Introduction
Ovarian cancer ranks 7th among the most common
cancer types globally, and is the 8th leading cause of
cancer-associated mortality in women, as reported in 2013 (1), with a 5-year overall survival rate
<45% (2). Ovarian cancer
frequently occurs during perimenopause and has the highest
mortality rate among different types of gynecological cancer, due
to unclear manifestations and lack of effective diagnosis in the
early stages (3). The following long
non-coding RNAs (lncRNAs), Human Satellite II (4), Antisense Non-coding RNA in the INK4
Locus (5) and Colon Cancer
Associated Transcript 1 (6,7) have been demonstrated to participate in
the occurrence and progression of ovarian cancer, which suggests
their role as biomarkers for early cancer diagnosis. However, this
knowledge does not decrease the threat of ovarian cancer to female
health. Thus, the molecular mechanisms underlying ovarian cancer
progression need to be further investigated to determine novel
effective therapeutic targets and prolong the survival of patients
with ovarian cancer.
lncRNAs containing >200 nucleotides are a type of
novel transcript that cannot code proteins (8,9). lncRNAs
have been reported to influence crucial cell functions, including
histone modification, chromatin rearrangement, regulation of gene
expression and alternative splicing gene modification (10–12).
They also participate in dosage compensation (13), genomic imprinting (14), cell differentiation, organogenesis
and tumor formation (15–17). A number of lncRNAs influence the
onset and progression of ovarian cancer and may serve as
therapeutic targets and markers for its diagnosis and prognosis
(18–20). Small nucleolar RNA host gene 9
(SNHG9) is located on chromosome 16p13.3, and while its involvement
in the progression of ovarian cancer remains unclear, it is a novel
prognostic marker for pancreatic cancer (21).
Yu et al (22)
reported that CRY2 suppresses proliferation and migration in
osteosarcoma cells. Fang et al (23) demonstrated that CRY2 degradation
influences chemoresistance in colorectal cancer. Furthermore,
Tokunaga et al (24)
suggested that CRY2 may serve a role in the progression of ovarian
cancer. Hence, CRY2 was selected for further analyses in the
present study.
The present study aimed to determine the biological
role and potential molecular mechanism of SNHG9 in ovarian cancer.
SNHG9 expression levels in ovarian cancer cell lines and tissues
were analyzed by performing in vitro cell function
assays.
Materials and methods
Ethical compliance
The present study was approved by the Institutional
Ethics Committee of The Second People's Hospital of Taizhou City
(Taizhou, China) and written informed consent was obtained from all
patients prior to the study start.
Human tissue specimens
A total of 25 pairs of ovarian cancer and
paracancerous normal tissue samples (mean age, 53 years; age range,
41–69 years) were collected from patients undergoing surgery at The
Second People's Hospital of Taizhou City (Taizhou, China). Patients
treated with radiotherapy and chemotherapy prior to surgery were
excluded from the study. Fresh tissues were collected during
surgery, frozen in liquid nitrogen within 5 min following
resection, and stored at −80°C until further experimentation. The
clinicopathological characteristics of patients with ovarian cancer
are presented in Table I.
 | Table I.Association between SNHG9 expression
and clinicopathological characteristics of patients with ovarian
cancer. |
Table I.
Association between SNHG9 expression
and clinicopathological characteristics of patients with ovarian
cancer.
| Characteristic | Low expression,
n=13 | High expression,
n=12 | P-value |
|---|
| Age, years |
|
|
|
|
<50 | 6 | 4 |
|
| ≥50 | 7 | 8 | 0.6882 |
| Histological
subtype |
|
|
|
|
Serous | 10 | 8 |
|
|
Others | 3 | 4 | 0.6728 |
| FIGO stage |
|
|
|
|
I–II | 1 | 6 |
|
|
III–IV | 12 | 6 | 0.0302a |
| Residual tumor
diameter, cm |
|
|
|
|
<1 | 6 | 7 |
|
| ≥1 | 7 | 5 | 0.6951 |
| Lymph node
metastasis |
|
|
|
|
Absent | 10 | 4 |
|
|
Present | 3 | 8 | 0.0472a |
Cell lines and cell culture
A total of three ovarian cancer cell lines (SKOV3,
OVCAR-3 and A2780) and a normal ovarian cell line (IOSE-80) were
purchased from Shanghai Institute for Biological Science and
cultured in DMEM (HyClone; GE Healthcare Life Sciences)
supplemented with 10% FBS and 1% penicillin/streptomycin (Gibco;
Thermo Fisher Scientific Inc.) at 37°C with 5% CO2.
Reverse transcription-quantitative
(RT-q)PCR
microRNAs (miRNAs/miR) were harvested from the
cultured cells and tumor tissues using a mirVana™ miRNA Isolation
kit (Ambion; Thermo Fisher Scientific, Inc.). miR-214-5p expression
levels were measured using the SYBR Primescript miRNA RT-qPCR kit
and SYBR1 Green I (Toyobo Life Science), according to
manufacturer's protocol. Total RNA was extracted from the ovarian
cancer cells and tissues using TRIzol® reagent (Thermo
Fisher Scientific, Inc.) and reverse transcribed into cDNA using a
RT kit (Takara Biotechnology Co., Ltd.) according to the
manufacturer's protocols. The following primer sequences (Takara
Biotechnology Co., Ltd.) were used for qPCR: SNHG9 forward,
5′-GACTGCAGACCCCTAACCTT-3′ and reverse, 5′-ACCCGCATGCAGTGAGTTA-3′;
cryptochrome circadian regulator 2 (CRY2) forward,
5′-TCCCAAGGCTGTTCAAGGAAT-3′ and reverse,
5′-TGCATCCCGTTCTTTCCCAAA-3′; GAPDH forward,
5′-GTCAACGGATTTGGTCTGTATT-3′ and reverse,
5′-AGTCTTCTGGGTGGCAGTGAT-3′; miR-214-5p forward,
5′-ACACTCCAGCTGGGCGTGTCGTTCACATCT-3′ and reverse,
5′-CUACAAAGGGAAGCGACAGGCA-3′ and U6 forward,
5′-CTCGCTTCGGCAGCACA-3′ and reverse, 5′-AACGCTTCACGAATTTGCGT-3′.
The following thermocycling conditions were used: Initial
denaturation step (95°C for 20 sec), followed by 40 cycles of
denaturation (95°C for 3 sec) and annealing (60°C for 30 sec).
miR-214-5p and SNHG9 mRNA expression levels were measured using the
2−ΔΔCq method (25) and
normalized to the internal reference genes U6 and GAPDH,
respectively.
Cell transfection
Short hairpin (sh)RNAs synthesized against SNHG9
(sh-SNHG9) and matched negative control sequences (sh-NC) were
subcloned into the pGPU6/GFP/Neo vector (Shanghai GenePharma Co.,
Ltd.). Subsequently, SKOV3 cells were transfected with 100 pmol
sh-SNHG9 or sh-NC using Lipofectamine® 2000 (Thermo
Fisher Scientific Inc.), according to the manufacturer's protocol.
Stable cell clones were selected using 0.5 mg/ml of G418
(Sigma-Aldrich; Merck KGaA). Thereafter, the cells were transfected
with 0.1 nmol miR-214-5p mimics, negative control (NC) mimics
(miR-NC), miR-214-5p inhibitors (anti-miR-214-5p) or anti-miR-NC
(Guangzhou RiboBio Co., Ltd.) using Lipofectamine® 2000,
according to the manufacturer's protocol. The sequences of
miR-214-5p mimics are as follows: Sense,
5′-UGCCUGUCUACACUUGCUGUGC-3′ and antisense,
5′-ACAGCAAGUGUAGACAGGCAUU-3′. The sequences of miR-214-5p inhibitor
are as follows: Sense, 5′-GCACAGCAAGUGUAGACAGGCA-3′. Subsequent
experimentation was performed 48 h post-transfection.
Cytotoxicity assay
A total of 5×103 of the transfected SKOV3
cells were seeded into a 96-well plate and incubated with CCK-8
reagent (Beyotime Institute of Biotechnology) for 2 h at 37°C in
the dark, at 24, 48, 72 and 96 h post-transfection, according to
the manufacturer's protocol. Cell proliferation was analyzed as a
wavelength of 450 nm using a Benchmark Plus™ Microplate
spectrometer (Bio-Rad Laboratories, Inc.). All experiments were
performed in triplicate.
Colony formation assay
SKOV3 cells that demonstrated a stable decrease in
SNHG9 expression were seeded into a 6-well plate at a density of
1×103 cells/well and incubated at 37°C with 5%
CO2 for 2 weeks. Cells were fixed with 96% ethanol for
10 min and subsequently stained with 1% crystal violet for 5 min at
room temperature (both from Sigma-Aldrich; Merck KGaA). Cells were
counted and the visible colonies were imaged using an inverted
light microscope (magnification, ×10; Olympus Corporation).
Cell cycle distribution analysis
The transfected SKOV3 cells were fixed with 70%
ethanol (Beyotime Institute of Biotechnology) overnight at 4°C and
rinsed twice with PBS, prior to incubation with 0.1 mg/ml of
propidium iodide and 1 mg/ml of RNase A (Sigma-Aldrich; Merck KGaA)
at room temperature for 30 min. Cell cycle distribution was
assessed using a FACScan flow cytometer (BD Biosciences) and cell
percentages in the G0/G1, S and
G2/M phases were determined using FlowJo software
(version 7.6.1; Tree Star, Inc.).
Wound healing assay
The transfected SKOV3 cells were seeded into a
6-well plate and incubated with DMEM at 37°C with 5% CO2
until they reached ~100% confluence. Cells were rinsed three times
with PBS and serum-starved for 24 h. Subsequently, the cell
monolayer was scratched using a 10 µl pipette tip and the migratory
cells were imaged at 0 and 24 h using an inverted light microscope
(magnification, ×10; Olympus Corporation). AxioVision software
(version 4.7; Carl Zeiss AG) was used to assess wound healing.
Transwell and Matrigel assays
Transwell chambers (Corning Inc.) coated with
Matrigel at 37°C for 30 min (BD Biosciences) were applied to assess
cell invasion. A total of 2.0×104 of the transfected
SKOV3 cells were plated in the upper chambers of Transwell plates
in DMEM without FBS, while the lower Transwell chamber was
incubated in DMEM supplemented with 10% FBS as a chemo-attractant.
Following incubation for 48 h at 37°C, the non-invasive cells in
the upper chamber were removed using cotton swabs, and the invasive
cells in the lower chamber were fixed in 4% paraformaldehyde for 20
min and subsequently stained with 1% crystal violet for 15 min at
room temperature. Stained cells were counted in five
randomly-selected fields using an inverted light microscope
(magnification, ×20; Olympus Corporation).
Subcellular fractionation
location
The location of SNHG9 in SKOV3 cells was determined
using the PARIS kit (Thermo Fisher Scientific, Inc.), whereby the
nuclear and cytoplasmic fractions were isolated, according to the
manufacturer's protocol. SNHG9 expression levels were quantified
via RT-qPCR analysis (as above), and normalized to the internal
reference genes GAPDH (cytoplasmic control) and U6 (nuclear
control), respectively.
Dual-luciferase reporter assay
Putative miRNA binding sites on SNHG9 and CRY2
sequences were speculated using miRcode (http://www.mircode.org) and confirmed via the
dual-luciferase assay. Briefly, SNHG9 and CRY2 3′-untranslated
region (UTR) sequences with the predicted wild-type (WT) or mutant
(MUT) miR-214-5p binding sites (Shanghai GenePharma Co., Ltd.) were
subcloned into the following firefly luciferase reporter psiCHECK-2
vectors; SNHG9-WT, SNHG9-MUT, CRY2-3′-UTR-WT and CRY2-3′-UTR-MUT
(Promega Corporation). miR-214-5p mimics or miR-NC mimics, SNHG9-WT
or SNHG9-MUT and CRY2-3′-UTR-WT or CRY2-3′-UTR-MUT reporter
plasmids were co-transfected in SKOV3 cells using
Lipofectamine® 2000 (Beyotime Institute of
Biotechnology). Following incubation at 37°C for 48 h, cells were
collected and firefly and Renilla luciferase activities were
detected using a Dual-luciferase reporter assay system (Promega
Corporation) according to the manufacturer's protocol. Firefly
luciferase activity was normalized to Renilla luciferase
activity.
RNA immunoprecipitation (RIP)
assay
The RIP assay was performed using the EZ-Magna RIP
kit (EMD Millipore), according to the manufacturer's protocol.
SKOV3 cells were lysed using RIP lysis buffer, prior to incubation
with magnetic beads conjugated with anti-Ago2 antibody (1:30; cat.
no. ab186733; Abcam) or anti-IgG antibody (1:150; cat. no. 12-370;
Merck Millipore), which was used as a control. RNAs were purified
using the TRIzol™ Plus RNA Purification kit (cat. no. 12183555,
Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol and detected via RT-qPCR analysis.
Statistical analysis
Statistical analysis was performed using SPSS
software (version 18.0; SPSS, Inc.). Data are presented as the mean
± standard deviation. The paired Student's t-test was performed to
detect the differential expression of SNHG9, miR-214-5p, and
CRY2 between ovarian cancer tissues and paracancerous normal
tissues. The difference was evaluated by unpaired Student's t-test
(other 2-group comparisons) or one-way ANOVA followed by the post
hoc Bonferroni's correction (multigroup comparisons) as
appropriate. The target genes of miR-214-5p were screened using
TargetScan 7.2 (http://www.targetscan.org) and the miRDB database
(http://mirdb.org). Patients were divided into low and
high SNHG9 expression groups based on the median expression in
ovarian cancer tumor tissues (cut-off=0.055), and Kaplan-Meier
survival curves were generated, and overall survival rates were
analyzed via the log-rank test. The association between SNHG9,
miR-214-5p and CRY2 expression were validated by Pearson's
correlation analysis. Association between SNHG9 expression and
clinicopathological characteristics of patients with ovarian cancer
were analyzed using χ2 or Fisher's exact tests.
P<0.05 was considered to indicate a statistically significant
difference. All experiments were performed in triplicate.
Results
SNHG9 is downregulated in ovarian
cancer cell lines and tissues
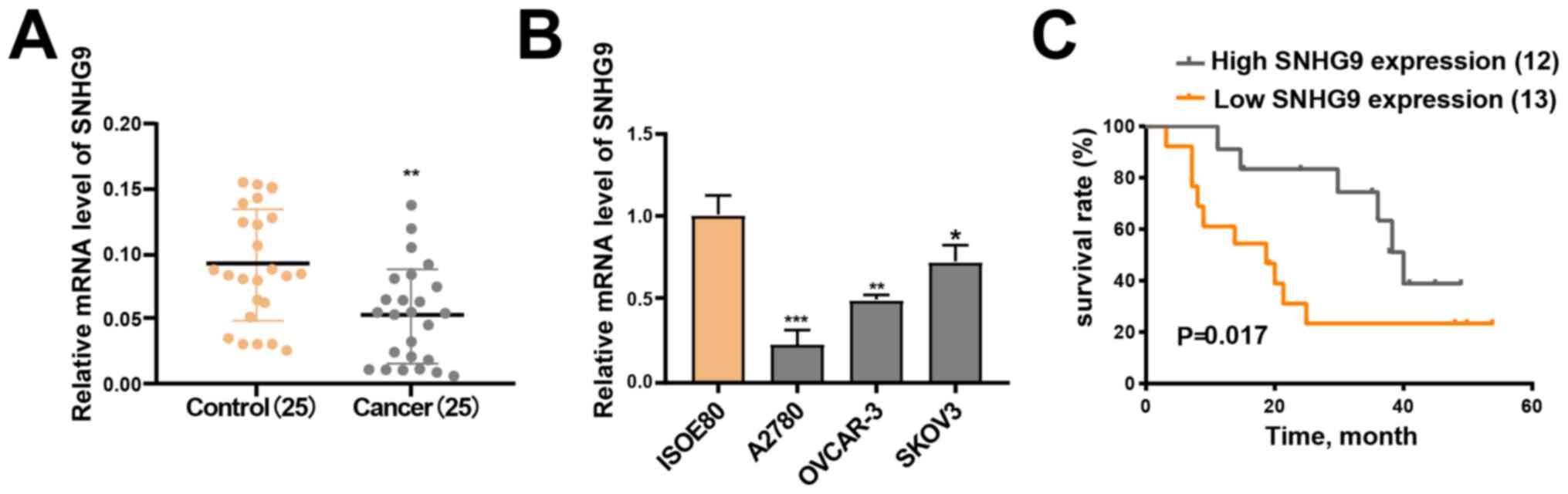
RT-qPCR analysis demonstrated that SNHG9 expression
was significantly downregulated in the 25 ovarian cancer tissue
samples compared with the matched paracancerous normal tissue
samples (P<0.01; Fig. 1A).
Furthermore, low SNHG9 expression was observed across all three
ovarian cancer cell lines (SKOV3, OVCAR-3 and A2780) compared with
IOSE-80 cells (all P<0.05; Fig.
1B), of which SKOV3 demonstrated the highest SNHG9 expression.
Thus, this cell line was chosen for subsequent experimentation.
SNHG9 expression is associated with
clinicopathological characteristics and good prognosis in ovarian
cancer
Analysis between SNHG9 expression and
clinicopathological characteristics revealed that SNHG9 expression
was significantly associated with FIGO stage (26) (P=0.0302) and lymph node metastasis
(P=0.0472). However, no significant associations were observed
between SNHG9 expression and other clinicopathological
characteristics (Table I).
Patients were divided into low and high SNHG9
expression groups based on mean median expression level in ovarian
cancer tumor tissues (cut-off=0.055), in order to determine the
association between SNHG9 expression and the outcomes of ovarian
cancer. The Kaplan-Meier curve demonstrated that patients in the
high SNHG9 expression group had higher survival rates compared with
patients in the low SNHG9 expression group (P=0.017; Fig. 1C). The current results suggest that
SNHG9 may serve as a tumor suppressor gene in ovarian cancer.
SNHG9-knockdown induces migration,
proliferation and invasiveness of ovarian cancer cells
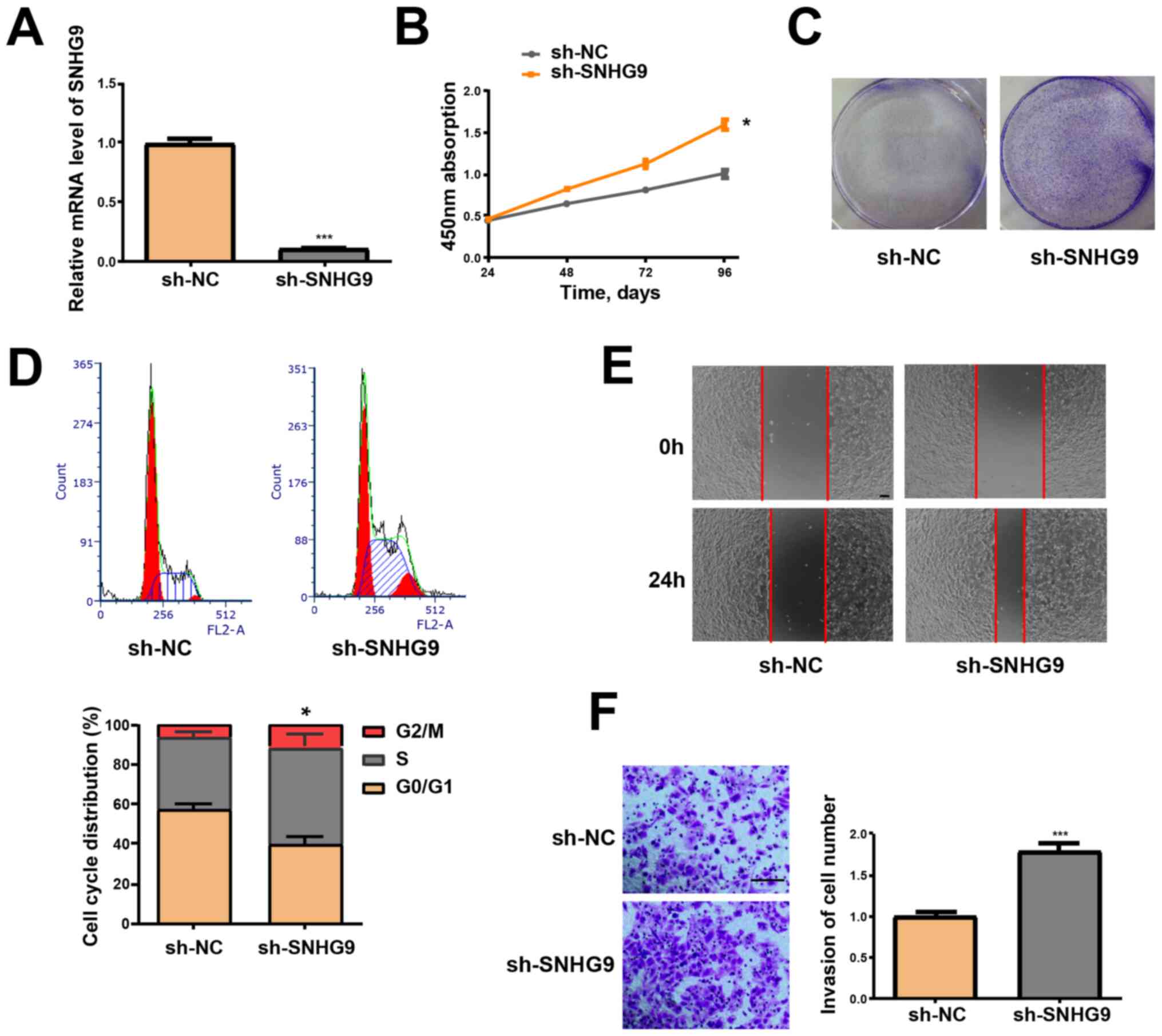
SNHG9-knockdown, via transfection of SKOV3 cells
with sh-SNHG9, was performed to determine the effect of SNHG9 in
tumor formation of ovarian cancer. RT-qPCR analysis demonstrated
that SNHG9 expression was significantly decreased in the sh-SNHG9
group compared with the sh-NC group (P<0.001; Fig. 2A). Furthermore, the CCK-8 assay
revealed that low SNHG9 expression induced the viability of SKOV3
cells (P<0.05; Fig. 2B). Cells in
the sh-SNHG9 group demonstrated a stronger capacity to form
colonies compared with the sh-NC group (Fig. 2C). Flow cytometry analysis
demonstrated that low SNHG9 expression decreased the percentage of
SKOV3 cells in the G1 phase but increased the number of
cells in the S phase (P<0.05, Fig.
2D). The wound healing assay results revealed that
SNHG9-knockdown significantly enhanced the migrative ability of
SKOV3 cells (Fig. 2E). Results from
the Transwell and Matrigel assays verified that low SNHG9
expression significantly induces the invasive ability of SKOV3
cells (P<0.001; Fig. 2F).
miR-214-5p is a direct target of SNHG9
in ovarian cancer cells
lncRNAs function in accordance to their subcellular
distribution (27). Thus, RT-qPCR
was performed to determine SNHG9 expression in the nucleus and
cytoplasm of SKOV3 cells. SNHG9 was demonstrated to be primarily
expressed in the cytoplasm of SKOV3 cells (Fig. 3A). This indicates that SNHG9 may
participate in the development of ovarian cancer via
post-transcriptional regulation, and thus was assumed to serve as a
competing endogenous RNA in the progression of ovarian cancer. This
result was confirmed using miRcode software, which identified the
underlying miRNA target of SNHG9. As the SNHG9 transcript contains
a putative miR-214-5p binding site, miR-214-5p was hypothesized to
be a potential binding target of the gene (Fig. 3B).
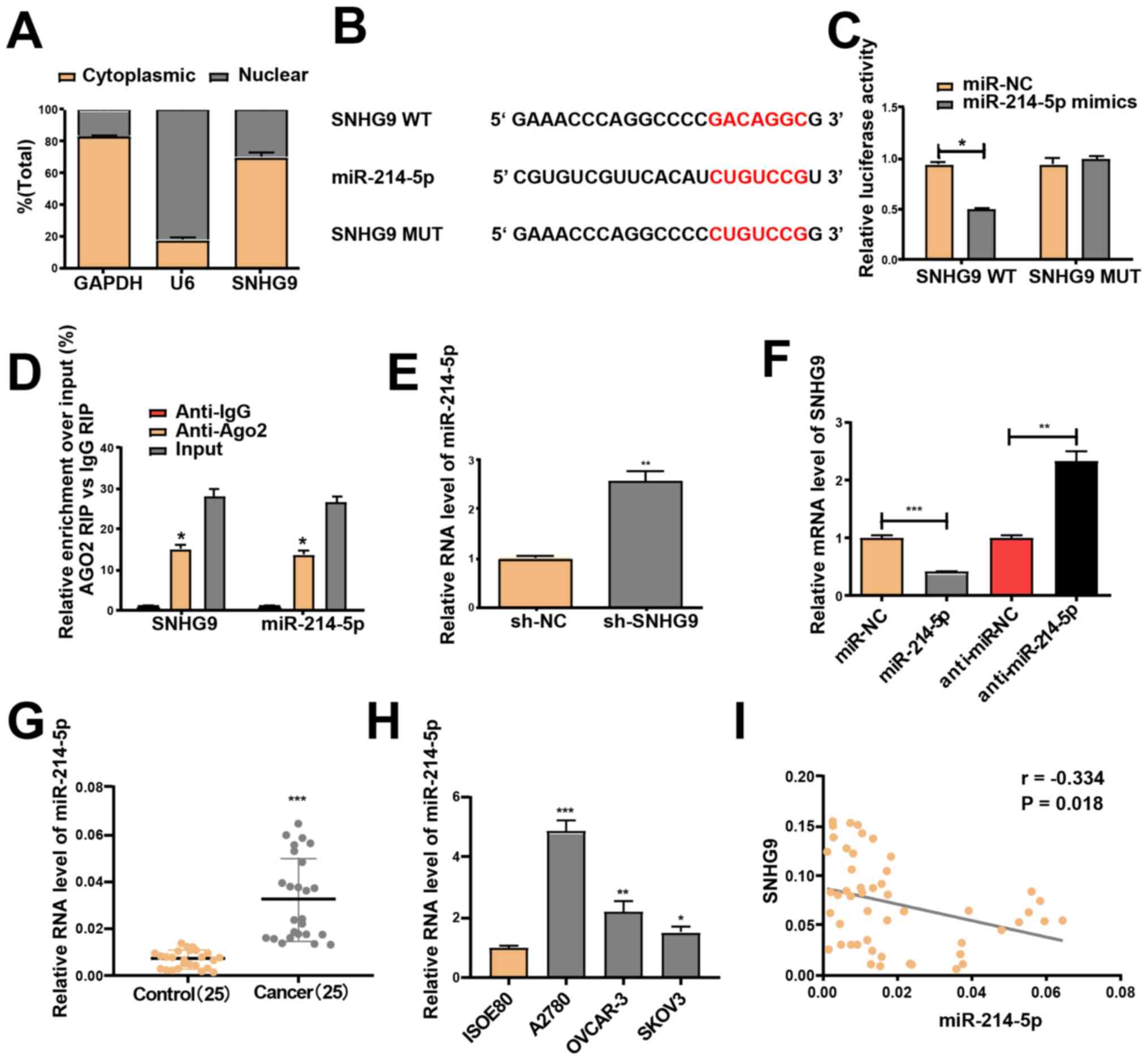 | Figure 3.miR-214-5p is a target of SNHG9. (A)
Reverse transcription-quantitative PCR was performed to evaluate
SNHG9 expression in the nucleus and cytoplasm of SKOV3 cells, with
U6 as the nuclear control and GAPDH as the cytoplasmic control. (B)
Mutant and binding sites between miR-214-5p and SNHG9. (C)
Following co-transfection with SNHG9-WT or SNHG9-MUT reporter
plasmids, and miR-214-5p/miR-NC mimics, SKOV3 cells were subjected
to a dual-luciferase reporter assay. (D) Anti-Ago2 RIP assay with
miR-214-5p mimics demonstrated that miR-214-5p and SNHG9
accumulated in Ago2 precipitates compared with the control (IgG) in
SKOV3 cells. (E) miR-214-5p expression in SKOV3 cells subjected to
transfection with sh-SNHG9 or sh-NC. (F) SNHG9 expression in SKOV3
cells treated with miR-214-5p/miR-NC or
anti-miR-214-5p/anti-miR-NC. (G) miR-214-5p expression in ovarian
cancer tissues and paracancerous normal tissues (n=25). (H)
miR-214-5p expression in ovarian cancer and normal ovarian cell
lines. (I) Pearson's correlation analysis demonstrated a negative
correlation between the mRNA expression levels of SNHG9 and
miR-214-5p in ovarian cancer tissues. All experiments were
performed in triplicate and data are presented as the mean ±
standard deviation. *P<0.05; **P<0.01; ***P<0.001 vs.
anti-igG, sh-NC, control group or ISOE80 group. miR, microRNA;
SNHG9, small nucleolar RNA host gene 9; WT, wild type; MUT, mutant;
NC, negative control; RIP, RNA immunoprecipitation; sh, short
hairpin. |
A Dual-luciferase reporter assay was performed to
determine whether miR-214-5p is a downstream target of SNHG9.
Transfection with miR-214-5p mimics did not change the luciferase
activity of the SNHG9 -MUT but significantly decreased the activity
of the SNHG9 WT in SKOV3 cells (P<0.05, Fig. 3C). miR-214-5p mimics were used for
the anti-Ago2 RIP assay, and the results revealed that SNHG9 and
miR-214-5p were enriched in the immunoprecipitation pulled down by
the anti-Ago2 antibody in SKOV3 cells (Fig. 3D). Furthermore, low SNHG9 levels
prominently elevated miR-214-5p expression in SKOV3 cells (Fig. 3E), and transfection with the
miR-214-5p mimics significantly downregulated SNHG9 expression,
which was reversed by transfection with miR-214-5p inhibitors in
SKOV3 cells (Fig. 3F). Significantly
increased miR-214-5p expression in ovarian cancer cell lines and
tissues, relative to normal samples, was confirmed via RT-qPCR
analysis (Fig. 3G and H). Pearson's
correlation analysis indicated a negative correlation between the
expression levels of miR-214-5p and SNHG9 (r=−0.334; P<0.05;
Fig. 3I).
SNHG9 regulates CRY2 expression via
miR-214-5p
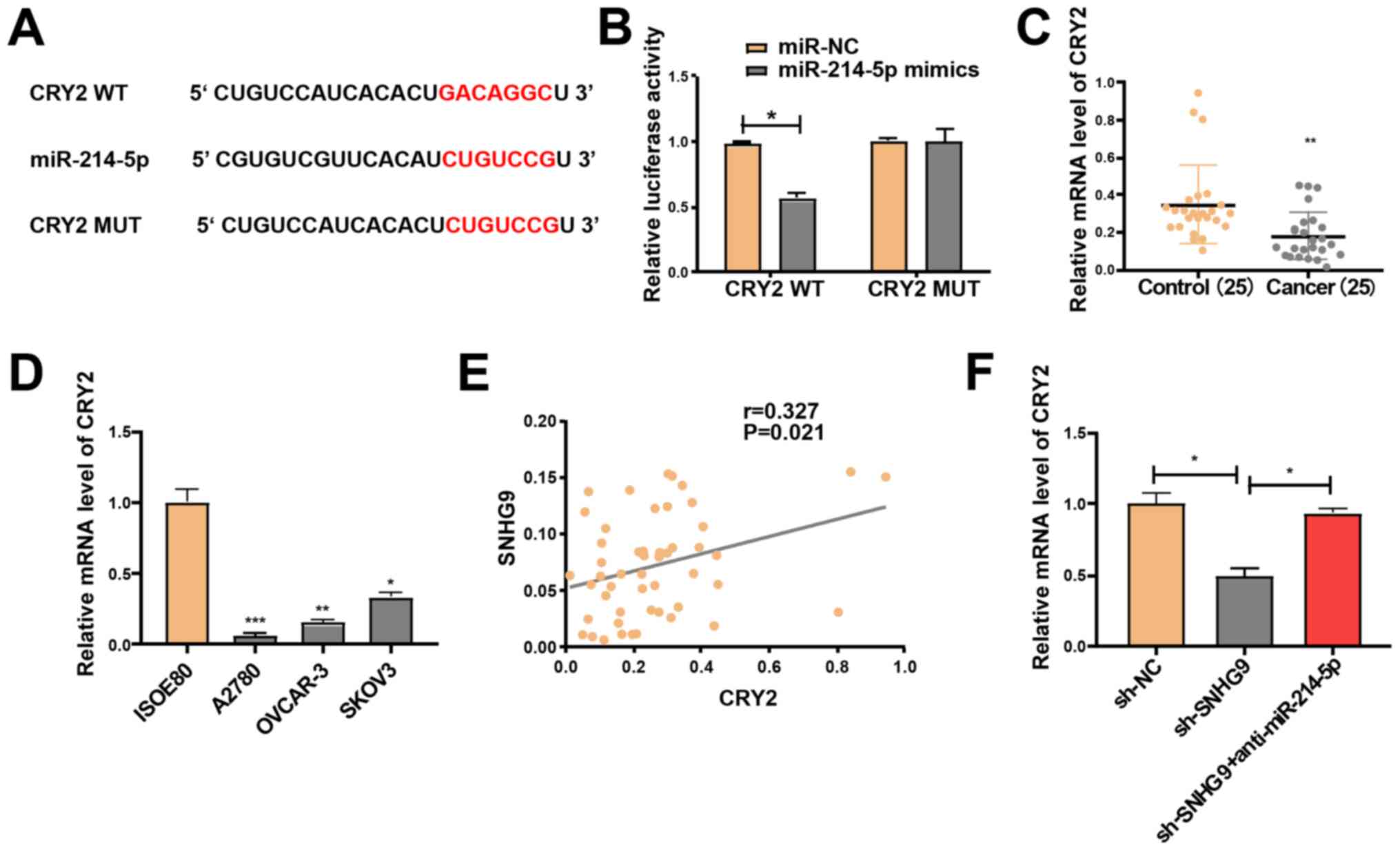
The target genes of miR-214-5p were screened using
TargetScan software and the miRDB database, in order to investigate
the potential role of miR-214-5p in the progression of ovarian
cancer. Following intersection of the target genes (score, >85)
from both databases, a total of 11 genes were selected, including
CRY2, which is closely associated with cancer progression (23,28).
Following their construction, the luciferase plasmids
pGL3-CRY2-3′UTR-WT and pGL3-CRY2-3′UTR-MUT were respectively
co-transfected with miR-214-5p mimics or NC in SKOV3 cells
(Fig. 4A). The luciferase activity
of the WT reporter was inhibited; however, the MUT reporter group
remained unchanged (Fig. 4B). Hence,
CRY2 may be a potential target gene of miR-214-5p. RT-qPCR analysis
demonstrated that CRY2 expression significantly declined in the
ovarian cancer cell lines and tissues (Fig. 4C and D). Pearson's correlation
analysis revealed that SNHG9 expression was positively correlated
with CRY2 expression (r=0.327; P=0.021; Fig. 4E). Subsequently, SKOV3 cells were
treated with sh-NC and sh-SNHG9, or co-transfected with sh-SNHG9
and anti-miR-214-5p. RT-qPCR analysis indicated that low SNHG9
expression significantly decreased CRY2 expression compared with
the sh-NC group; however, this effect was partially reversed by
miR-214-5p inhibitors in SKOV3 cells (P<0.05; Fig. 4F). Taken together, these results
suggest that SNHG9 regulates CRY2 expression via interaction with
miR-214-5p in ovarian cancer cells.
SNHG9/miR-214-5p axis regulates the
properties of ovarian cancer cells
SKOV3 cells were treated with sh-NC and sh-SNHG9, or
co-transfected with sh-SNHG9 and anti-miR-214-5p to determine
whether the inhibitory role of low SNHG9 expression in ovarian
cancer is mediated by miR-214-5p. Cell viability was detected via a
CCK-8 assay, while colony formation was assessed via a colony
formation assay. Subsequently, flow cytometry was performed to
assess cell cycle progression, and wound healing and Transwell
assays were performed to examine cell migration and invasion
abilities. The effects of low SNHG9 expression on cell migration,
colony formation, proliferation, cell cycle progression and
invasiveness were partially reversed by transfection with
anti-miR-214-5p (Fig. 5A-E), thus
suggesting that anti-miR-214-5p may regulate the roles of decreased
SNHG9 in ovarian cancer cells to a certain extent.
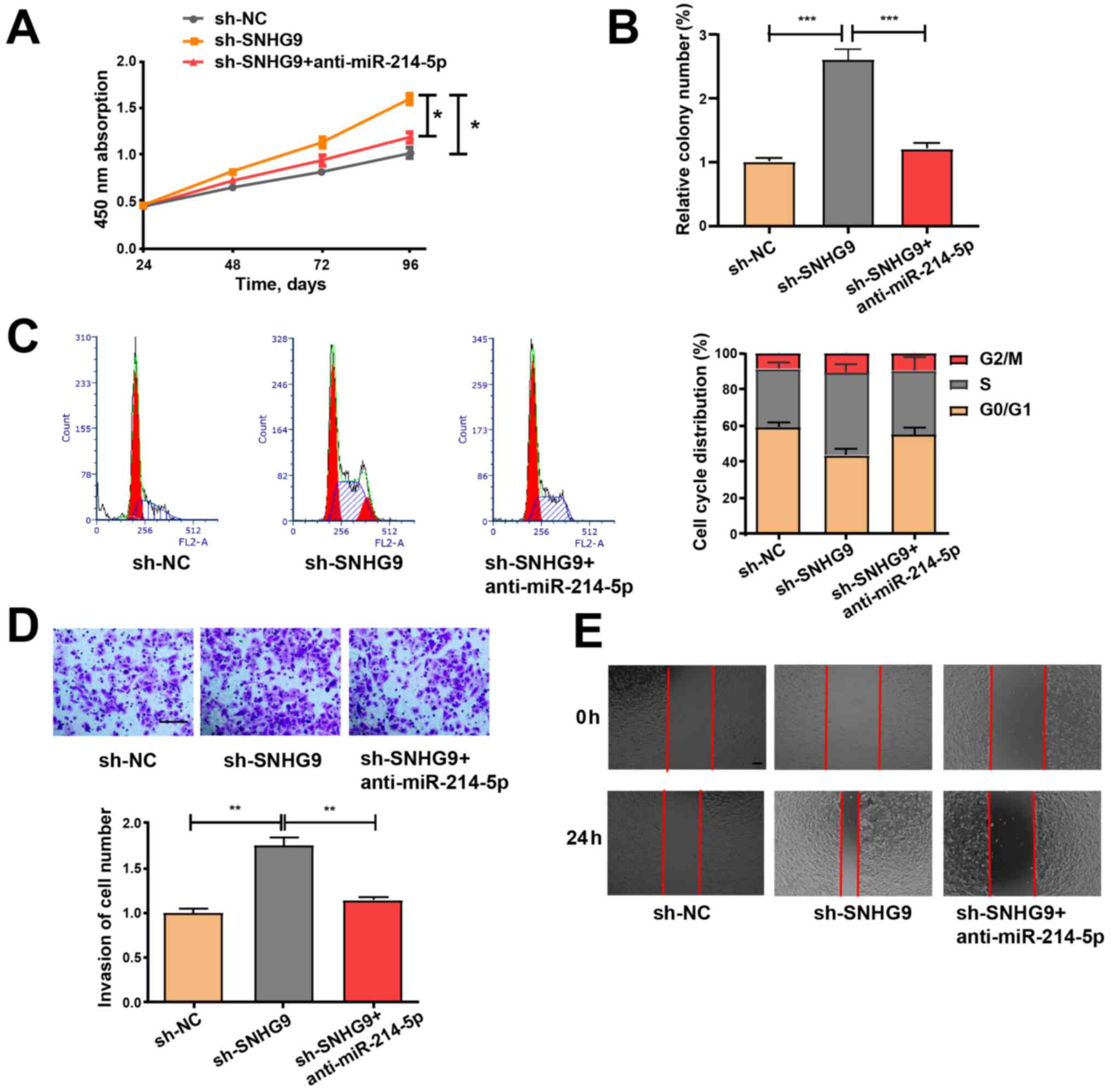 | Figure 5.miR-214-5p inhibitor partially
reverses the promoting effect of sh-SNHG9 on cell function. (A)
Cell Counting Kit-8, (B) colony formation, (C) cell cycle
progression, (D) Matrigel invasion (scale bar, 100 µm) and (E)
wound healing (scale bar, 100 µm) assays were performed on SKOV3
cells to assess cell proliferation, cycle, invasiveness and
migration. All experiments were performed in triplicate and data
are presented as the mean ± standard deviation. *P<0.05;
**P<0.01; ***P<0.001. miR, microRNA; sh, short hairpin;
SNHG9, small nucleolar RNA host gene 9; NC, negative control. |
Discussion
Aberrant regulation of lncRNAs is associated with
tumor development and carcinogenesis, and lncRNAs may be applied as
diagnostic, prognostic or therapeutic biomarkers for several types
of cancer, including gastric cancer (29), osteosarcoma (30) and ovarian cancer (31–33). For
example, Zhao et al (34)
reported that the lncRNA LINC00092 induces glycolysis in
fibroblasts associated with cancer, and influences the progression
of ovarian cancer. Furthermore, Zhang et al (35) demonstrated that the lncRNA, MIR4697HG
induces cell growth and metastasis in ovarian cancer. In the
present study, SNHG9 expression levels were detected in three
ovarian cancer cell lines (SKOV3, OVCAR-3 and A2780) and 25
patients with ovarian cancer. The results demonstrated that SNHG9
expression was notably downregulated in ovarian cancer cell lines
and tissues compared with the normal cell line and tissue samples,
respectively. Furthermore, decreased SNHG9 expression was indicated
to be associated with low overall survival time in patients with
ovarian cancer. In vitro analysis demonstrated that
transfection with sh-SNHG9 induced the migration, colony formation,
proliferation and invasion of ovarian cancer cells.
Subsequent experimentations were performed to
determine the underlying molecular mechanisms of SNHG9.
Dual-luciferase reporter assays, bioinformatics analyses and the
RIP assay verified that miR-214-5p is a direct target of SNHG9 in
ovarian cancer cells, and that CRY2 may be an underlying target of
miR-214-5p. Consistent with these results, miR-214-5p expression
increased in ovarian cancer tissues compared with paracancerous
normal tissues; however, CRY2 expression decreased in ovarian
cancer tissues compared with paracancerous normal tissues. Notably,
Pearson's correlation analysis demonstrated a negative association
between miR-214-5p and SNHG9, while a positive association was
indicated between CRY2 and SNHG9 expression levels in ovarian
cancer tissues, and transfection with a miR-214-5p inhibitor
partially reversed the effects of sh-SNHG9 on the proliferation,
cell cycle, migration and invasiveness of ovarian cancer cells.
Taken together, the present results suggest that SNHG9 may regulate
the expression of CRY2 by competitively binding to miR-214-5p,
thereby affecting the pathophysiological process of ovarian
cancer.
The current study presents a number of limitations.
First, a larger population size is required to further investigate
the clinical value of SNHG9. Secondly, the biological role of SNHG9
in ovarian cancer should be investigated using more ovarian cancer
cell lines. Thirdly, more target genes or miRNAs should be
investigated to evaluate their interaction with SNHG9. Furthermore,
the function of SNHG9 in the human body should be verified, and the
molecular mechanism of SNHG9 in ovarian cancer should be
investigated via further assays. For example, whether SNHG9
regulates the progression of ovarian cancer via the PI3K/AKT, NF-kB
or Wnt signaling pathways should be assessed.
In summary, SNHG9 was demonstrated to be
downregulated in human ovarian cancer cell lines and tissues, and
its expression was positively associated with overall survival in
patients with ovarian cancer. Decreased SNHG9 expression in ovarian
cancer cells was demonstrated to promote cell migration, colony
formation, proliferation, and invasion in vitro. Taken
together, the results suggest that SNHG9 may participate in ovarian
cancer development and progression by regulating the
miR-214-5p/CRY2 axis.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed in the present
study are available from the corresponding author upon reasonable
request.
Authors' contributions
YL made substantial contributions to the conception
and design of the present study. GC, ZL, YC and YL contributed to
the acquisition, analysis and interpretation of data, drafted the
initial manuscript and critically revised it for important
intellectual content. All authors read and approved the final
manuscript. and agreed to be accountable for all aspects of the
work in ensuring that questions related to the accuracy or
integrity of any part of the work are appropriately investigated
and resolved (according to the ICMJE).
Ethics approval and consent to
participate
The present study was approved by the Institutional
Ethics Committee of The Second People's Hospital of Taizhou City
(Taizhou, China). All population-associated experiments were
performed in accordance with The Declaration of Helsinki, and
written informed consent was provided by all patients prior to the
study start.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Tew WP: Ovarian cancer in the older woman.
J Geriatr Oncol. 7:354–361. 2016. View Article : Google Scholar
|
|
2
|
Webb PM and Jordan SJ: Epidemiology of
epithelial ovarian cancer. Best Pract Res Clin Obstet Gynaecol.
41:3–14. 2017. View Article : Google Scholar
|
|
3
|
Dong X, Men X, Zhang W and Lei P: Advances
in tumor markers of ovarian cancer for early diagnosis. Indian J
Cancer. 51 (Suppl 3):e72–e76. 2014. View Article : Google Scholar
|
|
4
|
Ting DT, Lipson D, Paul S, Brannigan BW,
Akhavanfard S, Coffman EJ, Contino G, Deshpande V, Iafrate AJ,
Letovsky S, et al: Aberrant overexpression of satellite repeats in
pancreatic and other epithelial cancers. Science. 331:593–596.
2011. View Article : Google Scholar
|
|
5
|
Qiu JJ, Lin YY, Ding JX, Feng WW, Jin HY
and Hua KQ: Long non-coding RNA ANRIL predicts poor prognosis and
promotes invasion/metastasis in serous ovarian cancer. Int J Oncol.
46:2497–2505. 2015. View Article : Google Scholar
|
|
6
|
Lai XJ and Cheng HF: lncRNA colon
cancer-associated transcript 1 (CCAT1) promotes proliferation and
metastasis of ovarian cancer via miR-1290. Eur Rev Med Pharmacol
Sci. 22:322–328. 2018.
|
|
7
|
Liu SP, Yang JX, Cao DY and Shen K:
Identification of differentially expressed long non-coding RNAs in
human ovarian cancer cells with different metastatic potentials.
Cancer Biol Med. 10:138–141. 2013.
|
|
8
|
Nikpayam E, Tasharrofi B, Sarrafzadeh S
and Ghafouri-Fard S: The role of long non-coding RNAs in ovarian
cancer. Iran Biomed J. 21:3–15. 2017. View Article : Google Scholar
|
|
9
|
Soudyab M, Iranpour M and Ghafouri-Fard S:
The role of long non-coding RNAs in breast cancer. Arch Iran Med.
19:508–517. 2016.
|
|
10
|
Cech TR and Steitz JA: The noncoding RNA
revolution-trashing old rules to forge new ones. Cell. 157:77–94.
2014. View Article : Google Scholar
|
|
11
|
Bunch H: Gene regulation of mammalian long
non-coding RNA. Mol Genet Genomics. 293:1–15. 2018. View Article : Google Scholar
|
|
12
|
Zhou HL, Luo G, Wise JA and Lou H:
Regulation of alternative splicing by local histone modifications:
Potential roles for RNA-guided mechanisms. Nucleic Acids Res.
42:701–713. 2014. View Article : Google Scholar
|
|
13
|
Yang L, Kirby JE, Sunwoo H and Lee JT:
Female mice lacking Xist RNA show partial dosage compensation and
survive to term. Genes Dev. 30:1747–1760. 2016. View Article : Google Scholar
|
|
14
|
Kanduri C: Long noncoding RNAs: Lessons
from genomic imprinting. Biochim Biophys Acta. 1859:102–111. 2016.
View Article : Google Scholar
|
|
15
|
Malakootian M, Mirzadeh Azad F, Fouani Y,
Taheri Bajgan E, Saberi H and Mowla SJ: Anti-differentiation
non-coding RNA, ANCR, is differentially expressed in different
types of brain tumors. J Neurooncol. 138:261–270. 2018. View Article : Google Scholar
|
|
16
|
Ranzani V, Arrigoni A, Rossetti G, Panzeri
I, Abrignani S, Bonnal RJP and Pagani M: Next-generation sequencing
analysis of long noncoding RNAs in CD4+ T cell differentiation.
Methods Mol Biol. 1514:173–185. 2017. View Article : Google Scholar
|
|
17
|
Dong J, Xu J, Wang X and Jin B: Influence
of the interaction between long noncoding RNAs and hypoxia on
tumorigenesis. Tumour Biol. 37:1379–1385. 2016. View Article : Google Scholar
|
|
18
|
Lou Y, Jiang H, Cui Z, Wang L, Wang X and
Tian T: Linc-ROR induces epithelial-to-mesenchymal transition in
ovarian cancer by increasing Wnt/β-catenin signaling. Oncotarget.
8:69983–69994. 2017. View Article : Google Scholar
|
|
19
|
Zheng ZG, Xu H, Suo SS, Xu XL, Ni MW, Gu
LH, Chen W, Wang LY, Zhao Y, Tian B and Hua YJ: The essential role
of H19 contributing to cisplatin resistance by regulating
glutathione metabolism in high-grade serous ovarian cancer. Sci
Rep. 6:260932016. View Article : Google Scholar
|
|
20
|
Gao C, Zhao D, Zhao Q, Dong D, Mu L, Zhao
X, Guo M, Xu A, Fang L, Liu Q and Che J: Microarray profiling and
co-expression network analysis of lncRNAs and mRNAs in ovarian
cancer. Cell Death Discov. 5:932019. View Article : Google Scholar
|
|
21
|
Zhang B, Li C and Sun Z: Long non-coding
RNA LINC00346, LINC00578, LINC00673, LINC00671, LINC00261, and
SNHG9 are novel prognostic markers for pancreatic cancer. Am J
Transl Res. 10:2648–2658. 2018.
|
|
22
|
Yu Y, Li Y, Zhou L, Yang G, Wang M and
Hong Y: Cryptochrome 2 (CRY2) suppresses proliferation and
migration and regulates clock gene network in osteosarcoma cells.
Med Sci Monit. 24:3856–3862. 2018. View Article : Google Scholar
|
|
23
|
Fang L, Yang Z, Zhou J, Tung JY, Hsiao CD,
Wang L, Deng Y, Wang P, Wang J and Lee MH: Circadian clock gene
CRY2 degradation is involved in chemoresistance of colorectal
cancer. Mol Cancer Ther. 14:1476–1487. 2015. View Article : Google Scholar
|
|
24
|
Tokunaga H, Takebayashi Y, Utsunomiya H,
Akahira JI, Higashimoto M, Mashiko M, Ito K, Niikura H, Takenoshita
SI and Yaegashi N: Clinicopathological significance of circadian
rhythm-related gene expression levels in patients with epithelial
ovarian cancer. Acta Obstet Gynecol Scand. 87:1060–1070. 2008.
View Article : Google Scholar
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
26
|
Helder-Woolderink JM, Blok EA, Vasen HF,
Hollema H, Mourits MJ and De Bock GH: Ovarian cancer in Lynch
syndrome; a systematic review. Eur J Cancer. 55:65–73. 2016.
View Article : Google Scholar
|
|
27
|
Miao H, Wang L, Zhan H, Dai J, Chang Y, Wu
F, Liu T, Liu Z, Gao C, Li L and Song X: A long noncoding RNA
distributed in both nucleus and cytoplasm operates in the
PYCARD-regulated apoptosis by coordinating the epigenetic and
translational regulation. PLoS Genet. 15:e10081442019. View Article : Google Scholar
|
|
28
|
Liu L, Shen H and Wang Y: CRY2 is
suppressed by FOXM1 mediated promoter hypermethylation in breast
cancer. Biochem Biophys Res Commun. 490:44–50. 2017. View Article : Google Scholar
|
|
29
|
Li P, Tong L, Song Y, Sun J, Shi J, Wu Z,
Diao Y, Li Y and Wang Z: Long noncoding RNA H19 participates in
metformin-mediated inhibition of gastric cancer cell invasion. J
Cell Physiol. 234:4515–4527. 2019. View Article : Google Scholar
|
|
30
|
Yao P, Ni Y and Liu C: Long non-coding RNA
691 regulated PTEN/PI3K/AKT signaling pathway in osteosarcoma
through miRNA-9-5p. Onco Targets Ther. 13:4597–4606. 2020.
View Article : Google Scholar
|
|
31
|
Yu F, Zhou G, Huang K, Fan X, Li G, Chen
B, Dong P and Zheng J: Serum lincRNA-p21 as a potential biomarker
of liver fibrosis in chronic hepatitis B patients. J Viral Hepat.
24:580–588. 2017. View Article : Google Scholar
|
|
32
|
Birgani MT, Hajjari M, Shahrisa A,
Khoshnevisan A, Shoja Z, Motahari P and Farhangi B: Long non-coding
RNA SNHG6 as a potential biomarker for hepatocellular carcinoma.
Pathol Oncol Res. 24:329–337. 2018. View Article : Google Scholar
|
|
33
|
Fu LL, Li CJ, Xu Y, Li LY, Zhou X, Li DD,
Chen SX, Wang FG, Zhang XY and Zheng LW: Role of lncRNAs as novel
biomarkers and therapeutic targets in ovarian cancer. Crit Rev
Eukaryot Gene Expr. 27:183–195. 2017. View Article : Google Scholar
|
|
34
|
Zhao L, Ji G, Le X, Wang C, Xu L, Feng M,
Zhang Y, Yang H, Xuan Y, Yang Y, et al: Long noncoding RNA
LINC00092 acts in cancer-associated fibroblasts to drive glycolysis
and progression of ovarian cancer. Cancer Res. 77:1369–1382. 2017.
View Article : Google Scholar
|
|
35
|
Zhang LQ, Yang SQ, Wang Y, Fang Q, Chen
XJ, Lu HS and Zhao LP: Long noncoding RNA MIR4697HG promotes cell
growth and metastasis in human ovarian cancer. Anal Cell Pathol
(Amst). 2017:82678632017.
|