequencer for targeted next
generation sequencing of solid tumors in a clinical laboratory.
PLoS One. 12:e01819682017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ballester LY, Fuller GN, Powell SZ, Sulman
EP, Patel KP, Luthra R and Routbort MJ: Retrospective analysis of
molecular and immunohistochemical characterization of 381 primary
brain tumors. J Neuropathol Exp Neurol. 76:179–188. 2017.PubMed/NCBI
|
|
36
|
Taher MM, Hassan AA, Saeed M, Jastania RA,
Nageeti TH, Alkhalidi H, Dairi G, Abduljaleel Z, Athar M,
Bouazzaoui A, et al: Next generation DNA sequencing of atypical
choroid plexus papilloma of brain: Identification of novel
mutations in a female patient by Ion Proton. Oncol Lett.
18:5063–5076. 2019.PubMed/NCBI
|
|
37
|
Butt M, Alyami S, Nageeti T, Saeed M,
AlQuthami K, Bouazzaoui A, Athar M, Abduljaleel Z, Al-Allaf F and
Taher M: Mutation profiling of anaplastic ependymoma grade III by
Ion Proton by next generation DNA sequencing. F1000Res. 8:6132019.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sanson M, Marie Y, Paris S, Idbaih A,
Laffaire J, Ducray F, El Hallani S, Boisselier B, Mokhtari K,
Hoang-Xuan K and Delattre JY: Isocitrate dehydrogenase 1 codon 132
mutation is an important prognostic biomarker in gliomas. J Clin
Oncol. 27:4150–4154. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lu Y, Kwintkiewicz J, Liu Y, Tech K, Frady
LN, Su YT, Bautista W, Moon SI, MacDonald J, Ewend MG, et al:
Chemosensitivity of IDH1-mutated gliomas due to an impairment in
PARP1-mediated DNA repair. Cancer Res. 77:1709–1718. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Reitman ZJ and Yan H: Isocitrate
dehydrogenase 1 and 2 mutations in cancer: Alterations at a
crossroads of cellular metabolism. J Natl Cancer Inst. 102:932–941.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hartmann C, Meyer J, Balss J, Capper D,
Mueller W, Christians A, Felsberg J, Wolter M, Mawrin C, Wick W, et
al: Type and frequency of IDH1 and IDH2 mutations are related to
astrocytic and oligodendroglial differentiation and age: A study of
1,010 diffuse gliomas. Acta Neuropathol. 118:469–474. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Hayden JT, Frühwald MC, Hasselblatt M,
Ellison DW, Bailey S and Clifford SC: Frequent IDH1 mutations in
supratentorial primitive neuroectodermal tumors (sPNET) of adults
but not children. Cell Cycle. 8:1806–1807. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Mukasa A, Takayanagi S, Saito K, Shibahara
J, Tabei Y, Furuya K, Ide T, Narita Y, Nishikawa R, Ueki K and
Saito N: Significance of IDH mutations varies with tumor histology,
grade, and genetics in Japanese glioma patients. Cancer Sci.
103:587–592. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Mellai M, Piazzi A, Caldera V, Monzeglio
O, Cassoni P, Valente G and Schiffer D: IDH1 and IDH2 mutations,
immunohistochemistry and associations in a series of brain tumors.
J Neurooncol. 105:345–357. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Yang H, Ye D, Guan KL and Xiong Y: IDH1
and IDH2 mutations in tumorigenesis: Mechanistic insights and
clinical perspectives. Clin Cancer Res. 18:5562–5571. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Alassiri AH, Alkhaibary A, Al-Sarheed S,
Alsufani F, Alharbi M, Alkhani A and Aloraidi A:
O6-methylguanine-DNA methyltransferase promoter
methylation and isocitrate dehydrogenase mutation as prognostic
factors in a cohort of Saudi patients with glioblastoma. Ann Saudi
Med. 39:410–416. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wang J, Zhao Y, Li JF, Guo CC, Chen FR, Su
HK, Zhao HF, Long YK, Shao JY, To Ss and Chen ZP: IDH1 mutation
detection by droplet digital PCR in glioma. Oncotarget.
6:39651–39660. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Shin H, Sa JK, Bae JS, Koo H, Jin S, Cho
HJ, Choi SW, Kyoung JM, Kim JY, Seo YJ, et al: Clinical targeted
next-generation sequencing panels for detection of somatic variants
in gliomas. Cancer Res Treat. 52:41–50. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Ali MAM, Ahmed EK, Assem MMA and Helwa R:
The synonymous isocitrate dehydrogenase 1 315C>T SNP confers an
adverse prognosis in egyptian adult patients with
NPM1-/CEBPA-negative acute myeloid leukemia. Indian J Hematol Blood
Transfus. 34:240–252. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wang HY, Tang K, Liang TY, Zhang WZ, Li
JY, Wang W, Hu HM, Li MY, Wang HQ, He XZ, et al: The comparison of
clinical and biological characteristics between IDH1 and IDH2
mutations in gliomas. J Exp Clin Cancer Res. 35:862016. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Wartenberg M, Cibin S, Zlobec I, Vassella
E, Eppenberger-Castori S, Terracciano L, Eichmann MD, Worni M,
Gloor B, Perren A and Karamitopoulou E: Integrated Genomic and
Immunophenotypic classification of pancreatic cancer reveals three
distinct subtypes with prognostic/predictive Significance. Clin
Cancer Res. 24:4444–4454. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Watanabe T, Nobusawa S, Kleihues P and
Ohgaki H: IDH1 mutations are early events in the development of
astrocytomas and oligodendrogliomas. Am J Pathol. 174:1149–1153.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Nobusawa S, Watanabe T, Kleihues P and
Ohgaki H: IDH1 mutations as molecular signature and predictive
factor of secondary glioblastomas. Clin Cancer Res. 15:6002–6007.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Wikstrand CJ, McLendon RE, Friedman AH and
Bigner DD: Cell surface localization and density of the
tumor-associated variant of the epidermal growth factor receptor,
EGFRvIII. Cancer Res. 57:4130–4140. 1997.PubMed/NCBI
|
|
55
|
Johnson LA, Scholler J, Ohkuri T, Kosaka
A, Patel PR, McGettigan SE, Nace AK, Dentchev T, Thekkat P, Loew A,
et al: Rational development and characterization of humanized
anti-EGFR variant III chimeric antigen receptor T cells for
glioblastoma. Sci Transl Med. 7:275ra222015. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Heimberger AB, Sun W, Hussain SF, Dey M,
Crutcher L, Aldape K, Gilbert M, Hassenbusch SJ, Sawaya R,
Schmittling B, et al: Immunological responses in a patient with
glioblastoma multiforme treated with sequential courses of
temozolomide and immunotherapy: Case study. Neuro Oncol. 10:98–103.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Binder DC, Ladomersky E, Lenzen A, Zhai L,
Lauing KL, Otto-Meyer SD, Lukas RV and Wainwright DA: Lessons
learned from rindopepimut treatment in patients with
EGFRvIII-expressing glioblastoma. Transl Cancer Res. 7 (Suppl
4):S510–S513. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Kwatra MM: A rational approach to target
the epidermal growth factor receptor in glioblastoma. Curr Cancer
Drug Targets. 17:290–296. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Kastenhuber ER, Huse JT, Berman SH,
Pedraza A, Zhang J, Suehara Y, Viale A, Cavatore M, Heguy A,
Szerlip N, et al: Quantitative assessment of intragenic receptor
tyrosine kinase deletions in primary glioblastomas: Their
prevalence and molecular correlates. Acta Neuropathol. 127:747–759.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Felsberg J, Hentschel B, Kaulich K,
Gramatzki D, Zacher A, Malzkorn B, Kamp M, Sabel M, Simon M,
Westphal M, et al: Epidermal growth factor receptor variant III
(EGFRvIII) positivity in EGFR-amplified glioblastomas: Prognostic
role and comparison between primary and recurrent tumors. Clin
Cancer Res. 23:6846–6855. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Carvalho PO, Uno M, Oba-Shinjo SM,
Rosemberg S, Wakamatsu A, da Silva CC, Teixeira MJ and Marie SK:
Activation of EGFR signaling from pilocytic astrocytomas to
glioblastomas. Int J Biol Markers. 29:e120–e128. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Wong A, Mitra S, Del Vecchio CA and
Skirboll S: Expression of EGFRvIII in brain tumor stem cells. J
Clin Oncol. 26 (Suppl 15):S2002. 2008. View Article : Google Scholar
|
|
63
|
Katsanis SH and Katsanis N: Molecular
genetic testing and the future of clinical genomics. Nat Rev Genet.
14:415–426. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Di Resta C, Galbiati S, Carrera P and
Ferrari M: Next-generation sequencing approach for the diagnosis of
human diseases: Open challenges and new opportunities. EJIFCC.
29:4–14. 2018.PubMed/NCBI
|
|
65
|
Liang A, Zhou B and Sun W: Integrated
genomic characterization of cancer genes in glioma. Cancer Cell
Int. 17:902017. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Jonsson P, Lin AL, Young RJ, DiStefano NM,
Hyman DM, Li BT, Berger MF, Zehir A, Ladanyi M, Solit DB, et al:
Genomic correlates of disease progression and treatment response in
prospectively characterized gliomas. Clin Cancer Res. 25:5537–5547.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Ceccarelli M, Barthel FP, Malta TM,
Sabedot TS, Salama SR, Murray BA, Morozova O, Newton Y, Radenbaugh
A, Pagnotta SM, et al: Molecular profiling reveals biologically
discrete subsets and pathways of progression in diffuse glioma.
Cell. 164:550–563. 2016. View Article : Google Scholar : PubMed/NCBI
|