Introduction
Colorectal cancer (CRC) is a common malignant tumor
of the digestive system that seriously endangers human lives
(1). CRC is the second- and
third-most common cancer in women and men, respectively. In 2012,
614,000 women (9.2% of all new cancer cases) and 746,000 men (10.0%
of new cancer cases) were diagnosed with colorectal cancer
worldwide (2). Incidence varies
geographically, with the highest incidence in Australia and New
Zealand, whereas Western Africa has the lowest incidence (2). In total, 40–50% of patients with CRC
develop tumor relapse, despite improvements in the diagnostic and
therapeutic methods, leading to a poor prognosis (3). There is a need to clarify the molecular
mechanisms underlying the occurrence and metastasis of CRC.
Meanwhile, developing effective biomarkers and improving
therapeutic strategies for CRC are the focuses of clinical
research.
Long non-coding RNAs (lncRNAs) are transcripts
longer than 200 nucleotides that cannot be translated into proteins
(4); they are involved in the
regulation of transcriptional, post-transcriptional and epigenetic
cell events (5,6). Growing evidence has identified that
lncRNAs participate in various biological processes, including
embryonic stem cells development, helper T-cell differentiation,
autophagy, myocardial infarction, cellular senescence, apoptosis
and metastasis of cancer cells, and chemotherapeutic resistance
(7,8). In addition, dysregulated lncRNAs are
closely linked to tumor relapse and poor prognosis (9). To date, several lncRNAs have been
revealed to be abnormally expressed in CRC profiling, such as
CCAT1, HOTAIR and H19 (10–12). The present study aimed to identify
novel lncRNAs that were dysregulated in the development of CRC. To
comprehensively characterize those aberrantly expressed lncRNAs, we
analyzed CRC and normal tissue RNA sequencing data downloaded from
The Cancer Genome Atlas (TCGA). In order to identify CRC-associated
lncRNAs, the literature was searched to identify a novel lncRNA
named LINC00565 has already been discovered in gastric cancer
(13). LINC00565 spans 2,478 bp
length and is located on chromosome 13, which is significantly
upregulated in gastric cancer (GC) tissues (13). LINC00565 drives the progression of
ovarian cancer by targeting GAS6 (14). In addition, LINC00565 promotes
proliferation and inhibits apoptosis of GC by targeting
miR-665/AKT3 axis. however, its role in CRC is still unclear
(13).
Enhancer of zeste homolog 2 (EZH2) is a histone
methyltransferase that functions as the catalytic subunit of the
polycomb repressive complex 2 (PRC2) (15). PRC2 methylates lysine 27 on histone 3
(H3K27), resulting in the transcriptional silencing of target
genes. EZH2 is responsible for methylating lysine 27 of histone 3
to generate H3K27me3, a histone mark associated with a more
condensed chromatin and transcriptional gene repression (15). A previous study suggested that
silencing EZH2 can induce senescence of tumor cells, including
breast cancer, ovarian cancer, prostate cancer and non-Hodgkin
lymphoma (16). In CRC tissues, EZH2
is highly expressed and its expression level is known to be
associated with the prognosis of CRC (17). The binding between EZH2 and lncRNAs
triggers the specific binding of lncRNAs to proteins, thus
regulating protein activity and function (18,19).
Recent studies have reported that lncRNAs recruit polycomb-group
proteins to target genes (18,19). A
total of 20% of all human lncRNAs have been shown to physically
associate with PRC2, suggesting that lncRNAs may have a general
role in recruiting polycomb-group proteins to their target genes
(18,19). At present, to the best of our
knowledge, no studies have shown whether LINC00565 can interact
with EZH2 to influence the occurrence and development of CRC.
Therefore, the present study aimed to explore the role of LINC00565
in the malignant progression of CRC and its interaction with EZH2,
in order to provide new strategies for the clinical diagnosis and
treatment of CRC.
Patients and methods
Samples
Samples of CRC were collected from 42
patients undergoing radical resection between May 2017 and May 2019
in The Affiliated Jiangyin Hospital of Southeast University Medical
College (Jiangyin, China). There were 31 males and 11 females with
an average age of (52.11±8.99) years, and the patient age ranged
from 37–75 years old. The inclusion criteria used were as follows:
i) Diagnosis of patients based on the diagnostic criteria for CRC;
and ii) Patients having had no prior CRC treatment. The exclusion
criteria were as follows: i) Patients undergoing retreatment after
cancer recurrence; and ii) Patients with other malignant tumors.
The tumor node metastasis (TNM) stage of CRC was based on the
American Joint Committee on Cancer CRC 7th edition TNM stage
(20). Histology grade was
determined based on a previous report (2). Normal tissues resected ≥5 cm away from
the edge of CRC tissues were also collected, and these were
pathologically confirmed to lack tumor invasive tissue. None of the
recruited patients had undertaken CRC treatment. Intraoperatively
collected tissue samples were immediately placed in liquid nitrogen
and stored at −80°C before use. The present study was approved by
the Ethics Committee of The Affiliated Jiangyin Hospital of
Southeast University Medical College (approval number
JYH-17-03-11-932), and written informed consent was obtained from
each subject. The clinical data for the patients is shown in
Table SI.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from the tissues and cells
using the TRIzol kit (Invitrogen; Thermo Fisher Scientific Inc.),
respectively, followed by measurement of RNA concentration using an
ultraviolet spectrophotometer (Hitachi Ltd.). The complementary
Deoxyribose Nucleic Acid (cDNA) was synthesized according to the
instructions of the PrimeScript™ RT MasterMix kit (Invitrogen;
Thermo Fisher Scientific Inc.). qPCR was subsequently conducted
following the recommendations of SYBR Premix Ex Taq™ (Takara Bio,
Inc.). The following primer sequences were used for qPCR: GAPDH
forward, 5′-CACCCACTCCTCCACCTTTG-3′ and reverse,
5′-CCACCACCCTGTTGCTGTAG-3′; LINC00565 forward,
5′-GGCCTGAGCATTGCATAACG-3′ and reverse, 5′-CTGATGGAGCGACCGTCTAC-3′;
and EZH2 forward, 5′-TTGTTGGCGGAAGCGTGTAAAATC-3′ and reverse,
5′-TCCCTAGTCCCGCGCAATGAGC-3′; and U6 forward,
5′-GCGCGTCGTGAAGCGTTC-3′ and reverse, 5′-GTGCAGGGTCCGAGGT-3′. GAPDH
was served as the internal control. The thermocycling conditions
included pre-denaturation at 94°C for 5 min, followed by 40 cycles
of denaturation at 94°C for 30 sec, annealing at 55°C for 30 sec
and extension at 72°C for 90 sec. Gene expression levels were
quantitatively analyzed using the 2−ΔΔCq method
(21).
Cell culture
CRC cell lines (HCT116, SW480, HT-29 and LoVo) and
the normal human colonic epithelial cell line (HCoEpiC) (cat. no.
BSC-5005479458-01) were purchased from the American Type Culture
Collection. Cells were cultured in Dulbecco's modified Eagle's
medium (DMEM; Gibco; Thermo Fisher Scientific, Inc.) containing 10%
fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.) and
1% penicillin in a humidified incubator with 5% CO2 at
37°C. Medium was replaced every 24 h. Cell passage was performed
using 0.25% trypsin when cell density was >70%.
Cell transfection
CRC cell lines and HCoEpiC were resuspended in
antibiotic-free DMEM and re-seeded into 6-well plates at a density
of 3×105 cells/well. CRC cell lines and HCoEpiC were
incubated for 18–24 h at 20°C and transfected using a
Lipofectamine® 2000 kit (Sigma-Aldrich; Merck KGaA).
Cell transfection was performed at 20°C for 48 h. Transfection
efficiency was assessed by RT-qPCR. Liposomes, si-NC (empty
vector), si-LINC00565 and pcDNA-EZH2 overexpression plasmid and
pcDNA-NC (negative control) were all purchased from Shanghai
Genechem Co. Ltd. Sequences of LINC00565 small interfering RNAs
(siRNAs) were as follows: si-LINC00565 1, GCCTCCTGGTTTATAGCATGA;
si-LINC00565 2, GGAGGACGCATCAATCCTTCT; and si-LINC00565 3,
GCTTTGCGAAGACTTTCTTCT. siRNA at the concentration of 50 nM was
added to each well and then incubated for 48 h following which
subsequent experimentation was performed.
Cell Counting Kit-8 (CCK-8) assay
Transfected CRC cell lines and HCoEpiC were cultured
in 96-well plates at a concentration of 2×103
cells/well. Six replicates were set in two groups (si-NC and
si-LINC00565). At 6, 24, 48, 72 and 96 h, 10 µl CCK-8 solution
(Dojindo Molecular Technologies, Inc.) was added per well and
incubated for 4 h. The CCK-8 was used according to the
manufacturer's protocol. Optical density at 450 nm was measured
using a microplate reader.
Transwell migration assay
Transfected cells were prepared as a suspension of
1×109 cells/ml in antibiotic-free DMEM. Next, 200 µl of
this cell suspension was added to the top of Transwell chambers
(EMD Millipore), which were pre-inserted in a 24-well plate. In the
bottom chambers, 600 µl of antibiotic-free DMEM containing 10% FBS
was added. The difference in serum concentration was the inductive
factor for cell chemotaxis. After incubation for 48 h at 37°C, the
invasive cells were fixed in 4% paraformaldehyde, dyed with crystal
violet for 2 min at 37°C and counted using a light microscope
(BX-42; Olympus Corporation). Penetrating cells were counted in ten
randomly selected fields per sample (magnification, ×10). Transwell
migration assay was conducted following the same procedures except
for Matrigel pre-coating (1 h at room temperature).
Chromatin fractionation
A total of 1×106 cells in the bottle were
fully lysed in 200 µl of lysis buffer (cat. no. QC25-05099;
Qincheng Biotech Ltd.) and centrifuged at 4°C, 4,500 × g for 5 min
to separate the cellular components. The supernatant contained the
cytoplasmic fraction. Subsequently, the pellet was incubated in
buffer SK and absolute ethanol. Nuclear RNAs were obtained by
column (Invitrogen; Thermo Fisher Scientific, Inc.) centrifugation
at 4°C, 4,500 × g for 10 min. Transfection efficiency was assessed
by RT-qPCR. Liposomes, si-NC (empty vector), si-LINC00565 and
pcDNA-EZH2 overexpression plasmid were all purchased from Shanghai
Genechem Co. Ltd. Cytoplasmic and nuclear RNA levels were then
determined by RT-qPCR.
RNA immunoprecipitation (RIP)
CRC cell lines and HCoEpiC were treated following
the manufacturer's instructions of Millipore Magna RIP RNA Binding
Protein Immunoprecipitation kit (EMD Millipore). Cell debris was
removed from 140 µl of HepG2 2.2.15 supernatant or patient plasma
by high-speed centrifugation (20,800 × g at 4°C for 5 min). Samples
were subsequently precleared by incubation with 50 µl of Protein G
MicroBeads (Protein G-Agarose; cat. no. SC-2003; Santa Cruz
Biotechnology Inc.) at room temperature for 2 h. The samples
extracted (10 µl) was used as input control. After application on a
µMACS column, the flow-through pool was incubated with 2 µg of
rabbit monoclonal enhancer of zeste homolog 2 (EZH2) antibody
(1:500; cat. no. ab191080; Abcam) or anti-IgG antibody (1:2,000;
cat. no. ab6721; Abcam) at 4°C for 6 h. Following application on a
new µMACS column and washing with phosphate-buffered saline,
bead-bound fractions were eluted from the columns by using lysis
buffer (Nanjing Jiancheng Bioengineering Institute) for RNA
isolation. A protein RNA complex was captured and digested with 0.5
mg/ml proteinase K containing 0.1% SDS (Nanjing Jiancheng Bittech
Company) to extract RNA. The magnetic beads were repeatedly washed
with RIP washing buffer to remove non-specific adsorption as much
as possible. Finally, the extracted RNA was subjected to mRNA level
determination using RT-qPCR.
Statistical analysis
SPSS 19.0 software (IBM Corp.) was used for data
analysis. Data were expressed as the mean ± standard deviation.
Each set of experiments was repeated three times. The diagnostic
value of LINC00565 in CRC was assessed by depicting receiver
operating characteristic (ROC) curves. Pearson's correlation test
was conducted to analyze the correlation between LINC00565 and EZH2
expression. Comparison of pairwise differences between groups was
performed using Student's t-test. Comparison between multiple
groups was performed using one-way ANOVA followed by the post hoc
Bonferroni test. P<0.05 was considered to indicate a
statistically significant difference.
Results
LINC00565 is upregulated in CRC
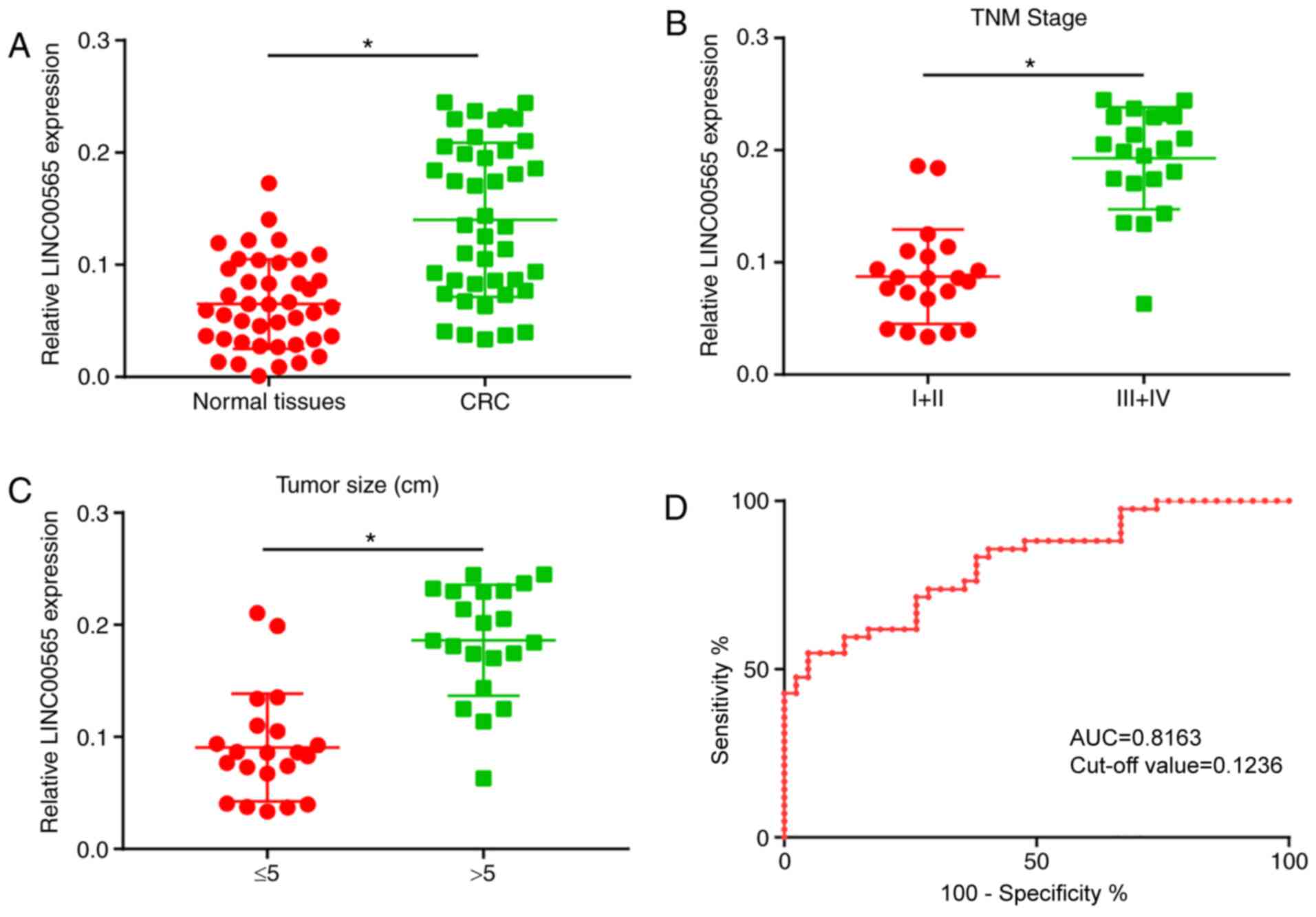
Firstly, the LINC00565 level in CRC tissues compared
with normal tissues was assessed. As shown by the RT-qPCR results,
LINC00565 was upregulated in CRC tissues compared with that in
normal tissues (Fig. 1A). In
addition, the classification of CRC tissues based on
Tumor-Node-Metastasis (TNM) staging and tumor size, indicated that
CRC tissues from more advanced TNM stages, such as stage III+IV,
exhibited a higher level of LINC00565 than those from stage I+II
tissues (Fig. 1B). Furthermore, CRC
tissues with large tumor sizes (>5 cm) expressed a higher
abundance of LINC00565 compared with those with small lesions
(Fig. 1C). These results suggested
that LINC00565 may be involved in the progression of CRC as an
oncogene. To confirm the diagnostic value of LINC00565 in CRC, ROC
curves were depicted to determine the area under the curve (AUC).
The results of the ROC curves demonstrated that LINC00565 has
diagnostic value in CRC (AUC, 0.8163; P<0.05; cut-off value,
0.1236) (Fig. 1D).
LINC00565 promotes proliferative and
migratory abilities in CRC
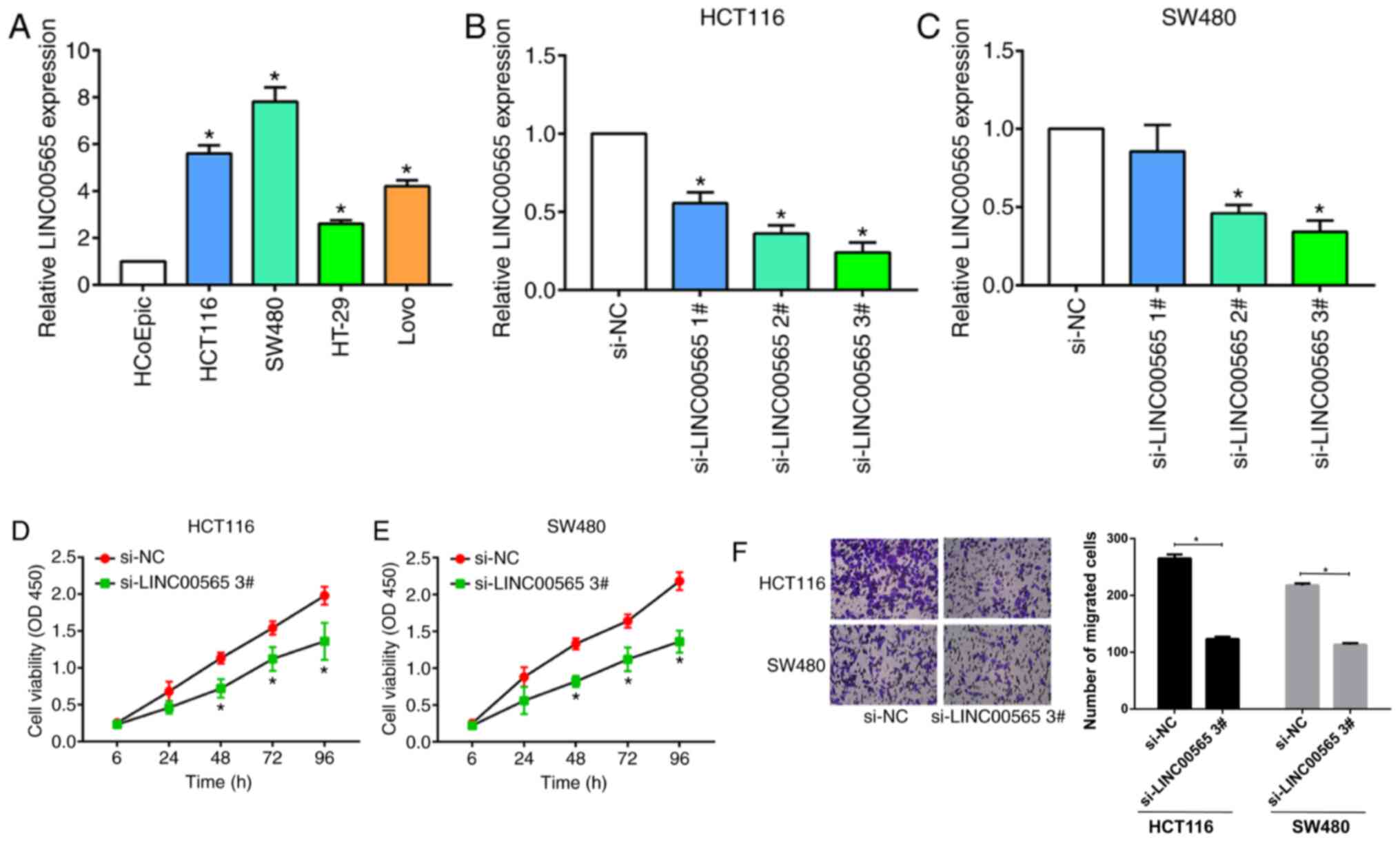
To investigate the effects of LINC00565 on
proliferative and migratory abilities in CRC in vitro
experiments were conducted to clarify the role of LINC00565 in
regulating cell phenotypes of CRC. LINC00565 was highly expressed
in CRC cell lines, especially SW480 and HCT116 cells, when compared
with HCoEpiC control cells (Fig.
2A). In order to silence LINC00565 expression in SW480 and
HCT116 cells, three siRNAs sequences were used. LINC00565
expression was inhibited by the transfection of all of the
sequences (Fig. 2B and C), but
si-LINC00565 3# presented the best efficacy and was selected for
the following experiments. Knockdown of LINC00565 markedly
decreased proliferation in the HCT116 and SW480 cells (Fig. 2D and E). In addition, the migratory
ability of the CRC cells was suppressed by the transfection of
cells with si-LINC00565 3# (Fig.
2F). These results suggested that knockdown of LINC00565 could
statistically inhibit proliferative and migratory potentials in
CRC.
LINC00565 positively regulates the
expression levels of EZH2
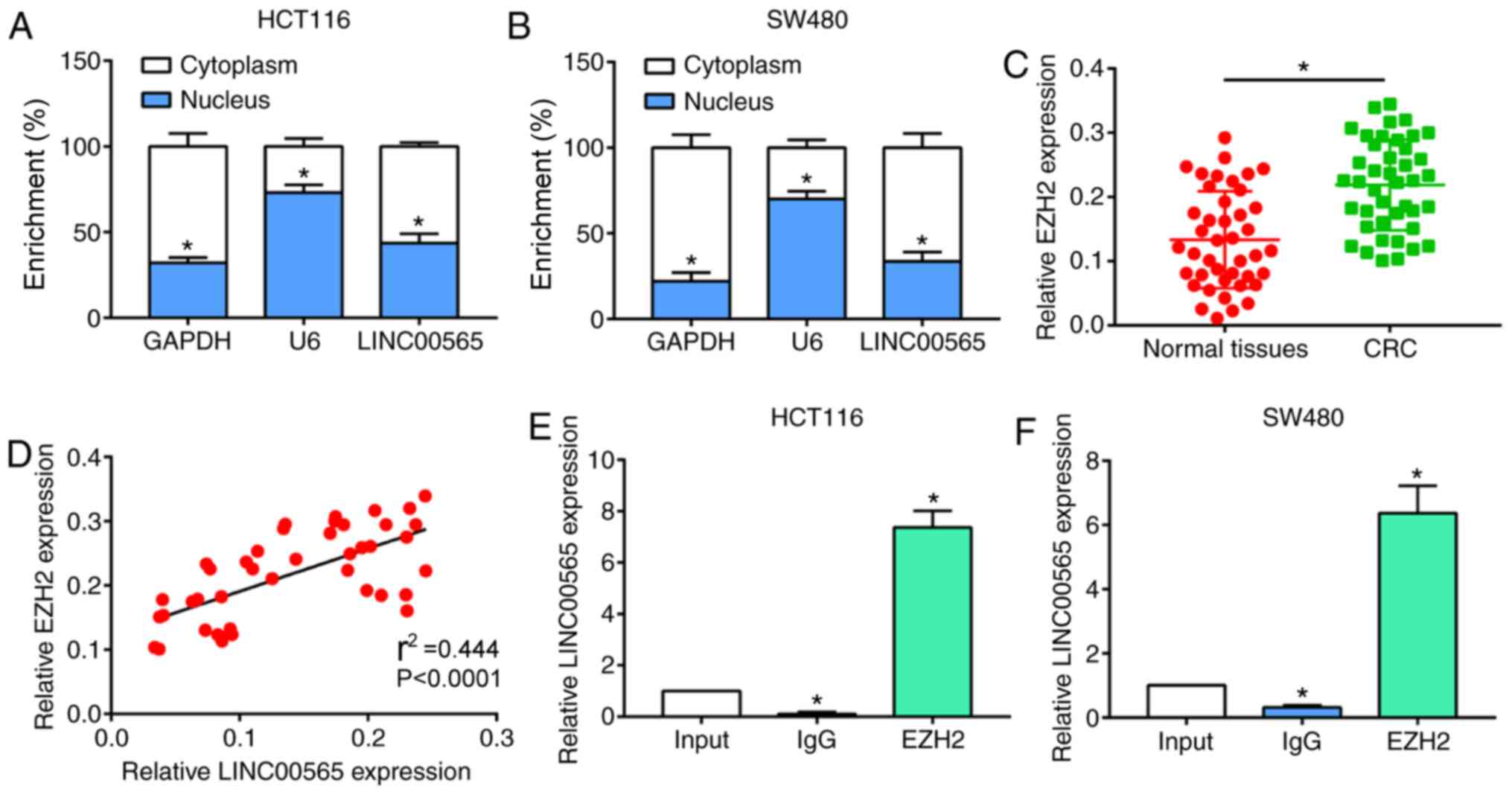
Subsequently, whether EZH2 was involved in the
progression of CRC was investigated. Subcellular distribution of
LINC00565 was analyzed. LINC00565 was mainly expressed in the
cytoplasm of HCT116 and SW480 cells (Fig. 3A and B). EZH2 is one member of PRC2
family (15). Compared with normal
tissues, EZH2 expression levels were upregulated in CRC tissues and
were positively correlated with the expression levels of LINC00565
(Fig. 3C and D). This result
suggested that EZH2 may also be involved in the progression of CRC.
The interaction between LINC00565 and EZH2 was determined by RIP.
Furthermore, LINC00565 was found to be largely enriched in EZH2,
indicating that there is an interaction between LINC00565 and EZH2
(Fig. 3E and F).
Effect of LINC00565 in CRC progression
requires EZH2
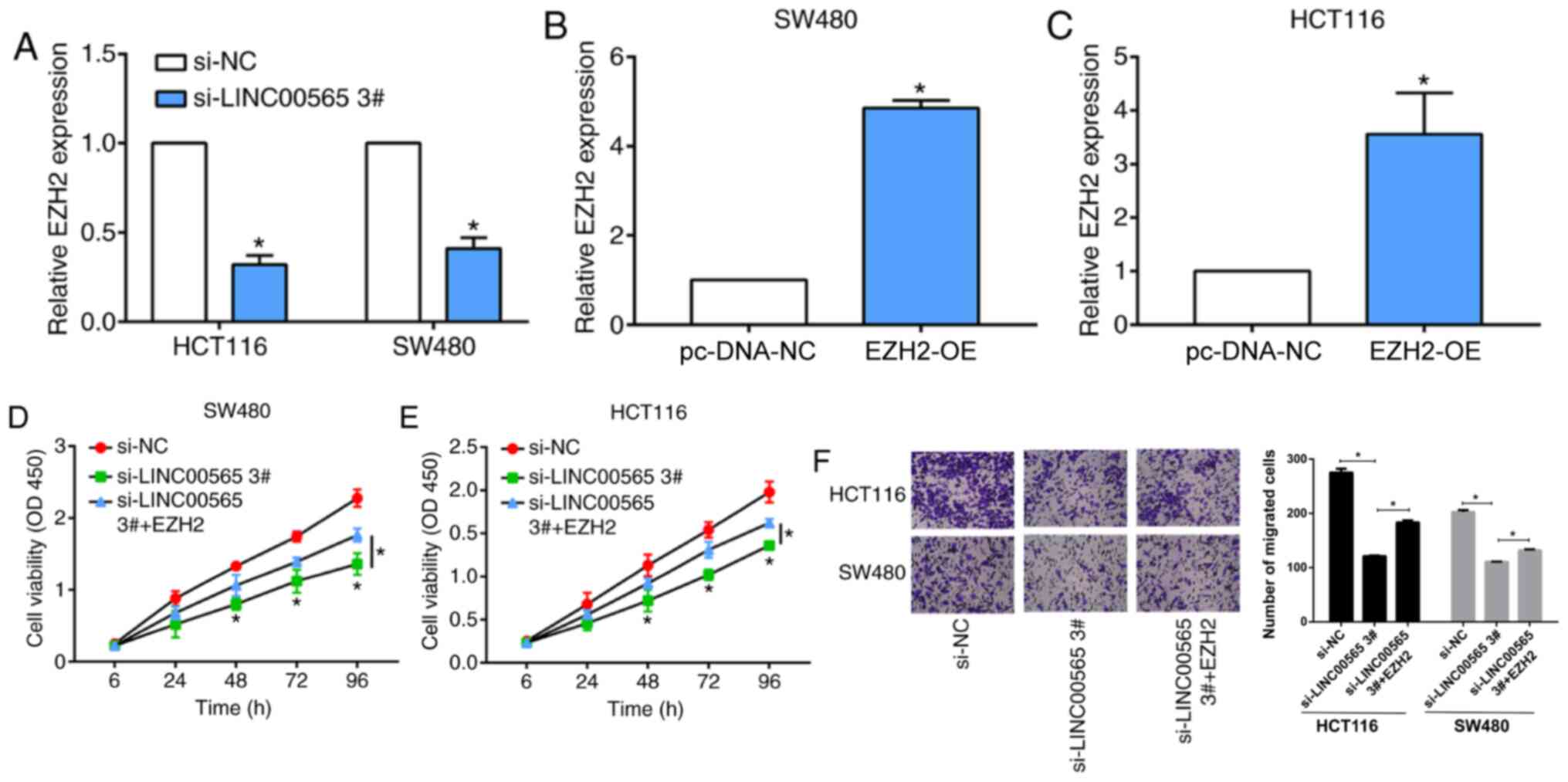
Next, whether the effects of LINC00565 on CRC
progression require EZH2 was studied. EZH2 was found to be
downregulated upon transfection of si-LINC00565 3# in CRC cells
(Fig. 4A). Next, pcDNA-EZH2
overexpression plasmid was used to transfect cells, which resulted
in a marked increase of EZH2 expression levels in SW480 and HCT116
cells (Fig. 4B and C). EZH2
overexpression was found to partially reverse the decreased
proliferation observed with LINC00565-knockdown in CRC cells
(Fig. 4D and E). In addition, the
reduced migratory ability of CRC cells transfected with
si-LINC00565 3# was also attenuated by the overexpression of EZH2
(Fig. 4F). These results indicated
that LINC00565 promoted proliferative and migratory potentials in
CRC by positively regulating EZH2 expression levels.
Discussion
CRC is a prevalent tumor, ranking globally as the
third and second most common tumor in men and women, respectively
(2). In 2012, ~1.4 million
individuals were diagnosed with CRC, causing 693,900 deaths
(22). According to the latest
report, the incidence and mortality rates of CRC in the United
States were in the top three among other cancers in 2012 (1). Great improvements have been made with
regards to the diagnosis and treatment of CRC. However, the
incidence of CRC in Asia still shows an upward trend (22). As a result, early diagnosis and the
development of CRC biomarkers are of clinical significance.
lncRNAs were initially detected from DNA transcripts
by Okazaki et al (23).
Currently, a large number of tumor-associated lncRNAs have been
identified. For example, Kogo et al (11) demonstrated that the lncRNA HOTAIR was
upregulated in CRC tissues when compared with non-tumoral tissues.
Highly expressed HOTAIR also predicted liver metastasis and a poor
prognosis in CRC. KEGG pathway analysis revealed that HOTAIR
induces gene silencing by interacting with the PRC2 family
(including SUZ12, EZH2 and H3K27me3) and LSD1 (9). Thus, it is possible that CRC-associated
lncRNAs can be utilized as diagnostic and therapeutic targets.
However, the regulatory mechanisms of CRC-specific lncRNAs in
cancer progression should be further validated. LINC000565 has been
reported to be significantly increased in ovarian cancer and to be
closely associated with the prognosis of ovarian cancer (14). Notably, in the present study,
LINC00565 was found to play a similar role in CRC, with LINC00565
being upregulated in CRC tissues when compared with normal tissues,
and higher levels of LINC00565 being detected in advanced stages of
CRC and in those with larger tumor sizes. In addition, in
vitro assays showed the promotive effects of LINC00565 on the
proliferative and migratory potential of CRC cells. The results
suggested that LINC00565 might be an oncogene involved in CRC
progression.
EZH2 was also upregulated in CRC tissues in the
present study. Pearson's correlation analysis showed a positive
correlation between the mRNA expression levels of LINC00565 and
EZH2. A number of studies have shown that EZH2 may serve a crucial
role in the occurrence and development of CRC. In CRC tissues, EZH2
is highly expressed and its expression level is known to be
associated with the prognosis of CRC (17). Xie et al (24) found that MALAT1 functioned as a
competing endogenous RNA to promote CRC development and EZH2
expression through sponging miR-363-3p in vitro and in
vivo. As a catalytic subunit of PRC2, EZH2 silences target
genes by catalyzing H3K27me3 and the methylation of lysine at
position 9 of histone H3 through its highly conserved region in
histone methyltransferase (15).
Functionally, EZH2 regulates gene activities at the chromosome
level, which are responsible for mediating cell growth,
differentiation and other cellular functions (25). For example, 20% of all human lncRNAs
have been demonstrated to physically associate with EZH2,
suggesting that lncRNAs may have a general role in recruiting
polycomb-group proteins to their target genes (18,19).
Previous studies have reported that upregulated EZH2 in CRC samples
is closely related to its degree of invasiveness, lymphatic
metastasis and differentiation level. In addition, EZH2 is
considered as an independent factor influencing the prognosis in
CRC (26,27). In the present study, EZH2 was shown
to interact with LINC00565. Importantly, it was able to attenuate
the regulatory effects of LINC00565 on CRC cell phenotypes.
Collectively, LINC00565 and EZH2 synergistically stimulated the
aggravation of CRC, indicating their potential as CRC biomarkers to
monitor disease progression.
In summary, to the best of our knowledge, the
present study is the first to find that LINC00565 is upregulated in
CRC tissues, which stimulates the aggravation of CRC by
upregulating EZH2. The current findings indicated that EZH2 was
involved in the oncogenic role of LINC00565 in CRC. However, the
specific mechanism of LINC00565 regulating EZH2 in the occurrence
and development of CRC remains to be further studied.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XS, TZ, LX and LD designed the study and performed
the experiments, JH, YZ and JZ analyzed the data, XS, LX, YZ, TZ
and LD prepared the manuscript. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
This study was approved by the ethics committee of
The Affiliated Jiangyin Hospital of Southeast University Medical
College (approval. number. JYH-17-03-11-932). Written informed
consent was obtained from the patients and/or guardians.
Patients consent for publication
Patients or their guardians have provided written
informed consent for publication.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
Statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dekker E, Tanis PJ, Vleugels J, Kasi PM
and Wallace MB: Colorectal cancer. Lancet. 394:1467–1480. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Weiser MR, Landmann RG, Kattan MW, Gonen
M, Shia J, Chou J, Paty PB, Guillem JG, Temple LK, Schrag D, et al:
Individualized prediction of colon cancer recurrence using a
nomogram. J Clin Oncol. 26:380–385. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mercer TR and Mattick JS: Structure and
function of long noncoding RNAs in epigenetic regulation. Nat
Struct Mol Biol. 20:300–307. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Deniz E and Erman B: Long noncoding RNA
(lincRNA), a new paradigm in gene expression control. Funct Integr
Genomics. 17:135–143. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Vance KW and Ponting CP: Transcriptional
regulatory functions of nuclear long noncoding RNAs. Trends Genet.
30:348–355. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Huarte M: The emerging role of lncRNAs in
cancer. Nat Med. 21:1253–1261. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shi T, Gao G and Cao Y: Long noncoding
RNAs as novel biomarkers have a promising future in cancer
diagnostics. Dis Markers. 2016:90851952016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yildirim E, Kirby JE, Brown DE, Mercier
FE, Sadreyev RI, Scadden DT and Lee JT: Xist RNA is a potent
suppressor of hematologic cancer in mice. Cell. 152:727–742. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
He X, Tan X, Wang X, Jin H, Liu L, Ma L,
Yu H and Fan Z: C-Myc-activated long noncoding RNA CCAT1 promotes
colon cancer cell proliferation and invasion. Tumour Biol.
35:12181–12188. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kogo R, Shimamura T, Mimori K, Kawahara K,
Imoto S, Sudo T, Tanaka F, Shibata K, Suzuki A, Komune S, et al:
Long noncoding RNA HOTAIR regulates polycomb-dependent chromatin
modification and is associated with poor prognosis in colorectal
cancers. Cancer Res. 71:6320–6326. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liang WC, Fu WM, Wong CW, Wang Y, Wang WM,
Hu GX, Zhang L, Xiao LJ, Wan DC, Zhang JF and Waye MM: The lncRNA
H19 promotes epithelial to mesenchymal transition by functioning as
miRNA sponges in colorectal cancer. Oncotarget. 6:22513–22525.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hu J, Ni G, Mao L, Xue X, Zhang J, Wu W,
Zhang S, Zhao H, Ding L and Wang L: LINC00565 promotes
proliferation and inhibits apoptosis of gastric cancer by targeting
miR-665/AKT3 axis. Onco Targets Ther. 12:7865–7875. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gong M, Luo C, Meng H, Li S, Nie S, Jiang
Y, Wan Y, Li H and Cheng W: Upregulated LINC00565 accelerates
ovarian cancer progression by targeting GAS6. Onco Targets Ther.
12:10011–10022. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li Z, Hou P, Fan D, Dong M, Ma M, Li H,
Yao R, Li Y, Wang G, Geng P, et al: The degradation of EZH2
mediated by lncRNA ANCR attenuated the invasion and metastasis of
breast cancer. Cell Death Differ. 24:59–71. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tsang DP and Cheng AS: Epigenetic
regulation of signaling pathways in cancer: Role of the histone
methyltransferase EZH2. J Gastroenterol Hepatol. 26:19–27. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bremer S, Conradi LC, Mechie NC, Amanzada
A, Mavropoulou E, Kitz J, Ghadimi M, Ellenrieder V, Ströbel P,
Hessmann E, et al: Enhancer of zeste homolog 2 in colorectal cancer
development and progression. Digestion. 6:1–9. 2019. View Article : Google Scholar
|
|
18
|
Hirata H, Hinoda Y, Shahryari V, Deng G,
Nakajima K, Tabatabai ZL, Ishii N and Dahiya R: Long noncoding RNA
MALAT1 promotes aggressive renal cell carcinoma through Ezh2 and
interacts with miR-205. Cancer Res. 75:1322–1331. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cerase A, Smeets D, Tang YA, Gdula M,
Kraus F, Spivakov M, Moindrot B, Leleu M, Tattermusch A, Demmerle
J, et al: Spatial separation of xist RNA and polycomb proteins
revealed by superresolution microscopy. Proc Natl Acad Sci USA.
111:2235–2240. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hashiguchi Y, Hase K, Kotake K, Ueno H,
Shinto E, Mochizuki H, Yamamoto J and Sugihara K: Evaluation of the
seventh edition of the tumour, node, metastasis (TNM)
classification for colon cancer in two nationwide registries of the
United States and Japan. Colorectal Dis. 14:1065–1074. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Okazaki Y, Furuno M, Kasukawa T, Adachi J,
Bono H, Kondo S, Nikaido I, Osato N, Saito R, Suzuki H, et al:
Analysis of the mouse transcriptome based on functional annotation
of 60,770 full-length cDNAs. Nature. 420:563–573. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xie JJ, Li WH, Li X, Ye W and Shao CF:
LncRNA MALAT1 promotes colorectal cancer development by sponging
miR-363-3p to regulate EZH2 expression. J Biol Regul Homeost
Agents. 33:331–343. 2019.PubMed/NCBI
|
|
25
|
Kondo Y, Shen L, Cheng AS, Ahmed S,
Boumber Y, Charo C, Yamochi T, Urano T, Furukawa K, Kwabi-Addo B,
et al: Gene silencing in cancer by histone H3 lysine 27
trimethylation independent of promoter DNA methylation. Nat Genet.
40:741–750. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mimori K, Ogawa K, Okamoto M, Sudo T,
Inoue H and Mori M: Clinical significance of enhancer of zeste
homolog 2 expression in colorectal cancer cases. Eur J Surg Oncol.
31:376–380. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang CG, Ye YJ, Yuan J, Liu FF, Zhang H
and Wang S: EZH2 and STAT6 expression profiles are correlated with
colorectal cancer stage and prognosis. World J Gastroenterol.
16:2421–2427. 2010. View Article : Google Scholar : PubMed/NCBI
|