Introduction
Worldwide, prostate cancer (PCa) is the second most
commonly diagnosed cancer in men, and there were ~1.1 million
diagnoses in 2012 (1). In China, the
incidence of PCa is lower compared with that in Western countries
(2). However, the incidence of PCa
is increasing in China, with an estimated 49,000 new diagnoses in
2012, and most of these patients were diagnosed with locally
advanced or metastatic PCa (2).
Unfortunately, due to late-stage diagnosis, these patients could
not undergo radical surgery, and androgen deprivation therapy (ADT)
was the standard systemic therapy for this group (2).
Initially, PCa progression is successfully
suppressed by ADT, as its growth is dependent on androgens
(3). However, almost all cases of
advanced PCa inevitably progresses toward castration-resistant PCa
(CRPC) (3). There are several drugs
used to treat CRPC. In 2004, docetaxel was approved by the Food and
Drug Administration to treat CRPC. In 2010, cabazitaxel and
immunotherapy were approved, enzalutamide was approved in 2012 and
abiraterone was approved in 2013 (4). Compared with mitoxantrone, docetaxel
only extends overall survival (OS) time by 2–2.9 months (5,6). In a
randomized phase III trial (PREVAIL) (7), enzalutamide was evaluated in 1,717
patients with chemo-naïve metastatic castration-resistant PCa
(mCRPC), and OS was significantly improved but was only extended by
2.2 months [32.4 vs. 30.2 months; hazard ratio (HR)=0.706; 95%
confidence interval (CI), 0.6–0.84; P<0.001). In the phase III
trial COU-AA-302 (8,9), abiraterone was evaluated in 1,088
patients with chemo-naïve, asymptomatic or mildly symptomatic
mCRPC, and OS was significantly improved but was only extended by
4.4 months (34.7 vs. 30.3 months; HR=0.81; 95% CI, 0.70–0.93;
P=0.0033). Overall, these results demonstrate that there is no
available curative therapy for CRPC. Notably, the lower the early
prostate-specific antigen (PSA) level after ADT treatment, the
longer the patient's response to ADT is (10). Therefore, treatments that decrease
the early PSA level combined with ADT and delay PCa progression to
CRPC may extend the OS of patients with PCa.
Previously, it was accepted that lysine methylation
was irreversible until lysine-specific demethylase 1 (LSD1) was
identified (11). Our previous
studies (12,13) and several others (14–16) have
demonstrated that overexpression of LSD1 is involved in PCa
initiation and progression. The androgen receptor (AR) signaling
axis serves an important role in the organogenesis, morphology and
normal functioning of the prostate (17). The vast majority of PCa cases express
high levels of AR and showed androgen-dependent growth (3). Hence, androgen ablation via castration
or administration of small chemical inhibitors is the most common
treatment for advanced PCa (3).
LSD1 forms chromatin-associated complexes with AR in
a ligand-dependent manner and demethylates the repressing histone
marks mono- and dimethyl H3-K9, thereby resulting in downregulation
of AR target genes, including PSA (14,16). PSA
is a major prognostic factor for PCa (10). Therefore, the present study aimed to
determine if downregulation of LSD1 expression could inhibit the AR
signaling axis and enhance the efficacy of ADT for PCa.
Materials and methods
Cell culture
LNCaP cells were obtained from the Cell Bank of the
Chinese Academy of Sciences. LNCaP cells were cultured in RPMI-1640
medium supplemented with 10% FBS (Gibco; Thermo Fisher Scientific,
Inc.) and 1% penicillin/streptomycin (Invitrogen; Thermo Fisher
Scientific, Inc.) at 37°C in a humidified incubator in the presence
of 5% CO2.
Cell transfection
LNCaP cells were transfected with 10 nM of small
interfering (si)RNA for LSD1 (siRNA-LSD1-LNCaP; cat. no. sc-60970;
Santa Cruz Biotechnology) or 10 nM of scrambled siRNA control using
Lipofectamine® 2000 (cat. no. 11668027; Invitrogen;
Thermo Fisher Scientific, Inc.), following the manufacturer's
instructions. siRNA-LSD1-LNCaP (siRNA-LSD1 group) and normal LNCaP
cells without treated with siRNA-LSD1 or scrambled siRNA (control
group) were used 48 h following transfection for the subsequent
experiments. The siRNA sequences against LSD1-1 were as follows:
forward 5′-CGGACAAGCUGUUCCUAAA-3′, reverse
3′-GCCUGUUCGACAAGGAUUU-5′; LSD1-2: forward
5′-GAACUCCAUCAGCAAUACA-3′, reverse 3′-CUUGAGGUAGUCGUUAUGU-5′.
Cell Counting Kit (CCK)-8 assay
The viability of the LNCaP cells was measured using
CCK-8 (Boster Biological Technology) according to the
manufacturer's instructions. The cells were seeded into a 96-well
microplate at a density of 5×103 cells/well and incubated at 37°C
for 24 h. Then, the cells were incubated with or without
bicalutamide (20 µM) for 0, 12, 24 or 48 h. Finally, the absorbance
of each well was measured at 490 nm using a Gemini XPS microplate
reader (Molecular Devices, LLC).
Migration and invasion activity
assay
The migration and invasion ability of
siRNA-LSD1-LNCaP or LNCaP cells were evaluated using the modified
Boyden chamber method. Cell migration assay was performed using
Transwell inserts without Matrigel (24 wells, 8-µm; cat. no.
354480; Corning Inc.). While, cell invasion assay was performed
using Transwell inserts with Matrigel (24 wells, 8-µm; cat. no.
354480; Corning Inc.). For the invasion assay, 5×104
cells were suspended and plated on inserts pre-coated with Matrigel
matrix at 37°C for 2 h. Outer wells were set as a chemoattractant
filling with 800 µl RPMI-1640 containing 10% FBS. After 48 h of
incubation at 37°C (13), the
invading cells were the cells on the undersurface of the filter
which were fixed with paraformaldehyde at room temperature for 10
min, stained with crystal violet at room temperature for 3 min and
counted manually five fields per membrane under a light microscope
(magnification, ×400) (Olympus Corp.).
Flow cytometric analysis
The siRNA-LSD1-LNCaP or LNCaP cells
(5×105 cells/ml) were inoculated into 6-well plates.
Each group comprised of three double wells on the plate. Following
24-h incubation at 37°C, the cells were treated with bicalutamide
(20 µM) and incubated at 37°C for an additional 24 h. Flow
cytometric analysis was performed according to the protocol of the
Annexin V-Fluorescein Isothiocyanate Apoptosis Detection kit
(Abcam). The apoptotic rates (included both early and late stages
of apoptosis) were analyzed immediately using a FACSCalibur™ flow
cytometer and BD FACSuite v.1.0.538.41 software (BD Biosciences).
The data was analyzed using SPSS v.17.0 (SPSS, Inc.).
Xenograft tumor experiments
Xenograft tumors were established using
early-passage cells and maintained in male NOD/SCID mice. A total
of 40 male NOD/SCID mice (15–20 g) were used. All animals were
obtained from Beijing Institute of Health Medicine and maintained
in standard conditions according to the institutional guidelines.
The Committee for Experimental Animals of Wuhan University (Wuhan,
China) approved all experimental procedures, and the procedures
complied with the Guidelines for the Care and Use of Laboratory
Animals. For subcutaneous injection (s.c.) tumor growth,
5×107 LNCaP cells (control group, n=20) or
siRNA-LSD1-LNCaP (siRNA-LSD1 group, n=20) cells were suspended in
100 µl of RPMI-1640 plus 100 µl Matrigel (Collaborative Biomedical
Products; BD Bioscience) and injected via a 27-gauge needle into
the s.c. space of the back of the neck. Approximately 10 days after
the LNCaP cells were injected into the NOD/SCID mice, palpable
tumors were detected. Four weeks after the LNCaP cells were
injected into the NOD/SCID mice, these mice were anesthetized with
pentobarbital (45 mg/kg) and castrated (implemented with bilateral
orchidectomy) using the scrotum incision approach. These mice were
then sacrificed with cervical dislocation on day 8 after
castration. Tumor tissues were fixed in 10% phosphate-buffered
formalin at room temperature for 24 h or immediately frozen and
stored at −80°C for different procedures. Serial sections of 5-µm
were cut from the tissue blocks.
TUNEL assay
Apoptosis was examined using an TUNEL assay and an
in situ Apoptosis Detection kit (Roche Applied Science). TUNEL
reagent steps were performed at room temperature according to the
manufacturer's instructions. Briefly, tumor tissues were fixed in
10% buffered formaldehyde at room temperature for a week, embedded
in paraffin blocks and then sectioned at a thickness of 4-µm.
Tissue sections were deparaffinized and rehydrated with xylene and
an ethanol series (100, 90, 80 and 70% ethanol) and treated with 20
µg/ml proteinase K at 37°C for 30 min. TUNEL reaction mixture (50
µl/section) was added and incubated at 37°C for 1 h. Converter-POD
(50 µl/section) (1:500; anti-fluorescein antibody conjugated with
horseradish peroxidase; cat. no. MK1025; BOSTER Biological
Technology Co., Ltd) was added. Tissue sections were incubated at
37°C for 30 min and the signal was developed using 0.1%
diaminobenzidine tetrahydrochloride for 5 min. Tissue sections were
dehydrated prior to mounting (Bio Optica Milano SpA). The nuclear
stain used was diaminobenzidine (DAB). Cover specimen was treated
with 100 µl of diluted DAB solution and incubated at room
temperature for 15 min. TUNEL-positive cells were identified by
brown or tan staining of the nucleus. Five fields of light
microscopy view (magnification, ×200) were randomly selected and
the apoptosis index was calculated as the ratio of
apoptotic-to-total cells.
Reverse transcription (RT)
semi-quantitative PCR
For gene expression studies RT-PCR was performed.
Total RNA was isolated using TRIzol® reagent
(Invitrogen; Thermo Fisher Scientific, Inc.), RNA concentration was
obtained by spectrophotometer and reverse transcription was
performed with the Revert Aid TM H Minus M-uLV Reverse
Transcriptase kit (Fermentas; Thermo Fisher Scientific Inc.)
according to the manufacturer's instructions. RNA was reverse
transcribed into single-stranded cDNA at 70°C for 5 min using the
cDNA synthesis kit (Takara Bio, Inc.) according to the
manufacturer's procedures. The primers used were as follows: LSD-1,
Forward: 5′-GCCCAAAGAAACTGTGGTGTC-3′ and reverse:
5′-TGTGGC-TGGGTAGTTACGGAT-3′; β-actin, forward:
5′-CACCCAGCACAATGAAGATCAAGAT-3′ and reverse:
5′-CCAGTTTTTAAATCCTGAGTCAAGC-3′. A total of 40 cycles of
amplification was performed for each of the RT-PCR experiments.
β-actin was amplified as an internal control. Initial denaturation
was done at 94°C for 5 min followed by 35 cycles of amplification.
Amplification protocol was repeated cycles of denaturation (30 sec,
94°C), annealing (30 sec, 56°C), extension (1 min, 72°C) and final
extension (7 min, 72°C). Amplified products were analyzed on 1.5–3%
agarose gels with Goldview staining at room temperature for 5 min
(Beijing Baihao Biological Technology Company). Gels were
visualized under UV light, photographed and optical densities of
the bands were analyzed using Quantity One software version 4.6.6
(Bio-Rad, Laboratories Inc.).
Western blot analyses
The protein expression levels of LSD1, PSA,
caspase-3, Bax and Bcl-2 were examined using western blotting.
Briefly, the proteins (30–50 µg/lane) were loaded onto 8–12% gels,
resolved using SDS-PAGE gels and then transferred to nitrocellulose
membranes (Bio-Rad Laboratories, Inc.). The membranes were blocked
with 5% non-fat milk in TBST buffer (10 mmol/l Tris-HCl, 0.15 mol/l
NaCl and 0.05% Tween-20, pH 7.2) for 2 h and incubated with primary
antibodies overnight at 4°C. The primary antibodies used were
monoclonal mouse antibodies against LSD1 (1:100; cat. no. 271720;
Santa Cruz Biotechnology, Inc.) and monoclonal rabbit antibodies
against PSA (1:800; cat. no. 5365; Cell Signaling Technology,
Inc.), caspase 3 (1:1,000; cat. no. 9662), Bax (1:800; cat. no.
2772) and Bcl-2 (1:1,000; cat. no. 3498) (all Cell Signaling
Technology, Inc.). After extensive washing with TBST buffer, the
membranes were incubated with HRP-conjugated anti-mouse (1:2,000;
cat. no. sc-516178) or anti-rabbit (1:2,000; cat. no. sc-2357)
secondary antibodies (both 1:2,000; Santa Cruz Biotechnology,
Inc.). The proteins were detected using an enhanced
chemiluminescence system (ECL kit; Pierce; Thermo Fisher
Scientific, Inc.) and captured on light-sensitive X-ray film
(Kodak). Optical densities were detected using ImageJ software
version 1.48 (National Institutes of Health).
Statistical analysis
Each sample was run in triplicates in 3 independent
experiments. All data are presented as the mean ± SEM. P<0.05
was considered to indicate a statistically significant difference.
The means of the different groups were compared using unpaired
Student's t-test. Multiple comparisons were analyzed by one-way
ANOVA followed by a Bonferroni's post hoc test. Categorical data
were analyzed using Fisher's exact test. Statistical analysis was
performed using SPSS 17.0 software (SPSS, Inc).
Results
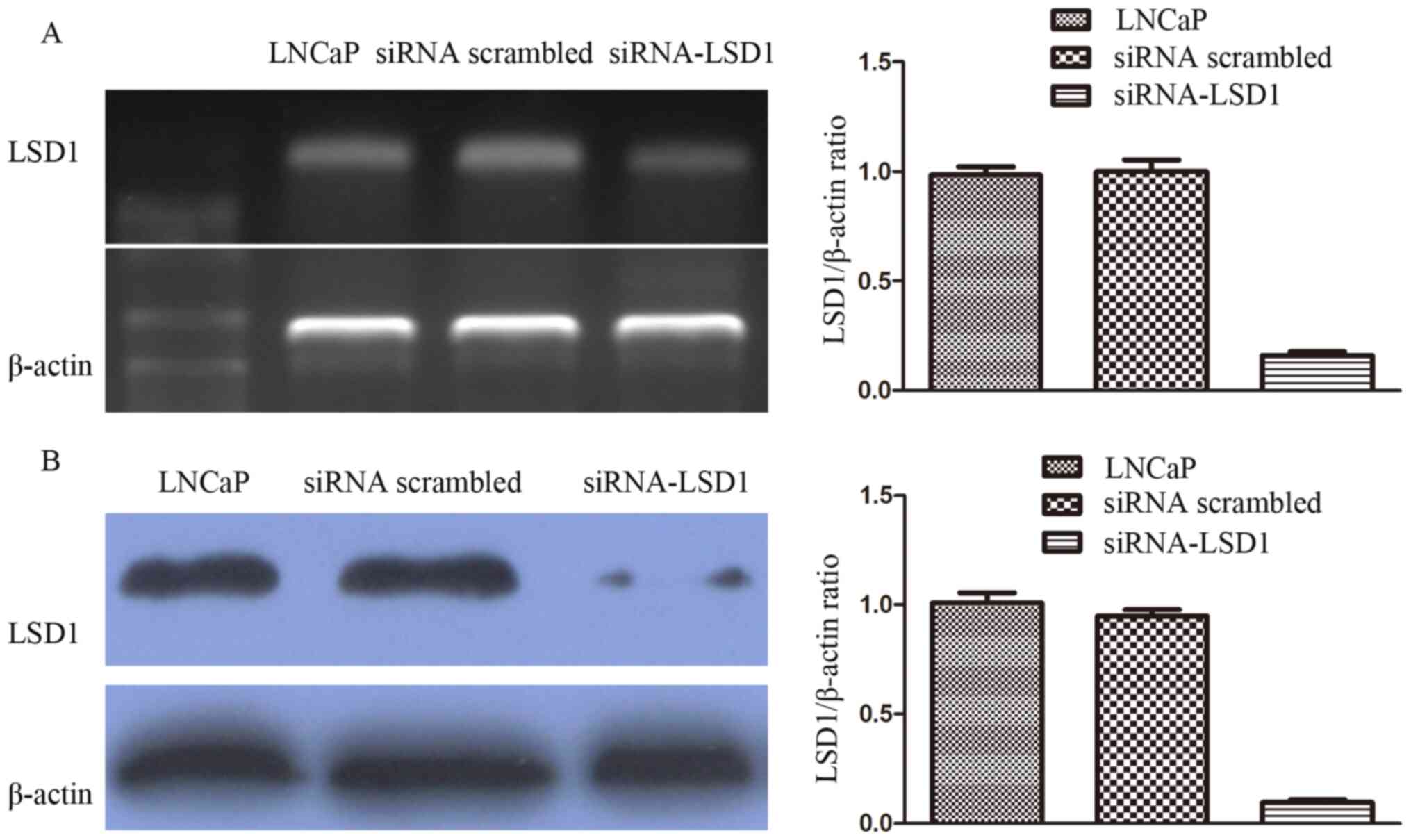
Transfection efficiency analyzed using
RT-PCR and western blotting
siRNA targeting LSD1 and scrambled siRNA were
constructed and transfected into LNCaP cells, and then RT-PCR and
western blotting were performed to evaluate transfection
efficiency. The results indicated that LSD1 mRNA levels were
markedly downregulated in LNCaP treated with siRNA-LSD1 compared
with those treated with scrambled siRNA and normal LNCaP cells
(LNCaP vs. siRNA scrambled vs siRNA-LSD1: 0.98±0.04 vs. 1.00±0.05
vs. 0.16±0.02; P<0.05; Fig. 1A).
The results also indicated that LSD1 protein levels were markedly
downregulated in LNCaP treated with siRNA-LSD1 compared with those
treated with scrambled siRNA and normal LNCaP cells (LNCaP vs.
siRNA scrambled vs. siRNA-LSD1: 1.01±0.05 vs. 0.95±0.03 vs.
0.10±0.01; P<0.05; Fig. 1B).
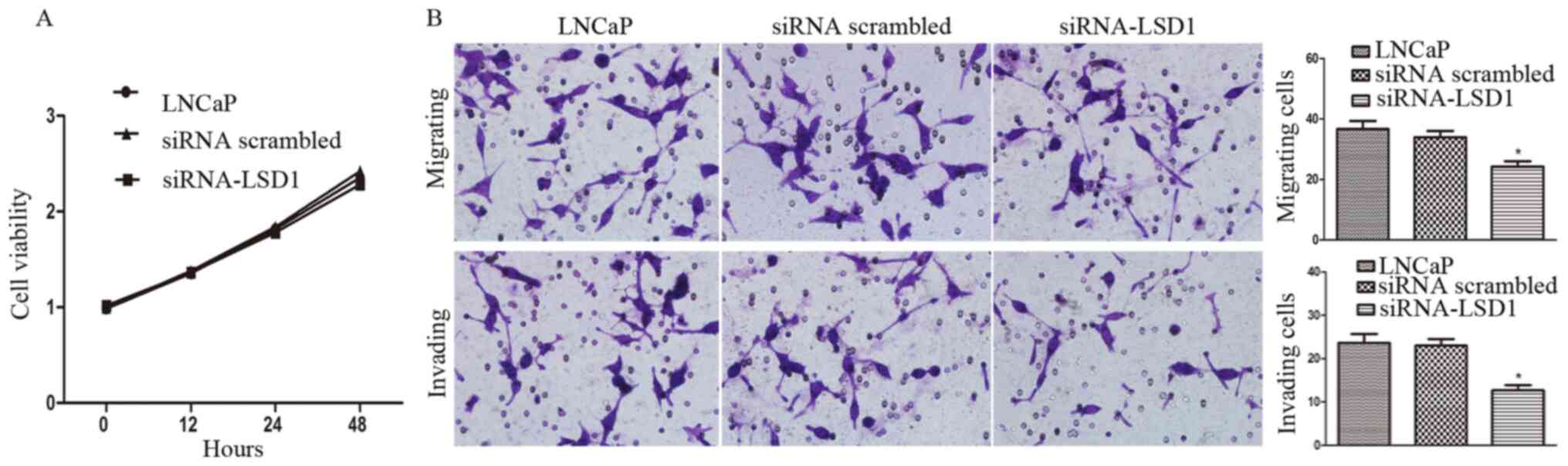
Knockdown of LSD1 attenuates the
invasion ability of LNCaP cells
Without bicalutamide treatment, LSD1-knockdown had
no significant impact on proliferation of LNCaP cells (P>0.05;
Fig. 2A). Furthermore, the effect of
LSD1-knockdown on the LNCaP cell migration and invasion was
examined using the Boyden chamber invasion assay. LSD1-knockdown
(siRNA-LSD1 group) significantly attenuated the migration
(P<0.05) and invasion (P<0.05) of LNCaP cells, compared with
LNCaP group and siRNA scrambled group, as shown in Fig. 2B, consistent with the findings of our
previous study via inhibition of LSD1 by its inhibitor pargyline
(13).
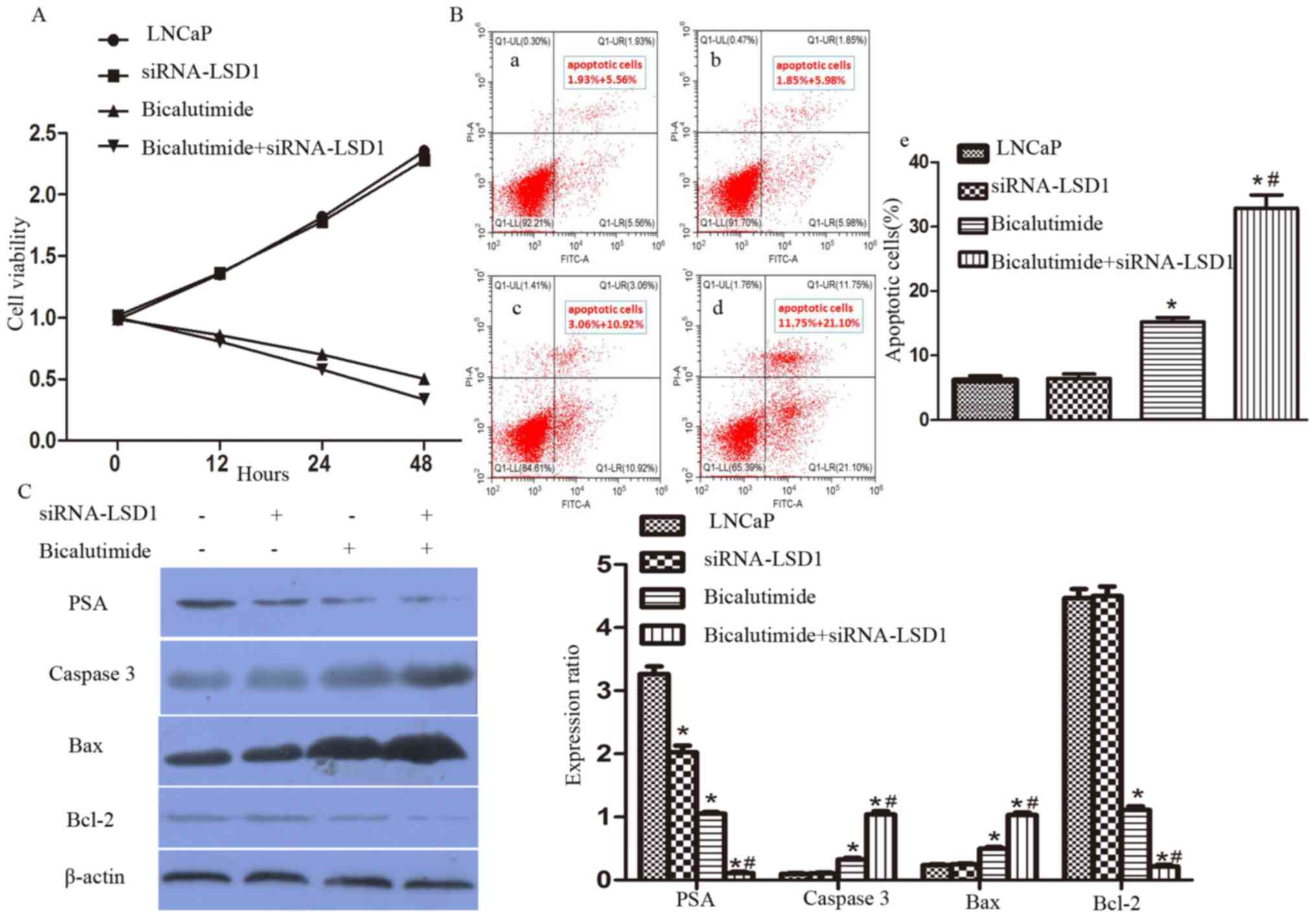
Knockdown of LSD1 enhances the
bicalutamide-induced apoptosis of LNCaP cells
Combined with bicalutamide treatment, LSD1-knockdown
(Bicalutimide + siRNA-LSD1 group) significantly decreased the
proliferation of LNCaP cells, compared with LNCaP group, siRNA
scrambled group and bicalutimide group (P<0.05; Fig. 3). Annexin V/PI-stained flow
cytometric analysis was used to determine whether the decreased
cell viability was due to apoptosis. The proportion of apoptotic
cells treated with bicalutamide increased significantly for the
cells treated with siRNA compared with the normal LNCaP cells
(bicalutamide group vs. bicalutamide+siRNA-LSD1 group, 16.78±0.90
vs. 33.16±2.16%, respectively (P<0.05; Fig. 3). Moreover, PSA and the
apoptosis-associated proteins caspase 3, Bax and Bcl-2 were
detected using western blot analyses (Fig. 3). Bicalutamide treatment combined
with LSD1-knockdown in the LNCaP cells (Bicalutimide + siRNA-LSD1
group) resulted in downregulation of PSA (P<0.05), upregulation
of caspase 3 (P<0.05) and Bax (P<0.05) and downregulation of
Bcl-2 (P<0.05) compared with the LNCaP group, siRNA scrambled
group and bicalutimide group.
Knockdown of LSD1 decreases the
tumorigenicity of LNCaP cells but has no significant impact on
tumor growth
Palpable tumors were detected in 75% of the NOD/SCID
mice 3 weeks after injection of the control LNCaP cells (LNCaP
group). However, palpable tumors were detected in only 35% of the
NOD/SCID mice 3 weeks after injection of the siRNA-LSD1-LNCaP cells
(siRNA-LSD1 group). In the siRNA-LSD1 group, the tumor sizes were
not significantly different from those in the LNCaP group (LNCaP
group vs. siRNA-LSD1 group: 813±6 vs. 807±12 mm3;
P=0.60; Table I), and there were no
significant differences in the maximum diameter of each tumor
during between LNCaP group (9.5–13.5 mm) and siRNA-LSD1 group (9–14
mm) (P>0.05; Fig. S1).
 | Table I.Effects of LSD1 on tumorigenicity and
tumor size. |
Table I.
Effects of LSD1 on tumorigenicity and
tumor size.
|
| Tumorigenicity |
|
|
|
|---|
|
|
|
|
|
|
|---|
| Group | Yes | No | P-value | Tumor size,
mm3 | P-value |
|---|
| LNCaP | 15 | 5 | 0.02 | 813±6 | 0.60 |
| siRNA-LSD1 | 7 | 13 |
|
807±12 |
|
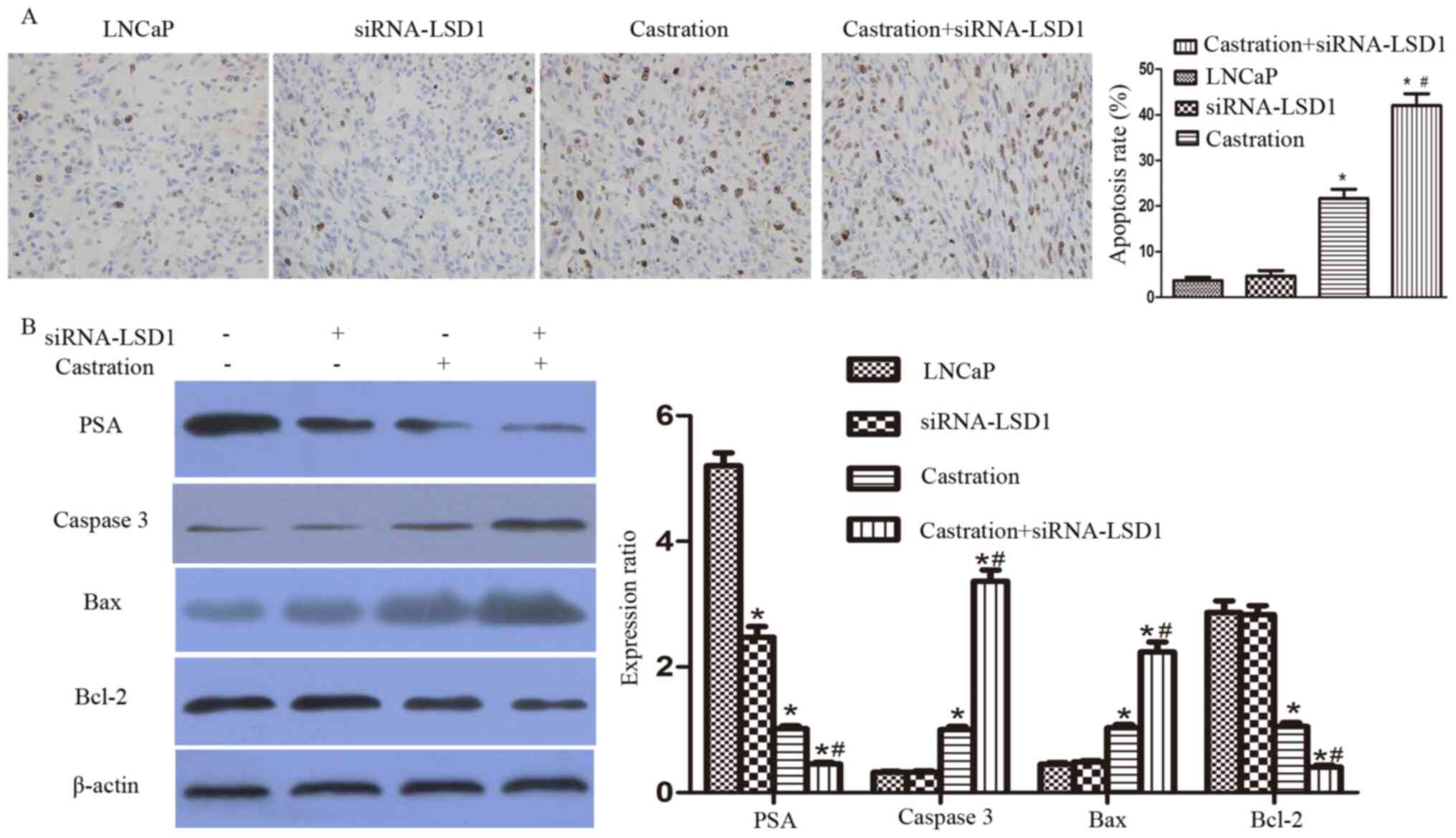
Knockdown of LSD1 enhances the
castration-induced apoptosis of LNCaP cells
Four weeks after injection of the LNCaP cells, the
mice with xenograft tumors were castrated via a bilateral
orchidectomy. These mice were then sacrificed on day 8 after
castration. The tumor tissues were used for the TUNEL assays and
western blot analyses. Consistent with the in vitro results
of the present study, the proportion of apoptotic cells in the
tumor tissues treated with castration and siRNA-LSD1 (Bicalutimide
+ siRNA-LSD1 group) increased significantly compared with the LNCaP
group, siRNA-LSD1 group and castration group (P<0.05; Fig. 4). Castration treatment combined with
LSD1-knockdown (Bicalutimide + siRNA-LSD1 group) showed significant
downregulation of PSA, upregulation of caspase 3 and Bax and
downregulation of Bcl-2, compared with the LNCaP group, siRNA-LSD1
group and castration group (P<0.05; Fig. 4).
Discussion
The present study demonstrated that knockdown of
LSD1 could effectively inhibit the invasion of LNCaP cells.
Moreover, knockdown of LSD1 combined with bicalutamide further
decreased PSA expression and proliferation and promoted apoptosis
of LNCaP cells in vitro. Furthermore, it was observed that
knockdown of LSD1 combined with castration enhanced the apoptosis
of LNCaP implant tumor cells, downregulating the expression of PSA
and Bcl-2 and upregulating the expression of caspase 3 and Bax.
AR is a steroid receptor and a member of the larger
nuclear receptor superfamily of proteins (18). The AR signaling pathway plays a
central role in normal prostate development and in PCa initiation
and progression (18). The exact
roles of AR in PCa are still unclear, but AR is involved in
stimulating the expression of a series of genes, such as cyclin D
proteins, mTOR and retinoblastoma tumor suppressor protein, that
regulate the PCa cell cycle, proliferation and survival (19,20). In
the early stage PCa cells are dependent on androgens for
proliferation (3,18). Therefore, ADT has been well accepted
as the standard treatment for patients with no indications of
radical prostatectomy (18).
LSD1 is involved in numerous pathological processes
of cancer, such as carcinogenesis, proliferation, apoptosis and
metastasis (21–25). Notably, multiple studies (14,16,26) have
demonstrated that LSD1 is a critical AR coregulator and plays a key
role in regulating AR signaling through its activator function.
These findings provided a strong molecular basis for knockdown of
LSD1 expression or LSD1 inhibitors to treat PCa. Hence, the present
study investigated the effect of the combination of ADT with
knockdown of LSD1 in preclinical models of hormone-sensitive PCa
using LNCaP cells and LNCaP xenograft tumors.
AR signaling regulates the transcription of protein
products that are required for prostate function, including PSA
(10,18). PSA is still the most commonly used
factor for PCa diagnosis, prognosis and follow-up (10,14,16). The
current study showed that LSD1-knockdown, combined with
bicalutamide treatment, significantly decreased the expression of
PSA in LNCaP cells. Furthermore, these results demonstrated that
LSD1-knockdown, combined with castration, significantly reduced the
expression of PSA in LNCaP xenograft tumors.
Apoptosis is characterized by a series of
physiological and pathological changes, such as cytoplasmic
blebbing, chromatin condensation and DNA fragmentation, which might
be induced by numerous different stimuli (27). Hormone-sensitive PCa cells are
dependent on AR signaling and are sensitive to AR antagonists
(3,18). Bicalutamide, the most commonly used
AR antagonist, effectively induces the apoptosis of PCa cells
(3). Members of the Bcl-2 family
(28,29), including Bax and Bcl-2, play a key
role in monitoring the mitochondrial pathway of apoptosis.
Furthermore, caspase 3 has been recognized as a critical downstream
effector of apoptosis (30). The
present study reported that knockdown of LSD1, combined with an AR
antagonist in vitro or with castration in vivo, could
significantly promote apoptosis of PCa cells, inhibit the
expression of the antiapoptotic Bcl-2 protein, and increase the
levels of the proapoptotic proteins Bax and caspase-3 in
vitro and in vivo.
The present experiments were conducted using only
one cell line, LNCaP, which is a limitation of the present study,
and use of clinical prostate cancer tissues or other cell lines may
not necessarily result in the same outcomes. The VCaP and C4-2 cell
lines should be used in future to validate the present
findings.
LSD1 functions as a transcriptional coregulator of
AR to enhance transcriptional activation or suppression of target
genes in a ligand-dependent manner (14,16,31). The
present results showed that knockdown of LSD1 could further
downregulate PSA expression and increase apoptosis of LNCaP cells
induced by bicalutamide in vitro, and enhance the apoptosis
of LNCaP xenograft tumor cells induced by castration. In
conclusion, knockdown of LSD1 expression might serve as a potential
therapeutic intervention to enhance the efficacy of ADT for PCa by
repressing AR signaling and promoting apoptosis.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This study was supported by grants from The National
Natural Science Foundation of China (grant no. 81972408), The
Province Natural Science Foundation of Hubei (grant no.
2019CFB302), The Province Natural Science Foundation of Hubei
(grant no. 2016CFB 114), The Science Foundation of Wuhan (grant no.
20150601010100 49), The Application and Basic Research Project Of
Wuhan City (grant no. 2018060401011321) and supported by the
Fundamental Research Funds for the Central Universities (grant no.
2042017kf0089).
Availability of data and materials
The datasets analyzed during the current study are
available from the corresponding author on reasonable request.
Authors' contributions
MW and XL conducted the experiments, analyzed the
data, and supervised and wrote the manuscript. MW, ZC and LZ
conducted the experiments and analyzed the data. MW, XW and LZ
performed the experiments and collated the data from them and
performed the statistical analysis. XL helped to design the study
and interpreted data. All the authors read and approved the final
version of the manuscript.
Ethics approval and consent to
participate
The present study was approved by The Ethics
Committee of the Renmin Hospital of Wuhan University (Wuhan,
China). Informed consent was obtained from all participants
(approval no. RM2018Y018-06). All animal study protocols were
approved by The Institutional Animal Use and Care Committee of
China Medical University (Shenyang, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
LSD1
|
lysine-specific demethylase 1
|
|
AR
|
androgen receptor
|
|
PCa
|
prostate cancer
|
|
ADT
|
androgen deprivation therapy
|
|
CRPC
|
castration-resistant prostate
cancer
|
|
OS
|
overall survival
|
References
|
1
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen W, Zheng R, Zeng H, Zou X, Zhang S
and He J: Report of Cancer Incidence and Mortality in China, 2011.
China Cancer. 24:1–10. 2012. View Article : Google Scholar
|
|
3
|
Larsson R, Mongan NP, Johansson M,
Shcherbina L, Abrahamsson PA, Gudas LJ, Sterner O and Persson JL:
Clinical trial update and novel therapeutic approaches for
metastatic prostate cancer. Curr Med Chem. 18:4440–4453. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rice MA, Malhotra SV and Stoyanova T:
Second-Generation Antiandrogens: From Discovery to Standard of Care
in Castration Resistant Prostate Cancer. Front Oncol. 9:8012019.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tannock IF, de Wit R, Berry WR, Horti J,
Pluzanska A, Chi KN, Oudard S, Théodore C, James ND, Turesson I, et
al TAX 327 Investigators, : Docetaxel plus prednisone or
mitoxantrone plus prednisone for advanced prostate cancer. N Engl J
Med. 351:1502–1512. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Scher HI, Morris MJ, Stadler WM, Higano C,
Basch E, Fizazi K, Antonarakis ES, Beer TM, Carducci MA, Chi KN, et
al: Trial design and objectives for castration-resistant prostate
cancer: updated recommendations from the prostate cancer clinical
trials working group 3. J Clin Oncol. 34:1402–1418. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Higano CS, Beer TM, Taplin ME, Efstathiou
E, Hirmand M, Forer D and Scher HI: Long-term safety and antitumor
activity in the Phase1-2 study of enzalutamide in pre- and
post-docetaxel castration-resistant prostate cancer. Eur Urol.
68:795–801. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ryan CJ, Smith MR, de Bono JS, Molina A,
Logothetis CJ, de Souza P, Fizazi K, Mainwaring P, Piulats JM, Ng
S, et al COU-AA-302 Investigators, : Abiraterone in metastatic
prostate cancer without previous chemotherapy. N Engl J Med.
368:138–148. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ryan CJ, Smith MR, Fizazi K, Saad F,
Mulders PF, Sternberg CN, Miller K, Logothetis CJ, Shore ND, Small
EJ, et al COU-AA-302 Investigators, : Abiraterone acetate plus
prednisone versus placebo plus prednisone in chemotherapy-naive men
with metastatic castration-resistant prostate cancer (COU-AA-302):
Final overall survival analysis of a randomised, double-blind,
placebo-controlled phase 3 study. Lancet Oncol. 16:152–160. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hussain M, Tangen CM, Higano C,
Schelhammer PF, Faulkner J, Crawford ED, Wilding G, Akdas A, Small
EJ, Donnelly B, et al Southwest Oncology Group Trial 9346
(INT-0162), : Absolute prostate-specific antigen value after
androgen deprivation is a strong independent predictor of survival
in new metastatic prostate cancer: Data from Southwest Oncology
Group Trial 9346 (INT-0162). J Clin Oncol. 24:3984–3990. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shi Y, Lan F, Matson C, Mulligan P,
Whetstine JR, Cole PA, Casero RA and Shi Y: Histone demethylation
mediated by the nuclear amine oxidase homolog LSD1. Cell.
119:941–953. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang M, Liu X, Jiang G, Chen H, Guo J and
Weng X: Relationship between LSD1 expression and E-cadherin
expression in prostate cancer. Int Urol Nephrol. 47:485–490. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang M, Liu X, Guo J, Weng X, Jiang G,
Wang Z and He L: Inhibition of LSD1 by Pargyline inhibited process
of EMT and delayed progression of prostate cancer in vivo. Biochem
Biophys Res Commun. 467:310–315. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Metzger E, Wissmann M, Yin N, Müller JM,
Schneider R, Peters AH, Günther T, Buettner R and Schüle R: LSD1
demethylates repressive histone marks to promote
androgen-receptor-dependent transcription. Nature. 437:436–439.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Metzger E, Imhof A, Patel D, Kahl P,
Hoffmeyer K, Friedrichs N, Müller JM, Greschik H, Kirfel J, Ji S,
et al: Phosphorylation of histone H3T6 by PKCbeta(I) controls
demethylation at histone H3K4. Nature. 464:792–796. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kahl P, Gullotti L, Heukamp LC, Wolf S,
Friedrichs N, Vorreuther R, Solleder G, Bastian PJ, Ellinger J,
Metzger E, et al: Androgen receptor coactivators lysine-specific
histone demethylase 1 and four and a half LIM domain protein 2
predict risk of prostate cancer recurrence. Cancer Res.
66:11341–11347. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Schaeffer EM, Marchionni L, Huang Z,
Simons B, Blackman A, Yu W, Parmigiani G and Berman DM:
Androgen-induced programs for prostate epithelial growth and
invasion arise in embryogenesis and are reactivated in cancer.
Oncogene. 27:7180–7191. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Green SM, Mostaghel EA and Nelson PS:
Androgen action and metabolism in prostate cancer. Mol Cell
Endocrinol. 360:3–13. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Knudsen KE, Arden KC and Cavenee WK:
Multiple G1 regulatory elements control the androgen-dependent
proliferation of prostatic carcinoma cells. J Biol Chem.
273:20213–20222. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xu Y, Chen SY, Ross KN and Balk SP:
Androgens induce prostate cancer cell proliferation through
mammalian target of rapamycin activation and post-transcriptional
increases in cyclin D proteins. Cancer Res. 66:7783–7792. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Huang J, Sengupta R, Espejo AB, Lee MG,
Dorsey JA, Richter M, Opravil S, Shiekhattar R, Bedford MT,
Jenuwein T, et al: p53 is regulated by the lysine demethylase LSD1.
Nature. 449:105–108. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Scoumanne A and Chen X: The
lysine-specific demethylase 1 is required for cell proliferation in
both p53-dependent and -independent manners. J Biol Chem.
282:15471–15475. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Schulte JH, Lim S, Schramm A, Friedrichs
N, Koster J, Versteeg R, Ora I, Pajtler K, Klein-Hitpass L,
Kuhfittig-Kulle S, et al: Lysine-specific demethylase 1 is strongly
expressed in poorly differentiated neuroblastoma: Implications for
therapy. Cancer Res. 69:2065–2071. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hayami S, Kelly JD, Cho HS, Yoshimatsu M,
Unoki M, Tsunoda T, Field HI, Neal DE, Yamaue H, Ponder BA, et al:
Overexpression of LSD1 contributes to human carcinogenesis through
chromatin regulation in various cancers. Int J Cancer. 128:574–586.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bennani-Baiti IM, Machado I,
Llombart-Bosch A and Kovar H: Lysine-specific demethylase 1
(LSD1/KDM1A/AOF2/BHC110) is expressed and is an epigenetic drug
target in chondrosarcoma, Ewing's sarcoma, osteosarcoma, and
rhabdomyosarcoma. Hum Pathol. 43:1300–1307. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cai C, He HH, Gao S, Chen S, Yu Z, Gao Y,
Chen S, Chen MW, Zhang J, Ahmed M, et al: Lysine-specific
demethylase 1 has dual functions as a major regulator of androgen
receptor transcriptional activity. Cell Rep. 9:1618–1627. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
McConkey DJ, Orrenius S and Jondal M:
Cellular signalling in programmed cell death (apoptosis). Immunol
Today. 11:120–121. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Strobel T, Swanson L, Korsmeyer S and
Cannistra SA: BAX enhances paclitaxel-induced apoptosis through a
p53-independent pathway. Proc Natl Acad Sci USA. 93:14094–14099.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Antonsson B and Martinou JC: The Bcl-2
protein family. Exp Cell Res. 256:50–57. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gross A, McDonnell JM and Korsmeyer SJ:
BCL-2 family members and the mitochondria in apoptosis. Genes Dev.
13:1899–1911. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lin T, Ponn A, Hu X, Law BK and Lu J:
Requirement of the histone demethylase LSD1 in Snai1-mediated
transcriptional repression during epithelial-mesenchymal
transition. Oncogene. 29:4896–4904. 2010. View Article : Google Scholar : PubMed/NCBI
|