Introduction
Colorectal cancer (CRC) is the fourth most common
cancer worldwide (6.1% of total cases in 2018) (1). Despite advancements in its diagnosis
and treatment strategies, CRC remains the leading cause of
cancer-associated mortality worldwide (9.2% of the total
cancer-associated deaths in 2018) (1). Thus, better understanding the molecular
mechanism of CRC progression and identifying novel effective
therapeutic strategies remains essential.
Gut microbiome is closely associated with the
development of CRC (2). Recently,
the Gram-negative anaerobic bacteria, Fusobacterium
nucleatum (Fn), has gained great interest. Previous
studies have demonstrated that Fn is enriched in CRC tissues
compared with adjacent normal tissues, and associated with poor
prognosis (3,4). In addition, Fn promotes the
development of CRC by activating autophagy of cancer cells,
protecting tumors from immune attack and creating an inflammatory
microenvironment (5,6). Although it has been reported that using
berberine and targeting Fn Fap2 may decrease Fn
potentiation of CRC (7,8), methods for suppressing the carcinogenic
properties of Fn remain largely unknown.
L-Fucose (Fucose), a natural monosaccharide present
in foods or bodies, plays an important role in sustaining the gut
homeostasis (9). Our previous
studies demonstrated that fucose alleviates dextran sulfate
sodium-induced acute and chronic colitis by regulating immune
responses and affecting the intestinal microenvironment (10,11).
Notably, fucose has been reported to affect microbial metabolic
pathways, and decrease pathogen virulence (12). However, the role of fucose in CRC
remains unknown.
Thus, the present study aimed to investigate the
effects of fucose on the functional regulations of Fn and
determine its underlying molecular mechanism in CRC. Taken
together, the results of the present study suggest that fucose may
ameliorate the carcinogenic properties of Fn.
Materials and methods
Cell lines, bacterial strain culture
and groups
The HCT116 and SW480 human colon cancer cell lines
were purchased from the American Type Culture Collection and
maintained in high glucose DMEM supplemented with 10% fetal bovine
serum (FBS) and 100 U/ml streptomycin/penicillin (all purchased
from Gibco; Thermo Fisher Scientific, Inc.), at 37°C in 5%
CO2.
Fn (25586) was purchased from the American
Type Culture Collection and cultured at Wuhan Research Institute of
First Light Industry (Wuhan, China). The methods of bacterial
pellets and conditioned medium were performed, as previously
described (13). The following
groups were classified: Control group, Fn group (supernatant
of Fn was added to the cells) and 0.5% fucose+Fn
(Fnf) group. Fucose (Sigma-Aldrich; Merck KGaA) was
added when Fn was cultured and the supernatant was
subsequently added to the cells. In total, 100 nM S3I-201 (Selleck
Chemicals), an inhibitor of stat3, was incubated for 1 h prior to
bacteria treatment to access the role of stat3 pathway.
Cell Counting Kit-8 (CCK-8) assay
The CCK-8 assay was performed to assess cell
proliferation. HCT116 and SW480 cells were seeded into a 96-well
plate at a density of 1×104 with 100 µl conditioned
medium and allowed to adhere overnight. Following treatment with
Fn or Fnf for 12 h at 37°C, the culture medium was
removed and 10 µl CCK-8 solution (Beyotime Institute of
Biotechnology) along with 90 µl medium was added to each well.
Following incubation for 1 h at 37°C, cell proliferation was
analyzed at a wavelength of 450 nm, using a microplate reader
(Biotek Instruments, Inc.).
Colony formation assay
The colon cancer cells were stimulated with
Fn or Fnf for 12 h at 37°C and subsequently seeded
into 6-well plates at a density of 500 cells/well. Cells were
cultured for 14 days at 37°C in 5% CO2. Following
incubation, cells were washed with PBS for three times, fixed with
4% paraformaldehyde for 30 min and stained with 0.5% crystal violet
for 15 min both at room temperature. Cell colonies were counted
using ImageJ1 software (National Institutes of Health).
Wound healing assay
Cells from each group were respectively seeded into
6-well plates at a density of 5×105 with serum-free DMEM
medium and incubated overnight at 37°C. Cell monolayers were
scratched using a sterile pipette tip. Cells were washed with PBS
three times to remove the cellular debris, and cell migration was
subsequently analyzed after 24 h, using ImageJ1 software (National
Institutes of Health).
Transwell migration and invasion
assays
For the cell migration assay, cells from each group
were resuspended with serum-free DMEM medium and plated in the
upper chambers of 24-well Transwell plates at a density of
1×105. DMEM medium supplemented with 10% FBS was plated
in the lower chambers. Following incubation for 24 h at 37°C, the
migratory cells were fixed with 4% paraformaldehyde for 30 min and
stained with 0.5% crystal violet for 15 min both at room
temperature. Cells were observed under a light microscope
(magnification ×200; Olympus Corporation). For the cell invasion
assay, Transwell membranes were precoated with Matrigel at 37°C for
4 h.
Western blotting
Total protein was extracted from cells or tissues
using RIPA lysis buffer (Beyotime Institute of Biotechnology)
supplemented with phenylmethyl sulfonyl fluoride protease inhibitor
(Beyotime Institute of Biotechnology) and phosphatase inhibitor
(Beyotime Institute of Biotechnology). Total protein was quantified
using the BCA assay (Thermo Fisher Scientific, Inc.) and equal
amounts of protein (40 µg/lane) were separated by 10% SDS-PAGE. The
separated proteins were subsequently transferred onto PVDF
membranes (EMD Millipore) and blocked with 5% BSA at room
temperature for 1 h. The membranes were incubated with primary
antibodies (all 1:1,000 dilution) against: GAPDH (ABclonal Biotech
Co., Ltd.; cat. no. AC001), ACTB (ABcloanl Biotech Co., Ltd.; cat.
no. AC006), phospho-stat3 (Tyr705; Cell Signaling Technology, Inc.;
cat. no. 9145S), stat3 (ABclonal Biotech Co., Ltd.; cat. no.
A11216), phosphor-jak2 (Tyr1007/1008; ABclonal Biotech Co., Ltd.;
cat. no. AP0531), jak2 (ABclonal Biotech Co., Ltd.; cat. no.
A7694), β-catenin (Cell Signaling Technology, Inc.; cat. no.
8480S), E-cadherin (Cell Signaling Technology, Inc.; cat. no.
3195S), N-cadherin (GeneTex, Inc.; cat. no. GTX127345) and Vimentin
(Cell Signaling Technology, Inc.; cat. no. 5741S) overnight at 4°C.
Following the primary incubation, membranes were incubated with
HRP-labelled secondary antibodies (AntGene; cat. no. ANT020;
1:2,000) at room temperature for 1 h. Protein bands were visualized
using enhanced chemiluminescent reagents (Beyotime Institute of
Biotechnology).
Statistical analysis
Statistical analysis was performed using SPSS 25.0
software (IBM Corp.) and GraphPad Prism 7.0 software (GraphPad
Software, Inc.). All experiments were performed in triplicate and
data are presented as the mean ± standard error of the mean.
One-way analysis of variance and Tukey's post hoc test were used to
compare differences between multiple groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
Fucose ameliorates the
pro-proliferation characteristic of Fn on colon cancer cells
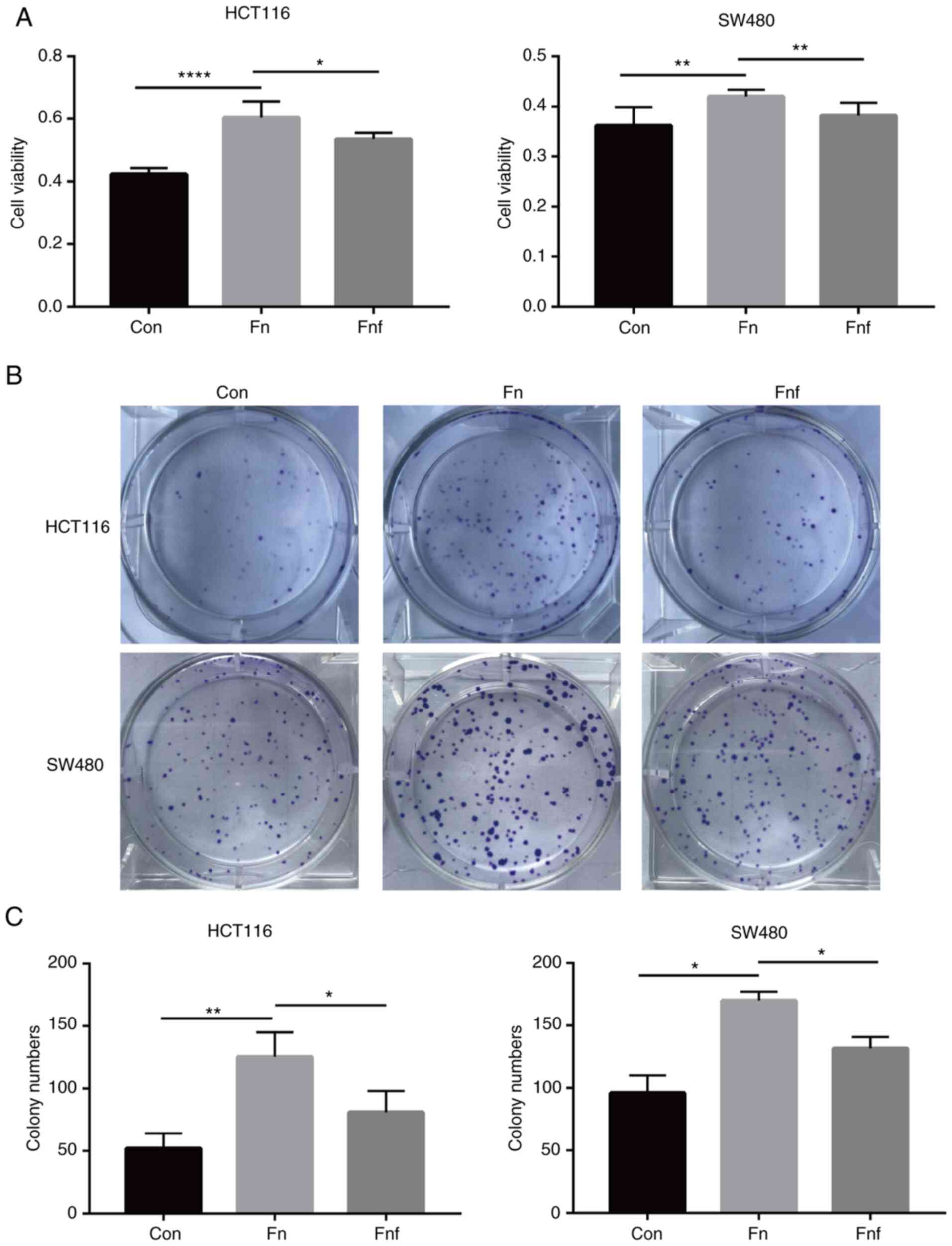
To investigate whether there were differences
between the impacts that Fn and Fnf exerted on colon
cancer cells, the CCK-8 assay was performed on HCT116 and SW480
cells following treatment for 12 h. The results demonstrated that
cells treated with Fn proliferated faster compared with the
control group (HCT116 control, 0.42±0.01; Fn, 0.60±0.02,
P<0.0001; SW480 control, 0.36±0.02; Fn, 0.42±0.01,
P=0.005; Fig. 1A). However,
following treatment with fucose, Fn exerted a weaker
pro-proliferative ability (HCT116 Fn, 0.60±0.02; Fnf,
0.54±0.01, P=0.014; SW480 Fn, 0.42±0.01; Fnf,
0.38±0.01, P=0.009; Fig. 1A). The
colony formation assay was subsequently performed to assess the
long-term effect on cell proliferation. The results demonstrated
that the number of HCT116 colonies treated with Fn increased
(control, 52.0±7.0; Fn, 125.3±11.3; P=0.005), while less
colonies were observed in the Fnf group (Fn,
125.3±11.3; Fnf, 81.0±9.9, P=0.042; Fig. 1B and C). Similar results were
observed in SW480 cells (control, 96±10; Fn, 170±5;
Fnf, 131.5±6.5; P=0.022 and P=0.043; Fig. 1B and C). Collectively, these results
suggest that L-fucose may ameliorate the pre-proliferative ability
of Fn on colon cancer cells.
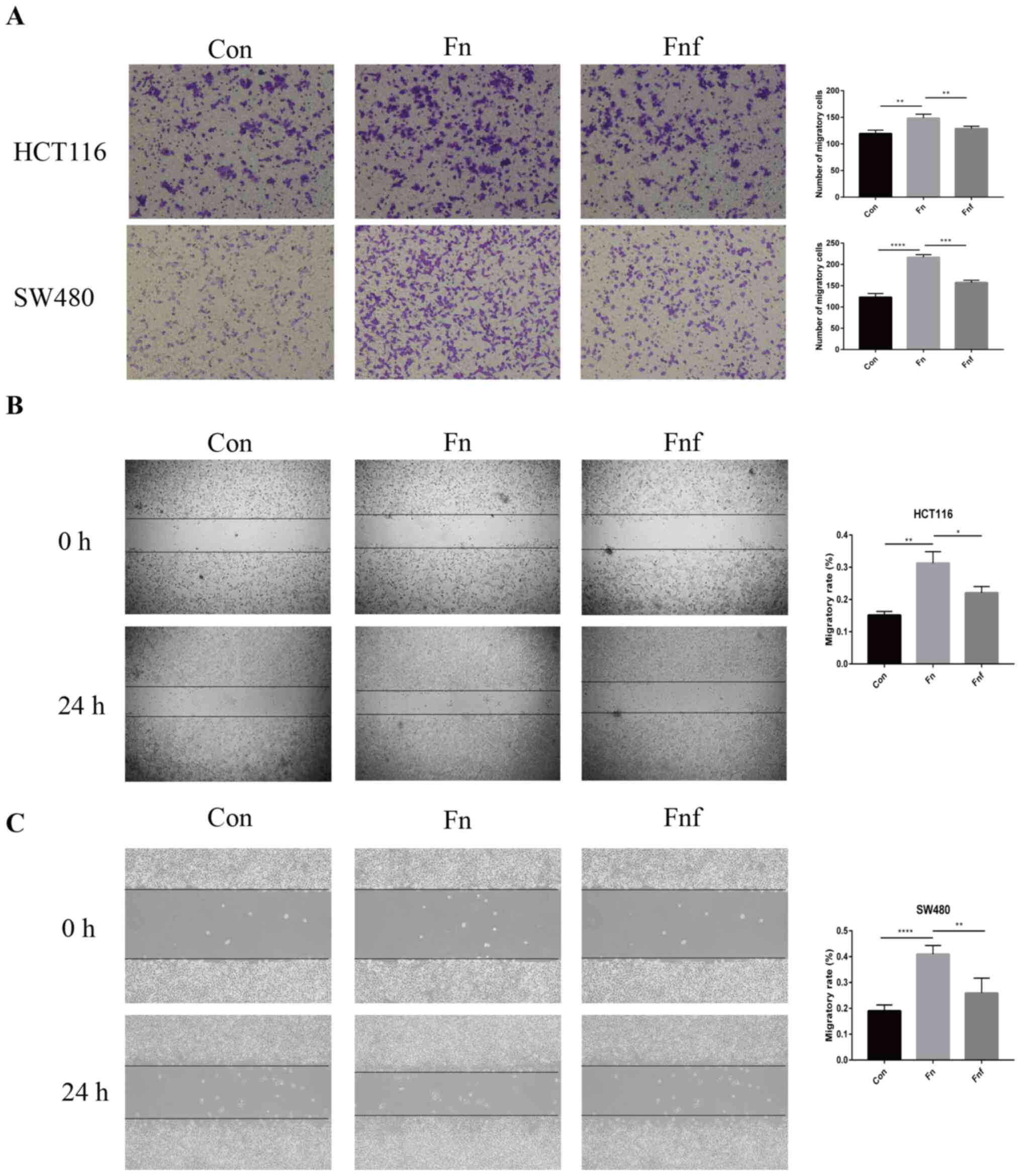
Fucose ameliorates the pro-migratory
ability of Fn on colon cancer cells
The migratory ability of colon cancer cells is a
functional characteristic to assess aggressiveness (14). The Transwell migration assay was
performed to assess the effect of Fn and Fnf on the
migratory ability of HCT116 and SW480 cells. The results
demonstrated that treatment with Fn significantly enhanced
the migratory ability of HCT116 and SW480 cells (HCT116 control,
119.30±3.84; Fn, 148.30±4.41, P=0.008; SW480 control,
123.00±4.93; Fn, 216.50±3.30; P<0.0001; Fig. 2A), while the migratory ability
decreased in cells treated with Fnf (HCT116 Fn,
148.30±4.41; Fnf, 128.80±2.29, P=0.008; SW480 Fn,
1216.50±3.30; Fnf, 157.70±2.96, P=0.002; Fig. 2A).
The results of the wound healing assay demonstrated
that treatment with Fn increased the migratory rate in
HCT116 cells compared with both the control cells and Fnf
treated cells (control, 0.152±0.006; Fn, 0.313±0.020;
Fnf, 0.221±0.011; P=0.002 and P=0.018; Fig. 2B). Similar trends were observed in
SW480 cells (control, 0.190±0.011; Fn, 0.410±0.016;
Fnf, 0.259±0.029; P<0.0001 and P=0.004; Fig. 2C). Taken together, these results
suggest that L-fucose may inhibit the pro-migratory ability of
Fn on colon cancer cells.
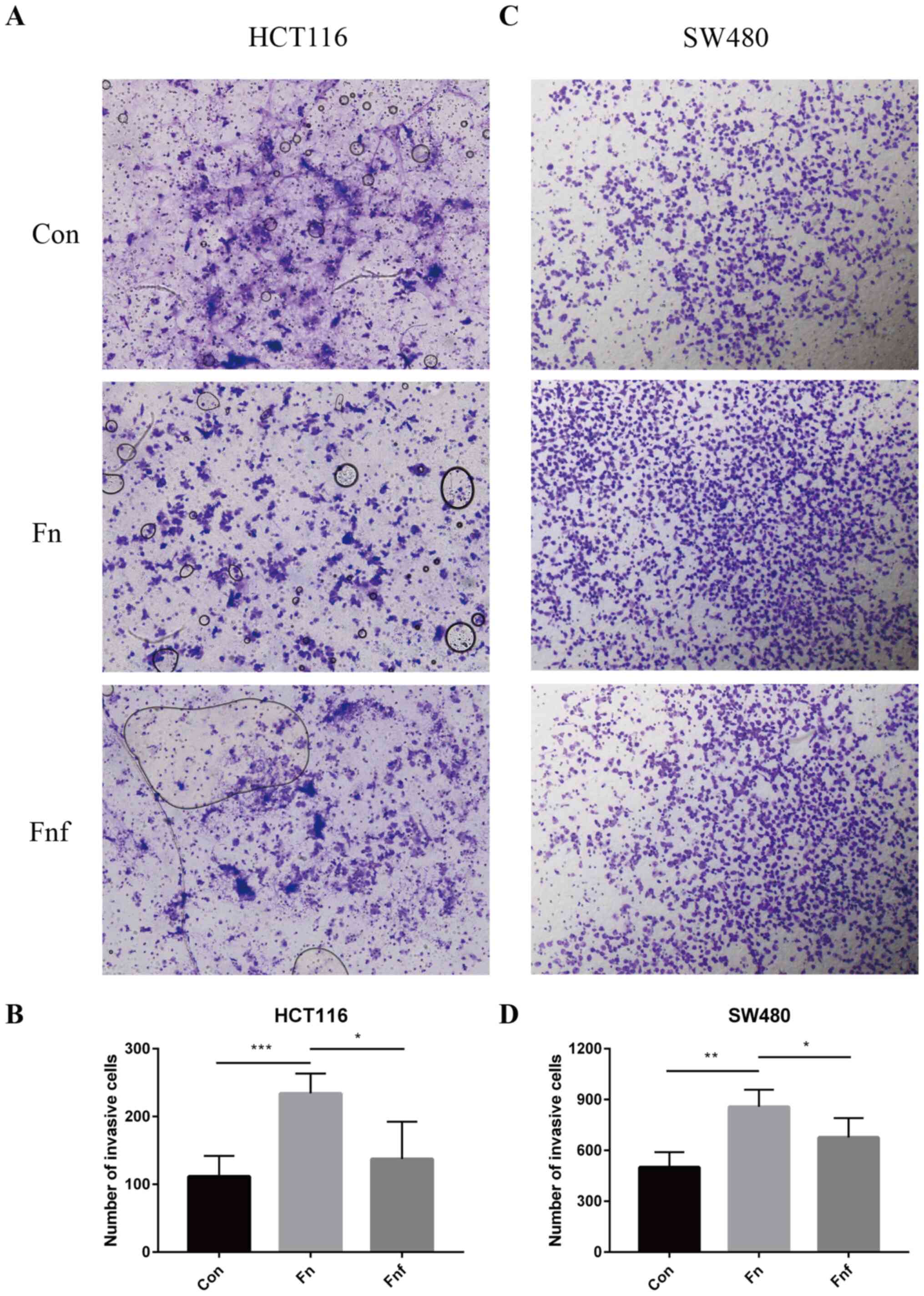
Fucose ameliorates the pro-invasive
ability of Fn on colon cancer cells
The Transwell invasive assay was performed to assess
the effect of Fn and Fnf on the invasive ability of
HCT116 and SW480 cells. The number of HCT116 and SW480 cells that
passed through the membrane was significantly higher in the
Fn group compared with the control group (HCT116 control,
111.50±15.16; Fn, 234.00±14.70, P=0.001; Fig. 3A and B; SW480 control, 500.00±44.77;
Fn, 857.80±50.50, P=0.002; Fig.
3C and D). As expected, the number of cells in the Fnf
group significantly decreased compared with the Fn group
(HCT116 Fn, 234.00±14.70; Fnf, 137.30±27.53, P=0.021;
Fig. 3A and B; SW480 Fn,
857.80±50.50; Fnf, 676.00±51.14, P=0.042; Fig. 3C and D). Collectively, these results
suggest that L-fucose may ameliorate the pro-invasive ability of
Fn on colon cancer cells.
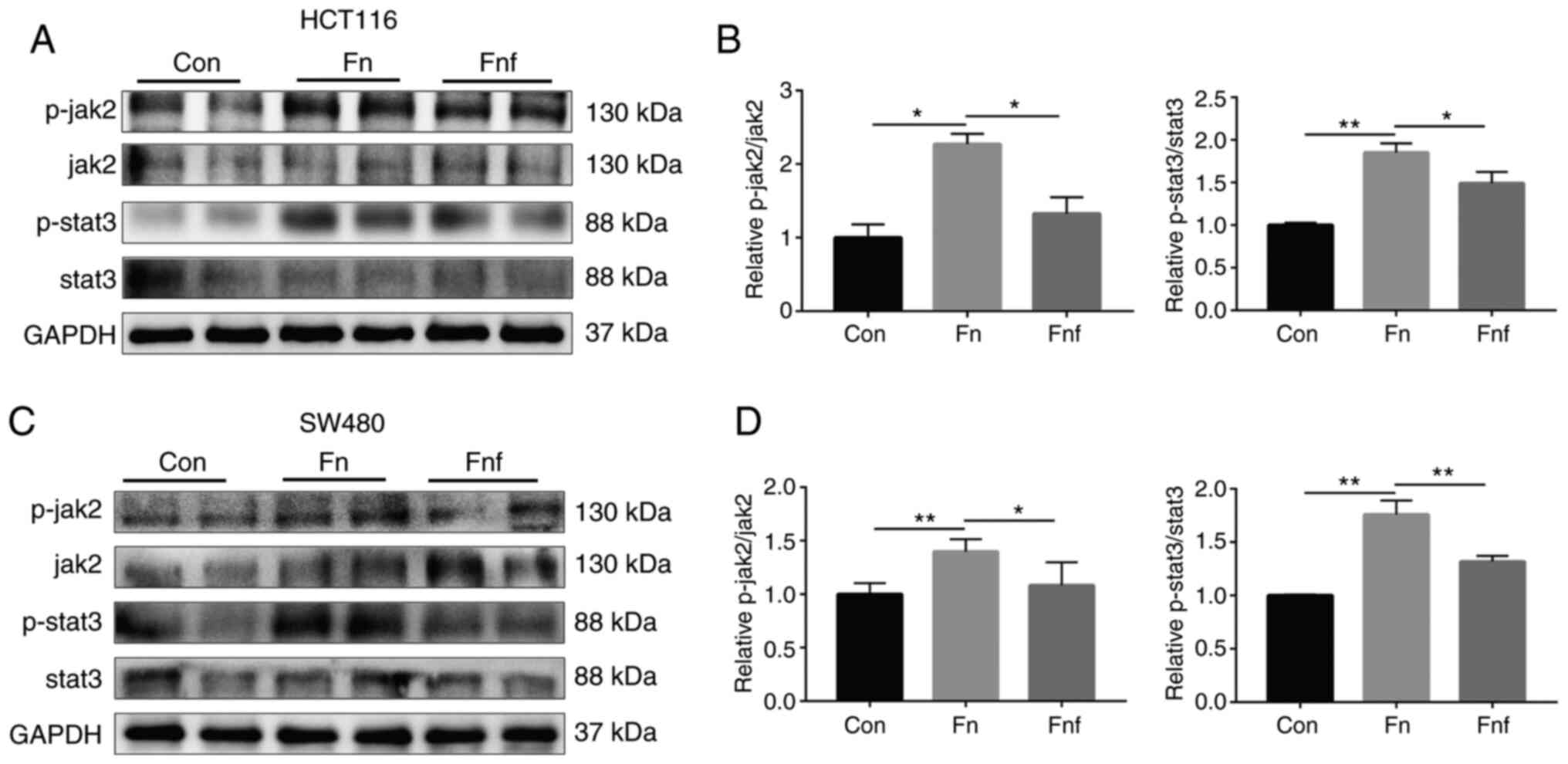
Fucose ameliorates the carcinogenic
property of Fn to activate stat3 and EMT
The effect of fucose on Fn potentiation on
activation of the pathway that promotes CRC progression was
assessed in the present study. Stat3 plays a crucial role in
tumorigenesis, and progression and invasion of cancer cells
(15). Thus, the expression and
phosphorylation of stat3 were detected via western blot analysis.
As presented in Fig. 4, the protein
levels of p-stat3 and p-jak2 significantly increased following
treatment with Fn (for p-stat3/stat3, HCT116 control,
1.000±0.020; Fn, 1.851±0.063; P=0.002; SW480 control,
1.000±0.006; Fn, 1.759±0.075; P=0.004. For p-jak2/jak2,
HCT116 control, 1.000±0.127; Fn, 2.272±0.098; P=0.016; SW480
control, 1.000±0.052; Fn, 1.400±0.057; P=0.002). However,
the protein levels significantly decreased following treatment with
fucose (for p-stat3/stat3, HCT116 Fn, 1.851±0.063;
Fnf, 1.490±0.095; P=0.044; SW480 Fn, 1.759±0.075;
Fnf, 1.318±0.031; P=0.005. For p-jak2/jak2, HCT116
Fn, 2.272±0.098; Fnf, 1.327±0.157; P=0.036; SW480
Fn, 1.400±0.057; Fnf, 1.082±0.108; P=0.040).
Stat3 activation is upstream of
epithelial-to-mesenchymal transition (EMT) in CRC, which is
associated with tumor progression (16). The present study assessed the
specific protein markers of EMT, which exist in different types of
tumors, such as prostate and colon cancer, and contribute to tumor
metastasis (17). As presented in
Fig. 5A and B, the expression levels
of N-cadherin, β-catenin and vimentin were higher in HCT116 cells
treated with Fn compared with the control group (N-cadherin
control, 1.00±0.01; Fn, 1.34±0.03, P=0.007; β-catenin
control, 1.00±0.02; Fn, 1.44±0.03, P=0.006; vimentin
control, 1.00±0.01; Fn, 2.19±0.09; P=0.006). As expected,
the expression levels of N-cadherin, β-catenin and vimentin
significantly decreased following treatment with fucose (N-cadherin
Fn, 1.34±0.03; Fnf, 1.20±0.01, P=0.048; β-catenin
Fn, 1.44±0.03; Fnf, 1.26±0.03, P=0.042; vimentin
Fn, 2.19±0.09; Fnf, 1.67±0.07, P=0.045). E-cadherin
was expressed at low levels in HCT116 cells treated with Fn
compared with the other two groups, which demonstrated the change
of the EMT pathway (E-cadherin control, 1.00±0.04; Fn,
0.37±0.03; Fnf, 0.72±0.07; P=0.005 and P=0.040). Similar
findings were observed in SW480 cells (Fig. 5C and D).
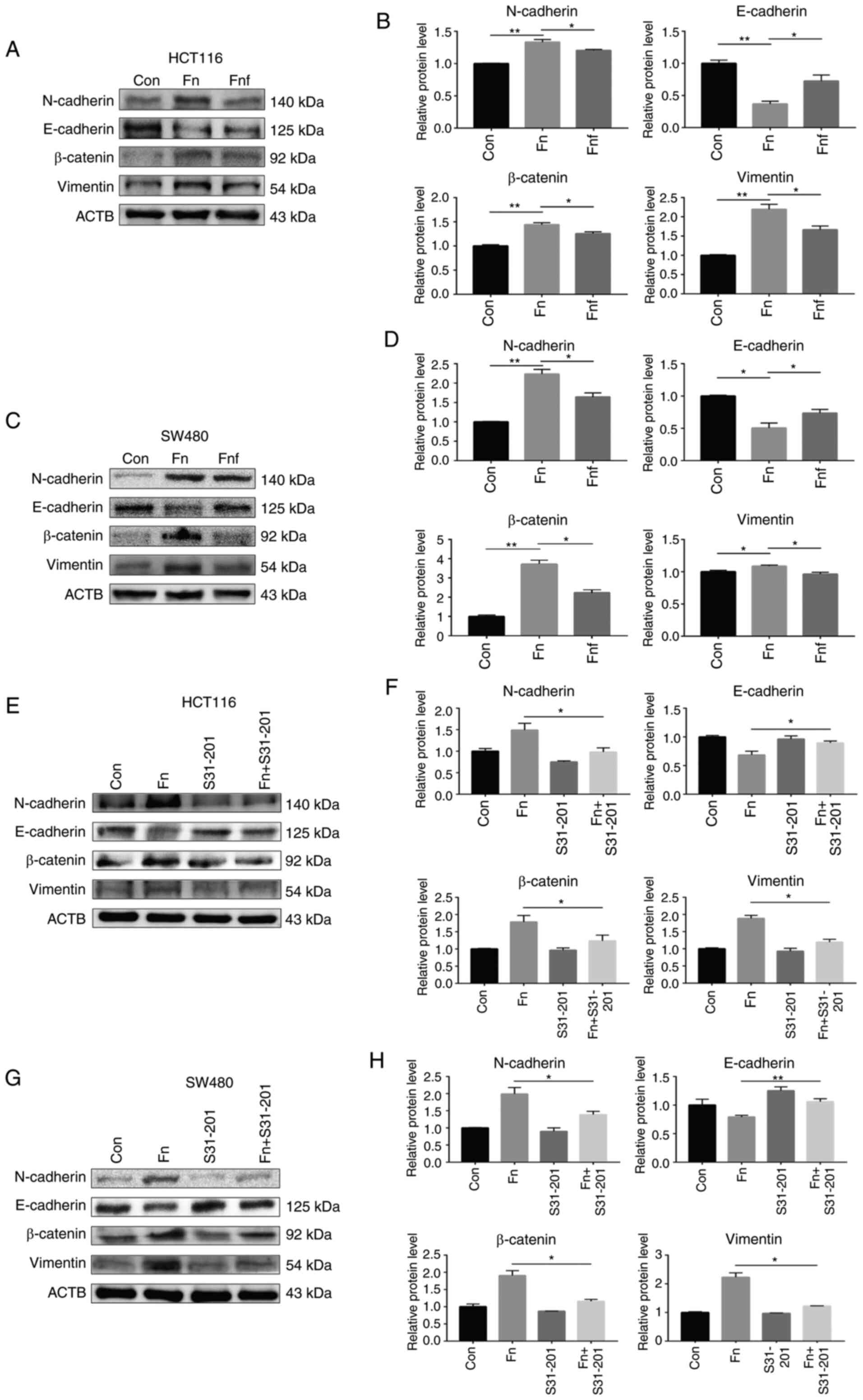 | Figure 5.Fucose ameliorates the carcinogenic
properties of Fn to activate epithelial-to-mesenchymal
transition. (A) Representative N-cadherin, E-cadherin, β-catenin
and vimentin immunoblots of HCT116 cells. Cells were treated with
supernatant of Fn or Fnf for 12 h. ACTB was used as
the loading control. (B) Relative protein levels are presented in
histograms. (C) Representative N-cadherin, E-cadherin, β-catenin
and vimentin immunoblots of SW480 cells. Cells were treated with
supernatant of Fn or Fnf for 12 h. ACTB was used as
the loading control. (D) Relative protein levels are presented in
histograms. (E) Representative N-cadherin, E-cadherin, β-catenin
and vimentin immunoblots of HCT116 cells. Cells were treated with
or without S3I-201 (100 nM) and supernatant of Fn. (F)
Relative protein levels are presented in histograms. (G)
Representative N-cadherin, E-cadherin, β-catenin and vimentin
immunoblots of SW480 cells. Cells were treated with or without
S3I-201 (100 nM) and supernatant of Fn. (H) Relative protein
levels are presented in histogram. Data are presented as the mean ±
standard error of the mean of at least three repeated experiments.
*P<0.05, **P<0.01. Fn, Fusobacterium nucleatum;
Fnf, Fn+L-fucose; Con, control. |
The stat3 inhibitor, S3I-201, was used to assess the
association between stat3 activation and EMT following treatment
with Fn. Western blot analysis demonstrated that inhibition
of stat3 activation significantly suppressed the activation of EMT
in both HCT116 and SW480 cells (Fig.
5E-H). Taken together, these results suggest that fucose
inhibits the carcinogenic properties of Fn to activate the
stat3 pathway and EMT.
Discussion
Recently, several studies have focused on the
molecular mechanisms of carcinogenesis of Fn (5,18,19);
however, methods for resisting the carcinogenic properties remain
largely unknown. The present study revealed a distinct role of
fucose in ameliorating the carcinogenic properties of Fn in
vitro.
The results of the present study demonstrated that
Fn significantly promoted proliferation, migration and
invasion of colon cancer cells. Fn was first observed in
oral cavity contributing to periodontal diseases (20). Recently, increasing evidence suggests
an association between Fn and CRC. For example, Mima et
al (21) reported that Fn
was detected in 76/598 (13.0%) colorectal carcinomas (stages I–IV)
and 19/558 (3.4%) adjacent non-tumor tissues. Furthermore, highly
enriched Fusobacterium in CRC tissues is associated with
microsatellite instability-high status (22). Fn enrichment may augment
myeloid-derived immune cells in CRC, which can inhibit T-cell
proliferation and induce apoptosis (23). In addition, Fn activates
toll-like receptor 4 signaling and downstream PAK1 and NF-κB in
CRC, thus increasing the proliferative and invasive abilities
(18,19). Consistent with these findings, the
results of the present study demonstrated the carcinogenic
properties of Fn.
Currently, as our research team discovered that
fucose is testified to competent in different types of diseases,
including colitis, renal ischemia/reperfusion injury and high-fat
diet-induced obesity and hepatic steatosis (10,24,25), it
has also been demonstrated to impact the microbial ecosystem
(11). Notably, fucose may decrease
pathogen virulence through certain bacterium's metabolic pathway,
such as Salmonella enterica (12). Thus, the present study assessed
whether L-fucose can restrain the ability of Fn to promote
CRC progression. The results demonstrated that Fn exhibited
less tendency to facilitate the proliferation, migration and
invasion of colon cancer cells following treatment with L-fucose.
To the best of our knowledge, the present study was the first to
demonstrate that L-fucose suppresses flora associated with colon
cancer.
Mechanistically, previous studies have demonstrated
that Fn induces stat3 expression in macrophages, resulting
in M2 polarization and increased tumor-immune cytokine secretion,
which alters the tumor microenvironment and promotes colorectal
tumor development (8,26). The expression of stat3 signaling was
assessed in the present study. Given that stat3 activation can
promote EMT (27), and EMT is an
important factor to drive carcinogenesis (16), the change in the expression levels of
EMT markers was assessed in the present study. The results
demonstrated that the protein expression levels of the EMT markers
significantly increased following treatment with Fn,
suggesting that Fn may promote EMT in colon cancer cells. In
addition, activation of stat3 signaling and EMT weakened following
treatment with L-fucose.
However, there were some limitations in this study.
On the one hand, in vivo experiments could further confirm
the conclusion and make the study more complete. On the other hand,
further studies are needed to explore the mechanism that how fucose
impair the carcinogenic properties of Fn. One hypothesis is
that it may alter its metabolism.
In conclusion, the results of the present study
demonstrated that L-fucose ameliorated the carcinogenic properties
of Fn by suppressing its ability to activate stat3 and EMT
of colon cancer cells in vitro. Thus, L-fucose may serve as
a novel therapeutic strategy of microflora-related colon
cancer.
Acknowledgements
Not applicable.
Funding
The present study was financially supported by the
National Natural Science Foundation of China (grant nos. 81800467,
81330014, 81720108006 and 81974062).
Availability of data and materials
The datasets used and analyzed during the present
study are available from the corresponding author upon reasonable
request.
Authors' contributions
CD performed the experiments, analyzed the data and
drafted the initial manuscript. XT, WW and WQ helped analyze the
data and revised the manuscript for important intellectual content.
XF and XD helped culture the bacteria. XH and CH designed the
present study, provided funding and obtained the grants. All
authors have read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
O'Keefe SJ: Diet, microorganisms and their
metabolites, and colon cancer. Nat Rev Gastroenterol Hepatol.
13:691–706. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mima K, Nishihara R, Qian ZR, Cao Y,
Sukawa Y, Nowak JA, Yang J, Dou R, Masugi Y, Song M, et al:
Fusobacterium nucleatum in colorectal carcinoma tissue and
patient prognosis. Gut. 65:1973–1980. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shang FM and Liu HL: Fusobacterium
nucleatum and colorectal cancer: A review. World J Gastrointest
Oncol. 10:71–81. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen Y, Chen Y, Zhang J, Cao P, Su W, Deng
Y, Zhan N, Fu X, Huang Y and Dong W: Fusobacterium nucleatum
promotes metastasis in colorectal cancer by activating autophagy
signaling via the upregulation of CARD3 expression. Theranostics.
10:323–339. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Brennan CA and Garrett WS: Gut microbiota,
inflammation, and colorectal cancer. Annu Rev Microbiol.
70:395–411. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Abed J, Emgård JE, Zamir G, Faroja M,
Almogy G, Grenov A, Sol A, Naor R, Pikarsky E, Atlan KA, et al:
Fap2 mediates fusobacterium nucleatum colorectal
adenocarcinoma enrichment by binding to tumor-expressed Gal-GalNAc.
Cell Host Microbe. 20:215–225. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yu YN, Yu TC, Zhao HJ, Sun TT, Chen HM,
Chen HY, An HF, Weng YR, Yu J, Li M, et al: Berberine may rescue
fusobacterium nucleatum-induced colorectal tumorigenesis by
modulating the tumor microenvironment. Oncotarget. 6:32013–32026.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Goto Y, Uematsu S and Kiyono H: Epithelial
glycosylation in gut homeostasis and inflammation. Nat Immunol.
17:1244–1251. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
He R, Li Y, Han C, Lin R, Qian W and Hou
X: L-Fucose ameliorates DSS-induced acute colitis via inhibiting
macrophage M1 polarization and inhibiting NLRP3 inflammasome and
NF-κB activation. Int Immunopharmacol. 73:379–388. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ke J, Li Y, Han C, He R, Lin R, Qian W and
Hou X: Fucose ameliorate intestinal inflammation through modulating
the crosstalk between bile acids and gut microbiota in a chronic
colitis murine model. Inflamm Bowel Dis. 26:863–873. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pickard JM, Maurice CF, Kinnebrew MA, Abt
MC, Schenten D, Golovkina TV, Bogatyrev SR, Ismagilov RF, Pamer EG,
Turnbaugh PJ and Chervonsky AV: Rapid fucosylation of intestinal
epithelium sustains host-commensal symbiosis in sickness. Nature.
514:638–641. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Neish AS: Microbes in gastrointestinal
health and disease. Gastroenterology. 136:65–80. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kyoung AK, Ryu YS, Piao MJ, Shilnikova K,
Kang HK, Yi JM, Boulanger M, Paolillo R, Bossis G, Yoon SY, et al:
DUOX2-mediated production of reactive oxygen species induces
epithelial mesenchymal transition in 5-fluorouracil resistant human
colon cancer cells. Redox Biol. 17:224–235. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yu H, Lee H, Herrmann A, Buettner R and
Jove R: Revisiting STAT3 signalling in cancer: New and unexpected
biological functions. Nat Rev Cancer. 14:736–746. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fan Y, Mao R and Yang J: NF-kB and STAT3
signaling pathways collaboratively link inflammation to cancer.
Protein Cell. 4:176–185. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen T, You Y, Jiang H and Wang ZZ:
Epithelial-mesenchymal transition (EMT): A biological process in
the development, stem cell differentiation, and tumorigenesis. J
Cell Physiol. 232:3261–3272. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen Y, Peng Y, Yu J, Chen T, Wu Y, Shi L,
Li Q, Wu J and Fu X: Invasive fusobacterium nucleatum
activates beta-catenin signaling in colorectal cancer via a
TLR4/P-PAK1 cascade. Oncotarget. 8:31802–31814. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang Y, Weng W, Peng J, Hong L, Yang L,
Toiyama Y, Gao R, Liu M, Yin M, Pan C, et al: Fusobacterium
nucleatum increases proliferation of colorectal cancer cells
and tumor development in mice by activating toll-like receptor 4
signaling to nuclear factor-κB, and up-regulating expression of
MicroRNA-21. Gastroenterology. 152:851–866.e824. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Brennan CA and Garrett WS:
Fusobacterium nucleatum-symbiont, opportunist and
oncobacterium. Nat Rev Microbiol. 17:156–166. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mima K, Sukawa Y, Nishihara R, Qian ZR,
Yamauchi M, Inamura K, Kim SA, Masuda A, Nowak JA, Nosho K, et al:
Fusobacterium nucleatum and T cells in colorectal carcinoma.
JAMA Oncol. 1:653–661. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nosho K, Sukawa Y, Adachi Y, Ito M,
Mitsuhashi K, Kurihara H, Kanno S, Yamamoto I, Ishigami K, Igarashi
H, et al: Association of fusobacterium nucleatum with
immunity and molecular alterations in colorectal cancer. World J
Gastroenterol. 22:557–566. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kostic AD, Chun E, Robertson L, Glickman
JN, Gallini CA, Michaud M, Clancy TE, Chung DC, Lochhead P, Hold
GL, et al: Fusobacterium nucleatum potentiates intestinal
tumorigenesis and modulates the tumor-immune microenvironment. Cell
Host Microbe. 14:207–215. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Howard MC, Nauser CL, Farrar CA, Wallis R
and Sacks SH: l-Fucose prevention of renal ischaemia/reperfusion
injury in Mice. FASEB J. 34:822–834. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wu G, Niu M, Tang W, Hu J, Wei G, He Z,
Chen Y, Jiang Y and Chen P: L-Fucose ameliorates high-fat
diet-induced obesity and hepatic steatosis in mice. J Transl Med.
16:3442018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen T, Li Q, Wu J, Wu Y, Peng W, Li H,
Wang J, Tang X, Peng Y and Fu X: Fusobacterium nucleatum
promotes M2 polarization of macrophages in the microenvironment of
colorectal tumours via a TLR4-dependent mechanism. Cancer Immunol
Immunother. 67:1635–1646. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Rokavec M, Öner MG, Li H, Jackstadt R,
Jiang L, Lodygin D, Kaller M, Horst D, Ziegler PK, Schwitalla S, et
al: IL-6R/STAT3/miR-34a feedback loop promotes EMT-mediated
colorectal cancer invasion and metastasis. J Clin Invest.
124:1853–1867. 2014. View Article : Google Scholar : PubMed/NCBI
|