Introduction
The basement membrane is a highly specialized
extracellular matrix (ECM) that is located around all cell
monolayers and multi-layered epithelia within the body (1–4). Its
functions include structural support, regulation of cell signalling
behaviors and providing a protective barrier. The main components
of basement membranes include collagen IV, laminin, perlecan and
nidogen. Of these components, it is known that collagen IV and
laminin can self-assemble into superstructures individually, which
plays a vital role in the stability of BMs. Perlecan and nidogen
(NID) influence the structural integrity through connecting the
collagen IV and laminin networks, increasing stability (1–4). NID,
also known as entactin, mimics the function of perlecan by
connecting the superstructures of collagen IV and laminin (4). Collagen IV contains a triple helical
domain that fuses to the G2 domain of NID, while the γ-3 chain of
laminin binds to the G3 domain of NID, effectively linking the two
components (1,2,4).
Only a small number of studies have investigated the
impact of NID1 in cancer with only 3 studies evaluating breast
cancer (5–7). A study by Alečković et al
(5) performed a secretome analysis
of lung metastatic sublines of the human breast cancer cell line
MDA-MB-231 and identified NID1 as potential regulator of
metastasis. The importance of NID1 in lung cancer metastasis was
further examined in MDA-MB-231 overexpressing NID1.
Overexpression of NID1 increased lung metastases 10-fold and
reduced survival in an experimental metastasis assay as well as
promoting invasion and migration in a transwell assay (5). These investigators also showed that
NID1 expression in breast cancer patients was associated
with lung metastasis and poor patient prognosis (5). However, a recent study by Ferraro et
al (7) showed the opposite;
secretion of NID1 from endothelial cells inhibited the migration of
the human breast cancer cell line SK-BR-3. Therefore, the impact of
NID1 on breast cancer migration is inconclusive. The third study
evaluating the function of NID1 in breast cancer found that a
disintegrin and metalloproteinase with thrombospondin repeats 1
(ADAMTS1) can cleave NID1 inhibiting its function. ADAMTS1 levels
were shown to be reduced in breast cancer samples and this was
associated with reduced NID1 proteolysis and increased deposition
of NID1 into tumor blood vessels (6).
A previous study from our lab looking at miR-200
levels found that re-expression of the miR-200b/200a/429 cluster
impaired tumor growth and metastasis (8). RNA-sequencing revealed that Nid1
was the most significantly reduced gene in miR-200 transfected
cells compared to control cells (8).
Given that a higher Nid1 expression was associated with
increased tumorigenicity and metastatic capacity this study
evaluated the impact of Nid1 on mammary tumor cell
proliferation and migration/invasion.
Materials and methods
Cell lines
RJ423 cells have been previously described (8–11). To
reduce Nid1 expression, RJ423 cells were transfected with
four unique shRNAs targeting Nid1 (cat. no. TL501474;
Origene Technologies Inc.) or a control shRNA (cat. no. TR30021;
Origene Technologies Inc.). Stable clones were selected using 5
µg/ml puromycin (InvivoGen). Two knockdown lines
(RJ423shNid1A and RJ423shNid1D) were identified as
having the greatest Nid1 mRNA knockdown relative to a
control line (RJ423con). All cells were cultured as described in
(8). RJ423shNid1A,
RJ423shNid1D and RJ423con cells were maintained in 5 µg/ml
puromycin.
RNA isolation and RT-qPCR
RNA isolation and RT-qPCR were performed as
previously described (8). Primers
for Nid1 (qMmuCID0018281), Cdh1 (qMmuCED0044197),
Vim (qMmuCID0005527), Snai1 (qMmuCID0024342),
Snai2 (qMmuCED0046072), Twist1 (qMmuCED0004065),
Twist2 (qMmuCID0009652), Zeb1 (qMmuCID0009095),
Zeb2 (qMmuCID0014662) and Hprt (qMmuCID0005679) were
purchased from Bio-Rad Laboratories (Canada) Ltd. Gene expression
was quantified and normalized to Hprt using the normalized
expression (ΔΔCq) (12) mode in
CFX-Manager 3.1 software [Bio-Rad Laboratories (Canada) Ltd.].
Collection of conditioned media
Cells were seeded in 10 mm culture dishes at a
density of 2×106 cells per dish overnight in fully
supplemented media. The next day, the fully supplemented media was
removed, the cells were washed twice with phosphate-buffered saline
(PBS) and 10 ml of serum-free media containing sodium pyruvate,
glutamine and antibiotic-antimycotic was added to each dish.
Twenty-four hours later, conditioned media was collected and either
used immediately or stored at −20°C.
Western blotting
Western blotting was performed as previously
described (13). NID1 protein was
detected using a primary antibody against NID1 (cat. no.
NBP1-97701; Novus Biologicals) at a 1:200 dilution while β-actin
was detected using a 1:5,000 dilution of a primary antibody against
β-actin (cat. no. 8457; Cell Signaling Technologies, Inc.). The
appropriate HRP-linked secondary antibody (Cell Signaling
Technology, Inc.) was used at a 1:2,000 dilution.
Cell proliferation
Immunofluorescence for phospho-histone H3 (PH3) was
used to determine cell proliferation as previously described
(8). For the conditioned media
experiments, cells were allowed to attach to the coverslips
overnight in fully supplemented media. The next day, the fully
supplemented media was removed, the cells washed twice with PBS,
and then the cells were cultured in conditioned media for 24 h
prior to assessing proliferation.
Cell apoptosis
Apoptosis was assessed using the Annexin V-FITC
Apoptosis Detection kit (eBioscience) according to the
manufacturer's protocol. A BD Accuri C6 flow cytometer (BD
Biosciences) was used to measure fluorescence at a flow rate less
than 400 events/second and analyzed using the Accuri C6 software
(BD Biosciences).
Transwell invasion chamber assay
Transwell assays were performed as previously
described (8) with minor
alterations. Since Matrigel contains NID1, the Cultrex®
24 Well Collagen IV Cell Invasion Assay kit (cat. no. 3458-024-K)
was used for the transwell inserts. The amount of cell migration
was analyzed using ImageJ software (National Institutes of Health).
To determine whether NID1 served as a chemoattractant, cells were
cultured in serum-free media in the upper chamber with conditioned
media from the various cell lines in the lower chamber. To
determine whether NID1 directly interacted with cells to alter
migration, cells were cultured in conditioned media in the upper
chamber and fully supplemented media was placed in the lower
chamber.
Statistical analysis
Statistical analysis was conducted using one-way
ANOVA followed by Tukey's test using GraphPad Prizm 8 (GraphPad
Software, Inc.). Statistical significance (P<0.05) is indicated
by asterisks.
Results
Nid1 shRNA reduces NID1 protein levels
in conditioned media
Our previous worked revealed that re-expressing the
miR-200b/200a/429 cluster in murine mammary tumor cells
(RJ423-200ba429) suppressed tumor growth and metastasis in
vivo (8). RNA sequencing of the
RJ423-200ba429 cells revealed that Nid1 was the most
significant, downregulated gene (Nid1 expression was
decreased 3,821-fold, P-value 3.5×10−268, GSE113162).
Consistent with this finding, Nid1 was expression was
reduced 6,654-fold (P-value 3.5×10−240) in the poorly
tumorigenic, non-metastatic murine mammary tumor cell line RJ345
compared to the highly tumorigenic, highly metastatic murine
mammary tumor cell line RJ423. Since both RJ345 cells and
RJ423-200ba429 cells are poorly tumorigenic, non-metastatic
(8) and express almost no
Nid1 (Table I) this study
examined whether reducing Nid1 in RJ423 cells could
negatively impact proliferation and/or migration.
 | Table I.Characteristics of murine mammary
tumor cells. |
Table I.
Characteristics of murine mammary
tumor cells.
| Cell lines | miR-200
expression | Nid1
expression | Tumorigenicity | Metastatic |
|---|
| RJ345 | High | Low | Low | No |
| RJ423 | Low | High | High | Yes |
| RJ423-200ba429 | High | Low | Low | No |
| RJ423EV | Low | High | High | Yes |
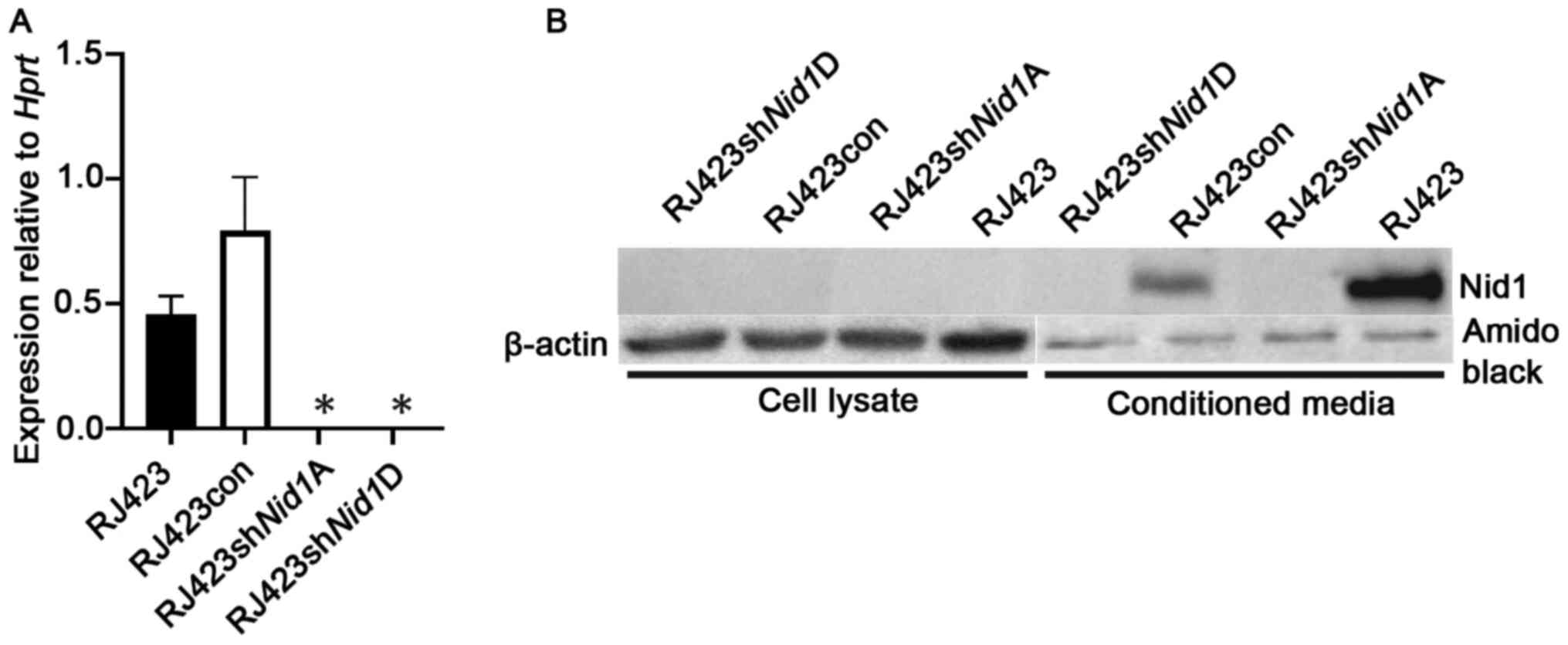
To reduce the expression of Nid1, RJ423 cells
were stably transfected with 4 unique shRNA constructs (A-D)
targeting mouse Nid1 or a control shRNA construct. The two
most effective Nid1 shRNA constructs, shNid1A and
shNid1D, reduced. Nid1 mRNA levels over 99% compared
to RJ423 cells stably expressing the control shRNA construct
(RJ423con; Fig. 1A).
To confirm NID1 was suppressed at the protein level
western blotting was performed on cell lysates from RJ423,
RJ423con, RJ423shNid1A and RJ423shNid1D cells. The
NID1 antibody failed to detect NID1 protein in any of the cell
lysates (Fig. 1B). Since NID1 is a
secreted protein conditioned media was collected from all four cell
lines. As shown in Fig. 1B, NID1 was
detected in the condition media from RJ423 cells and RJ423con cells
but not from either RJ423shNid1A or RJ423shNid1D
cells.
Nid1 knockdown inhibits proliferation
but not apoptosis
The impact of Nid1 suppression on cell
proliferation was assessed using PH3 immunofluorescence. Both
Nid1 shRNA constructs significantly reduced proliferation
compared to the control cell line, RJ423con (Fig. 2A). The impact of Nid1
suppression on apoptosis was evaluated using Annexin V flow
cytometry. RJ423, RJ423con, RJ4223shNid1A and
RJ423shNid1D cells all had very low levels of apoptosis
(<1%) and this level of basal apoptosis when cultured in fully
supplemented media (Fig. 2B). Since
there was no significant difference in apoptosis this cell response
was not further evaluated.
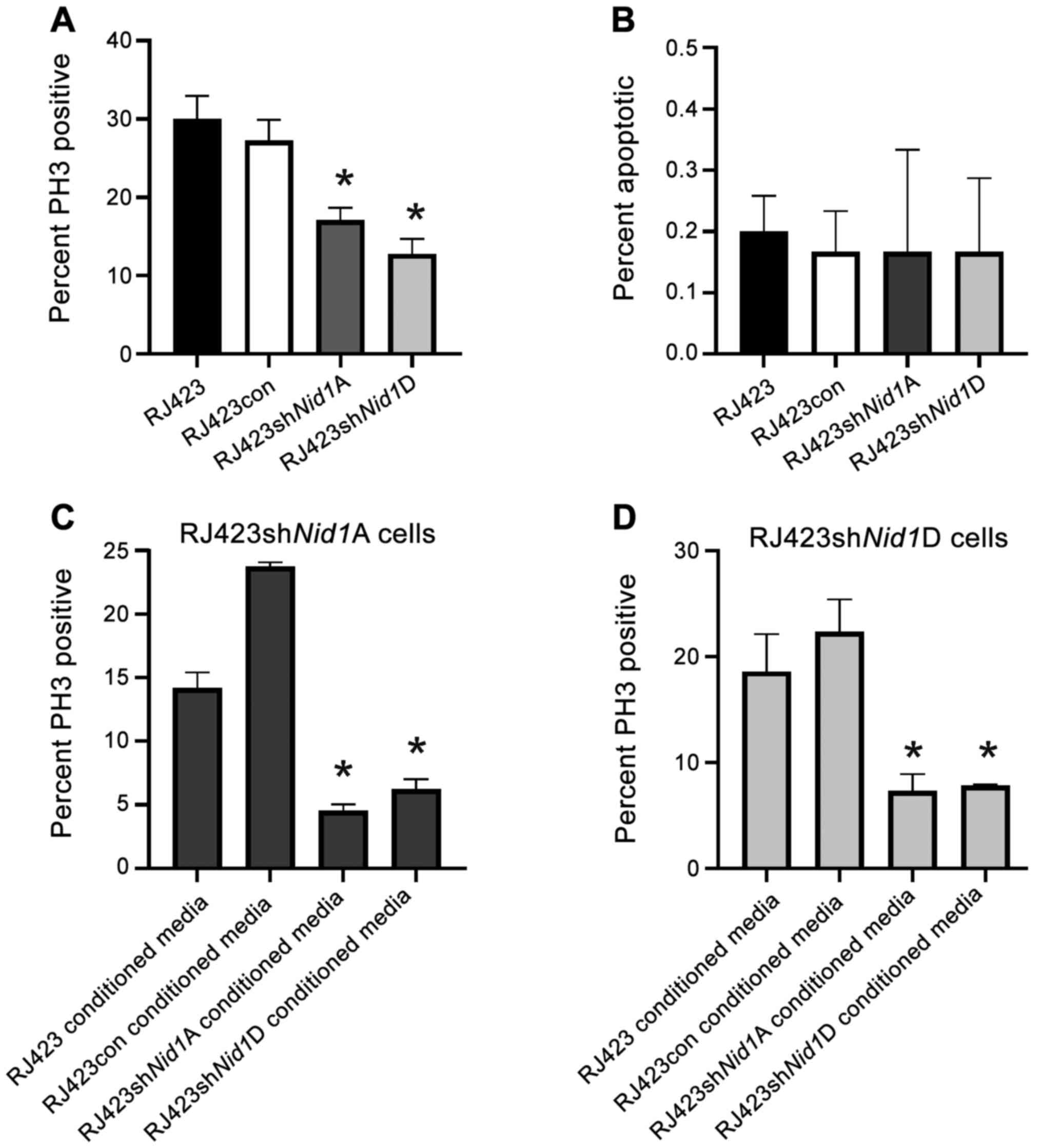 | Figure 2.Impact of NID1 conditioned media on
cell proliferation and apoptosis. (A) Percentage of PH3-positive
cells in parental RJ423, RJ423con, RJ423shNid1A and
RJ423shNid1D cells. (B) Percentage of apoptotic cells in
RJ423, RJ423con, RJ423Nid1A and RJ423Nid1D cells.
Percentage of PH3-positive cells from (C) RJ423shNid1A and
(D) RJ423shNid1D cells treated with conditioned media from
RJ423, RJ423con, RJ423shNid1A or RJ423shNid1D cells
for 24 h. *P<0.05 vs. RJ423con cells (n=3). Nid1, nidogen
1; con, control; sh, short hairpin RNA; PH3, phospho-histone
H3. |
To confirm the decrease in proliferation resulted
from Nid1 suppression, RJ423shNid1A and
RJ423shNid1D cells were cultured in the presence of
conditioned media from RJ423 cells, RJ423con cells, their own
conditioned media and conditioned media from the other knockdown
line. It was anticipated that conditioned media from RJ423 or
RJ423con cells would increase proliferation in the Nid1
knockdown cells as both RJ423 and RJ423con cells secrete NID1 into
the media. As shown in Fig. 2C and D
proliferation of RJ423shNid1A and RJ423shNid1D cells
was significantly increased when the cells were grown in
conditioned media from RJ423con cells compared to when these cells
were grown in conditioned media from either Nid1 knockdown
cell line. Proliferation of RJ423shNid1A cells was also
significantly increased when the cells were grown in conditioned
media from RJ423 cells compared to when these cells were grown in
conditioned media from either Nid1 knockdown cell line while
the proliferation in the RJ423shNid1D cells was close to
being significant when these cells were grown in condition media
from RJ423 cells compared to condition media from
RJ423shNid1A (P=0.058) or RJ423shNid1D (P=0.081)
cells.
Nid1 knockdown impairs cell
migration
Next, migration and invasion were assessed using a
transwell assay. The transwell inserts were coated with collagen IV
rather than Matrigel as Matrigel contains NID1. Fig. 3A-D shows representative images of the
stained cells that invaded and migrated through the collagen IV
coated inserts. Quantification of these images (Fig. 3E) revealed that RJ423shNid1A
and RJ423shNid1D cells were significantly less effective at
invading and migrating than either the RJ423 or RJ423con cells.
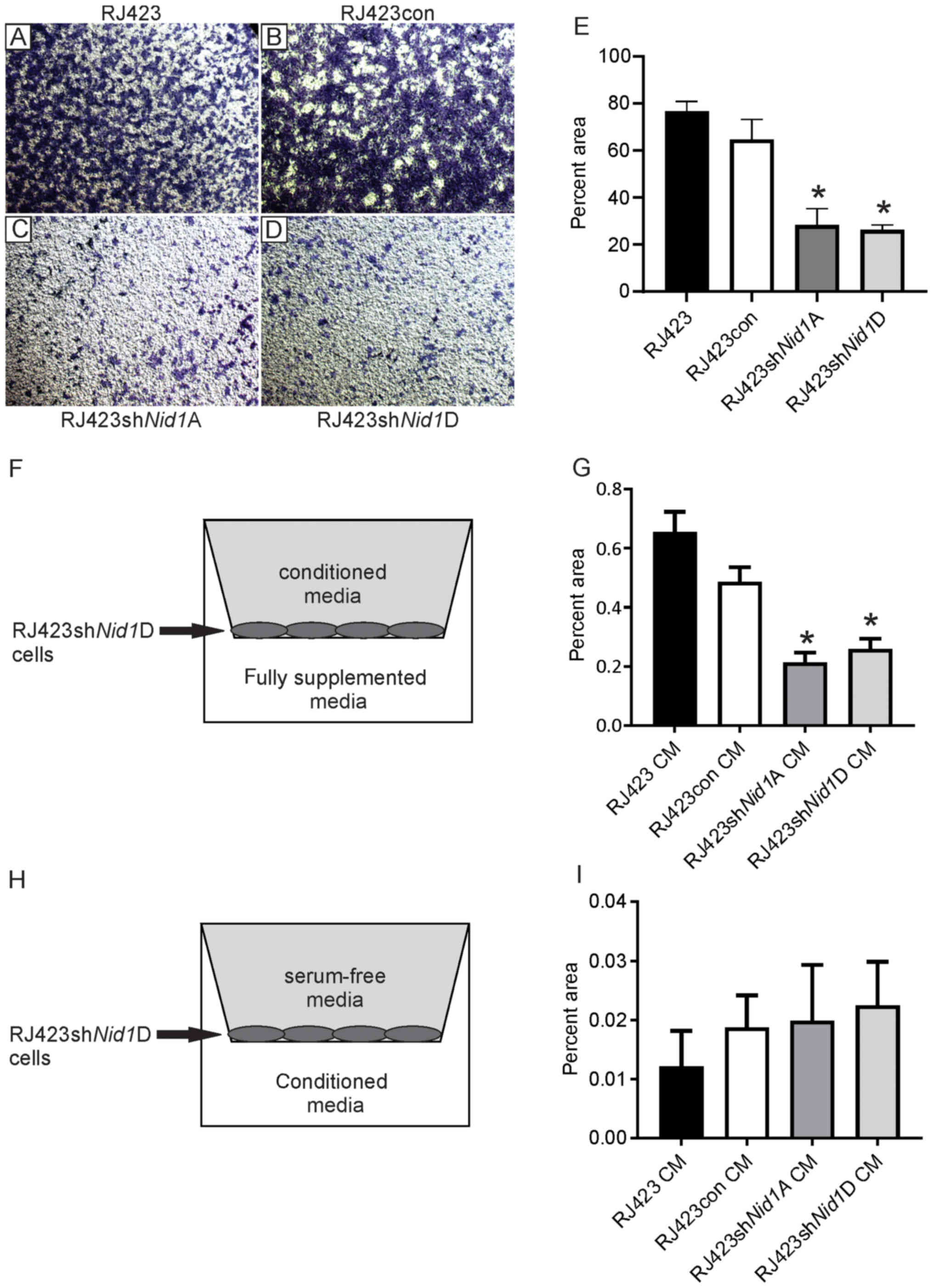 | Figure 3.Impact of NID1 conditioned media on
cell migration. Representative images of toluidine blue stained
cells on the bottom of collagen-coated transwell inserts for (A)
parental RJ423, (B) RJ423con, (C) RJ423shNid1A and (D)
RJ423shNid1D cells (magnification, ×4). (E) Quantification
of the percent area occupied by parental RJ423, RJ423con,
RJ423shNid1A and RJ423shNid1D cells. Migration and
invasion of RJ423shNid1D cells when conditioned media from
RJ423, RJ423con, RJ423shNid1A or RJ423shNid1D cells
were placed (F and G) in the upper chamber of the well with the
cells or (H and I) in the lower chamber. (F and H) Schematic
indicating the location of the conditioned media and cells. (G and
I) Quantitative data from these experiments. *P<0.05 vs.
RJ423con cells (n=3). Nid1, nidogen 1; con, control; sh, short
hairpin RNA; CM, conditioned media. |
To confirm that migration and invasion was dependent
on NID1, migration of RJ423shNid1D cells was assessed using
conditioned media from RJ423, RJ423con, RJ423shNid1A or
RJ423shNid1D cells in the upper chamber and fully
supplemented media in the lower chamber (Fig. 3F). The presence of conditioned media
from RJ423 and RJ423con cells significantly increased invasion and
migration of RJ423shNid1D cells compared the conditioned
media from either RJ423shNid1A or RJ423shNid1D cells
(Fig. 3G).
To assess whether NID1 served as a chemoattractant,
RJ423shNid1D cells were grown in the upper chamber in
serum-free media and conditioned media from RJ423, RJ423con,
RJ423shNid1A or RJ423shNid1D cells was placed in the
lower chamber (Fig. 3H). As shown in
Fig. 3I, RJ423shNid1A cells
invaded and migrated very poorly independent of the conditioned
media used as a chemoattractant and there were no significant
differences observed.
Enhanced migration and invasion are often associated
with increased expression of mesenchymal genes and loss of
epithelial genes in a process known as epithelial to mesenchymal
transition (EMT). It was anticipated that one or more mesenchymal
genes would be expressed at significantly reduced levels or an
increase in expression of the epithelial gene, Cdh1, would
be observed in the Nid1 knockdown lines compared to RJ423con
and RJ423 parental cells. The only gene significantly altered was
Vim in the Nid1 knockdown lines, however Vim
expression in the Nid1 knockdown lines was almost identical
to the parental RJ423 cells (Fig.
4).
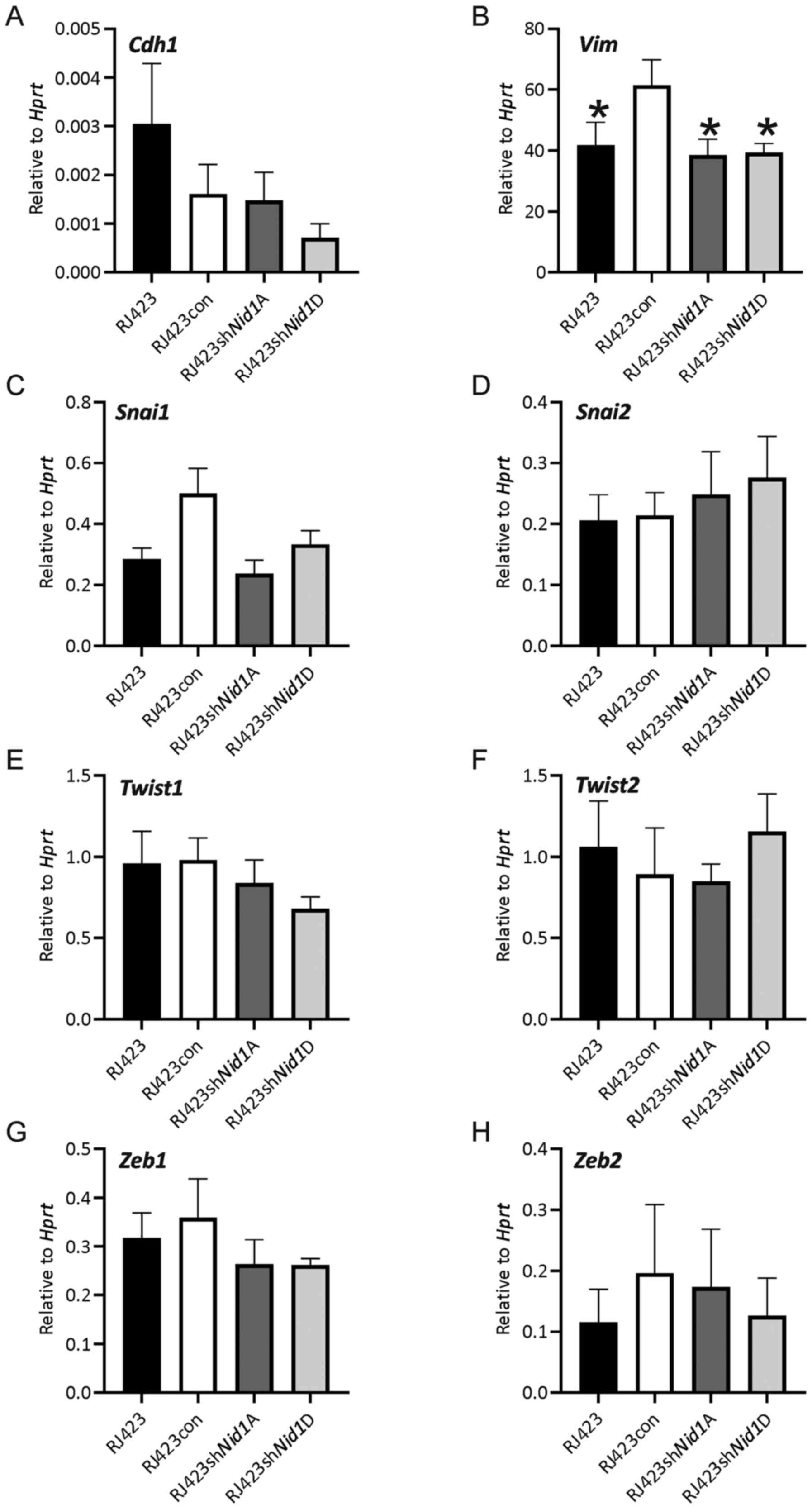 | Figure 4.Expression levels of epithelial and
mesenchymal genes following Nid1 knockdown. mRNA expression
levels of (A) Cdh1, (B) Vim, (C) Snai1, (D)
Snai2, (E) Twist1, (F) Twist2, (G) Zeb1
or (H) Zeb2 relative to Hprt. *P<0.05 vs. RJ423con cells
(n=5). Nid1, nidogen 1; Hprt, hypoxanthine
phosphoribosyltransferase; con, control; sh, short hairpin RNA. |
Discussion
Our previous work looking at highly tumorigenic and
metastatic murine mammary tumor cells compared to weakly
tumorigenic, non-metastatic tumor cells identified Nid1 as a
potential promoter of tumor development and metastasis (8). RJ423 cells are murine mammary tumor
cells with characteristics of human claudin-low breast cancer that
express high levels of Nid1. RJ423 cells rapidly form tumors
when injected into the mammary fat pad of mice and induce lung
metastases in mice following tail vein injection (8).
Suppression of Nid1 expression in RJ423 cells
significantly reduced cell proliferation and this decrease in
proliferation was rescued by incubating cells in conditioned media
containing NID1. While it is possible that the conditioned media
from RJ423 and RJ423con cells contain proteins other than NID1,
presumably the conditioned media from RJ423con and
RJ423shNid1A/RJ423shNid1D cells should be highly
similar as the only difference between these cells was the stable
knockdown of Nid1. While treating the cells with recombinant
NID1 would confirm the impact of NID1 on proliferation,
commercially available, full-length recombinant murine NID1 could
not be found. In a study by Alečković et al (5), proliferation of MDA-MB-231 cells
overexpressing NID1 cultured in vitro was not
examined. However, this group did assess tumor size in vivo
and found that overexpression of NID1 in MDA-MB-231 did not
increase primary tumor growth. No other study investigated the
impact of NID1 on breast cancer cell proliferation in
vitro.
NID1 did not impact basal levels of apoptosis as
RJ423con, RJ423shNid1A, and RJ423shNid1D cells. As no
study has investigated the impact of NID1 on breast cancer cell
apoptosis this is the first study showing that a reduction of NID1
does not increase mammary tumor cell apoptosis.
Reducing NID1 levels significantly impaired
migration and invasion through collagen IV coated membranes and
migration/invasion could be rescued when conditioned media
containing NID1 was placed in the upper chamber with the
RJ423shNid1D cells but not when the conditioned media
containing NID1 was placed in the lower chamber. These findings
suggest that NID1 is interacting directly with the cancer cells and
not acting as a chemoattractant. It is not clear how NID1 alters
cell migration and invasion, but it could be through binding to
receptors on the cell and influencing signaling. NID1 has been
reported to bind to integrins αvβ3 and α3β1 (14–16) and
RNA sequencing by our lab has shown the presence of Itga3,
Itgb3, and Itgb1 but not Itgav in RJ423 cells
(GSE113162). Therefore, NID1 is presumably signaling through
integrin α3β1 or another receptor that has yet to be identified.
The observation that NID1 promotes migration and invasion is
consistent with the findings of Alečković et al (5) who showed that MDA-MB-231 cells
overexpressing NID1 increased migration and invasion in
vitro and lung metastatic colonization in vivo. However,
Ferraro et al (7) found that
NID1 secreted by endothelial cells inhibited invasion a migration
of the human breast cancer cell line, SK-BR-3. The main difference
between the study by Ferraro et al (7) and our study and the study by Alečković
et al (5) was that while the
Ferraro et al (7) study
utilized breast cancer cells with HER2 overexpression while our
study (9) and the study by Alečković
et al (5) used tumor cells
with characteristics of claudin-low breast cancer. Thus, NID1 may
promote migration and invasion of claudin-low breast cancer cells
but not in other breast cancer subtypes.
Enhanced invasion and migration are often
accompanied by an increase in mesenchymal gene expression, reduced
epithelial gene expression and a more spindled-shape morphology.
Moreover, NID1 has been implicated in inducing EMT in colorectal
cancer cells (17) and ovarian
cancer cells (18) but has not been
investigated in breast cancer. No consistent increase in
mesenchymal gene expression or decrease in epithelial gene
expression was observed in either RJ423shNid1A or
RJ423shNid1D cells compared to RJ423con cells suggesting
that NID1 cannot restore epithelial gene expression patterns in
claudin-low mammary tumor cells.
Acknowledgements
Not applicable.
Funding
The present study was supported by a CIHR operating
grant (MOP-136970) and a CIHR project grant (PJT-162218) to
RAM.
Availability of data and materials
The datasets generated and/or analyzed during the
current study are available in the Gene Expression Omnibus
repository, https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE113162
Authors' contributions
RJ performed reverse transcription-quantitative PCR,
western blotting for NID1, proliferation assays, apoptosis assays
and Transwell migration assays. CJM performed reverse
transcription-quantitative PCR on some of the samples for
Nid1 and some of the epithelial and mesenchymal genes. RAM
created the cell lines, edited the manuscript and ran the project.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Jayadev R and Sherwood DR: Basement
membranes. Curr Biol. 27:R207–R211. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
LeBleu VS, Macdonald B and Kalluri R:
Structure and function of basement membranes. Exp Biol Med
(Maywood). 232:1121–1129. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chang J and Chaudhuri O: Beyond proteases:
Basement membrane mechanics and cancer invasion. J Cell Biol.
218:2456–2469. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yurchenco PD: Basement membranes: cell
scaffoldings and signaling platforms. Cold Spring Harb Perspect
Biol. 3:a0049112011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Alečković M, Wei Y, LeRoy G, Sidoli S, Liu
DD, Garcia BA and Kang Y: Identification of Nidogen 1 as a lung
metastasis protein through secretome analysis. Genes Dev.
31:1439–1455. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Martino-Echarri E, Fernández-Rodríguez R,
Rodríguez-Baena FJ, Barrientos-Durán A, Torres-Collado AX,
Plaza-Calonge Mdel C, Amador-Cubero S, Cortés J, Reynolds LE,
Hodivala-Dilke KM and Rodríguez-Manzaneque JC: Contribution of
ADAMTS1 as a tumor suppressor gene in human breast carcinoma.
Linking its tumor inhibitory properties to its proteolytic activity
on nidogen-1 and nidogen-2. Int J Cancer. 133:2315–2324. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ferraro DA, Patella F, Zanivan S, Donato
C, Aceto N, Giannotta M, Dejana E, Diepenbruck M, Christofori G and
Buess M: Endothelial cell-derived nidogen-1 inhibits migration of
SK-BR-3 breast cancer cells. BMC Cancer. 19:3122019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Watson KL, Jones RA, Bruce A and Moorehead
RA: The miR-200b/200a/429 cluster prevents metastasis and induces
dormancy in a murine claudin-low mammary tumor cell line. Exp Cell
Res. 369:17–26. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jones R, Watson K, Bruce A, Nersesian S,
Kitz J and Moorehead R: Re-expression of miR-200c suppresses
proliferation, colony formation and in vivo tumor growth of murine
claudin-low mammary tumor cells. Oncotarget. 8:23727–23749. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Campbell CI and Moorehead RA: Mammary
tumors that become independent of the type I insulin-like growth
factor receptor express elevated levels of platelet-derived growth
factor receptors. BMC Cancer. 11:4802011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Campbell CI, Thompson DE, Siwicky MD and
Moorehead RA: Murine mammary tumor cells with a claudin-low
genotype. Cancer Cell Int. 11:282011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chorner PM and Moorehead RA: A-674563, a
putative AKT1 inhibitor that also suppresses CDK2 activity,
inhibits human NSCLC cell growth more effectively than the pan-AKT
inhibitor, MK-2206. PLoS One. 13:e01933442018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dedhar S, Jewell K, Rojiani M and Gray V:
The receptor for the basement membrane glycoprotein entactin is the
integrin alpha 3/beta 1. J Biol Chem. 267:18908–18914.
1992.PubMed/NCBI
|
|
15
|
Dong LJ, Hsieh JC and Chung AE: Two
distinct cell attachment sites in entactin are revealed by amino
acid substitutions and deletion of the RGD sequence in the
cysteine-rich epidermal growth factor repeat 2. J Biol Chem.
270:15838–15843. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yi XY, Wayner EA, Kim Y and Fish AJ:
Adhesion of cultured human kidney mesangial cells to native
entactin: Role of integrin receptors. Cell Adhes Commun. 5:237–248.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Rokavec M, Bouznad N and Hermeking H:
Paracrine induction of epithelial-mesenchymal transition between
colorectal cancer cells and its suppression by a
p53/miR-192/215/NID1 axis. Cell Mol Gastroenterol Hepatol.
7:783–802. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhou Y, Zhu Y, Fan X, Zhang C, Wang Y,
Zhang L, Zhang H, Wen T, Zhang K, Huo X, et al: NID1, a new
regulator of EMT required for metastasis and chemoresistance of
ovarian cancer cells. Oncotarget. 8:33110–33121. 2017. View Article : Google Scholar : PubMed/NCBI
|