Introduction
Acute myeloid leukemia (AML) is a heterogeneous
disease that includes several discrete syndromes with
characteristic clinical, morphological, phenotypic and cytogenetic
features (1). According to the
French-American-British (FAB) Co-operative Group, the subgroups of
AML defined in the FAB classification are M0, M1, M2, M3, M4, M5,
M6 and M7 (2). For patients with AML
under 60 years of age who undergo induction chemotherapy, complete
remission (CR) rates of 60–70% are generally obtained, with a
5-year disease-free survival rate of ≤40% (3,4). By
contrast, patients over 60 years of age have CR rates of 40–50%,
with a 3-year disease-free survival rate of <20% (5). Therefore, further characterization of
the pathogenesis of AML is important for the development of new
therapies.
As a type of antisense RNA, microRNAs (miRNAs or
miRs) are a group of small noncoding RNAs of ~22 nucleotides in
length (6), which bind to
complementary sites in the 3′-untranslated regions (3′-UTRs) of
mRNA, and repress translation or induce mRNA degradation (7). Approximately 30% of all human
protein-coding genes are regulated by miRNAs (8), and abnormally expressed miRNAs can
function as tumor suppressors or oncogenes (9,10).
Increasingly, miRNAs have been evaluated in various types of
cancer, and have been found to participate in the regulation of
cancer cell behaviors, including proliferation, apoptosis,
differentiation, metabolism and tumor metastasis (11–13).
Notably, several aberrantly expressed tissue miRNAs have been
regarded as diagnostic indicators in multiple cancer types
(14–16).
Accumulating evidence has demonstrated that various
miRNAs are closely associated with AML, including miR-9 (17), miR-21 (18), miR-34a (19), miR-126 (20), miR-143 (21) and miR-155 (22). Among them, miR-21, which is one of
the best studied miRNAs, is highly expressed in numerous types of
human cancer, including lung, cervical, colorectal, pancreatic and
oral cancer (23–26), and displays oncogenic activities in
various steps of tumorigenesis such as cell invasion, cell
proliferation, metastasis and cell survival (27–29).
Moreover, a meta-analysis summarizing the global predicting role of
miR-21 for survival in patients with a variety of carcinomas in
2011 showed that elevated miR-21 expression is significantly
associated with poor survival (30).
Numerous miR-21 target genes have been identified, including
phosphatase and tensin homolog deleted on chromosome ten (PTEN),
programmed cell death 4 (PDCD4) and B cell translocation gene 2,
which play important roles in the oncogenic process (23,28,31,32). In
particular, several studies have investigated whether miR-21 is
highly expressed in multiple types of leukemia such as chronic
lymphocytic leukemia, chronic myelogenous leukemia, chronic myeloid
leukemia, childhood B cell acute lymphoblastic leukemia and AML
(32–36). In addition, a previous study
demonstrated that miR-21 is overexpressed in nucleophosmin 1-mutant
AML (36). These studies suggested
that miR-21 may play an important role in AML progression.
Recently, Li et al (18)
reported that miR-21 promoted proliferation through directly
regulating Kruppel-like factor 5 expression in AML cells. However,
the detailed regulatory mechanisms of miR-21 in AML progression
remain unknown.
B cell lymphoma/leukemia 11B (BCL11B) is a
Krüppel-like C2H2 zinc finger transcription factor located on
chromosome 14q32.2 that is required for normal T-cell development
(37). Loss of BCL11B function in
mice contributes to lymphomagenesis (38). BCL11B may have suppressive and
disruptive effects on the proliferation and differentiation of
myeloid cells (39).
miR-21 (miRBase Accession number: MI0000077), a
stem-loop precursor sequence, is processed into two mature miRNA
sequences, miR-21-5p (miRBase Accession number: MIMAT0000076) and
miR-21-3p (miRBase Accession number: MIMAT0004494), are derived
from 5′ and 3′ ends of miR-21, respectively (40). In the present study, it was
demonstrated that miR-21 was highly expressed in patients with AML
and in AML cell lines. Overexpression of miR-21 promoted the
proliferation of Thp-1 cells, which derive from acute mononuclear
leukemia peripheral blood, while downregulation of miR-21-5p
(sequence: 5′-UAGCUUAUCAGACUGAUGUUGA-3′) inhibited cell
proliferation. Specifically, it was observed that overexpression of
miR-21 could promote the entry of Thp-1 cells into the S and G2/M
phases of the cell cycle, while inhibition of miR-21-5p arrested
the cells in the S and G2/M phases. In addition, BCL11B was
identified as the functional target of miR-21-5p in Thp-1 cells.
This study provides a novel insightful understanding of miR-21 in
AML.
Materials and methods
The Cancer Genome Atlas (TCGA)
dataset
miRNA and mRNA expression data, and clinical data
for patients with AML, were obtained from TCGA data portal
(http://cancergenome.nih.gov). Both the
miRNA and mRNA expression data and clinical data, including the FAB
subtype information of TCGA AML patients deposited at the Data
Coordinating Center, are publicly available through open access.
The present study meets the publication guidelines provided by TCGA
(41). In total, data of 102 tumor
samples were obtained, which were classified into six types (M0,
M1, M2, M3, M4 and M5) according to their clinical data, excluding
subtypes with low number of cases such as M6 (n=1) and M7 (n=2).
The miRNA and mRNA expression data from 99 cases were available and
included in the datasets from the platforms. All datasets were
processed and calculated for kilo reads per million (KRPM).
Cell culture
Human bone marrow stromal HS-5 cells and human
leukemia cell lines, including HL-60, NB4 and Thp-1, were purchased
from American Type Culture Collection. Mycoplasma testing was
performed on all the cell lines used. The cells were maintained in
RPMI-1640 culture medium (Gibco; Thermo Fisher Scientific, Inc.)
containing 10% fetal bovine serum (Gibco; Thermo Fisher Scientific,
Inc.), and 2 mM L-glutamine and 1% penicillin-streptomycin solution
(10,000 U/ml penicillin and 10 mg/ml streptomycin, HyClone; GE
Healthcare Life Sciences) at 37°C in 5% CO2.
Lentivirus infection
Lentiviral vectors expressing hsa-miR-21 (LV-miR-21)
and hsa-miR-21-5p-inhibitor (LV-miR-21-5p inhibitor), as well as a
control vector (LV-control), were constructed by Shanghai GeneChem
Co., Ltd. For lentivirus-mediated miR-21, miR-21-5p-inhibitor or
control vector transfection in vitro, 1.0×105
cells/ml Thp-1 cells were infected with a lentivirus at a
multiplicity of infection of 50 in the presence of polybrene (5
µg/ml; Sigma-Aldrich; Merck KGaA). After 8–12 h of infection, the
culture media containing lentiviruses was removed after
centrifugation at 300 × g for 5 min at room temperature, and the
cells were incubated with RPMI-1640 complete culture media for
additional 48–72 h. Next, the cells were screened with 2 µg/ml
puromycin to obtain stable cell lines for subsequent
experiments.
Cell proliferation assay
MTT assay was used to evaluate cell proliferation.
Normal Thp-1 cells, as well as cells transfected with LV-miR-21,
LV-miR-21-5p inhibitor and LV-control, were plated in a 96-well
plate at a density of 1.5×104 cells/well. After 24 or 48
h, 20 µl MTT solution (5 mg/ml, Sigma-Aldrich; Merck KGaA) was
added to each well, and the cells were cultured for 4 h.
Subsequently, 200 µl DMSO was added to each well, and the
absorbance was measured at 492 nm with a microplate reader
(Multiskan Ascent; Thermo Fisher Scientific, Inc.).
Cell cycle assay
Flow cytometry was used to conduct cell cycle
analysis. Normal Thp-1 cells, as well as cells transfected with
LV-miR-21, LV-miR-21-5p inhibitor and LV-control, that were in the
exponential phase of cell proliferation were centrifuged at 1,000 ×
g for 5 min at room temperature, and the cell pellets were
harvested. Next, the cells were washed once with PBS for 2 min and
fixed in 70% ethanol overnight at 4°C. Subsequently, the cells were
washed with PBS and treated with 0.1 mg/ml RNase A (Sigma-Aldrich;
Merck KGaA) at 37°C for 30 min. Finally, the cells were stained
with propidium iodide (Sigma-Aldrich; Merck KGaA) at 4°C for 30 min
in the dark. Fluorescence was then detected with a FACS 420 system
(BD Biosciences) at 488 nm. ModFit 3.0 software (BD Biosciences)
was used to analyze the DNA content.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from Thp-1 cells with
TRIzol® reagent (Thermo Fisher Scientific, Inc.).
RT-qPCR for miRNA analysis was performed with cDNA, which was
generated from 1 µg total RNA using the All-in-One™ miRNA
First-Strand cDNA Synthesis Kit (GeneCopoeia, Inc.) and a miRNA
RT-qPCR detection kit (GeneCopoeia, Inc.). RT-qPCR analysis was
performed using the U6 gene as an internal control for
normalization and the LightCycler® 96 Real-Time PCR
System (Roche Diagnostics) with the following conditions: A heating
step at 95°C for 10 min, followed by 40 cycles of 95°C for 10 sec,
60°C for 20 sec and 72°C for 20 sec. PCR specificity was checked by
melting curve analysis. The primers against mature miRNA
hsa-miR-21-3p (cat. no. HmiRQP0315), hsa-miR-21-5p (cat. no.
HmiRQP0316), and U6 (cat. no. HmiRQP9001) were purchased from
GeneCopoeia, Inc. All reactions were conducted in triplicate, and
the 2−ΔΔCq method (42)
was used to quantify the level of miRNA expression.
Western blot analysis
Western blot analysis was carried out to detect the
protein expression of proliferating cell nuclear antigen (PCNA),
BCL11B and murine doubleminute 2 (MDM2) in normal Thp-1 cells, as
well as cells transfected with LV-miR-21, LV-miR-21-5p inhibitor
and LV-control. Cells were lysed in RIPA lysis buffer (Beyotime
Institute of Biotechnology) on ice. The cellular debris was
discarded by centrifugation at 4°C and 12,000 × g for 15 min. BCA
Protein Assay Kit (Beyotime Institute of Biotechnology) was used to
measure the protein concentration of the supernatants. Protein
samples (30 µg per lane) were separated on 12% SDS-PAGE and then
transferred to polyvinylidene difluoride membranes. The membranes
were blocked with 5% skim milk for 1.5 h at room temperature and
then incubated overnight at 4°C with primary antibodies against
PCNA (monoclonal rabbit anti-human, cat. no. ab92552; 1:1,000;
Abcam), BCL11B (monoclonal rabbit anti-human, cat. no. ab240636;
1:1,000; Abcam), MDM2 (monoclonal rabbit anti-human, cat. no.
ab259265; 1:1,000; Abcam) and GAPDH (monoclonal rabbit anti-human,
cat. no. ab181602; 1:1,000; Abcam), which served as a loading
control. Membranes were washed three times for ten minutes each
with Tris-buffered saline containing 0.1% (v/v) Tween-20 (TBS-T),
then were incubated withHRP-labeled goat anti-rabbit IgG (cat. no.
111-035-003; 1:10,000; Jackson ImmunoResearch Laboratories, Inc.)
for 1.5 h at room temperature. After washing three times for ten
minutes each with TBS-T, the bound antibodies were detected using
enhanced chemiluminescence reagents (Beyotime Institute of
Biotechnology) and visualized by a ChemiDoc-It Imaging System
(Analytik Jena AG). The image density of the immunoblots was
determined by Image Pro-Plus 6.0 software (Media Cybernetics,
Inc.).
Bioinformatic analysis of miR-21-5p
target genes
Putative miR-21-5p targets were predicted using
different algorithms, including miRanda (http://www.miranda.org) and TargetScan Release 7.2
(http://www.targetscan.org/vert_72).
The miRNA-target interactions and sequence conservation were
analyzed with TargetScan.
Dual-luciferase reporter assay
293 cells were obtained from The Cell Bank of Type
Culture Collection of The Chinese Academy of Sciences, and were
cultured in Dulbecco's modified Eagle's medium (Gibco; Thermo
Fisher Scientific, Inc.) at 37°C with 5% CO2. Cells were
plated in a 96-well plate at a density of 1.5×104
cells/well. A psiCHECK-2 reporter vector (Promega Corporation)
containing the wild-type (WT) 3′-UTR or the random mutations (MUT)
of BCL11B were co-transfected with miR-21-5p mimic, miR-21-5p
inhibitor or a negative control (NC) (Shanghai GenePharma Co.,
Ltd.) at a concentration of 50 nM into 293 cells using
Lipofectamine® 2000 reagent (Thermo Fisher Scientific,
Inc.). After 48 h, the firefly and Renilla luciferase
activities were detected by a Lumat LB9507 luminometer
(Titertek-Berthold) using a Dual-Luciferase Reporter Assay System
(Promega Corporation) according to the manufacturer's protocol. The
relative luciferase activity was calculated as the ratio of
Renilla signal/firefly signal.
Statistical analysis
All experiments were performed at least in
triplicate. The experimental data are expressed as the mean ±
standard deviation. SPSS software version 18.0 (SPSS, Inc.) was
used to perform statistical analyses. Differences among groups were
evaluated by one-way ANOVA followed by Tukey's post-hoc test.
Statistical analyses between two groups were evaluated with
Student's t-test. Two-sided P<0.05 was considered to indicate a
statistically significant difference.
Results
miR-21 is expressed at high levels in
patients with AML and in AML cell lines
miR-21 expression was analyzed in TCGA samples from
103 patients with AML. First, based on the patient information, the
data were classified as M0-M5 according to the FAB criteria, and
the expression levels of miR-21 in these samples were compared. As
shown in Fig. 1A, miR-21 expression
was significantly higher in M3, M4 and M5 than in M0, M1 and M2. No
differences were observed among the subgroups M3, M4 or M5.
Additionally, the levels of miR-21-3p and miR-21-5p in various cell
lines were examined. Human bone marrow stromal HS-5 cells were
selected as normal control cells. The RT-qPCR results showed that
the levels of miR-21-5p and miR-21-3p were significantly
upregulated in the Thp-1, HL-60 and NB4 cell lines compared with
their expression levels in HS-5 cells (Fig. 1B and C), suggesting that miR-21 was
expressed at high levels in AML cell lines. Of note, Thp-1 cells
appeared to exhibit the highest miR-21 expression; therefore, the
AML cell line Thp-1 was selected for subsequent in vitro
experiments.
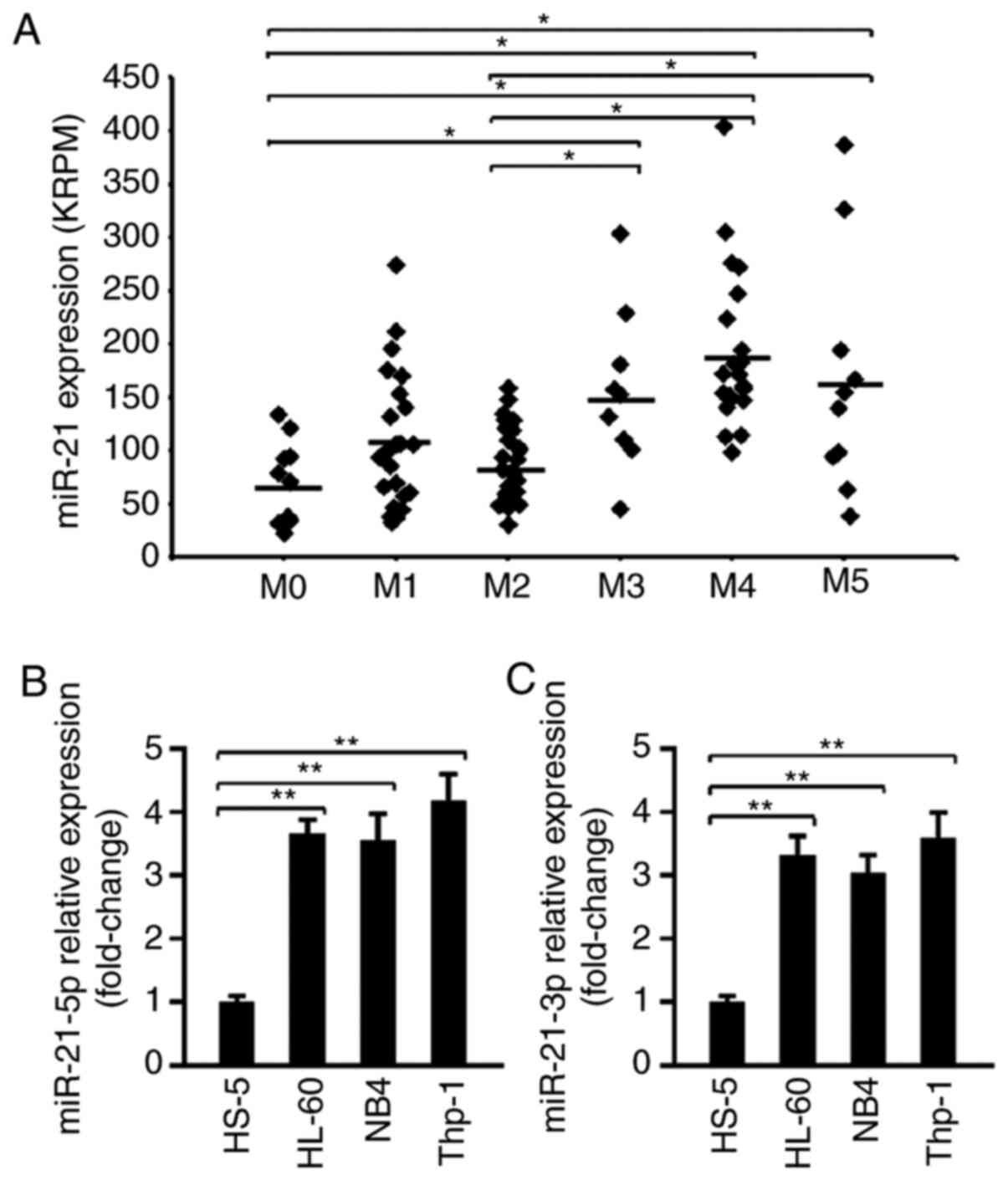 | Figure 1.miR-21 is highly expressed in
patients with AML and in AML cell lines. (A) The expression of
miR-21 in patients with AML from The Cancer Genome Atlas data
portal was analyzed. A total of 102 tumor samples were classified
into six types (M0, M1, M2, M3, M4 and M5) according to their
clinical data. All datasets were processed and calculated for KRPM.
*P<0.05. (B) The relative expression of miR-21-5p in the human
bone marrow stromal cell line HS-5 and in three human AML cell
lines (HL-60, NB4 and Thp-1) was determined by RT-qPCR (n=5).
**P<0.01. (C) The relative expression of miR-21-3p in HS-5,
HL-60, NB4 and Thp-1 cells was determined by RT-qPCR (n=5).
**P<0.01. AML, acute myeloid leukemia; miR, microRNA; RT-qPCR,
reverse transcription-quantitative PCR; KRPM, kilo reads per
million. |
miR-21-5p significantly promotes Thp-1
cell proliferation
To investigate the biological function of miR-21,
human Thp-1 cell lines that stably overexpressed miR-21
(LV-miR-21), miR-21-5p inhibitor (LV-miR-21-5p inhibitor) or NC
(LV-control) were constructed. As shown in Fig. S1, the expression of miR-21-3p and
miR-21-5p was markedly upregulated in cells infected with
LV-miR-21, while miR-21-5p, not miR-21-3p, was significantly
downregulated in cells infected with LV-miR-21-5p inhibitor,
suggesting that miR-21 overexpression and miR-21-5p inhibition were
successful in Thp-1 cells.
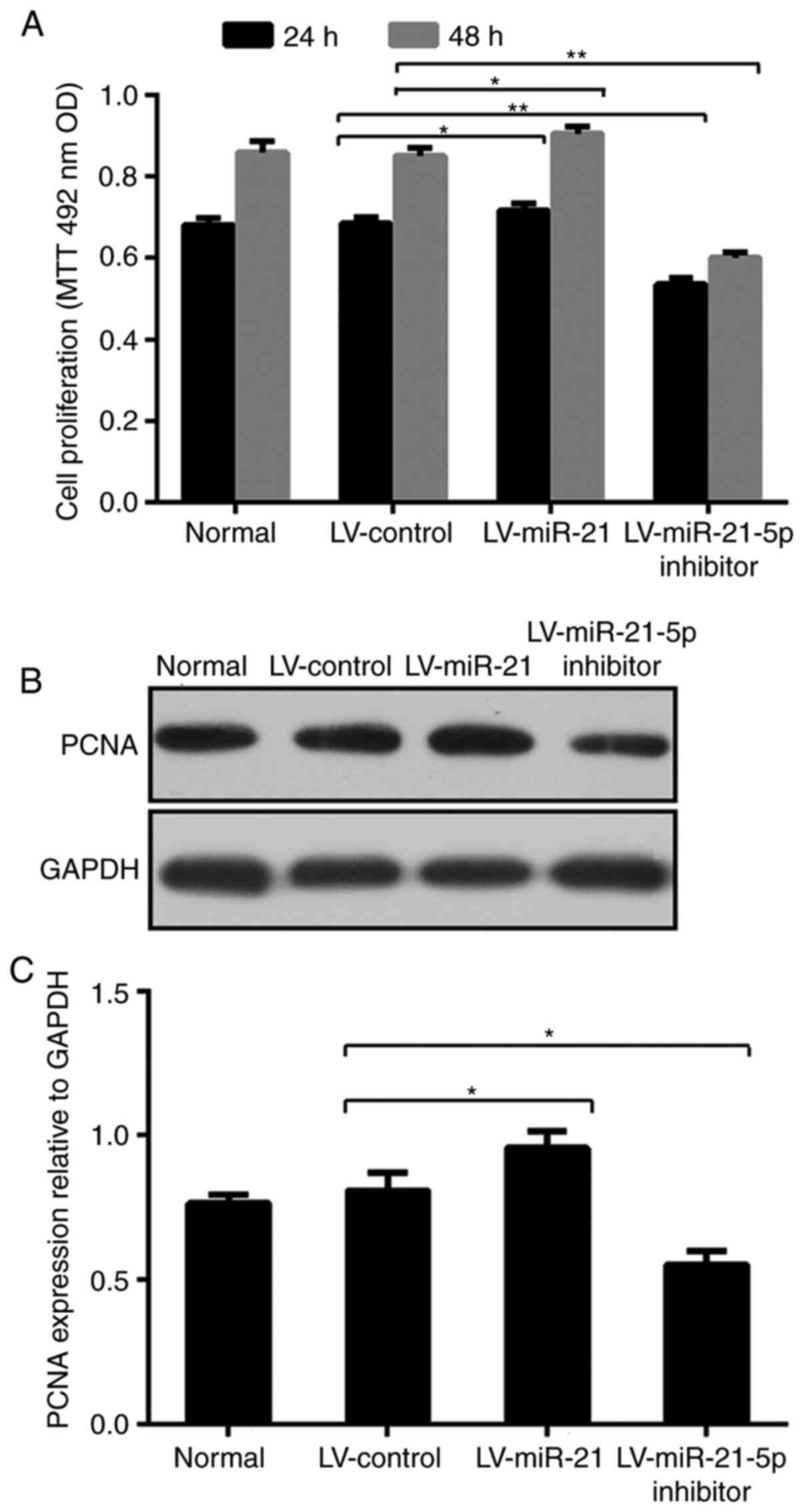
To evaluate the proliferation of normal cells and
cells transfected with LV-miR-21, LV-miR-21-5p inhibitor or
LV-control, an MTT assay was performed. As presented in Fig. 2A, the proliferation of cells
transfected with LV-miR-21-5p inhibitor was obviously reduced
compared with that of cells transfected with LV-control at 24 h
(0.684±0.014 and 0.535±0.014) and 48 h (0.856±0.025 and
0.601±0.012), respectively. Unlike that of cells transfected with
LV-miR-21-5p inhibitor, the proliferation of cells transfected with
LV-miR-21 was significantly higher compared with that of LV-control
cells at 24 and 48 h, indicating that miR-21-5p promoted Thp-1 cell
proliferation.
PCNA, a nuclear antigen in proliferating cells,
protein levels peak during the S phase of the cell cycle (43). PCNA is almost undetectable in other
phases of the cycle. Due to its unique expression, PCNA has been
extensively used in studies associating the prognosis of tumor
progression and neoplastic proliferation (44–46).
Therefore, the expression of PCNA was investigated by western
blotting in the present study. As observed in Fig. 2B and C, and consistent with the cell
proliferation assay, the PCNA protein levels were substantially
increased in Thp-1 cells after transfection with LV-miR-21, whereas
transfection with LV-miR-21-5p inhibitor reversed such increase.
Collectively, these data suggested that miR-21-5p promoted Thp-1
cell proliferation.
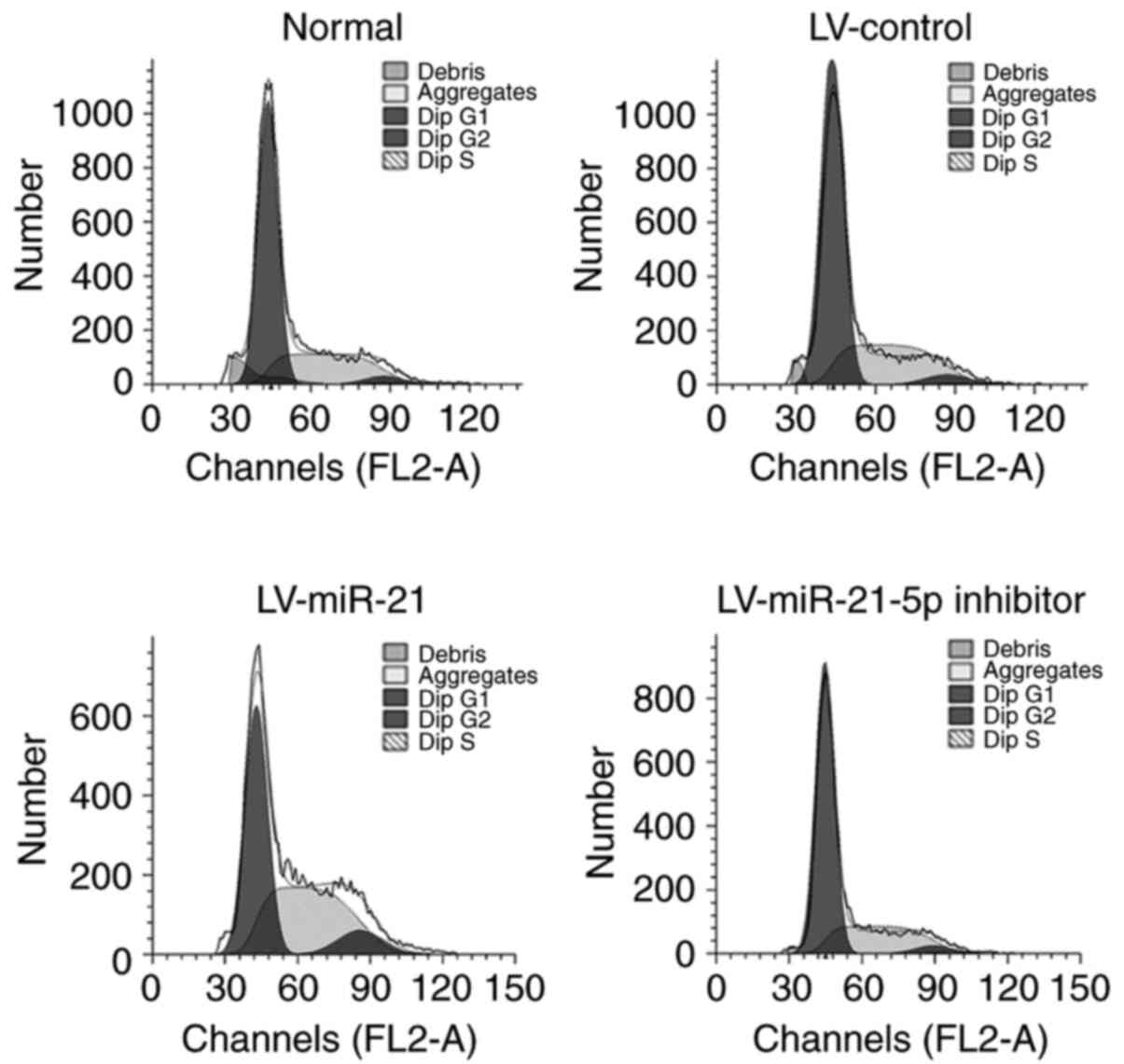
miR-21-5p promotes the transition of
Thp-1 cells into the proliferative phases of the cell cycle
To explore the possible mechanism by which miR-21
promotes the proliferation of Thp-1 cells, the distribution of
cells within the different stages of the cell cycle was determined
by flow cytometry. As shown in Table
I and Fig. 3, compared with that
of LV-control cells, the proportion of cells transfected with
LV-miR-21 in the G0/G1 phase was remarkably reduced by 17.0%, while
it was increased by ~22.5% in cells transfected with LV-miR-21-5p
inhibitor. The proportion of cells transfected with miR-21 in the S
phase was significantly increased by 15.9% compared with that of
the LV-control group, while it was reduced by ~26.8% in cells
transfected with LV-miR-21-5p inhibitor. The proportion of
LV-miR-21 cells in the G2/M phase increased by ~56.5% compared with
that of the LV-control group, whereas it decreased by ~33.2% in
LV-miR-21-5p inhibitor-transfected cells. Together, these results
indicated that overexpression of miR-21-5p promoted the transition
of Thp-1 cells into the proliferative phases of the cell cycle (as
indicated by the proportion of cells in the S and G2/M phases),
while inhibition of miR-21-5p arrested cells in the S and G2/M
phases.
 | Table I.Percentage of cells in cell cycle
phases by flow cytometry (mean ± standard deviation). |
Table I.
Percentage of cells in cell cycle
phases by flow cytometry (mean ± standard deviation).
| Group | G0/G1 | S | G2/M |
|---|
| Normal | 59.69±1.12 | 35.57±1.36 | 4.75±0.94 |
| LV-control | 55.08±0.18 | 39.38±0.28 | 5.54±0.10 |
| LV-miR-21 |
45.71±0.13a |
45.63±0.45a |
8.67±0.30b |
| LV-miR-21-5p
inhibitor |
67.48±0.18a |
28.82±0.27a |
3.70±0.08a |
BCL11B is the direct target gene of
miR-21-5p
To determine the underlying mechanisms by which
miR-21-5p contributes to AML, the bioinformatic algorithms miRanda
and TargetScan were used to predict the potential target genes of
miR-21-5p. According to the prediction analysis, BCL11B, an
important transcriptional regulator and a tumor suppressor gene
(47), has a putative
miR-21-5p-binding site (Fig. 4A).
The target sequence is highly conserved across species, including
human, chimp, rhesus, squirrel, mouse, rat, rabbit and pig
(Fig. 4B). To validate the
miRNA-target interactions, the 3′-UTR sequence of BCL11B containing
the putative binding site of miR-21-5p was cloned into the
psiCHECK-2 reporter vector. The constructed vector was then
co-transfected with miR-21-5p mimic, miR-21-5p inhibitor or NC into
293 cells. As shown in Fig. S2, the
expression of miR-21-5p was markedly upregulated in cells
transfected with miR-21-5p mimic, and significantly downregulated
in cells transfected with miR-21-5p inhibitor, suggesting that
miR-21-5p overexpression and inhibition were successful. As
presented in Fig. 4C, the
transfection of miR-21-5p mimic suppressed the luciferase activity,
while miR-21-5p inhibitor increased the activity of the WT-BCL11B
reporter compared with that of the NC. No significant change was
observed in the luciferase activity of the MUT-BCL11B reporter
between the miR-21-5p mimic and miR-21-5p inhibitor groups,
suggesting that BCL11B is a direct target gene of miR-21-5p.
 | Figure 4.BCL11B is a direct target of
miR-21-5p. (A) Schematic representation of the miR-21-5p putative
binding site in the human BCL11B 3’-UTR, and alignment of the WT
and MUT BCL11B 3’-UTR binding site of miR-21-5p. The BCL11B mutated
nucleotides are underlined. (B) The conserved miR-21-5p binding
site in the BCL11B 3’-UTR is shown in grey. (C) 293 cells were
co-transfected with a luciferase reporter carrying BCL11B-3’-UTR-WT
or BCL11B-3’-UTR-MUT, together with miR-21-5p mimic, miR-21-5p
inhibitor or a negative control. Luciferase activities were
measured at 48 h after transfection (n=5). *P<0.05. (D) BCL11B
protein expression was analyzed in Thp-1 normal cells, and in
LV-miR-21, LV-miR-21-5p inhibitor and LV-control cells by western
blotting. (E) The blots were quantified with Image Pro-Plus 6.0
software, and the results are represented as the BCL11B/GAPDH ratio
(n=3). *P<0.05. (F) The expression of BCL11B in patients with
acute myeloid leukemia from The Cancer Genome Atlas data portal was
analyzed. All datasets were processed and calculated for KRPM.
*P<0.05. miR, microRNA; LV, lentivirus; WT, wild-type; MUT,
mutant; UTR, untranslated region; NC, negative control; KRPM, kilo
reads per million; BCL11B, B-cell lymphoma/leukemia 11B. |
Furthermore, western blotting was performed to
determine whether miR-21-5p influenced the expression level of
BCL11B in Thp-1 cells. As revealed in Fig. 4D and E, the expression of BCL11B
protein was remarkably decreased after overexpression of miR-21-5p;
however, it was significantly increased in LV-miR-21-5p
inhibitor-transfected cells. The expression of BCL11B was further
analyzed in patients with AML from TCGA dataset. The results showed
that the expression of BCL11B in patients with FAB M5 was
significantly decreased compared with that of patients with M0 and
M4. No differences were observed among other subgroups (Fig. 4F). Together, these results suggested
that miR-21-5p suppressed BCL11B expression by directly targeting
its 3′-UTR in Thp-1 cells.
Discussion
The current study aimed to investigate the role of
miR-21 in AML cell proliferation and to explore its possible
mechanisms. It was found that miR-21 was expressed at high levels
in patients with AML and in AML cell lines. Further experiments
indicated that overexpression of miR-21 in Thp-1 cells promoted
proliferation, while downregulation of miR-21-5p inhibited
proliferation. Specifically, it was observed that overexpression of
miR-21 could promote Thp-1 cell transition into the S and G2/M
phases of the cell cycle, as shown by flow cytometry, while
inhibition of miR-21-5p arrested cells in the S and G2/M phases.
Finally, BCL11B was identified as a functional target of miR-21-5p
by luciferase assays.
miR-21 has been reported to be an oncogene, and to
be upregulated in various human cancer types such as glioblastoma,
and prostate, breast, pancreas, esophagus and liver cancer
(29,48–50).
Additionally, miR-21 is also highly expressed in various types of
blood cancer (51–54). Furthermore, several targets of miR-21
have been identified, including PDCD4 (27), myristoylated alanine-rich C-kinase
substrate (49) and leucine-rich
repeat (in FLII) interacting protein 1 (48). Previous studies have indicated that
miR-21 promotes the proliferation and growth of hepatocellular
carcinoma cells by targeting PTEN, which activates the AKT
signaling pathway (55). However,
there are limited reports on the function and targets of miR-21 in
AML. The present study showed that BCL11B is the direct target gene
of miR-21-5p in Thp-1 cells.
The BCL11 family has two members, BCL11A and BCL11B.
BCL11A is a zinc finger protein that is critically involved in the
normal differentiation of lymphocytes (56), while BCL11B is a Krüppel-like C2H2
zinc finger transcription factor that is required for normal T-cell
development (37). Abbas et
al (39) reported that BCL11B
may have suppressive and disruptive effects on the proliferation
and differentiation of myeloid cells. Loss of BCL11B was
demonstrated to cause a natural killer cell-like phenotype with
differentiation arrest, and was linked to a high proliferative
potential. Mechanistically, BCL11B binds to the GC-rich consensus
sequence of target genes, and/or interacts with the nucleosome
remodeling and histone deacetylase complex, thus repressing the
transcription of target genes (57).
Previous studies have found that BCL11B binds to the P2 promotor of
human double minute 2 promoter and inhibits HDM2 expression in a
p53-dependent manner (47). HDM2,
also known as MDM2, is an oncogene that encodes a nucleus-localized
E3 ubiquitin ligase that can promote tumor formation by targeting
tumor suppressor proteins such as p53. Activated p53 regulates
multiple p53 target genes, which results in various cellular
responses, including apoptosis, cell cycle arrest and DNA repair
(58). In addition, MDM2 also has
p53-independent oncogenic functions, including control of
proliferation, apoptosis and tumor invasion. For example, MDM2 can
interact with the tumor suppressor retinoblastoma protein, which is
a negative regulator of the cell cycle, thus inhibiting its
function in the regulation of the cell cycle and proliferation
(59). In our study, the MDM2
protein levels in LV-miR-21, LV-miR-21-5p inhibitor and LV-control
cells were analyzed. As shown in Fig.
S3, the expression of MDM2 was remarkably increased after
overexpression of miR-21; however, it was significantly decreased
in LV-miR-21-5p inhibitor-transfected cells, suggesting that miR-21
targets BCL11B and then regulates the expression of MDM2. Another
study showed that the expression of the CDK inhibitor p27 was
decreased in BCL11B-knockdown Jurkat cells, which are a type of
T-cell line used for studying the regulatory role of BCL11B in the
cell cycle. A decrease in p27 may promote cell cycle progression
during the S phase (60). In
addition, in BCL11B-knockdown cells, the activation of the cell
cycle checkpoint kinase Chk1 was deregulated, and activated Chk1
led to the arrest of the cell cycle at the S phase by Cdc25A
phosphorylation (61). The present
study confirmed that miR-21-5p regulated the expression of BCL11B.
In AML cells, downregulated expression of BCL11B may influence the
process of cell proliferation by changing the expression of MDM2 or
a CDK inhibitor, or by regulating the activation of Chk1. The exact
mechanism is not entirely clear yet.
In conclusion, our study demonstrated that miR-21
was expressed at high levels in patients with AML and in AML cell
lines. Functional and mechanistic studies suggested that miR-21-5p
promoted Thp-1 cell proliferation (as indicated by the proportion
of cells in the S and G2/M phases of the cell cycle) by targeting
BCL11B. The present study provides a new insight into the
mechanisms of AML, and suggests that miR-21 might be a potential
AML therapeutic target.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This work was supported by the National Natural
Science Foundation of China (grant nos. 81100193 and 31871441) and
the Shandong Provincial Natural Science Foundation (grant no.
ZR2017MH087).
Availability of data and materials
The datasets generated and analyzed during the study
are available from the corresponding author on reasonable
request.
Authors' contributions
LZ and LY conducted the cell culture and cell
proliferation experiments, as well as the cell cycle analyses, and
were major contributors in writing the manuscript. YL and SW
conducted the RT-qPCR and western blot experiments, and performed
the statistical analyses. ZH analyzed TCGA data and revised the
manuscript. JZ contributed to the study design. All authors read
and approved the final version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Vandenberghe H; Second MIC Cooperative
Study Group, : Morphologic, immunologic and cytogenetic (MIC)
working classification of the acute myeloid leukaemias. Br J
Haematol. 68:487–494. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Head DR: Revised classification of acute
myeloid leukemia. Leukemia. 10:1826–1831. 1996.PubMed/NCBI
|
|
3
|
Ravindranath Y, Chang M, Steuber CP,
Becton D, Dahl G, Civin C, Camitta B, Carroll A, Raimondi SC and
Weinstein HJ; Pediatric Oncology Group, : Pediatric Oncology Group
(POG) studies of acute myeloid leukemia (AML): A review of four
consecutive childhood AML trials conducted between 1981 and 2000.
Leukemia. 19:2101–2116. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Smith FO, Alonzo TA, Gerbing RB, Woods WG,
Arceci RJ and Grp Cs C; Children's Cancer Group, : Long-term
results of children with acute myeloid leukemia: a report of three
consecutive Phase III trials by the Children's Cancer Group: CCG
251, CCG 213 and CCG 2891. Leukemia. 19:2054–2062. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Estey EH: How I treat older patients with
AML. Blood. 96:1670–1673. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cummins JM and Velculescu VE: Implications
of micro-RNA profiling for cancer diagnosis. Oncogene.
25:6220–6227. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Filipowicz W, Bhattacharyya SN and
Sonenberg N: Mechanisms of post-transcriptional regulation by
microRNAs: Are the answers in sight? Nat Rev Genet. 9:102–114.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kumar MS, Lu J, Mercer KL, Golub TR and
Jacks T: Impaired microRNA processing enhances cellular
transformation and tumorigenesis. Nat Genet. 39:673–677. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Croce CM and Calin GA: miRNAs, cancer, and
stem cell division. Cell. 122:6–7. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gregory RI and Shiekhattar R: MicroRNA
biogenesis and cancer. Cancer Res. 65:3509–3512. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Silakit R, Loilome W, Yongvanit P, Chusorn
P, Techasen A, Boonmars T, Khuntikeo N, Chamadol N, Pairojkul C and
Namwat N: Circulating miR-192 in liver fluke-associated
cholangiocarcinoma patients: A prospective prognostic indicator. J
Hepatobiliary Pancreat Sci. 21:864–872. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang LG and Gu J: Serum microRNA-29a is a
promising novel marker for early detection of colorectal liver
metastasis. Cancer Epidemiol. 36:e61–e67. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ng EKO, Chong WWS, Jin H, Lam EK, Shin VY,
Yu J, Poon TC, Ng SS and Sung JJ: Differential expression of
microRNAs in plasma of patients with colorectal cancer: A potential
marker for colorectal cancer screening. Gut. 58:1375–1381. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen P, Price C, Li Z, Li Y, Cao D, Wiley
A, He C, Gurbuxani S, Kunjamma RB, Huang H, et al: miR-9 is an
essential oncogenic microRNA specifically overexpressed in mixed
lineage leukemia-rearranged leukemia. Proc Natl Acad Sci USA.
110:11511–11516. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li C, Yan H, Yin J, Ma J, Liao A, Yang S,
Wang L, Huang Y, Lin C, Dong Z, et al: MicroRNA-21 promotes
proliferation in acute myeloid leukemia by targeting Krüppel-like
factor 5. Oncol Lett. 18:3367–3372. 2019.PubMed/NCBI
|
|
19
|
Liu L, Ren W and Chen K: MiR-34a promotes
apoptosis and inhibits autophagy by targeting HMGB1 in acute
myeloid leukemia cells. Cell Physiol Biochem. 41:1981–1992. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
de Leeuw DC, Denkers F, Olthof MC, Rutten
AP, Pouwels W, Schuurhuis GJ, Ossenkoppele GJ and Smit L:
Attenuation of microRNA-126 expression that drives
CD34+38− stem/progenitor cells in acute
myeloid leukemia leads to tumor eradication. Cancer Res.
74:2094–2105. 2014. View Article : Google Scholar
|
|
21
|
Hartmann JU, Brauer-Hartmann D, Kardosova
M, Wurm AA, Wilke F, Schödel C, Gerloff D, Katzerke C, Krakowsky R,
Namasu CY, et al: MicroRNA-143 targets ERK5 in granulopoiesis and
predicts outcome of patients with acute myeloid leukemia. Cell
Death Dis. 9:8142018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wallace JA, Kagele DA, Eiring AM, Kim CN,
Hu R, Runtsch MC, Alexander M, Huffaker TB, Lee SH, Patel AB, et
al: miR-155 promotes FLT3-ITD-induced myeloproliferative disease
through inhibition of the interferon response. Blood.
129:3074–3086. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu ZL, Wang H, Liu J and Wang ZX:
MicroRNA-21 (miR-21) expression promotes growth, metastasis, and
chemo- or radioresistance in non-small cell lung cancer cells by
targeting PTEN. Mol Cell Biochem. 372:35–45. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhu Y, Han Y, Tian T, Su P, Jin G, Chen J
and Cao Y: MiR-21-5p, miR-34a, and human telomerase RNA component
as surrogate markers for cervical cancer progression. Pathol Res
Pract. 214:374–379. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Okugawa Y, Yao L, Toiyama Y, Yamamoto A,
Shigemori T, Yin C, Omura Y, Ide S, Kitajima T, Shimura T, et al:
Prognostic impact of sarcopenia and its correlation with
circulating miR-21 in colorectal cancer patients. Oncol Rep.
39:1555–1564. 2018.PubMed/NCBI
|
|
26
|
Chen S, Chen X, Shan T, Ma J, Lin W, Li W
and Kang Y: MiR-21-mediated Metabolic Alteration of
Cancer-associated Fibroblasts and Its Effect on Pancreatic Cancer
Cell Behavior. Int J Biol Sci. 14:100–110. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Luo F, Ji J, Liu Y, Xu Y, Zheng G, Jing J,
Wang B, Xu W, Shi L, Lu X, et al: MicroRNA-21, up-regulated by
arsenite, directs the epithelial-mesenchymal transition and
enhances the invasive potential of transformed human bronchial
epithelial cells by targeting PDCD4. Toxicol Lett. 232:301–309.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang Q, Xu E, Dai J, Wu J, Zhang S, Peng B
and Jiang Y: miR-21 regulates
N-methyl-N-nitro-N′-nitrosoguanidine-induced gastric tumorigenesis
by targeting FASLG and BTG2. Toxicol Lett. 228:147–156. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang G, Wang JJ, Tang HM and To SS:
Targeting strategies on miRNA-21 and PDCD4 for glioblastoma. Arch
Biochem Biophys. 580:64–74. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fu X, Han Y, Wu Y, Zhu X, Lu X, Mao F,
Wang X, He X and Zhao Y and Zhao Y: Prognostic role of microRNA-21
in various carcinomas: A systematic review and meta-analysis. Eur J
Clin Invest. 41:1245–1253. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Asangani IA, Rasheed SAK, Nikolova DA,
Leupold JH, Colburn NH, Post S and Allgayer H: MicroRNA-21 (miR-21)
post-transcriptionally downregulates tumor suppressor Pdcd4 and
stimulates invasion, intravasation and metastasis in colorectal
cancer. Oncogene. 27:2128–2136. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Espadinha AS, Prouzet-Mauléon V, Claverol
S, Lagarde V, Bonneu M, Mahon FX and Cardinaud B: A tyrosine
kinase-STAT5-miR21-PDCD4 regulatory axis in chronic and acute
myeloid leukemia cells. Oncotarget. 8:76174–76188. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ruiz-Lafuente N, Alcaraz-Garcia MJ,
Sebastian-Ruiz S, García-Serna AM, Gómez-Espuch J, Moraleda JM,
Minguela A, García-Alonso AM and Parrado A: IL-4 up-regulates
MiR-21 and the MiRNAs hosted in the CLCN5 gene in chronic
lymphocytic leukemia. Plos One. 10:e01249362015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Taverna S, Giallombardo M, Pucci M, Flugy
A, Manno M, Raccosta S, Rolfo C, De Leo G and Alessandro R:
Curcumin inhibits in vitro and in vivo chronic myelogenous leukemia
cells growth: A possible role for exosomal disposal of miR-21.
Oncotarget. 6:21918–21933. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Labib HA, Elantouny NG, Ibrahim NF and
Alnagar AA: Upregulation of microRNA-21 is a poor prognostic marker
in patients with childhood B cell acute lymphoblastic leukemia.
Hematology. 22:392–397. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Riccioni R, Lulli V, Castelli G, Biffoni
M, Tiberio R, Pelosi E, Lo-Coco F and Testa U: miR-21 is
overexpressed in NPM1-mutant acute myeloid leukemias. Leuk Res.
39:221–228. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liu P, Li P and Burke S: Critical roles of
Bcl11b in T-cell development and maintenance of T-cell identity.
Immunol Rev. 238:138–149. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wakabayashi Y, Inoue J, Takahashi Y,
Matsuki A, Kosugi-Okano H, Shinbo T, Mishima Y, Niwa O and Kominami
R: Homozygous deletions and point mutations of the Rit1/Bcl11b gene
in gamma-ray induced mouse thymic lymphomas. Biochem Biophys Res
Commun. 301:598–603. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Abbas S, Sanders MA, Zeilemaker A,
Geertsma-Kleinekoort WM, Koenders JE, Kavelaars FG, Abbas ZG,
Mahamoud S, Chu IW, Hoogenboezem R, et al: Integrated genome-wide
genotyping and gene expression profiling reveals BCL11B as a
putative oncogene in acute myeloid leukemia with 14q32 aberrations.
Haematologica. 99:848–857. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Landgraf P, Rusu M, Sheridan R, Sewer A,
Iovino N, Aravin A, Pfeffer S, Rice A, Kamphorst AO, Landthaler M,
et al: A mammalian microRNA expression atlas based on small RNA
library sequencing. Cell. 129:1401–1414. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
National Cancer Institute, . The Cancer
Genome Atlas Program. Accessed from:. simplehttp://cancergenome.nih.gov/publications/publicationguidelines
|
|
42
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−ΔΔC(T)) method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Morris GF and Mathews MB: Regulation of
proliferating cell nuclear antigen during the cell cycle. J Biol
Chem. 264:13856–13864. 1989.PubMed/NCBI
|
|
44
|
Zhong W, Peng J, He H, Wu D, Han Z, Bi X
and Dai Q: Ki-67 and PCNA expression in prostate cancer and benign
prostatic hyperplasia. Clin Invest Med. 31:E8–E15. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Juríková M, Danihel Ľ, Polák Š and Varga
I: Ki67, PCNA, and MCM proteins: Markers of proliferation in the
diagnosis of breast cancer. Acta Histochem. 118:544–552. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Kimos MC, Wang S, Borkowski A, Yang GY,
Yang CS, Perry K, Olaru A, Deacu E, Sterian A, Cottrell J, et al:
Esophagin and proliferating cell nuclear antigen (PCNA) are
biomarkers of human esophageal neoplastic progression. Int J
Cancer. 111:415–417. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Obata M, Kominami R and Mishima Y: BCL11B
tumor suppressor inhibits HDM2 expression in a p53-dependent
manner. Cell Signal. 24:1047–1052. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Li Y, Li W, Yang Y, Lu Y, He C, Hu G, Liu
H, Chen J, He J and Yu H: MicroRNA-21 targets LRRFIP1 and
contributes to VM-26 resistance in glioblastoma multiforme. Brain
Res. 1286:13–18. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Li T, Li D, Sha J, Sun P and Huang Y:
MicroRNA-21 directly targets MARCKS and promotes apoptosis
resistance and invasion in prostate cancer cells. Biochem Biophys
Res Commun. 383:280–285. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Kimura S, Naganuma S, Susuki D, Hirono Y,
Yamaguchi A, Fujieda S, Sano K and Itoh H: Expression of microRNAs
in squamous cell carcinoma of human head and neck and the
esophagus: MiR-205 and miR-21 are specific markers for HNSCC and
ESCC. Oncol Rep. 23:1625–1633. 2010.PubMed/NCBI
|
|
51
|
Volinia S, Galasso M, Costinean S,
Tagliavini L, Gamberoni G, Drusco A, Marchesini J, Mascellani N,
Sana ME, Abu Jarour R, et al: Reprogramming of miRNA networks in
cancer and leukemia. Genome Res. 20:589–599. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Pichiorri F, Suh SS, Ladetto M, Kuehl M,
Palumbo T, Drandi D, Taccioli C, Zanesi N, Alder H, Hagan JP, et
al: MicroRNAs regulate critical genes associated with multiple
myeloma pathogenesis. Proc Natl Acad Sci USA. 105:12885–12890.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Fulci V, Chiaretti S, Goldoni M, Azzalin
G, Carucci N, Tavolaro S, Castellano L, Magrelli A, Citarella F,
Messina M, et al: Quantitative technologies establish a novel
microRNA profile of chronic lymphocytic leukemia. Blood.
109:4944–4951. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Asangani IA, Rasheed SAK, Nikolova DA,
Leupold JH, Colburn NH, Post S and Allgayer H: MicroRNA-21 (miR-21)
post-transcriptionally downregulates tumor suppressor Pdcd4 and
stimulates invasion, intravasation and metastasis in colorectal
cancer. Oncogene. 27:2128–2136. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
He C, Dong X, Zhai B, Jiang X, Dong D, Li
B, Jiang H, Xu S and Sun X: MiR-21 mediates sorafenib resistance of
hepatocellular carcinoma cells by inhibiting autophagy via the
PTEN/Akt pathway. Oncotarget. 6:28867–28881. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Lee BS, Dekker JD, Lee BK, Iyer VR,
Sleckman BP, Shaffer AL III, Ippolito GC and Tucker PW: The BCL11A
transcription factor directly activates RAG gene expression and
V(D)J recombination. Mol Cell Biol. 33:1768–1781. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Cismasiu VB, Adamo K, Gecewicz J, Duque J,
Lin Q and Avram D: BCL11B functionally associates with the NuRD
complex in T lymphocytes to repress targeted promoter. Oncogene.
24:6753–6764. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Wu H, Pomeroy SL, Ferreira M, Teider N,
Mariani J, Nakayama KI, Hatakeyama S, Tron VA, Saltibus LF,
Spyracopoulos L, et al: UBE4B promotes Hdm2-mediated degradation of
the tumor suppressor p53. Nat Med. 17:347–355. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Kubbutat MH, Jones SN and Vousden KH:
Regulation of p53 stability by Mdm2. Nature. 387:299–303. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Go R, Takizawa K, Hirose S, Katsuragi Y,
Aoyagi Y, Mishima Y and Kominami R: Impairment in differentiation
and cell cycle of thymocytes by loss of a Bcl11b tumor suppressor
allele that contributes to leukemogenesis. Leuk Res. 36:1035–1040.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Kamimura K, Mishima Y, Obata M, Endo T,
Aoyagi Y and Kominami R: Lack of Bcl11b tumor suppressor results in
vulnerability to DNA replication stress and damages. Oncogene.
26:5840–5850. 2007. View Article : Google Scholar : PubMed/NCBI
|