Introduction
Glioma is a type of primary brain cancer, which is
characterized by high prevalence and gross invasion and is lethal
to human health (1,2). It is estimated that >19,000 new
cases of glioma are diagnosed each year, with an age-adjusted
average annual incidence rate of 6.24 per 100,000 individuals in
the United States between 2007 and 2011 (3). Glioma cells are aggressive and can
develop resistance to conventional therapies; treatment options
(such as surgery, radiotherapy and chemotherapy) exhibit poor
efficiency and the average survival time of patients with
high-grade glioma is <1 year (4).
With the development of molecular biology, the potential of
immunotherapy as a treatment for glioma has been investigated. It
is necessary to understand immunotolerance and immunomodulation of
glioma to develop an immunotherapy strategy. Toll-like receptors
(TLRs) have been identified as key pattern recognition receptors
(PRRs). The release of inflammatory mediators and the activation of
innate immunity are primarily caused by TLRs, which induce signal
transduction via recognition of pathogen-associated molecular
patterns (PAMPs) and certain endogenous ligands (5,6). TLRs
are also implicated in the pathogenesis of numerous types of
immune-associated disease in human (7–9). For
example, Tian et al (10)
reported that mulberry leaf decreased inflammation and insulin
resistance in type 2 diabetic mice via TLRs and the insulin
signaling pathway. A total of 10 types of TLR have been identified
in humans, among which TLR4 exhibits a significant association with
the biological behavior of tumors (11,12).
This finding is of interest from a tumor immunity perspective.
Although TLRs mediate innate immunity, they are not unique to the
innate immune system; in addition to immune cells
(monocytes/macrophages, T/B lymphocytes and dendritic cells), they
are also expressed in certain non-immune cells (endothelial, smooth
muscle and tumor cells) (13). TLR
expression levels in tumor cells are important for tumor genesis
and development, and is an area of interest for tumor research
(14). Among TLRs, TLR4 is more
commonly expressed in different types of tumor, such as colon
cancer and ovarian carcinoma, compared with other TLRs (15,16).
High expression levels of TLR4 accelerate tumor proliferation and
mediate tumor immune escape (17,18).
Hence, the present study aimed to investigate the association
between TLR4 and the development of human glioma, and to provide an
experimental basis for understanding of the molecular mechanism
underlying proliferation regulation of human glioma cells.
Materials and methods
Reagents and equipment
All-in-One™ quantitative (q)PCR Primer (cat. no.
HQP054754) and H-TLR4-short hairpin (sh)RNAs 1–3 (cat. nos.
HSH054754-CU6-a, b, c and CSHCTR001-CU6) were purchased from
GeneCopoeia, Inc. Lipofectamine® 2000 was obtained from
Invitrogen (Thermo Fisher Scientific, Inc.). High-purity Plasmid
Mini-Preps kit was obtained from Tiangen Bioctech Co., Ltd..
Minimum Essential Medium (MEM) and bovine serum albumin were
obtained from Gibco (Thermo Fisher Scientific, Inc.). RNAiso total
RNA extraction reagent, HiScript II 1st Strand complementary cDNA
Synthesis kit and qPCR SYBR Green Master mix were acquired from
Vazyme Biotech Co., Ltd.. Annexin V-PE/7-AAD kit and Matrigel
Matrix Basement Membrane were purchased from BD Biosciences. FC-500
flow cytometer (Beckman Coulter, Inc.), the PCR instrument
(Eppendorf) and Mx3000P fluorescence quantitative PCR instrument
(Agilent Technologies, Inc.) were also used.
Cell lines
The U-87MG cell line (cat. no. CL-0238) was obtained
from Procell Life Science & Technology Co., Ltd. STR profiling,
Y-chromosome paint and Q-band assay confirmed that the cell line is
male in origin (performed by Procell Life Science & Technology
Co., Ltd). Based on current literature, the cell line is likely a
glioblastoma of unknown origin (19). The U-87MG cell line had a stable
passage in the laboratory, and was cultured in MEM containing 10%
FBS, 100 U/ml penicillin and streptomycin. Cells were sustained in
an incubator at 37°C with 5% CO2.
RNA interference lentiviral vector
construction
GeneCopoeia, Inc. synthesized the interference
plasmids [cat. nos. HSH054754-CU6-a,b,c and CSHCTR001-CU6;
accession no. NM_003266.3 (Homo sapiens TLR4, mRNA)]. The
vector used was psi-U6.1 (GeneCopoeia, Inc.). A total of three
shRNAs were designed to interfere with the TLR4 gene. shRNA target
sequences are presented in Table
I.
 | Table I.Short hairpin RNA target
sequences. |
Table I.
Short hairpin RNA target
sequences.
| Clone name | Symbol | Location | Length, bp | Target sequence
(5′-3′) |
|---|
|
HSH054754-CU6-a(OS397039) | TLR4 | 890 | 21 |
GCTCACAATCTTATCCAATCT |
|
HSH054754-CU6-b(OS397040) | TLR4 | 678 | 21 |
GGTGTGAAATCCAGACAATTG |
|
HSH054754-CU6-c(OS397041) | TLR4 | 887 | 21 |
GTGGCTCACAATCTTATCCAA |
Transfection of shRNA-containing
plasmids into U-87MG cells
shRNA-containing plasmids (4 µg) were transfected
into U-87MG cells, in order to silence the TLR4 gene. For reagent
1, 244 µl MEM (FBS- and antibiotic-free) was used to buffer 6 µl
Lipofectamine® 2000. For reagent 2, 240 µl MEM (FBS- and
antibiotics-free) was used to dilute 10 µl shRNA plasmids
[containing 4 µg TLR4-sh1-3 and negative control (NC) plasmids].
Both reagents were placed at room temperature for 5 min.
U-87MG cells in the logarithmic phase of growth were
collected and seeded in 6-well plates at a density of
4×105 cells/well. Cells were transfected until they
reached 85–90% confluence. MEM (FBS- and antibiotics-free) was used
to wash cells twice, then 2 ml FBS- and antibiotic-free MEM was
added into each well. After placing the mixture containing reagents
1 and 2 at room temperature for 20 min, 200 µl of this mixture was
added into wells. After 6 h, FBS- and antibiotics-free MEM was
replaced with complete culture medium. Following transfection for
48 h, cells were observed under a fluorescence microscope
(magnification, ×100). Untreated cells were considered as the blank
control group. Cells only treated with Lipofectamine were
considered as the liposome group.
Identifying stably transfected cells
with puromycin
The TLR4-sh2 plasmid was chosen for subsequent siRNA
experiments. After U-87MG cells were transfected with the TLR4-sh2
plasmid and the negative control plasmid for 48 h, the cells were
diluted with MEM and cultured in a 6-well plate at a density of
5×104 cells/well. Subsequently, 600 ng/ml puromycin was
added into each well. The non-transfected U-87MG cells were used as
the blank control group. After the cells were cultured for 14 days
at 37°C with 5% CO2, the surviving cells were considered
as stably transfected cells. The cells transfected with TLR4-sh2
were named U-87MG-Sh, while the cells transfected with the negative
control (NC) plasmid were named U-87MG-NC.
Evaluating transfection efficiency via
flow cytometry
Following transfection for 48 h, cells were
collected and washed with PBS once. Next, flow cytometry was
performed using a FC-500 type flow cytometer (Beckman Coulter,
Inc.) to measure green fluorescence in cells. Transfection
efficiency was identified as the proportion of cells expressing
green fluorescence compared with all observed cells. Green
fluorescence was measured using the EXPO32 ADC v1.2 software
(Beckman Coulter, Inc.).
Evaluating TLR4 mRNA expression levels
via reverse transcription-quantitative (RT-q)PCR
Following transfection for 48 h, cells were
collected and washed with PBS twice. Next, based on the one-step
method (20), total RNA was
extracted by adding 1 ml RNA isolator (Vazyme Biotech Co., Ltd.).
As the template for PCR, cDNA was synthesized using the HiScript II
First Strand cDNA Synthesis kit according to the manufacturer's
protocol (Vazyme Biotech Co., Ltd.). The internal reference was
human GAPDH. The fluorescent signal was obtained using SYBR-Green
I. Based on the manufacturer's protocol of the qPCR SYBR-Green
Master Mix kit (Vazyme Biotech Co., Ltd.), qPCR was performed. The
thermocycling conditions were as follows: 95°C for 5 min
(pre-denaturation), followed by 40 cycles at 95°C for 10 sec and
60°C for 30 sec, and dissociation at 95°C for 15 sec, 60°C for 1
min and 95°C for 15 sec. For each sample, each experiment was
repeated three times. The forward primer for GAPDH was
3′-CACTACCGTACCTGACACCA-5′ and the reverse primer was
3′-ATGTCGTTGTCCCACCACCT-5′. The TLR4 sequence was synthesized by
GeneCopoeia, Inc. (cat. no. HQP054754). The relative expression
levels of TLR4 mRNA (ΔCq=CqTLR4-CqGAPDH) were
calculated using the 2−ΔΔCq method (21). RT-qPCR was used to evaluate the
silencing effect of shRNA TLR4 expression levels and to select the
interference plasmid with the greatest silencing effect.
Measuring TLR4 protein expression
levels via western blotting
U-87MG-Sh, U-87MG-NC and U-87MG cells were collected
in the logarithmic phase of growth and washed with cold PBS twice.
Total protein was collected by adding cell lysis solution
(RIPA:PMSF, 100:1; Beijing Solarbio Science & Technology Co.,
Ltd.). The protein concentration was evaluated via bicinchoninic
acid assay. Protein (50 µg/lane) was separated via 10% SDS-PAGE
then transferred to PVDF membranes before being blocked overnight
at 4°C with 5% skimmed milk. Next, rabbit-anti-human β-actin
(1:4,000; cat. no. ab8227; Abcam) or mouse-anti-human TLR4 (1:500;
cat. no. ab22048; Abcam) monoclonal antibodies were added for
incubation at room temperature for 1 h. The membrane was washed
three times, then incubated at room temperature for 2 h with
horseradish peroxidase-conjugated goat anti-mouse and anti-rabbit
secondary antibodies (1:20,000; cat. nos. ab205719 and ab205718,
respectively; Abcam). The antigen-antibody reaction was visualized
by detection using an ECL assay (EMD Millipore). Relative
expression levels of TLR4 protein were normalized to those of
β-actin using Odyssey v3.0 software (LI-OR Biosciences). A total of
three independent repeats were performed.
Flow cytometric analysis of cell
cycle
U-87MG-Sh, U-87MG-NC and U-87MG cells were collected
in the logarithmic phase of growth and washed with cold PBS twice.
Cells were fixed with 70% ethanol at 4°C for 24 h. Cells were
washed twice with PBS and cell density was adjusted to
1×107/ml. Then, 100 µl cell suspension was collected and
500 µl propidium iodide including a ribonuclease (BD Biosciences)
was added for 30 min at 4°C. Flow cytometry was performed using a
FC500 type flow cytometer (Beckman Coulter, Inc.), and the
Multicycle AV v6.0 software (Beckman Coulter, Inc.) was used to
estimate the percentage of cells in each stage of the cell cycle.
The proliferation index (PI) was calculated as the percentage of
cells in the S and G2/M phases against all cells,
representing cell proliferation activity.
Flow cytometry analysis of
apoptosis
U-87MG-Sh, U-87MG-NC and U-87MG cells were collected
in the logarithmic phase of growth and washed with cold PBS twice.
Then, 1×106 cells were collected and washed with PBS
once and suspended in 100 µl 1X binding buffer. Next, cells were
incubated on ice with 10 µl Annexin V-PE for 15 min. Then, 380 µl
1X binding buffer and 10 µl 7-AAD were separately added, before
being placed on ice for 15 min in the dark, washed and suspended in
1 ml PBS. Flow cytometry (FC500 type flow cytometer; Beckman
Coulter, Inc.) was used to evaluate cell apoptosis.
Immunofluorescence data and apoptosis rate were measured using the
EXPO32 ADC v1.2 software (Beckman Coulter, Inc.).
Measuring cell migration by wound
healing assay
On the back of the 6-well culture plate, lines
(5/well) at 0.5 cm intervals were drawn. Then, ~5×105
cells (U-87MG-Sh, U-87MG-NC and U-87MG) were seeded into each well
and cultivated overnight at 37°C to 100% confluence. The next day,
scratches perpendicular to the lines on the back of the 6-well
plate were drawn on cells using a 10-µl pipette tip. In order to
remove unattached cells, cells were washed with PBS three times and
cultured for 24 h at 37°C in serum-free MEM. The cell culture
condition was 37°C with 5% CO2, and images were captured
at 0 and 24 h using an inverted light microscope (TE2000; Nikon
Corporation; magnification, ×100). The area of wound healing was
assayed using ImageJ 1.42 software (National Institutes of
Health).
Detecting cell viability by cell
counting Kit-8 (CCK-8) assay
Monolayer cultured U-87MG-Sh, U-87MG-NC and U-87MG
cells were digested at 37°C for 30 sec using 0.25% trypsin. Then,
MEM supplemented with 10% FBS (Gibco; Thermo Fisher Scientific,
Inc.) was used to prepare the cell suspension. The cell density was
adjusted to 1×104 cells/ml and cells were seeded into
96-well plates before being cultured in the incubator for cell
adherence. Following incubation at 37°C with 5% CO2 for
24 h, 20 µl CCK-8 (Wuhan Boster Biological Technology, Ltd.) was
added to each well and the plate was incubated for 4 h at 37°C
according to the manufacturer's protocol. The 450 nm optical
density wavelength was read via a microplate spectrophotometer,
calibrated using the blank well. Growth inhibitory rate was
calculated as 1-A450 U-87MG-Sh, U-87MG-NC or U-87MG cells/A450
U-87MG cells) ×100%.
Statistical analysis
All data are presented as the mean ± SD (n=3).
One-way ANOVA was performed to compare multiple samples, followed
by Tukey's post hoc test. Data were analyzed using SPSS software
(version 21; IBM Corp.). P<0.05 was considered to indicate a
statistically significant difference.
Results
Silencing of TLR4 in U-87MG cells
using shRNA plasmid transfection and evaluation of transfection
efficiency and TLR4 mRNA expression levels via flow cytometry and
fluorescence RT-qPCR
Green fluorescence expression levels in U-87MG cells
transfected with H_TLR4-sh1, H_TLR4-sh2, H_TLR4-sh3 and NC plasmid
were observed via fluorescence microscope. No green fluorescence
was detected in non-transfected cells (blank control) and U-87MG
cells in the liposome group.
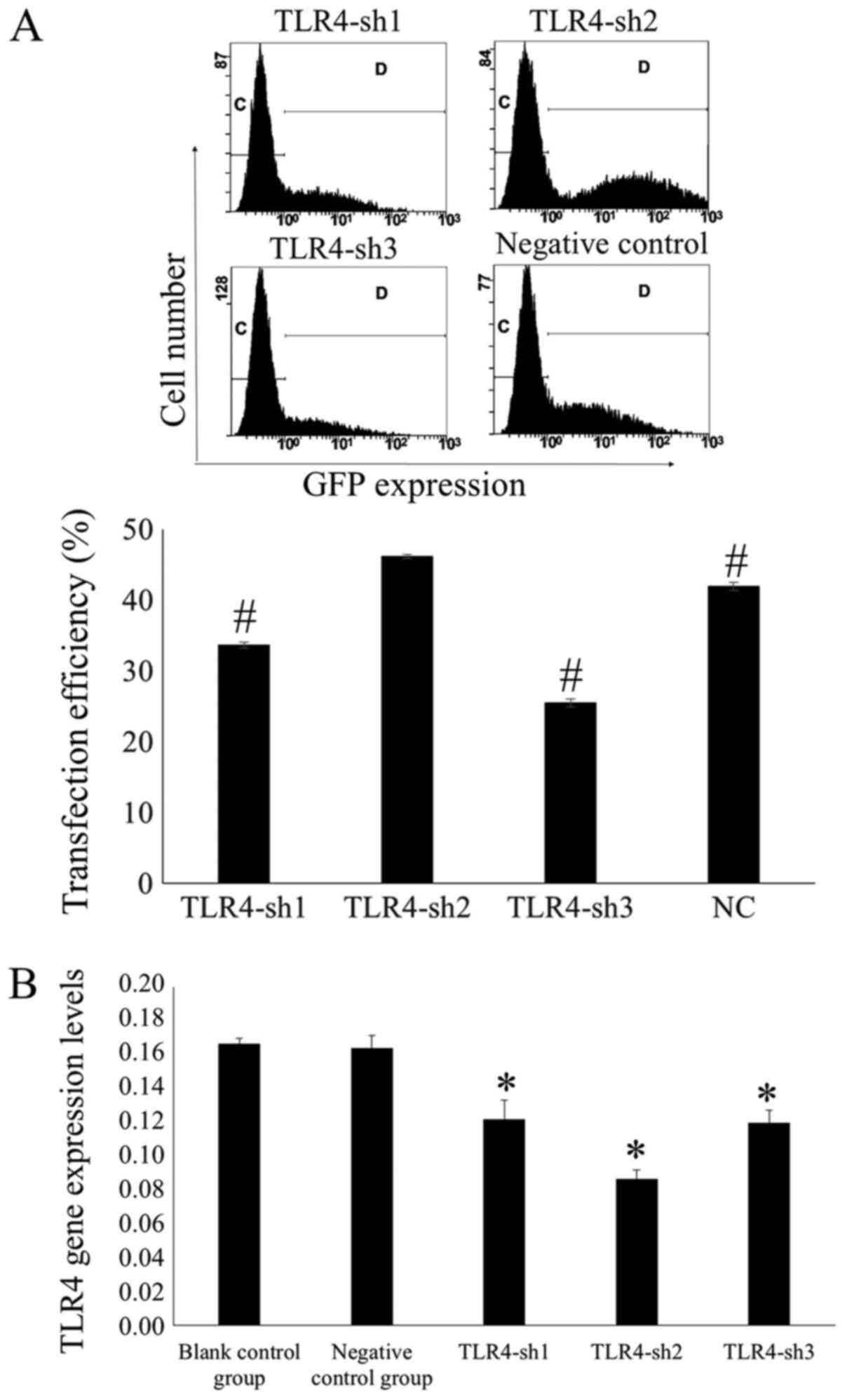
Flow cytometry was used to measure transfection
efficiency after 48 h. Results demonstrated that the transfection
efficiency was 33.66±0.40, 46.20±0.31, 25.49±0.60 and 41.94±0.55%
in the H_TLR4-sh1, H_TLR4-sh2, H_TLR4-sh3 and NC groups,
respectively. In the H_TLR4-sh2 group, the transfection efficiency
was significantly higher compared with the other groups (P<0.01;
Fig. 1A).
It was indicated that the TLR4 mRNA expression
levels in U-87MG cells transfected with H_TLR4-sh1, H_TLR4-sh2,
H_TLR4-sh3 and NC plasmid were 0.1219±0.0116, 0.0867±0.0054,
0.1201±0.0071 and 0.1643±0.0074, respectively, whereas that in the
blank control group was 0.1669±0.0029. The TLR4 mRNA expression
levels were significantly lower in U-87MG cells transfected with
H_TLR4-sh2 than in the other groups (P<0.05; Fig. 1B). This illustrated that H_TLR4-sh2
possessed the largest silencing effect. Hence, H_TLR4-sh2 was
selected for subsequent experiments.
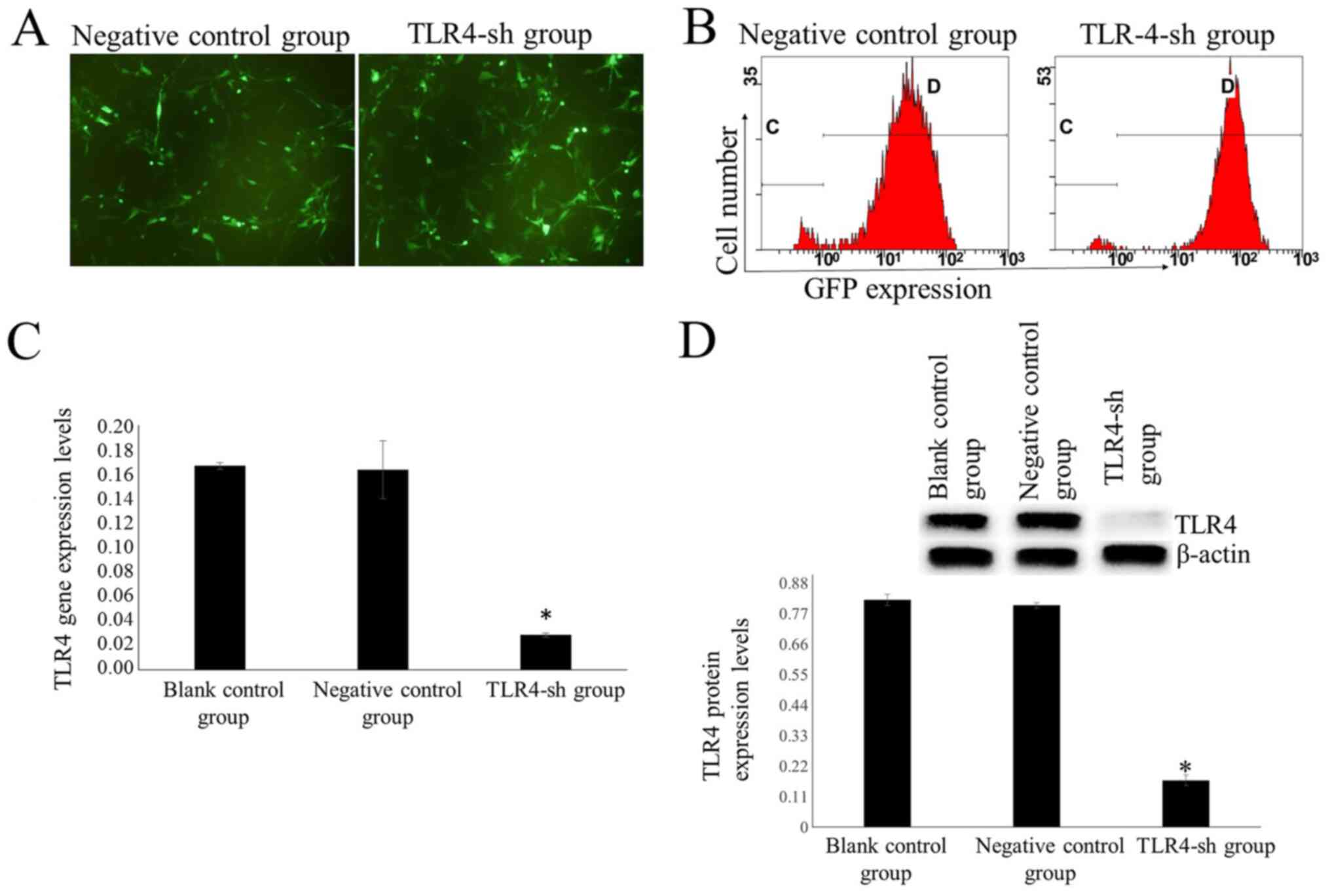
TLR4 silencing decreases TLR4 mRNA
expression levels
Following treatment with 600 ng/ml puromycin for 14
days, all cells in the blank control group died. A notable number
of cells transfected with TLR4-sh2 and NC plasmids were alive
(Fig. 2A). Surviving cells
represented stably transfected cells. Cells transfected with
TLR4-sh2 and NC plasmids were labeled as U-87MG-Sh and U-87MG-NC
cells, respectively.
Transfection efficiency was measured via flow
cytometry. In the TLR4-sh and NC group, transfection efficiency was
95.05±0.17 and 94.75±0.72%, respectively (Fig. 2B). Fluorescence RT-qPCR results
demonstrated that the TLR4 mRNA expression levels were
significantly lower in the TLR4-sh group than that in NC and blank
control groups (P<0.01; Fig.
2C).
TLR4 protein expression levels are
decreased in TLR4-sh-transfected cells
Western blot results demonstrated that TLR4 protein
expression levels were significantly lower in the TLR4-sh group
than in NC and blank control groups (P<0.01; Fig. 2D).
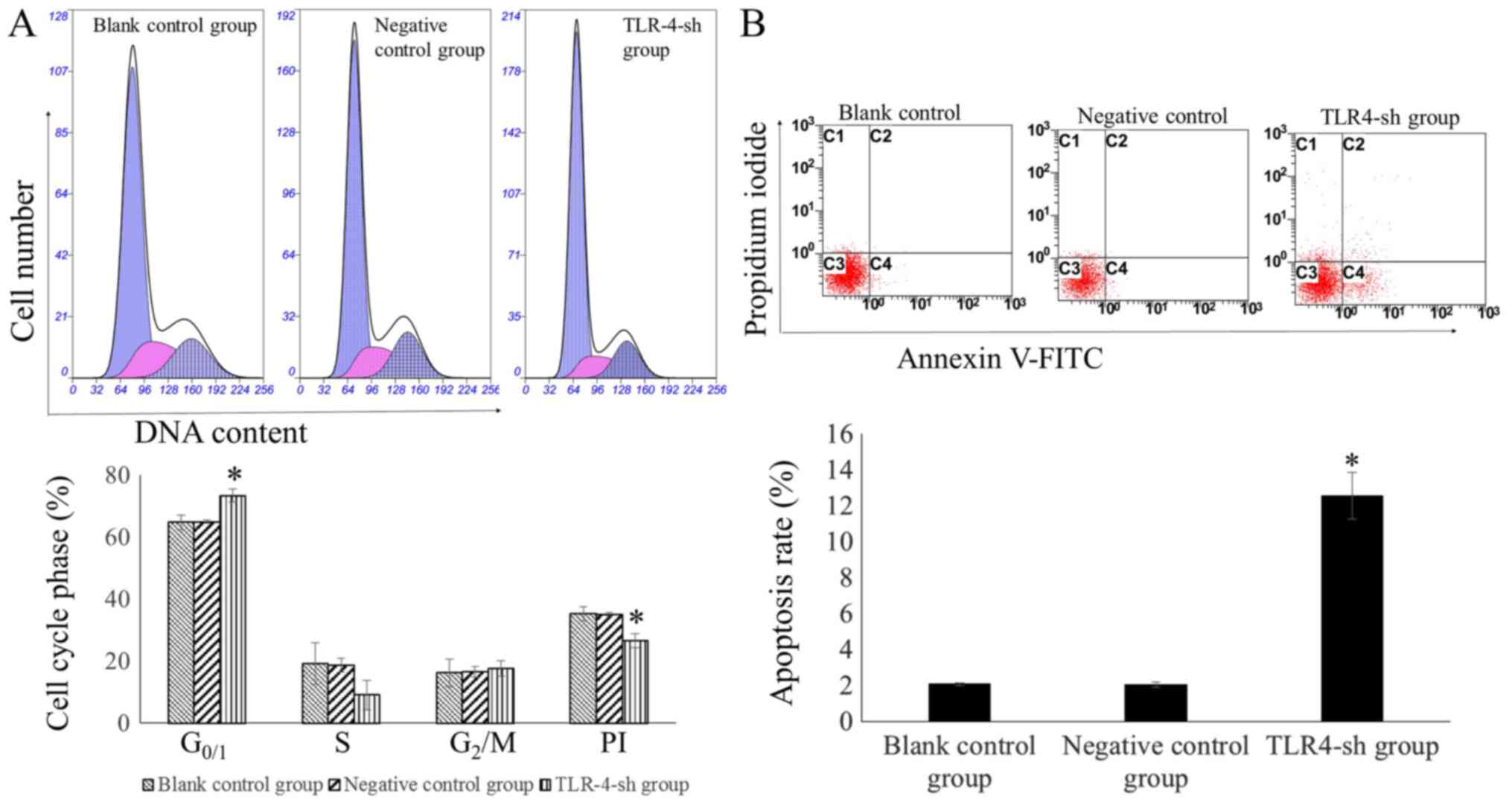
Measuring cell cycle and apoptosis
rate of transfected cells via flow cytometry
The percentage of cells in the G0/1 phase
was significantly higher in the TLR4-sh than that in NC and blank
control groups (P<0.01). By contrast, the PI was significantly
lower in U-87MG-Sh cells than in U-87MG-NC and U-87MG cells
(P<0.01). The percentage of the cells in the G0/1
phase and PI in U-87MG-NC and U-87MG cells exhibited no significant
difference (P>0.05; Fig. 3A).
The cell apoptosis rate was significantly higher in
U-87MG-Sh cells than in U-87MG-NC and U-87MG cells (P<0.01)
whereas no significant difference was observed in the apoptosis
rates between U-87MG-NC and U-87MG cells (P>0.05; Fig. 3B).
TLR4 silencing decreases migration
ability
Wound healing assay indicated that migration ability
of the U-87MG-Sh cells was significantly lower than that of
U-87MG-NC and U-87MG cells (Fig.
4).
TLR4 silencing inhibits cell
proliferation
Results of the CCK-8 assay indicated that the
proliferation inhibitory rates were 0.40±1.47, 0.31±0.94 and
28.94±1.31% in the blank, NC and TLR4-sh groups, respectively. The
proliferation inhibitory rate of TLR4-sh group was significantly
higher than that of blank and NC groups (P<0.01) (data not
shown).
Discussion
Glioma is a type of primary brain cancer (22), which is characterized by high
prevalence and gross invasion and is lethal to human health
(1,2). Resistance to conventional therapy (such
as surgery, radiotherapy and chemotherapy) and the diffusely
aggressive nature of glioma mean overall efficiency of glioma
treatment is poor, and the average survival time of patients with
high-grade glioma is <1 year (23). With the development of molecular
biology, research has focused on immunotherapy against glioma
(23). Therefore, it is necessary to
determine the characteristics of immunotolerance and
immunomodulation of glioma to develop an effective immunotherapy
strategy.
TLRs have been investigated as key PRRs in natural
immunity and recognize certain highly conserved molecular
structures of pathogens (24).
Moreover, TLRs are the bridge between innate immunity and acquired
immunity and are important to prevent biological pathogenic factor
infection (25).
TLRs are type I transmembrane proteins in mammals
and they are a family of conserved transmembrane signaling
receptors regulating innate immunity. TLRs are a member of the PRR
family and are associated with the recognition of PAMPs (26). TLR4 was the first identified human
cell TLR homologous to Drosophila, and is therefore the most
studied TLR (27). To date, 13 types
of TLRs (TLR 1–13) have been identified in the human body (28).
Although TLRs mediate innate immunity, they are not
unique to the innate immune system: In addition to immune cells
[monocytes/macrophages, T/B lymphocytes and dendritic cells
(29)], they are also expressed in
certain non-immune cells, such as endothelial (30), smooth muscle (31) and tumor cells (32). TLR expression levels in tumor cells
are key for tumorigenesis and progression, and this is an important
topic in tumor research (33). Among
TLRs, TLR4 is closely associated with tumors; high expression
levels of TLR4 accelerate tumor proliferation and mediate tumor
immune escape, for example in colon cancer, lung cancer and oral
squamous cell carcinoma (34–36).
Pandey et al (37)
investigated TLR4 polymorphisms and expression level patterns and
reported that TLR4 expression levels in different types of solid
tumor tissue promote tumor growth, invasion and metastasis.
Therefore, TLR4 expression levels in different tumor types may
serve as a marker for tumor proliferation, differentiation,
metastasis, prognosis and patient survival. Chen et al
(38) reported that TLR4 is
associated with liver cancer development; the activated TLR4/Nanog
oncogenic pathway is associated with the suppression of cytostatic
TGF-β signaling and may serve as a therapeutic target for hepatitis
C virus-associated hepatocellular carcinoma (HCC). Yao et al
(39) reported that the TLR4/STAT3
signaling pathway participates in the carcinogenesis, development,
invasion and metastasis of liver cancer. The authors also
demonstrated that M2-polarized macrophages facilitate
the migration and epithelial-mesenchymal transition of HCC cells
via the TLR4/STAT3 signaling pathway, suggesting that TLR4 may be a
novel therapeutic target. Szajnik et al (40) demonstrated that the TLR4/MyD88
signaling pathway promotes ovarian cancer development, drug
resistance and immune escape. Zhang and Zhang (41) reported that Foxp3 and TLR4 are highly
expressed in cervical cancer cells and are associated with clinical
stage and lymph node metastasis; Foxp3 and TLR4 may be useful
biomarkers for predicting the prognosis of patients and treating
cervical cancer. Zhan et al (42) revealed that the molecular factors
TLR4 and TLR3 affect the progression of lung cancer by regulating
autophagy, which may be a potential treatment strategy for lung
cancer. Li et al (43)
reported that TLR4 is highly expressed in breast cancer tissue and
promotes breast cancer metastasis via the AKT/GSK3β/β-catenin
pathway, which is stimulated by lipopolysaccharide. Numerous
studies have proved that TLR4 is associated with tumors (44,45), but
there are few studies on the association between TLR4 and the
occurrence and development of glioma. Hence, the present study
aimed to elucidate the association between TLR4 and the occurrence
and development of glioma in vitro, and to provide
experimental support to further investigate the pathogenesis of
glioma.
In the present study, TLR4 gene expression levels in
the human glioma cell line U-87MG were silenced by gene
interference. The use of only one cell line was a potential
limitation of the present study: In future investigations, multiple
glioma cell lines should be used to verify the regulatory effect of
TLR4 on proliferation. In the present study, a total of three
interference plasmids were designed for the human TLR4 gene. After
48 h transfection, flow cytometry detection demonstrated that the
transfection and silencing efficiency of TLR4-sh2 was the highest.
Therefore, TLR4-sh2 interference plasmids were selected for
subsequent experiments. The transfection efficiency of TLR4-sh2
interference and NC plasmid were 95.05±0.17 and 94.75±0.72%,
respectively. Results of fluorescence RT-qPCR revealed that the
TLR4 gene expression levels in U-87MG-sh cells decreased
significantly, which revealed that TLR4 gene silencing in U-87MG
cells was successful.
The present study investigated the association
between TLR4 and U-87MG cell proliferation. After silencing TLR4
gene expression levels in U-87MG cells via gene interference, flow
cytometry indicated that the PI of U-87MG-sh cells declined and the
G0/1 phase of the cell cycle extended significantly,
demonstrating that TLR4 gene silencing significantly inhibited the
proliferation of U-87MG cells. Following TLR4 gene silencing, CCK-8
demonstrated that U-87MG cell proliferation was significantly
inhibited, which was consistent with the flow cytometry results.
The absence of colony formation assay was a potential limitation of
the present study: In future investigations, colony formation assay
should be used to verify the CCK-8 results. Following TLR4 gene
silencing in U-87MG cells, flow cytometry revealed that the
apoptotic rate of U-87MG cells increased significantly, indicating
that silencing the TLR4 gene inhibited growth and induced apoptosis
in the glioma cell line U-87MG. TLR4 may therefore represent a
target for the development of anti-glioma drugs and treatment
methods.
TLR4-sh interference plasmids were successfully
transfected into U-87MG cells via liposome transfection and stably
transfected cells were screened using puromycin to establish a
glioma cell model with low TLR4 gene expression levels. The present
study demonstrated that the TLR4 gene is associated with
proliferation and apoptosis regulation in U-87MG cells. Silencing
the TLR4 gene in U-87MG cells significantly inhibited cell
proliferation and induced U-87MG cell apoptosis, providing an
experimental basis for the development of anti-glioma drugs and
therapeutic methods targeting the TLR4 gene. The absence of
clinical data (expression levels in tissue, association with
patient prognosis) was a limitation of the present study. The
molecular mechanisms underlying growth inhibition and cell
apoptosis induction of U-87MG cells by silencing the TLR4 gene are
complex, necessitating further study in the future.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Excellent
Talent Project in Clinical Medicine funded by Hebei provincial
government in 2017 [grant no. Jicaishe(2017)46].
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YL performed the experiments and wrote the
manuscript. YJ performed the experiments and statistical analysis.
JL, YC and XH performed the experiments. LL designed the study,
performed the experiments and revised the manuscript. All authors
read and approved the final version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Davis ME: Epidemiology and overview of
gliomas. Semin Oncol Nurs. 34:420–429. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ostrom QT, Gittleman H, Stetson L, Virk SM
and Barnholtz-Sloan JS: Epidemiology of gliomas. Cancer Treat Res.
163:1–14. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ostrom QT, Gittleman H, Liao P, Rouse C,
Chen Y, Dowling J, Wolinsky Y, Kruchko C and Barnholtz-Sloan J:
CBTRUS statistical report: Primary brain and central nervous system
tumors diagnosed in the United States in 2007–2011. Neuro Oncol. 16
(Suppl 4):iv1–iv63. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rasmussen BK, Hansen S, Laursen RJ,
Kosteljanetz M, Schultz H, Nørgård BM, Guldberg R and Gradel KO:
Epidemiology of glioma: Clinical characteristics, symptoms, and
predictors of glioma patients grade I–IV in the the Danish
neuro-oncology registry. J Neurooncol. 135:571–579. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kawai T and Akira S: Toll-like receptors
and their crosstalk with other innate receptors in infection and
immunity. Immunity. 34:637–650. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kumar H, Kawai T and Akira S: Toll-like
receptors and innate immunity. Biochem Biophys Res Commun.
388:621–625. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hua Z and Hou B: TLR signaling in B-cell
development and activation. Cell Mol Immunol. 10:103–106. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kawai T and Akira S: TLR signaling. Semin
Immunol. 19:24–32. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Seydoux E, Liang H, Dubois Cauwelaert N,
Archer M, Rintala ND, Kramer R, Carter D, Fox CB and Orr MT:
Effective combination adjuvants engage both TLR and inflammasome
pathways to promote potent adaptive immune responses. J Immunol.
201:98–112. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tian S, Wang M, Liu C, Zhao H and Zhao B:
Mulberry leaf reduces inflammation and insulin resistance in type 2
diabetic mice by TLRs and insulin signalling pathway. BMC
Complement Altern Med. 19:3262019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dickinson SE and Wondrak GT: TLR4-directed
molecular strategies targeting skin photodamage and carcinogenesis.
Curr Med Chem. 25:5487–5502. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang YH, Cao YW, Yang XC, Niu HT, Sun LJ,
Wang XS and Liu J: Effect of TLR4 and B7-H1 on immune escape of
urothelial bladder cancer and its clinical significance. Asian Pac
J Cancer Prev. 15:1321–1326. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Monlish DA, Bhatt ST and Schuettpelz LG:
The role of toll-like receptors in hematopoietic malignancies.
Front Immunol. 7:3902016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dajon M, Iribarren K and Cremer I:
Toll-like receptor stimulation in cancer: A pro- and anti-tumor
double-edged sword. Immunobiology. 222:89–100. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Makkar S, Riehl TE, Chen B, Yan Y,
Alvarado DM, Ciorba MA and Stenson WF: Hyaluronic acid binding to
TLR4 promotes proliferation and blocks apoptosis in colon cancer.
Mol Cancer Ther. 18:2446–2456. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sun NK, Huang SL, Chang TC and Chao CC:
TLR4 and NFκB signaling is critical for taxol resistance in ovarian
carcinoma cells. J Cell Physiol. 233:2489–2501. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shetab Boushehri MA and Lamprecht A:
TLR4-based immunotherapeutics in cancer: A review of the
achievements and shortcomings. Mol Pharm. 15:4777–4800. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dapito DH, Mencin A, Gwak GY, Pradere JP,
Jang MK, Mederacke I, Caviglia JM, Khiabanian H, Adeyemi A,
Bataller R, et al: Promotion of hepatocellular carcinoma by the
intestinal microbiota and TLR4. Cancer Cell. 21:504–516. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Allen M, Bjerke M, Edlund H, Nelander S
and Westermark B: Origin of the U87MG glioma cell line: Good news
and bad news. Sci Transl Med. 8:354re32016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhou T, Li Y, Yang L, Liu L, Ju Y and Li
C: Silencing of ANXA3 expression by RNA interference inhibits the
proliferation and invasion of breast cancer cells. Oncol Rep.
37:388–398. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ostrom QT, Bauchet L, Davis FG, Deltour I,
Fisher JL, Langer CE, Pekmezci M, Schwartzbaum JA, Turner MC, Walsh
KM, et al: The epidemiology of glioma in adults: A ‘state of the
science’ review. Neuro Oncol. 16:896–913. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lim M, Xia Y, Bettegowda C and Weller M:
Current state of immunotherapy for glioblastoma. Nat Rev Clin
Oncol. 15:422–442. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Satoh T and Akira S: Toll-like receptor
signaling and its inducible proteins. Microbiol Spectr.
4:2016.PubMed/NCBI
|
|
25
|
Steinhagen F, Kinjo T, Bode C and Klinman
DM: TLR-based immune adjuvants. Vaccine. 29:3341–3355. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kumar V: Toll-like receptors in the
pathogenesis of neuroinflammation. J Neuroimmunol. 332:16–30. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Medzhitov R, Preston-Hurlburt P and
Janeway CA Jr: A human homologue of the Drosophila Toll
protein signals activation of adaptive immunity. Nature.
388:394–397. 1997. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Vijay K: Toll-like receptors in immunity
and inflammatory diseases: Past, present, and future. Int
Immunopharmacol. 59:391–412. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mikulic J, Longet S, Favre L, Benyacoub J
and Corthesy B: Secretory IgA in complex with Lactobacillus
rhamnosus potentiates mucosal dendritic cell-mediated Treg cell
differentiation via TLR regulatory proteins, RALDH2 and secretion
of IL-10 and TGF-β. Cell Mol Immunol. 14:546–556. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tang AT, Choi JP, Kotzin JJ, Yang Y, Hong
CC, Hobson N, Girard R, Zeineddine HA, Lightle R, Moore T, et al:
Endothelial TLR4 and the microbiome drive cerebral cavernous
malformations. Nature. 545:305–310. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yin Q, Jiang D, Li L, Yang Y, Wu P, Luo Y,
Yang R and Li D: LPS promotes vascular smooth muscle cells
proliferation through the TLR4/Rac1/Akt signalling pathway. Cell
Physiol Biochem. 44:2189–2200. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Fu HY, Li C, Yang W, Gai XD, Jia T, Lei YM
and Li Y: FOXP3 and TLR4 protein expression are correlated in
non-small cell lung cancer: Implications for tumor progression and
escape. Acta Histochem. 115:151–157. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Grimmig T, Matthes N, Hoeland K, Tripathi
S, Chandraker A, Grimm M, Moench R, Moll EM, Friess H, Tsaur I, et
al: TLR7 and TLR8 expression increases tumor cell proliferation and
promotes chemoresistance in human pancreatic cancer. Int J Oncol.
47:857–866. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Xiao T, Wu S, Yan C, Zhao C, Jin H, Yan N,
Xu J, Wu Y, Li C, Shao Q and Xia S: Butyrate upregulates the TLR4
expression and the phosphorylation of MAPKs and NK-κB in colon
cancer cell in vitro. Oncol Lett. 16:4439–4447.
2018.PubMed/NCBI
|
|
35
|
Li C, Li H, Jiang K, Li J and Gai X: TLR4
signaling pathway in mouse Lewis lung cancer cells promotes the
expression of TGF-β1 and IL-10 and tumor cells migration. Biomed
Mater Eng. 24:869–875. 2014.PubMed/NCBI
|
|
36
|
Ren WH, Zhang LM, Liu HQ, Gao L, Chen C,
Qiang C, Wang XL, Liu CY, Li SM, Huang C, et al: Protein
overexpression of CIRP and TLR4 in oral squamous cell carcinoma: An
immunohistochemical and clinical correlation analysis. Med Oncol.
31:1202014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Pandey N, Chauhan A and Jain N: TLR4
polymorphisms and expression in solid cancers. Mol Diagn Ther.
22:683–702. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chen CL, Tsukamoto H, Liu JC, Kashiwabara
C, Feldman D, Sher L, Dooley S, French SW, Mishra L, Petrovic L, et
al: Reciprocal regulation by TLR4 and TGF-β in tumor-initiating
stem-like cells. J Clin Invest. 123:2832–2849. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yao RR, Li JH, Zhang R, Chen RX and Wang
YH: M2-polarized tumor-associated macrophages facilitated migration
and epithelial-mesenchymal transition of HCC cells via the
TLR4/STAT3 signaling pathway. World J Surg Oncol. 16:92018.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Szajnik M, Szczepanski MJ, Czystowska M,
Elishaev E, Mandapathil M, Nowak-Markwitz E, Spaczynski M and
Whiteside TL: TLR4 signaling induced by lipopolysaccharide or
paclitaxel regulates tumor survival and chemoresistance in ovarian
cancer. Oncogene. 28:4353–4363. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhang H and Zhang S: The expression of
Foxp3 and TLR4 in cervical cancer: Association with immune escape
and clinical pathology. Arch Gynecol Obstet. 295:705–712. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhan Z, Xie X, Cao H, Zhou X, Zhang XD,
Fan H and Liu Z: Autophagy facilitates TLR4- and TLR3-triggered
migration and invasion of lung cancer cells through the promotion
of TRAF6 ubiquitination. Autophagy. 10:257–268. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Li J, Yin J, Shen W, Gao R, Liu Y, Chen Y,
Li X, Liu C, Xiang R and Luo N: TLR4 promotes breast cancer
metastasis via Akt/GSK3β/β-catenin pathway upon LPS stimulation.
Anat Rec (Hoboken). 300:1219–1229. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kim KH, Jo MS, Suh DS, Yoon MS, Shin DH,
Lee JH and Choi KU: Expression and significance of the TLR4/MyD88
signaling pathway in ovarian epithelial cancers. World J Surg
Oncol. 10:1932012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhou Q, Wang C, Wang X, Wu X, Zhu Z, Liu B
and Su L: Association between TLR4 (+896A/G and +1196C/T)
polymorphisms and gastric cancer risk: An updated meta-analysis.
PLoS One. 9:e1096052014. View Article : Google Scholar : PubMed/NCBI
|