Introduction
Amphibian antimicrobial peptides (AMPs) are
important components of the amphibian innate immune system
(1). AMPs have a wide range of
biological activities, including antibacterial, antiviral and
antiparasitic activities (2). In
addition, AMPs have antitumor activity (3). Previous studies have shown that
Temporin-La (T-La), a peptide isolated from a bullfrog skin cDNA
library, has antitumor activity and mild hemolytic activity
(4,5). Antitumor drugs exploit biological
characteristics of tumor cells (6)
to target the biological activity to specific tissues, cells or
organs (7), while sparing normal
cells, tissues and non-target organs (8). Improved targeting of antitumor drugs
will improve treatment efficacy, while reducing toxicity (9). Integrin αvβ3, which is highly expressed
on the surface of certain tumor cells, can be exploited to improve
antitumor drug targeting (5).
Integrin αvβ3 is also expressed on endothelial cells in the
basement membrane, which is traversed by migrating tumor cells
(10). The RGD peptide is a small
molecule peptide, containing three amino acids, that specifically
binds to integrin αvβ3 (11,12). Thus, the specific binding qualities
and small size of the RGD peptide can be exploited to improve the
targeting and efficacy of antitumor drugs.
The present study evaluated the antitumor properties
of the T-La peptide and T-La derivatives. To improve the targeting
of T-La to tumor cells and to decrease its cytotoxicity, the amino
acid sequence was altered and an RGD chimeric T-La peptide was
designed, called T-La (FS), using bioinformatics methods and
analyses. The secondary structure of T-La (FS) was determined.
Tumor targeting of the polypeptides was assessed in vivo and
the effects of the peptides on apoptosis-related protein caspase-3
and endothelial growth factors were evaluated. The inhibitory
effects of T-La (FS) on tumor growth were determined. The present
study provides the scientific basis for the application of small
molecule antimicrobial peptides as anti-tumor agents. Furthermore,
the current study aimed to establish the theoretical foundation for
the development and clinical application of these peptides as
antitumor drugs.
Materials and methods
Database
Peptide (Temporin-La) details were obtained from the
Antibacterial Peptide Database (http://aps.unmc.edu/AP/main.php), which characterizes
various antimicrobial peptides.
Software
Protein sequence analyses were performed using the
ExPasy sequence analysis and protein analysis tools (https://web.expasy.org/protparam/). The secondary
structure was analyzed using the Spiral online analysis software
(http://heliquest.ipmc.cnrs.fr/).
Chemical structure simulation was performed using the PyMoL
Molecular Graphics System software version 1.5.0.4 (Schrödinger,
LLC. http://sourceforge.net/projects/pymol).
Samples
Polypeptides were synthesized by Zhejiang Hongtuo
Biotechnology Co., Ltd. using solid phase synthesis. Samples were
desalted and purified (95% purity) using HPLC (Agilent high
performance liquid chromatography, Agilent Technology Co., Ltd.,
Waters 600 controller) reverse phase C18 column (Agilent ZORBAX;
cat. no. 300SB-C18 5 µm; 4.6×250 mm; Agilent Technology Co.,
Ltd.) chromatography. The whole liquid phase system was used at
room temperature and the composition of mobile phase was A, 0.1%
TFA in H2O and B, 0.1% TFA in 100% ACN; The sample
quantity of the system was 20 µl and the flowrate was 1.0 ml/min.
The molecular mass was determined using fast atom bombardment mass
spectrometry (Applied Biosystems 3200 QTRAP LC/MS/MS; Applied
Biosystems Inc.). The positive ionization mode was used in the mass
spectrometer and the details were as follows: nitrogen gas
temperature (350°C), nebulizer pressure (40 psi) and flow rate (0.3
ml/min).
Reagents
Reagents used in the present study included
RPMI-1640 (pH 7.2) medium, high-sugar DMEM medium (Beijing Solarbio
Science & Technology Co., Ltd.), South American fetal bovine
serum (Gibco; Thermo Fisher Scientific, Inc.), dimethyl sulfoxide
(DMSO), trypsin, penicillin-streptomycin, MTT, DAPI nuclear stain
(Beijing Zhongsheng Ruitai Technology Co., Ltd.), 3% glutaraldehyde
(Alfa Aesar; Thermo Fisher Scientific, Inc.) mouse Anti-Human CD61
PE-labeled antibody (Beijing Solarbio Science & Technology Co.,
Ltd.), cisplatin (Qilu Pharmaceutical), 10% sheep serum (NGS;
Shanghai Biosun Sci&Tech Co., Ltd.) and 0.05% DAB containing
0.03% hydrogen peroxide (protected from light). The cell
endothelial growth factor VEGF (1:100; cat. no. 251659) and
apoptosis-related protein caspase-3 (1:100; cat. no. 341034)
primary antibodies and the secondary goat anti-mouse antibodies IgG
were obtained from Zen BioScience Co., Ltd. (1:200; cat. no.
550047). Primary antibodies against cleaved caspase-3 (1:1,000;
cat. no. ab13847), GAPDH (1:1,000; cat. no. ab8245) and goat
anti-rabbit horseradish peroxidase secondary antibody (1:3,000;
cat. no. ab7621) were obtained from Abcam.
Experimental animals
All animal experiments were approved by The
Laboratory Animal Ethics Committee of Tianjin Medical University
(Tianjin, China). In total, 41 BALB/c female nude mice, aged 5–6
weeks, weighing 18–22 g, were used. The mice were reared in
separate cages to ensure stable atmospheric pressure difference,
with sufficient food and water and ambient temperature controlled
at 25±1°C. The cycle of light and dark was 12 h alternately. The
mice were obtained from a specific pathogen free facility (cat. no.
11400700271441; Beijing Vital River Laboratory Animal Technology
Co., Ltd.).
Main instruments used
The following instruments were used in this study:
Circular Chromatograph J-715 (JASCO), Microplate Reader (Bio-Rad
Laboratories, Inc.), Inverted Fluorescence Microscope (Nikon
ECLIPSE Ti; Nikon Corporation), Laser Confocal Microscope (Leica
TCS SP5 II; Leica Microsystems, Inc.), Scanning Electron Microscope
QUANTA 200 (Thermo Fisher Scientific, Inc.), small animal imaging
system (Kodak), automatic dyeing machine (LEICA ST5010) paraffin
embedding machine (LEICA EG1150H), and rotary slicer (LEICA RM2235)
(all Leica Microsystems, Inc.).
Cell lines
Melanoma cells (B16; Hunan Fengh Biotech Co., Ltd.),
hepatoma cells (Huh-7.5.1; Veterinary Laboratory of Tianjin
Agricultural University), mouse fibroblast cells (NIH-3T3), mouse
hepatoma cells (HEPA1-6), human lung cancer cells (NCI-H460), human
hepatoma cells (HepG2) and human breast cancer cells (MCF7) were a
gift from the Institute of Radiology, Chinese Academy of
Sciences.
Bioinformatics analysis of
antimicrobial peptide sequences
Peptide sequences were entered into http://www.expasy.ch/cgi-bin/protparam
and http://heliquest.ipmc.cnrsunitr/ to
analyze the physical and chemical properties of the antimicrobial
peptide molecules. The following parameters were obtained: Net
charge, hydrophobicity, hydrophobic torque and amphiphilicity.
Secondary structure prediction and 3D
structure simulation of polypeptides using circular dichroism (CD)
spectrum analysis
The CD spectrum was measured using a Jasco J-715
circular dichroic instrument equipped with a Peltier type
temperature control system. Molar ellipticity (θ) unit
deg/cm2 · dmol, was calculated using the formula
[θ]=100θ/(cl), where c is the molar concentration of the
polypeptide, and l is the thickness (cm) of the specific cuvette.
The CD spectrum was processed using Origin 7.5 software (Origin
Lab). The 3D structural simulation of several antitumor peptides
was plotted using the PyMoL Molecular Graphics System chemical
structure simulation software.
MTT assay to detect the inhibitory
activity of the polypeptides on tumor cells
Seven tumor cell lines (B16, huh7, 3T3, HEPA1-6,
H460, HepG2 and MCF7) were used in the present study. The cells
were cultured in 96-well plates at a concentration of
5×103 cells per well in a 100 µl. After 24 h
incubation at 37°C and 5% CO2, the four different
antitumor peptides were added at final concentrations of 200, 50,
10, 5 and 1 µg/ml in 200 µl per well. Our preliminary
experiment results demonstrated that the cell survival rate of 100
µg/ml peptides was basically 0 for tumor cells (data not
shown). In order to investigate the high concentration toxicity of
peptides to tumor cells, 200 µg/ml was chosen for the
present study. There were five experimental replicates per group.
Plates were incubated at 37°C and 5% CO2 for 24 h. MTT
(20 µl of 5 mg/ml) was added to each well, and the plates
were incubated at 37°C for 4 h. The culture medium was removed and
150 µl DMSO was added into each well. The crystals were
thoroughly dissolved and the absorbance (OD value) of each well
(wavelengths 490 or 630 nm) was measured with a microplate reader.
Each experiment was repeated three times, and the cell
proliferation inhibition rate was calculated according to the
following formula: Cell proliferation inhibition rate
(%)=(1-average OD value of the experimental group/average OD value
of the normal control group)×100%.
Real-time observation of the effects
of the polypeptides on cell morphology
The B16 melanoma cell line was sensitive to the
polypeptides, as indicated by the MTT assay. B16 cells were
inoculated on a laser confocal cell culture dish and cultured to
80% confluence at 37°C and 5% CO2. Fluorescein
isothiocyanate (FITC)-labeled polypeptides were incubated with the
tumor cells at a final concentration of 25 µg/ml. Then, DAPI
(nuclear stain) was added for 10 min at room temperature. Real-time
acquisition was performed using a confocal microscope (FluoView
FV1000; Olympus Corporation) (magnification, ×100). Images were
captured every 10–20 min for 2 h, while cells were maintained at
37°C.
Effects of polypeptide treatment on
the tumor cell membrane
B16 melanoma cells were added to 6-well plates
containing sterile cell slides at a density of 5×105
cells/well. Cells were cultured for 12 h (37°C and 5%
CO2) to reach 80% confluence. Antitumor peptides were
added to the cells at 50, 10, 5 and 1 µg/ml for 48 h. Cells
were then collected, washed with PBS three times and fixed with 3%
glutaraldehyde for 2 h at room temperature. After incubation at 4°C
for 15 min, cells were washed three times in PBS. Cells were
dehydrated using a step-by-step ethanol gradient at 30, 50, 70 and
90% for 15 min each step. Final dehydration in 100% ethanol was
performed three times for 10 min each. Samples were dried using the
critical point drying method (13)
and then the samples were coated and sprayed with gold, after
adhering to the sample table.
Detection of cell surface integrin
αvβ3 expression
Cells without polypeptide were selected as normal
controls, and 1×106 HepG2, 3T3, MCF7 or HEPA1-6 cells
were incubated with mouse anti-human CD61 PE-labeled antibody at
4°C for 10 min. The fluorescence intensity was measured by flow
cytometry (BD FACS Aria III; Becton, Dickinson and Company). BD
Accurate C6 software version 1.0.264.21 (Becton, Dickinson and
Company) was used for data analysis. Differences in the cell
surface expression of integrin αvβ3 were evaluated.
Detection of caspase-3 apoptotic
protein expression
After reaching ~80% confluency, B16 cells were
collected and diluted with cell culture medium (RPMI-1640) to
7×104 cells/ml. The cells were added to 6-well plates (2
ml per well) and treated with peptides at final concentrations of
10, 20, 50, and 100 µg/ml for 24 h. B16 cells which had PBS
added were the control group. After washing cells three times with
PBS, NP-40 was added and the cells were incubated at 4°C for 1 h.
The cells were then collected and stored at −80°C or used
immediately.
Samples were centrifuged (12,000 × g for 5 min at
4°C) and the supernatants were collected. The total protein
concentration was measured using the BCA method. Loading buffer was
added to each sample (50 ng total protein) and the samples were
boiled. Samples were separated using 15% SDS-PAGE electrophoresis
and transferred to a PVDF membrane using the semi-dry method
(14). After blocking with 5%
skimmed milk at 4°C overnight, cleaved caspase-3 and GAPDH primary
antibodies were added and the membrane was incubated for 1.5 h at
room temperature. After incubation with horseradish
peroxidase-labeled goat anti-rabbit secondary antibodies for 1 h at
room temperature, the bound secondary antibodies were detected with
enhanced chemiluminescence (ECL Chemiluminescent Substrate Reagent
kit; Thermo Fisher Scientific Inc.). Image J software (version 1.51
q and k; National Institutes of Health) was used to analyze the
gray value of each band, and the target protein to GAPDH ratio was
determined.
Reverse transcription-quantitative
(RT-q)PCR analysis of apoptotic gene mRNA expression
After reaching ~80% confluency, B16 cells were
collected and diluted with cell culture medium to 7×104
cells/ml. B16 cells were added to 6-well plates. After overnight
culture, the cells were treated with different concentrations of
polypeptides (20, 50 and 100 µg/ml) and the control group
was untreated. After 24 h, cells were collected for total RNA
extraction according to the RNA extraction kit (RNAprep pure Cell
kit; TIANGEN Biotech Co., Ltd.) instructions. The RNA concentration
was determined. RNA was reverse transcribed into cDNA for PCR
amplification, The reverse transcription program was 45°C for 10
min (UltraSYBR Mixture; ComWin Biosciences). The PCR reaction
procedure was as follows: pre-denaturation at 95°C for 10 min;
denaturation at 95°C for 15 sec, annealing/extension at 60°C for 1
min, total 35 cycles. The PCR primers sequences were as follows:
GAPDH forward, 5′-GCACAGTCAAGGCTGAGAATG-3′ and reverse
5′-ATGGTGGTGAAGACGCCAGTA-3′; caspase 3 forward,
5′-CTGGACTGTGGCATTGAGAC-3′ and reverse 5′-ACAAAGCGACTGGATGAACC-3′.
Quantitative analysis was performed on a real-time PCR instrument
using SYBR-Green dye and the final data was analyzed using the
2−ΔΔCq method (13).
Targeted distribution of RGD-chimeric
peptide in nude mice
Establishment of a liver tumor
model
In total, 15 BALB/c nude mice were randomly divided
into three groups, RGD-T-La (FS) group, T-La (FS) group and a blank
control group, with five mice in each group. The mouse model of
subcutaneous transplantation of HepG2 hepatoma cells was
established according to a previous study (12). The concentration of HepG2 cells was
adjusted to 106 cells/ml. The cells were injected
subcutaneously (100 µl/mouse) in the axillary posterior part of the
forelimbs. When tumors developed to ~5 mm in diameter, RGD-T-La
(FS) or T-La (FS) was injected into the tail vein at a dose of 100
µl/mouse (2 µmol/ml). The blank control group was
injected with PBS 100 µl/mouse.
Distribution of antitumor peptides in
mice
After injection of RGD-T-La (FS), T-La (FS) or PBS,
the heart, liver, spleen, lung, kidney and tumor tissues were
collected from mice at 30 min, 1, 2, 4 and 8 h post injection. The
fluorescence distribution and intensity of FITC-labeled antitumor
peptide was observed using a small animal imaging system and
compared with the blank group.
Making pathological slices of
subcutaneous liver tumors
Pathological sections of liver tumors were made
according to the method described by Ji and Zhang (13).
Therapeutic effects of RGD-chimeric
peptides on melanoma in a nude mouse model
Subcutaneous melanoma model in nude
mice
In total, 36 female BALB/c nude mice, aged 5–6
weeks, were randomly divided into six groups (n=6 per group): i)
T-La (FS); ii) RGD-T-La (FS); iii) T-La; iv) RGD-T-La; v) Cisplatin
(positive control); and vi) PBS (blank control). B16 melanoma cells
were adjusted to 5×105 cells/ml in saline suspension and
each mouse was inoculated subcutaneously into the forelimb axilla
with 0.1 ml of the cell suspension. The success of tumor growth was
confirmed visually when the tumor reached ~5×5 mm2, and
the tumor sizes in each group were measured. Mice in each group
were injected with the corresponding drug treatments (600
µg/ml of peptide). The positive control group was treated
with 1 mg/ml cisplatin solution prepared in saline and injected
through the tail vein.
Effects of antitumor peptides on
melanoma growth in vivo
Drug treatments were administered every 48 h for 10
days (five doses in total). Tumor growth in each group was
recorded. Nude mice were weighed, the tumor volume was measured
with an electronic Vernier caliper (Vmm3=1/2×length=x
width) before each drug administration, and the mental state of the
mice was observed. One week after the last administration, the
final weight and tumor measurement were collected. Mice were
sacrificed by cervical dislocation after taking blood by removing
the eyeball, and necropsy was performed as previously described
(6).
Immunohistochemical evaluation of VEGF
and caspase-3 expression
Sample slides were subjected to positive charge
treatment and the tissue was adsorbed onto the glass slide by
electrostatic action (3,6). Mouse tissues were fixed in 4%
paraformaldehyde for 24 h at room temperature. Tissues were
embedded in paraffin blocks and sectioned at a thickness of 4–5
µm per slice. The sections containing the intact tissue were
then placed in warm water (40°C) for spreading. After the tissue
was fully flattened, samples were dried at 60–65°C for dewaxing.
Tissue slices were then placed in sodium citrate, boiled for 2.5
min, and prepared at high temperature (210°C) and high pressure
(1.8×105Pa) for antigen retrieval. After antigen
retrieval, samples were cooled to room temperature, and the citrate
buffer was removed. Samples were washed twice with water and then
twice with PBS, for 5 min per wash. After drying, the tissue
sections were blocked with 10% NGS for 30 min at 37°C, followed by
incubation in a wet box for 30 min. Samples were then incubated
with primary antibody in 10% NGS (VEGF and apoptosis-related
protein caspase-3 antibodies, dilution ratio 1:100) in the wet box
overnight at 4°C. The following day, samples were placed at room
temperature for 1 h and washed three times with PBST, for 5 min
each wash. Then, samples were incubated with the secondary antibody
horseradish peroxidase labeled goat-anti-mouse IgG (diluted 1:500)
for 2 h at room temperature. After incubation with the secondary
antibody, the sample was washed three times with PBS for 5 min per
wash. According to the kit instructions, 50 µl concentrated
DAB solution was mixed with 1 ml of A solution. DAB coloring
solution was added to the washed slices and slices were incubated
for 5 min at room temperature. The color change of the tissue was
observed, and the tissue sample was placed in distilled water to
stop the reaction. The colored slices were counterstained with
hematoxylin for 30 sec at room temperature. The samples were
dehydrated with gradient diluted absolute ethanol (70, 80, 90, 95
and 100% twice) and then sealed. The sections were observed by
light microscope (magnification, ×200 and ×400).
Statistical analysis
The results were processed by Microsoft Excel
version 2016 (Microsoft Corporation), Origin version 7.5
(Originlab), and SPSS version 17 (SPSS Inc.). All statistical
comparisons were performed using SPSS and presented as the mean ±
standard error of the mean, (SEM) of three or more samples. One-way
ANOVA with Dunnett's post hoc test to compare with the control
group. P<0.05 was considered to indicate a statistically
significant difference.
Results
Amino acid sequence analysis of the
polypeptides
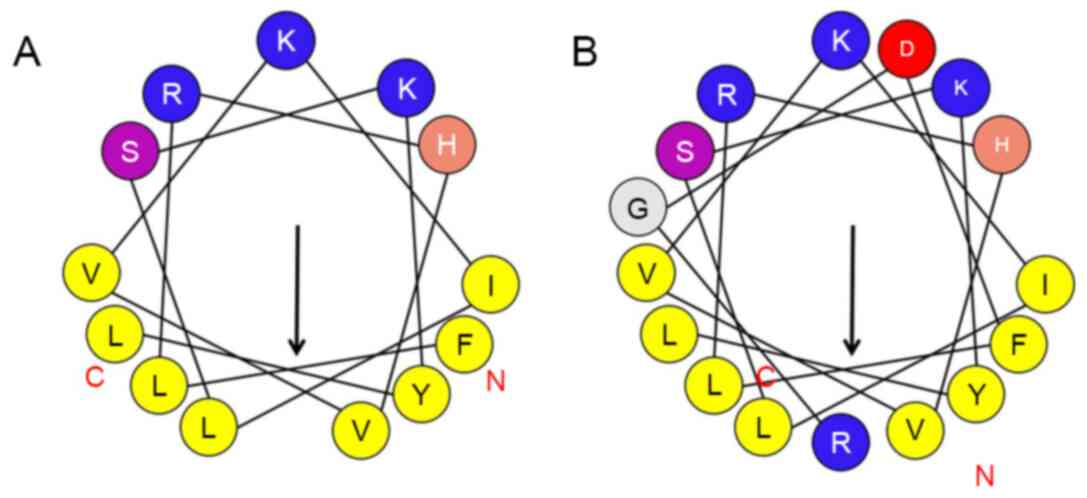
A novel antitumor peptide and an RGD-chimeric
peptide were designed and synthesized by analyzing the physical and
chemical properties, molecular structure and amphipathicity of the
sequence of the bullfrog antibacterial peptide T-La. The sequence,
protein structure and spiral properties (Fig. 1) of T-La were evaluated with analysis
software to obtain the net charge, hydrophobicity, hydrophobic
torque and amphiphilic parameters of several polypeptides (Table I). The net charge number of T-La was
+2. After sequence modification, the first leucine (L) and the
tenth glutamic acid (E) were replaced with phenylalanine (F) and
serine (S), respectively, resulting in a net charge of +3. The
result of the substitution was an increase in the average
hydrophobicity from 0.654 to 0.707 and a decrease in amphipathicity
from 0.754 to 0.733.
 | Table I.Sample name, sequence and
physicochemical properties of the antitumor peptides. |
Table I.
Sample name, sequence and
physicochemical properties of the antitumor peptides.
| Items | T-La | T-La (FS) | RGD-T-La | RGD-T-La (FS) |
|---|
| Sequence | LLRHV | RGD-LLRHV | F | RGD-F |
|
| VKILE |
| LRHVVKIL | LRHVVKIL S |
|
| KYL-NH2 | VKILEKYL-NH2 | S KYL-NH2 | KYL-NH2 |
| Amino acid
number | 13 | 13 | 16 | 16 |
| Molecular
number | 1,624.05 | 1,616.03 | 1,952.37 | 1,944.35 |
| Isoelectric
point | 9.70 | 10.29 | 9.70 | 10.28 |
| Total residue of
negative charge | 1 | 0 | 2 | 1 |
| Total residue of
positive charge | 3 | 3 | 4 | 4 |
| Net charge | 2 | 3 | 2 | 3 |
| Instability
index | 44.93 | 44.93 | 33.08 | 28.36 |
| Stability | Unstable | Unstable | Stable | Stable |
| Aliphatic
index | 194.62 | 164.62 | 158.12 | 133.75 |
| Total average
hydrophilicity | 0.600 | 0.731 | −0.037 | 0.069 |
| Hydrophobic, H | 0.654 | 0.707 | 0.420 | 0.463 |
| Hydrophobic torque,
µH | 0.754 | 0.733 | 0.594 | 0.578 |
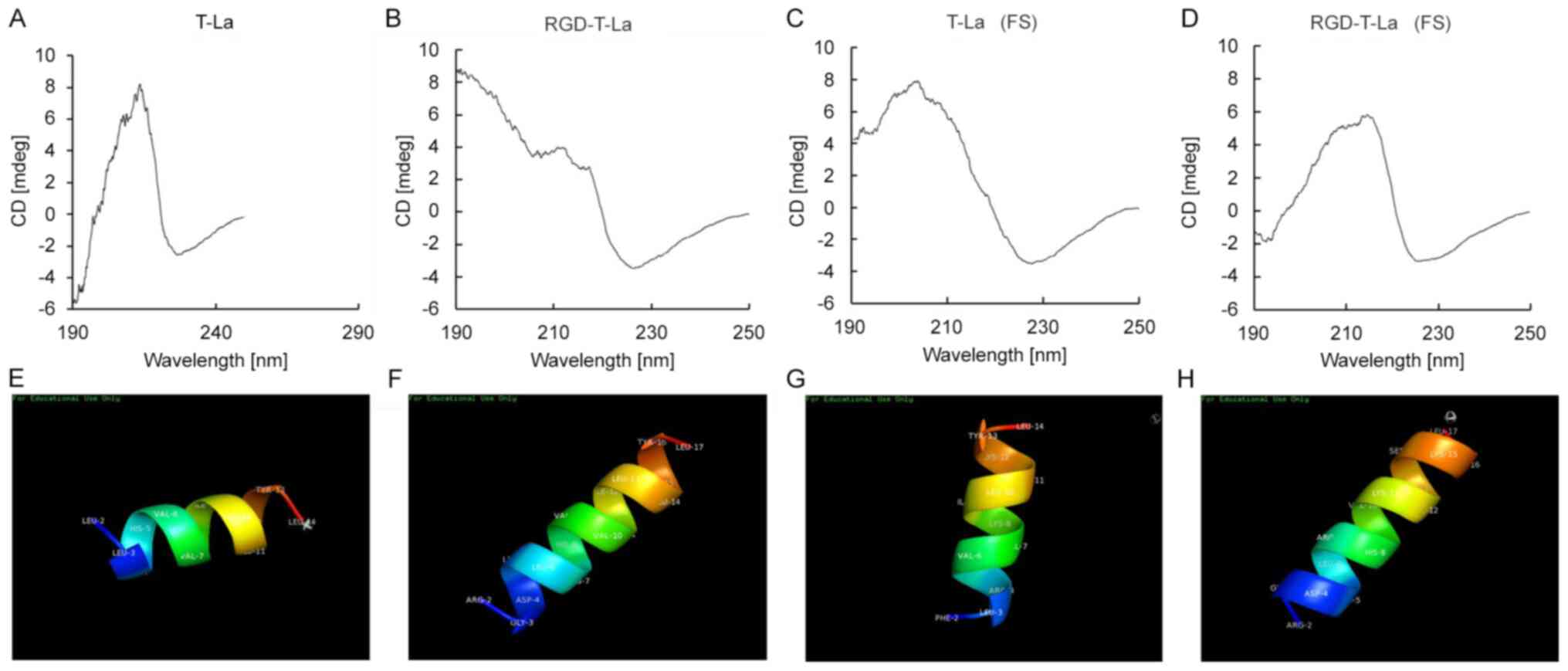
Determination of the secondary
structure and 3D structure of antitumor T-La peptides by CD
spectrum analysis
The secondary structures of the four peptides
derived from T-la were evaluated using CD spectrum (190-250 nm)
analysis (Fig. 2). CD spectrum
analysis revealed that T-La (FS), the two RGD-chimeras and the
bullfrog skin antitumor peptide T-La had α-helix structures,
similar to the natural antitumor peptide T-La (4,6,7). The sequences of the four antitumor
peptides were imported into chemical structure simulation software
and visualized using the PyMoL Molecular Graphics system. The
simulated 3D α-helical structures of T-La, T-La (FS) and the
chimeric peptides, RGD-T-La and RGD-T-La (FS), are shown in
Fig. 2E, F, G and H,
respectively.
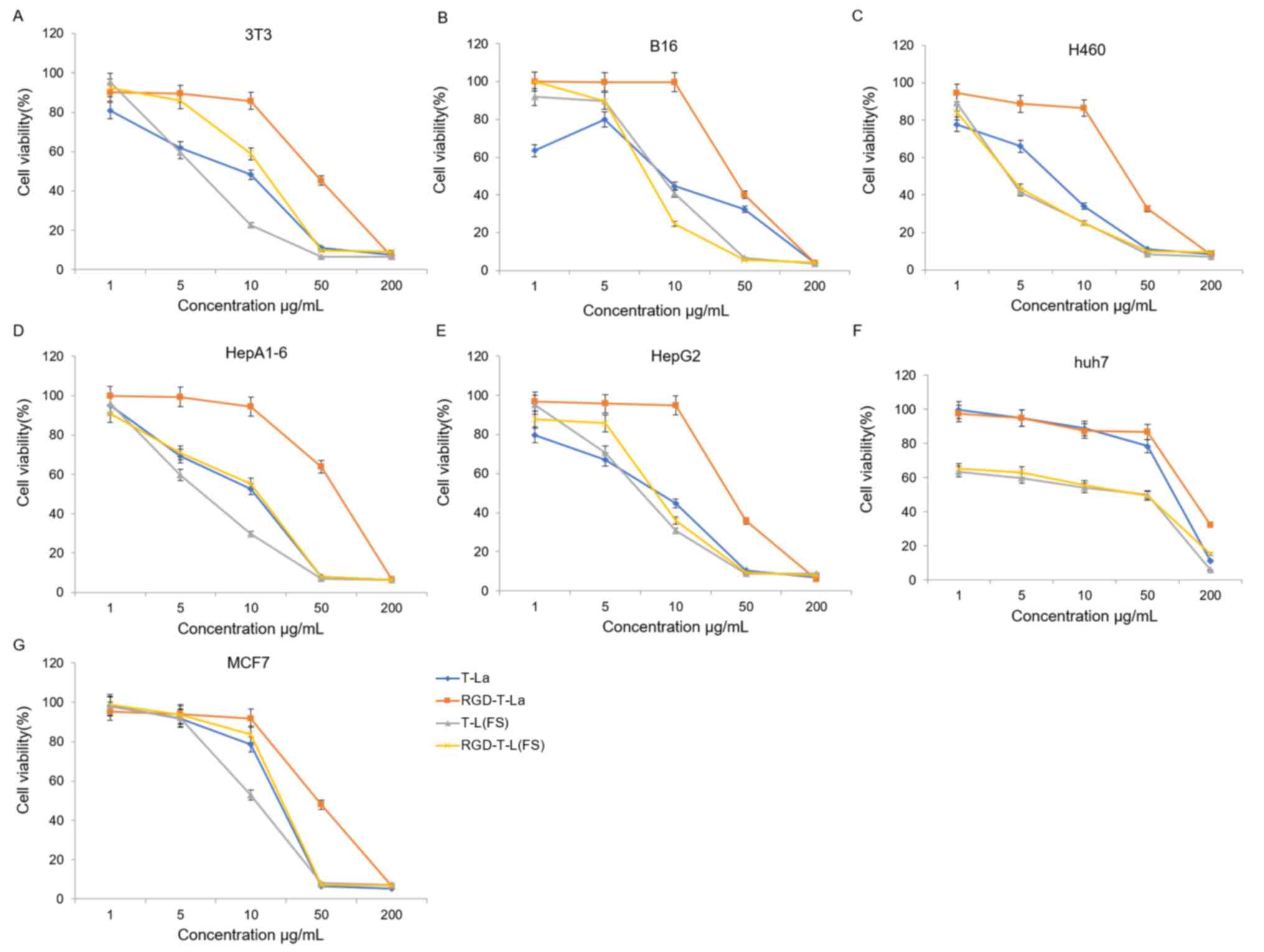
In vitro cytotoxicity of anti-tumor
peptides
The four antitumor peptides were evaluated for in
vitro cytotoxicity in seven tumor cell lines: B16, huh7, 3T3,
HEPA1-6, H460, HepG2 And MCF7. The dose-dependent effects of the
antitumor peptides and the RGD chimeric peptides on cell viability
in the seven tumor cell lines are shown in Fig. 3. All peptides, T-La, RGD-T-La, T-La
(FS) and RGD-T-La (FS), displayed dose-dependent cytotoxicity. For
certain tumor cells, such as B16 and Huh7, the effectiveness of
T-La (FS) was higher compared with T-La, and RGD-T-La (FS) was
higher compared with RGD-T-La. Among the peptides, RGD-T-La (FS)
had the strongest effect on B16 cells, resulting in the lowest cell
viability (24.65%; Fig. 3B).
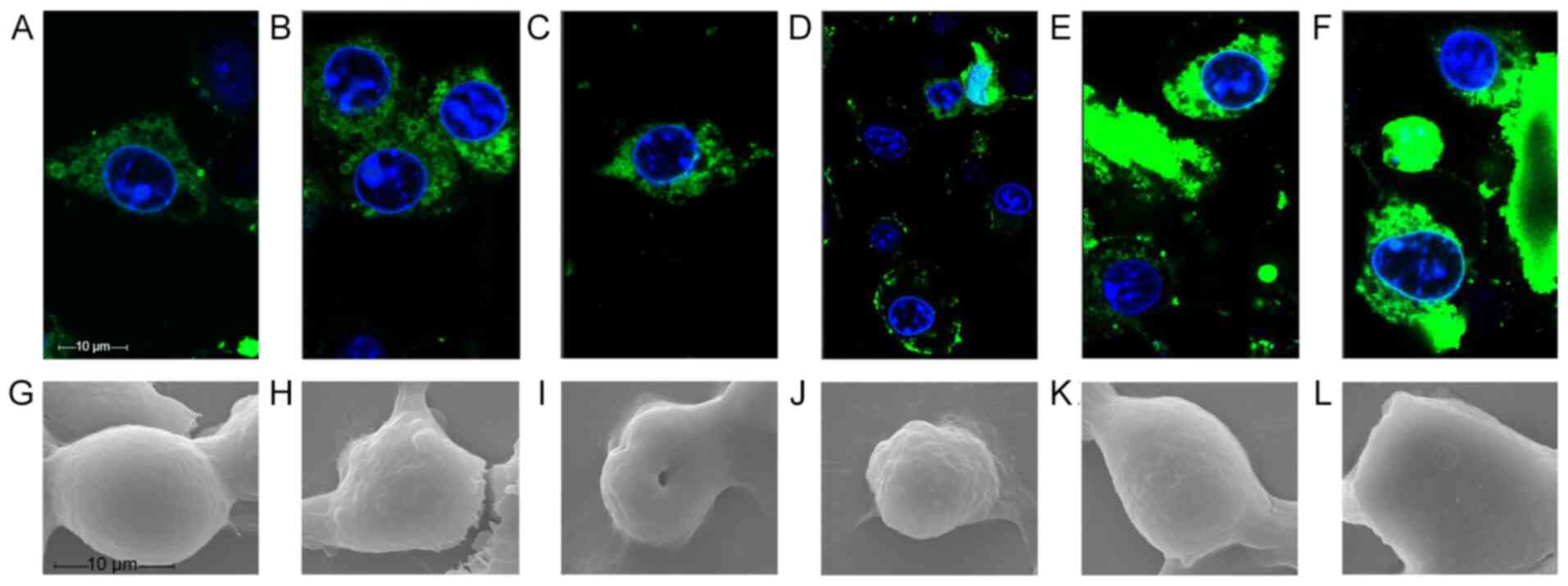
Disruption of tumor cell morphology
and evaluation of the cytotoxic mechanism of the RGD-chimera
peptides in tumor cells
After 20 min of treatment, both fluorescently
labeled polypeptides had entered the cells when observed by
confocal microscopy (Fig. 4). At 70
min, the fluorescence profile of FITC-RGD-T-La (FS)-infected cells
was incomplete and cellular debris appeared around the nucleus,
indicating that FITC-RGD-T-La (FS) disrupted the cell membrane of
B16 cells (Fig. 4D). By 110 min,
intact cellular morphology was difficult to detect and nuclear
shrinkage occurred in cells treated with FITC-RGD-T-La (FS)
(Fig. 4A-F).
B16 cells treated with the polypeptide were
evaluated by scanning electron microscopy. The structure of the B16
cells changed significantly with increasing polypeptide
concentration (Fig. 4G-L). B16 cells
treated with 50 µg/ml RGD-T-La (FS) exhibited cell shrinkage
and breakage between cells (Fig.
4H). Treatment of B16 cells with 200 µg/ml RGD-T-La (FS)
resulted in the isolation of individual cells, cytoplasm leakage,
loss of intact cell morphology and central cell depression
(Fig. 4I). Treatment of B16 cells
with 50 µg/ml T-La (FS) resulted in disruption in morphology
and partial loss of cytoplasm (Fig.
4K). There were few cellular features visible in B16 cells
treated with 200 µg/ml T-La (FS) (Fig. 4L).
Detection of cell surface expression
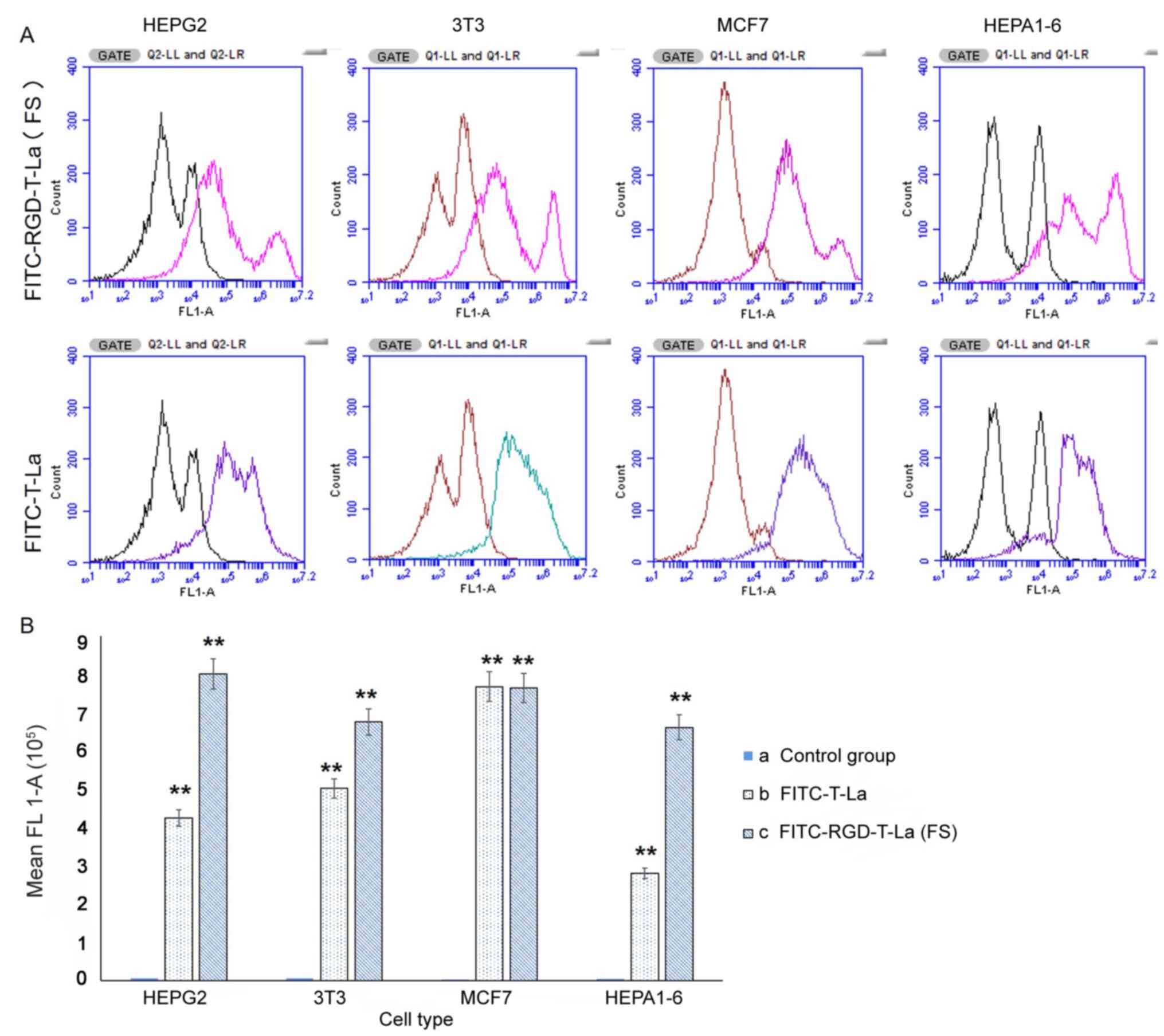
of integrin αvβ3
Expression of integrin αvβ3 was evaluated on the
cell surface of MCF7, HEPA1-6, HepG2 and 3T3 cells using flow
cytometry. The same number of events was recorded for each cell
line (1×106). Fluorescent profiles were different
amongst cell lines, indicating differences in expression of
integrin αvβ3 on the cell surface (Fig.
5A). The absorption intensity for HepG2, MCF7, 3T3 and HEPA1-6
cells for FITC-RGD-T-La (FS) was 800,950.70, 763,824.54, 675,429.46
and 661,462.48, respectively; the absorption intensity for
FITC-T-La was 424,694.19, 767,889.04, 502,185.53 and 279,975.88,
respectively. Absorption intensity in the blank control group was
5,773.83, 3,836.19, 6,755.40 and 4,793.72, respectively (Fig. 5B).
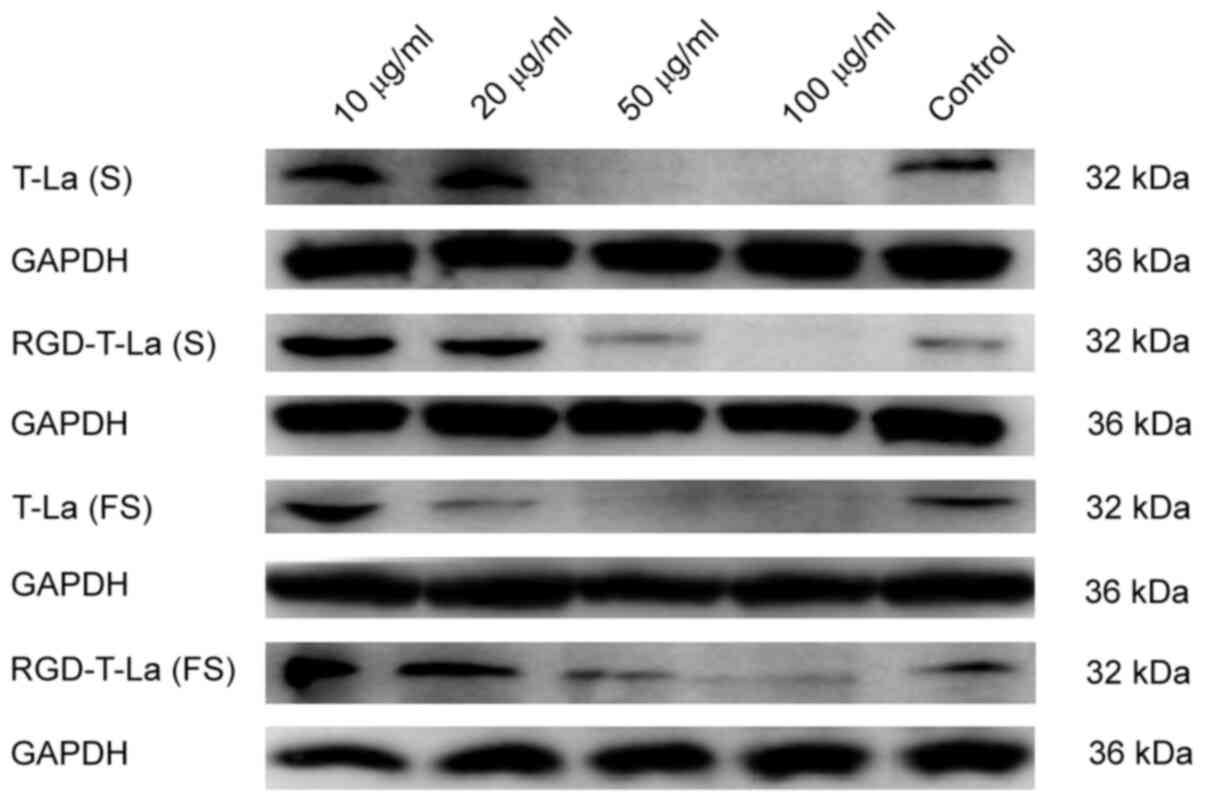
Western blot analysis of caspase-3
protein expression
B16 melanoma cells were treated with 0, 10, 20, 50
and 100 µg/ml of peptide for 24 h. A representative western
blot for apoptotic protein caspase-3 is shown in Fig. 6 and quantification is shown in
Fig. 7. Expression of caspase-3 was
more abundant after RGD-T-La (FS) treatment in B16 cells at 10 and
20 µg/ml compared with the control group and the expression
of caspase-3 was significantly higher compared with other peptides
groups at 50 µg/ml. With increasing peptide concentration,
the expression of caspase-3 protein gradually decreased, which may
have led to cell death.
RT-qPCR analysis of apoptotic gene
caspase-3 mRNA expression
The effects of peptide treatment on the expression
of caspase-3 mRNA in B16 cells were determined using qPCR. Compared
with the control group, 10 µg/ml of RGD-T-La (S), RGD-T-La
(FS) and T-La (FS) induced significantly higher expression of
caspase-3 apoptotic gene mRNA in B16 cells (P<0.05) (Table II). These results were consistent
with the western blotting results. However, treatment with T-La (S)
did not significantly alter caspase-3 mRNA levels compared with
controls (Table II).
| Table II.Effect of T-La (S) and RGD-T-La (S)
on mRNA expression of apoptosis gene in B16 cells. |
Table II.
Effect of T-La (S) and RGD-T-La (S)
on mRNA expression of apoptosis gene in B16 cells.
| Peptide | Caspase-3
expression (mean±SEM) |
|---|
| Control |
0.62±0.04 |
| T-La (S),
µg/ml |
|
| 10 |
0.83±0.13 |
| 20 |
0.78±0.20 |
| 50 |
0.61±0.09 |
| RGD-T-La (S),
µg/ml |
|
| 10 |
1.32±0.04b |
| 20 |
1.06±0.22a |
| 50 | 0.42±0.1 |
| T-La (FS),
µg/ml |
|
| 10 |
1.37±0.09b |
| 20 |
0.39±0.08 |
| 50 |
0.25±0.16 |
| RGD-T-La (FS),
µg/ml |
|
| 10 |
1.27±0.12a |
| 20 |
0.81±0.16 |
| 50 |
0.35±0.12 |
Distribution of antitumor peptide
RGD-T-La (FS) in different organs and tumor tissues
After 30 min post tail vein injection, the antitumor
peptide, RGD-T-La (FS), was distributed in the tumors, liver, lung
and kidney of nude mice. After 1 h post injection, fluorescence was
observed in the heart, liver, lung, kidney and tumors, but the
fluorescence intensity was strongest in the tumors. After 4 h post
injection, the fluorescence in tumors was higher compared with in
the fluorescence in heart, liver, spleen, lung and kidney. After 8
h, the fluorescence in heart, liver, spleen, lung and kidney
disappeared, but fluorescence was still observed in the tumor
tissues. In the T-La (FS) group and PBS group, no fluorescence was
observed (Fig. 8).
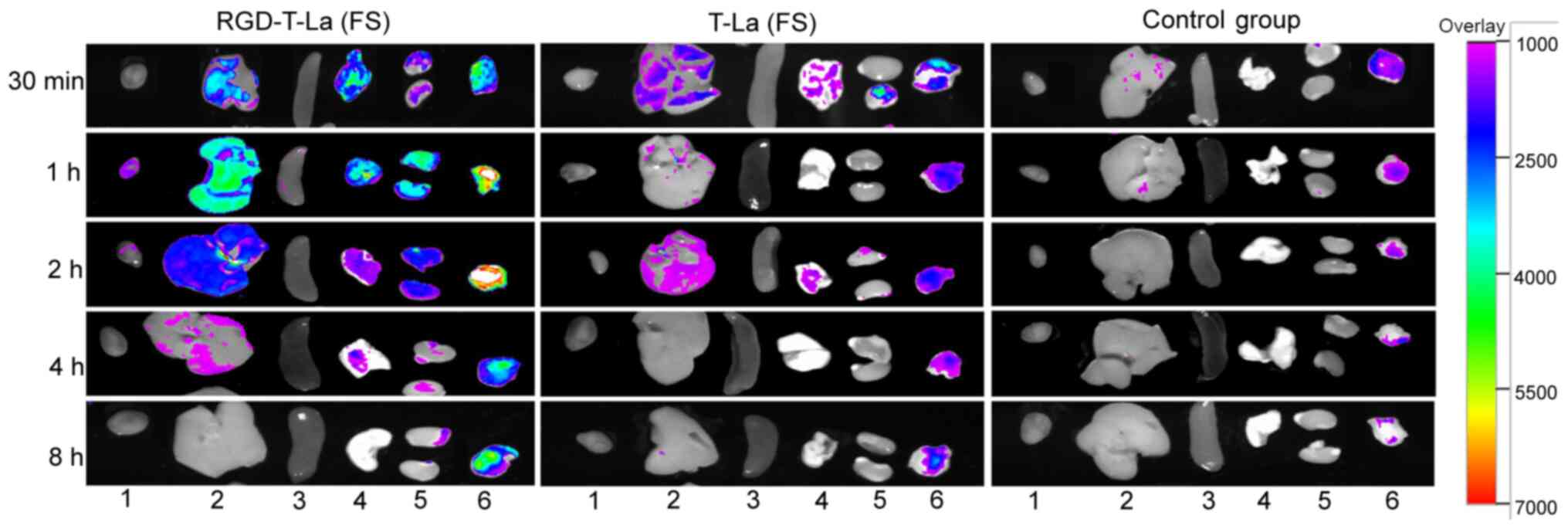 | Figure 8.Distribution of antineoplastic
peptides in different organs and tumor tissues of nude mice at
different times. Heart, liver, spleen, lung, kidney and tumor
tissues are denoted by 1, 2, 3, 4, 5 and 6, respectively. Change of
color in the tissue represents the intensity of fluorescence, and
purple to red indicates that the fluorescence intensity changes
from weak to strong. T-La, Temporin-La; (FS), antitumor. |
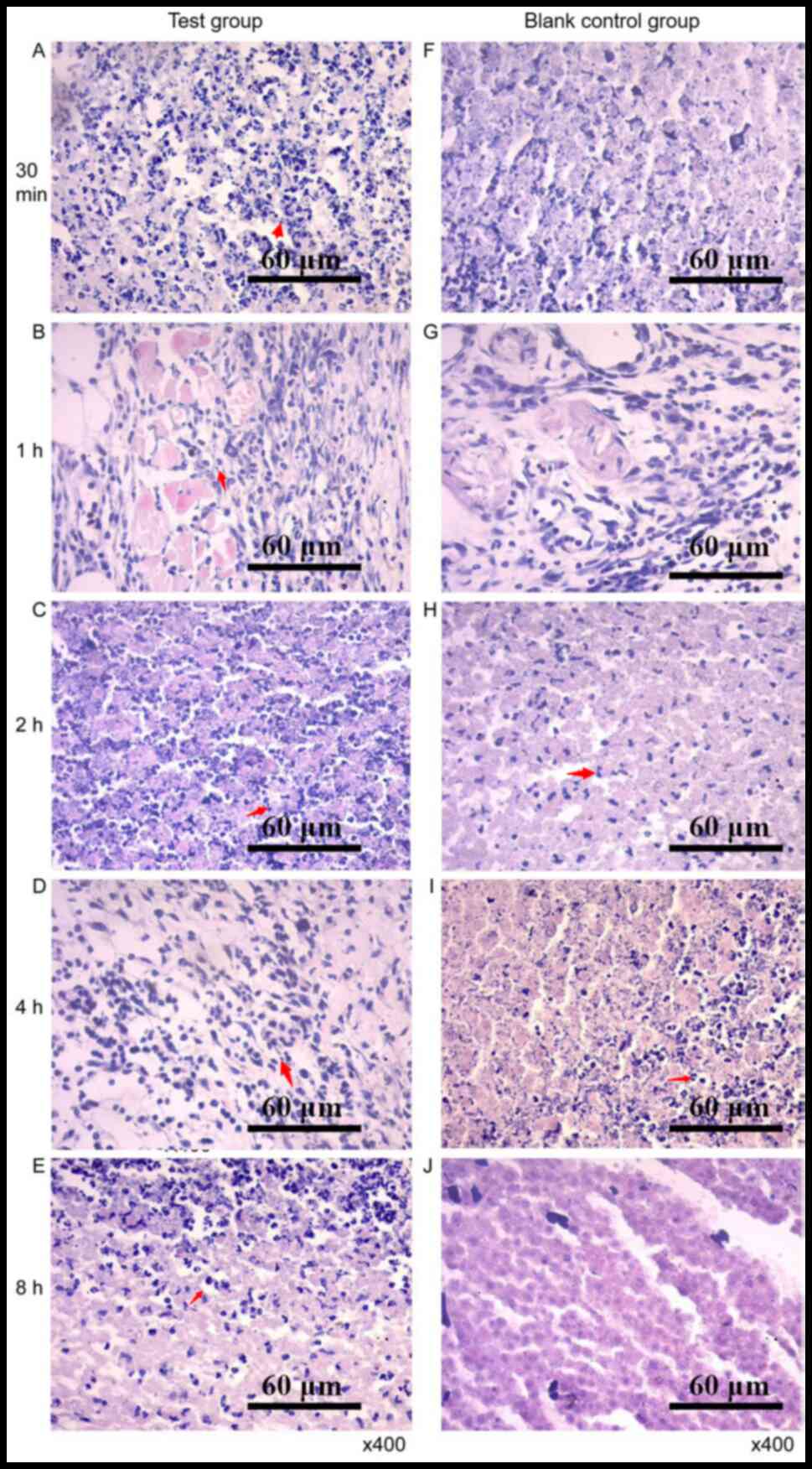
Histopathological observation of tumor
tissue
After RGD-T-La (FS) injection, tumor cell necrosis
was observed. The cellular structure was broken down and
fragmentation of the nucleus was observed. In the control group,
tumor cells with blurred margins, degeneration, necrosis and
mitotic phase were observed (Fig.
9). In addition, the four peptides had no side effects on
normal nude mice tissues, and the results of tissue sections are
shown in Figs. S1–S4.
Effects of antitumor peptides on mouse
body weight and melanoma growth
Body weights did not differ significantly between
the groups before treatment (Table
III). Treatment with the peptides resulted in positive weight
gain in tumor-bearing mice (Table
III). After the first four injections, the average weight gain
in each peptide treatment group was significantly higher compared
with that in the cisplatin and PBS groups (all P<0.05). One week
after the fifth drug administration, weight gain between the
treatment groups was not significantly different (P>0.05).
| Table III.Comparison of body weight before and
48 h after injection of nude mice with antitumor drugs. |
Table III.
Comparison of body weight before and
48 h after injection of nude mice with antitumor drugs.
|
|
| Weight increase
after administration, g |
|---|
|
|
|
|
|---|
| Drug | Initial weight,
g | First | Second | Third | Fourth | Fifth |
|---|
| T-La (FS) | 20.540± |
0.850± |
0.850± |
0.817± |
1.017± | 2.533± |
|
|
0.3855a |
0.1839a |
0.1821a |
0.3229a |
0.3038a |
0.6800a |
| RGD-T-La (FS) | 20.360± |
0.667± |
0.783± |
1.017± |
1.467± | 0.483± |
|
|
0.2874a |
0.1382a,b |
0.1138a |
0.3229a |
0.3792a |
1.4363a |
| T-La | 19.620± |
0.300± |
0.917± |
1.333± |
0.200± | 1.917± |
|
|
0.4128a |
0.2295a,b |
0.3449a |
0.3229a |
0.4933a |
0.4936a |
| RGD-T-La | 19.940± |
0.400± |
0.383± |
0.983± |
1.400± | 0.883± |
|
|
0.5680a |
0.1000a,b |
0.2496a |
0.3229a |
0.3286a |
0.3919a |
| Cisplatin positive
control | 19.680± | −0.050± | −0.950± | −1.983± | −3.683± | NA |
|
|
0.5152a |
0.3149b |
0.3871b |
0.3229b |
0.5043b |
|
| PBS blank
control | 20.400± | −1.067± | −0.667± |
0.017± | −2.017± | 1.267± |
|
|
0.4050a |
0.4917c |
0.4780b |
0.3229b |
0.4708c |
0.8123a |
As shown in Table
IV, the initial tumor volumes in each experimental group were
not significantly different (P=0.306). After the first treatment,
the tumor volume increased significantly in the PBS control group
compared with the other treatment groups (all P<0.05). There
were no significant differences in tumor volume changes between the
antitumor peptide groups and the cisplatin treatment group (all
P>0.05). After the second treatment, the tumor volume increase
in the RGD-T-La group was significantly smaller compared with the
PBS group P=0.018) (Table IV). The
tumor volume increases in the other treatment groups were not
significantly different (all P>0.05; Table IV).
| Table IV.Melanoma tumor volume in nude mice
before and 48 h after injection of anti-tumor drugs |
Table IV.
Melanoma tumor volume in nude mice
before and 48 h after injection of anti-tumor drugs
|
|
| Tumor volume, |
|---|
|
|
|
|
|---|
| Item | Primary tumor
volume, mm3 | First | Second | Third | Fourth | Fifth |
|---|
| T-La (FS) | 21.1022± | 25.3020± | 19.8162± | 20.6351± | 27.0552± | 134.1800± |
|
|
2.48719a |
3.04705b |
2.69010a,b |
3.42328a |
5.39173a |
14.22572a |
| RGD-T-L (FS) | 38.9477± | 38.5640± | 25.6673± | 12.9156± | 32.1026± |
82.8592± |
|
|
11.16416a |
6.22073b |
1.93695a |
15.98864a |
11.98846a |
25.75526a |
| T-La | 26.6849± | 33.3320± | 19.5228± | 20.0724± | 32.6486± | 100.7889± |
|
|
5.36854a |
8.72730b |
3.78134a,b |
3.76676a |
5.07784a |
10.08681a |
| RGD-T-La | 24.3425± | 27.9720± | 10.7225± | 18.8990± | 32.6512± |
83.0283± |
|
|
3.35537a |
5.69018b |
2.03661b |
2.16945a |
5.11254a |
18.52589a |
| Cisplatin positive
control | 23.9659± | 34.4100± | 14.2439± | 13.1352± |
3.5965± | / |
|
|
5.18682a |
7.04033b |
4.04242a,b |
5.14370a |
4.20833b |
|
| PBS blank
control | 32.2281± | 62.0960± | 25.9056± | 25.8578± | 22.9230± | 114.2617± |
|
|
2.09646a |
5.50607a |
7.16902a |
4.13521a |
3.37649a |
13.95786a |
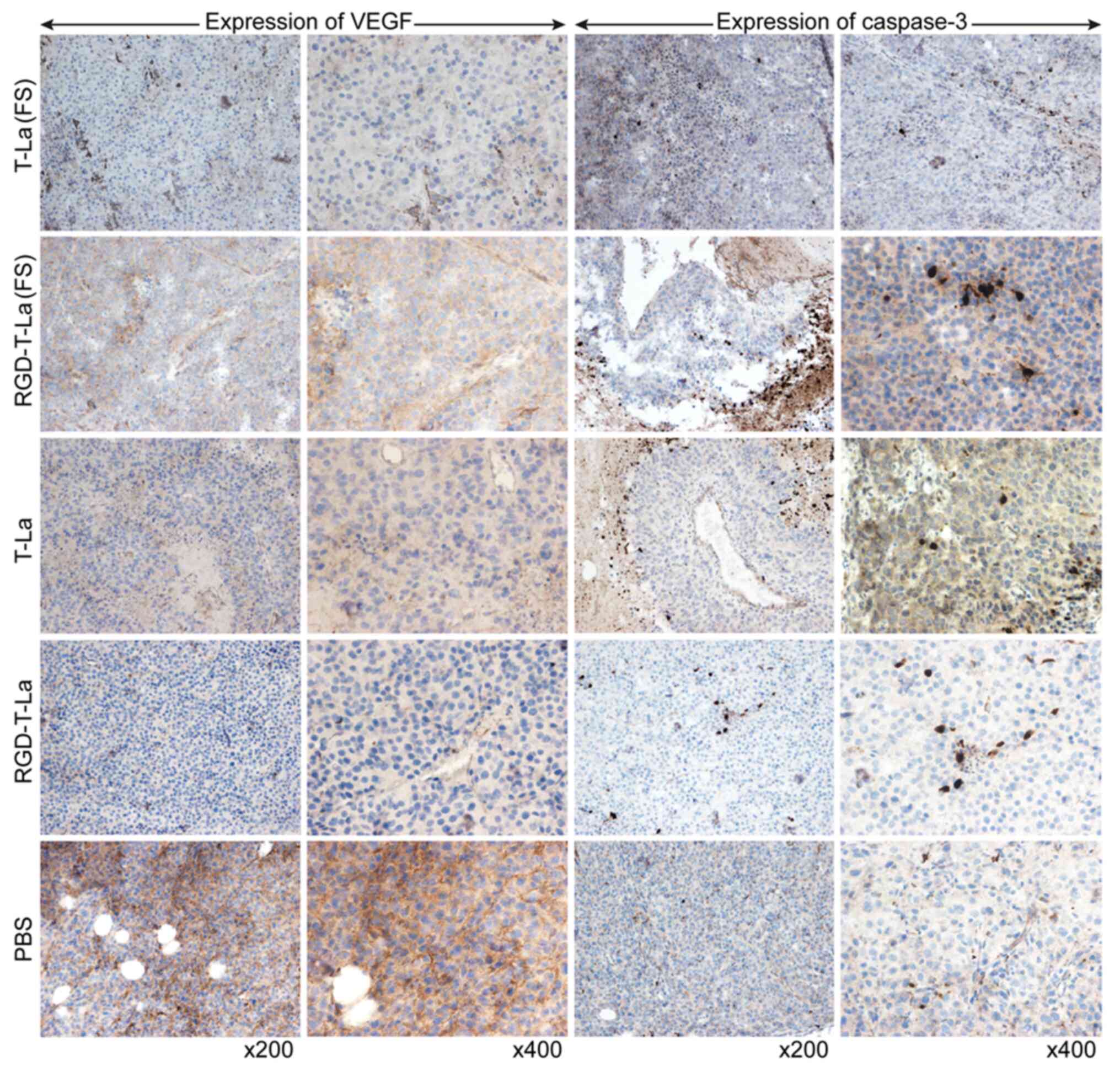
Evaluation of VEGF and caspase-3
expression using immunohistochemistry
Evaluation of tumor sections using
immunohistochemistry showed that, in melanoma cells, VEGF was
expressed mostly on the tumor cell membrane surface. VEGF
expression was higher in superficial tumor cells compared with in
cells toward the center of the tumor. Caspase-3 was primarily
expressed in the nucleus and around necrotic foci in tumor nodules.
The expression of VEGF was lower in all peptide treatment groups
[T-La (FS), RGD-T-La (FS), T-La and RGD-T-La] compared with the PBS
control group (Fig. 10). Expression
of caspase-3 was higher in the antitumor peptide treatment groups
[T-La (FS), RGD-T-La (FS), T-La and RGD-T-La] compared with in the
PBS treatment control group (Fig.
10).
Discussion
Analysis of the structure and activity of a large
number of antimicrobial peptides has revealed that the peptide
structure is related to its activity (15). A recent study has shown that the
positive charge number, hydrophobicity, α-helix, amphoteric
molecular structure, disulfide bonds and other factors of
antimicrobial peptides have important effects on their activity
(16). The present study focused on
four peptides, including T-La, T-La (FS) and the chimeric peptides,
RGD-T-La and RGD-T-La (FS). Glutamate (E) is a negatively charged
amino acid and serine is an uncharged polar amino acid (5). Both leucine and phenylalanine are
uncharged non-polar hydrophobic amino acids (5). Increased hydrophobicity has been shown
to increase antimicrobial activity of peptides (17). Likewise, an increase in
amphiphilicity can increase the activity of antimicrobial peptides
(18). In the present study, the
antitumor activity of T-La (FS) was higher compared with that of
T-La, and the antitumor activity of chimeric RGD-T-La (FS) was
higher compared with that of chimeric RGD-T-La. The effects of four
polypeptides at different concentrations on different cells were
compared, and it was demonstrated that the increase in charge was
the main reason for differences in antitumor activities of the
polypeptides. The increase in net charge increased the
concentration of polypeptide on the cell membrane surface, and
differences in hydrophobicity and amphiphilicity also contributed
to differences in antitumor activity. The present study provides a
scientific basis for the application of small molecule
antimicrobial peptides as antitumor agents. Furthermore, the
current study established the theoretical foundation for the
development and clinical application of these peptides as antitumor
drugs. By combining the targeting effect of the RGD peptide with
the antitumor effect of the T-La peptide, the antitumor effects
were improved.
Observation of polypeptide effects in real time
using laser confocal microscopy revealed that the peptides elicited
a very rapid effect on cells. The mechanism of action of the
polypeptides and RGD chimeras was observed using scanning electron
microscopy, and it was reported that the RGD-conjugated chimeric
peptide caused loss of cytoplasm and cell death by rapidly acting
on a specific target on the cell membrane. The non-chimeric
polypeptide acted on the entire cell membrane surface, causing cell
membrane lysis and cell death.
Integrin proteins on the surface of most tumor cell
membranes are transmembrane receptors that mediate cross-talk
between tumor cells and their external environment (19). In the process of contacting tumor
cells and the external environment, integrins not only play a
transmembrane role, but also participate in the transmission of
signals between cells and their external environment (20). Integrin αvβ3 serves an important role
in tumor angiogenesis and is highly expressed on a variety of tumor
surfaces and neovascular endothelial cells (21). Targeting αvβ3 for the detection and
diagnosis of a variety of tumors, such as melanoma has been widely
evaluated and used (22). The RGD
small molecule peptide used in the present study is a ligand for
integrin αvβ3 and was conjugated to the engineered antitumor
peptides to enhance the targeting of the peptides to tumor cells.
The current data may aid the development of targeted antitumor
drugs in this regard.
The weights of tumor-bearing nude mice in the
antitumor peptide treatment groups demonstrated an increasing trend
after the first three doses, while the weights of mice treated with
the cisplatin positive control and PBS negative control decreased.
These data suggested that the antitumor peptides had less severe
side effects on animal health. In addition, the four peptides have
no side effects on normal nude mice tissues. The cisplatin
treatment significantly inhibited melanoma tumor growth in nude
mice, but the dose of 100 µl/1 mg/ml caused harm to the
animals, as indicated by the weight loss. These data indicated that
the antitumor peptides exerted their inhibitory effect on melanoma
tumor growth in nude mice during the early stages of cancer
treatment.
Under non-pathological conditions, VEGF is expressed
mainly in the cytoplasm and on the membranes of vascular
endothelial cells (23,24). The expression of VEGF in tumor cells
treated with antitumor peptides was lower compared with that in the
PBS group, indicating that the peptides could effectively inhibit
VEGF expression (25,26). The apoptotic protein caspase-3
promotes apoptosis of tumor cells (27), and higher caspase-3 expression
indicates higher degrees of tumor cell apoptosis (28). The present study reported that
caspase-3 expression in melanoma tissues of nude mice treated with
the antitumor peptides was higher compared with that in the PBS
control group. These data suggested that the antitumor peptides
induce apoptosis through a combined effect on VEGF and caspase-3;
however, more experiments are needed to fully elucidate the
inhibitory effects of the peptides on tumor growth. For example,
repetitive testing of target animals in vivo. Compared with
previously reported articles (5,27,28), the
T-La peptide sequence was modified resulting in stronger antitumor
effects in vitro and in vivo. Coupling the modified
peptides with the RGD peptide increased their targeting ability.
Therefore, the current study highlight the potential of RGD-T-La
(S) and RGD-T-La (FS) as safe drugs in the treatment of
melanoma.
Overall, with the aim to improve the antineoplastic
activity of the antimicrobial peptide T-La from bullfrog skin, the
present study constructed a novel chimeric peptide by altering
several amino acids in the peptide and coupling the peptide to the
small molecular peptide, RGD. The resulting peptide, RGD-T-La (FS),
had altered physical and chemical properties, including changes in
the charge number, isoelectric point and hydrophobicity.
Furthermore, in vitro and in vivo experiments showed
that RGD-T-La (FS) had strong growth inhibitory effects in tumor
cells, acted on tumor tissues in nude mice for a longer time and
had significant inhibitory effects on melanoma.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was funded by grants from The
General Fund of Application Foundation & Advanced Technology
Program, Tianjing, China (grant nos. 14JCYBJC3000 and
18JCYBJC30100), The Veterinary ‘Innovative Talent People of Young
and Middle Age Key Members Project’ of Tianjin, China, The National
Natural Science Foundation of China (grant no. 31201886, 31572492)
and The ‘131’ Innovative Talents Training Project Third Level
Candidate Program, Tianjin.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ML and RZ participated in the study design, analyzed
the data and drafted the manuscript. ML and XJ carried out most of
the experiments and collected the data. CF and TJ were involved
with study design and analyzing data. TJ was involved in drafting
the manuscript. JM, SQ, LAL, YH and XZ analyzed data. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments were approved by The
Laboratory Animal Ethics Committee of Tianjin Medical University
(approval no. TMUaMEC 2018036) (Tianjin, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
T-La
|
Temporin-La
|
|
FITC
|
fluorescein isothiocyanate
|
|
T-La (FS)
|
Temporin-La (FS)
|
References
|
1
|
Li S, Hao L, Bao W, Zhang P, Su D, Cheng
Y, Nie L, Wang G, Hou F and Yang Y: A novel short anionic
antibacterial peptide isolated from the skin of Xenopus
laevis with broad antibacterial activity and inhibitory
activity against breast cancer cell. Arch Microbiol. 198:473–482.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhang F and Li MS: Antibacterial peptides'
roles in immune defense and anti-tumorogenesis. World Not Antibiot.
38:284–289. 2017.(In Chinese).
|
|
3
|
Gao J, Xie C and Lu WY: Progress in
research of antitumor activity of antimicrobial peptides. World
Clinical Drugs,. 10:122012.
|
|
4
|
Zhu LZR, Liu SS, Ma JF, Jin TM, Liu JF, Hu
Y and Liu MY: Biological activity and in vivo tissue targeting
distribution of catesbeianalctin. Chin J Vet Sci. 36:185–190.
2016.(In Chinese).
|
|
5
|
Diao Y, Han W, Zhao H, Zhu S, Liu X, Feng
X, Gu J, Yao C, Liu S, Sun C, et al: Designed synthetic analogs of
the α-helical peptide temporin-La with improved antitumor
efficacies via charge modification and incorporation of the
integrin αvβ3 homing domain. J Pept Sci. 18:476–486. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ma L, Cai ZL, Wang QR and Lei CX: Study on
construction of citrostatin and its bioactivity. Chin J Biochem
Pharm. 31:19–22. 2010.
|
|
7
|
Huang R, Long J and Zhang Y: Synthesise
and activity of dual targeted antineoplastic polypeptide
RGDSY-CTTHWGFTLC. Cancer Res Prev Treat. 41:531–535. 2014.
|
|
8
|
Ma Q, Dong N, Cao Y and Shan A: Rational
design of alpha-helical antimicrobial peptide with Val and Arg
residues. Acta Microbiol Sin. 51:346–451. 2011.
|
|
9
|
Wang XX, Niu B, Chen XJ, Xie J and Zhang
YH: Synthesis and activities of RGD peptide analog. J Chin Pharm
Sci. 43:462–464. 2008.(In Chinese).
|
|
10
|
Liu KY, Li QW and Liu GY: Study progress
of malignant tumor imaging of integrin αvβ3 expression with
radiolabelled RGD peptides. Medical Recapitulate. 12:757–759.
2006.(In Chinese).
|
|
11
|
Zhang CL, Yang M and Wang RF: The
structure-activity relationship of RGD peptides binding to αvβ3
integrin and radiolabeled ligand design. J Oncol. 15:76–81.
2009.
|
|
12
|
Yu YP, Wang Q, Liu YC and Xie Y: Molecular
basis for the targeted binding of RGD-containing peptide to
integrin αvβ3. Biomaterials. 35:1667–1675. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ji QZ and Zhang KW: Observation of DFRKN-1
isolate by scanning electron microscope. Anhui Agric Sci. 39:33–34.
2011.
|
|
14
|
Seo GY, Ha Y, Park AH, Kwon OW and Kim YJ:
Leathesia difformis extract inhibits α-MSH-induced
melanogenesis in B16F10 cells via down-regulation of CREB signaling
pathway. Int J Mol Sci. 20:E5362019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang CL, Wang RF and L Z: Design of
~(131)I labeling of a disulfide bridged RGD-peptide targeted to
integrin αvβ3 receptor and its biodistribution and imaging in tumor
bearing mice. J Oncol. 16:276–280. 2010.
|
|
16
|
Dia YW: Design for tumor-targeting
antimicrobial peptide chimera and its anticancer mechanism study
(unpublished PhD thesis). Jilin University; 2012
|
|
17
|
Li ZDLY and Zhen YS: Potentiation of
boanmycin antitumor activity by chemotactic peptide. Chin J Cancer
Res. 17:79–83. 2005.(In Chinese). View Article : Google Scholar
|
|
18
|
Zhao RL, Han JY, Han WY, Liu SS, Ma JF,
Lei LC, Feng X, Zhang AG and Wang X: The antibacterial and
antitumor function of a novel bio-active peptide Catesbeianin-1a of
Rana catesbeiana. Chin J Vet Sci. 33:1407–1411. 2013.(In
Chinese).
|
|
19
|
Wang LZ and Min Y: Advance of” 1” 8F
labeled RGD peptide as integrin α_Vβ_3 receptor imaging agents. J
Isot. 24:68–75. 2011.
|
|
20
|
Jiang Z, Vasil AI, Hale JD, Hancock RE,
Vasil ML and Hodges RS: Effects of net charge and the number of
positively charged residues on the biological activity of
amphipathic α-helical cationic antimicrobial peptides. Biopolymers.
90:369–383. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang J, Xiao X, Zhu J, Gao Z, Lai X, Zhu
X and Mao G: Lactoferrin- and RGD-comodified, temozolomide and
vincristine-coloaded nanostructured lipid carriers for gliomatosis
cerebri combination therapy. Int J Nanomedicine. 13:3039–3051.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen T, Chen L, He H, Peng J, Guo X, Zheng
Y and Shao C: Influence of HAnps carrier system with doping Mg and
grafting RGD on endocytosis of MG63 cells. J Cent South Univ.
48:625–634. 2017.
|
|
23
|
Habibi N, Kamaly N, Memic A and Shafiee H:
Self-assembled peptide-based nanostructures: Smart nanomaterials
toward targeted drug delivery. Nano Today. 11:41–60. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lambert E, Fuselier E, Ramont L, Brassart
B, Dukic S, Oudart JB, Dupont-Deshorgue A, Sellier C, Machado C and
Dauchez M: Conformation-dependent binding of a Tetrastatin peptide
to αvβ3 integrin decreases melanoma progression through
FAK/PI3 K/Akt pathway inhibition. Sci Rep. 8:98372018.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hou J, Diao Y, Li W, Yang Z, Zhang L, Chen
Z and Wu Y: RGD peptide conjugation results in enhanced antitumor
activity of PD0325901 against glioblastoma by both tumor-targeting
delivery and combination therapy. Int J Pharm. 505:329–340. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen G, Yu F, Sheng G, Wang J and Qu B:
Progress and future direction of vascular mimicry. J Mod Oncol.
1:138–141. 2017.
|
|
27
|
Cao QZ and Lin ZB: Antitumor and
anti-angiogenic activity of Ganoderma lucidum polysaccharides
peptide. Acta Pharmacol Sin. 25:833–838. 2004.PubMed/NCBI
|
|
28
|
Ren HD, Zhang L, Zhao Y, Si XH, Zhang L
and Wang C: Study on inhibitory effect of antimicrobial peptide
Temporin-1CEa and its analogues inhibit metastasis and invasion of
human melanoma A375 cells and reduce angiogenesis. Chin J Biochem
Pharm. 2:19–22. 2016.(In Chinese).
|