Introduction
Glioblastoma refers to an astrocytic tumor that is
typically observed in the nervous system (1). Glioblastoma has a 97% mortality rate
during advanced age (>60 years) in Finland (2), not only because the brain and spinal
cord are complex and difficult-to-understand, but also because the
existing surgical techniques have limited benefits (3). Additionally, glioblastoma has a poor
prognosis, thereby contributing to the low 10.1% 3-year survival
rate and the high recurrence rate of patients with this cancer
(4,5). Recently, the survival rate of patients
with glioblastoma has considerably improved with the use of
standard surgical treatments, conventional radiotherapy and
medication such as temozolomide (6–8).
Increasing research on immunotherapy has also provided novel
insights into glioblastoma treatments (9). While considerable effort and notable
progress have been made to improve the outcomes of patients with
glioblastoma, researchers are yet to determine the underlying
molecular mechanisms that trigger the development of
glioblastoma.
Regarded as small non-coding single-stranded RNAs
encoded by an endogenous gene, microRNAs (miRNAs/miRs) were
discovered to play crucial regulatory roles in the development of
cancer by regulating mRNA degradation or translational inhibition
of downstream target genes (10–12).
Previous studies have reported that miRNAs act as tumor suppressors
or promoters in the abnormal biological process of glioblastoma
(13–15). Despite the discovery of several
miRNAs associated with the development and progression of glioma
(16–18), researchers are yet to investigate the
effects of other miRNAs on glioblastoma cells (19–21). For
example, miR-640 inhibits the formation of capillaries (22); however, its role in cancer
development has not yet been investigated. In their research, Li
et al (23) and Zhai et
al (24) both demonstrated that
miR-640 exerts tumor-suppressive effects in ovarian cancer and
hepatocellular carcinoma. In another study, miR-640 was reported to
enhance the development of prostate cancer (25). However, the role of miR-640 in
glioblastoma remains unclear.
Slit guidance ligand 1 (SLIT1) is located on
chromosome 10q24.1 and consists of 37 exons (26). SLIT1 was first identified as a member
of the axon guidance molecule SLITs family, which is predominantly
located in the central nervous system, and mediates axon branching,
elongation and metastatic repulsion of nerve cells (27,28).
Previous studies have demonstrated that SLITs regulate the
signaling pathways of different types of cancer, including breast
cancer and colorectal cancer (29,30). It
has also been reported that low levels of SLIT1, caused by
methylation, can further deteriorate colorectal cancer (31). SLITs exhibit different expression
levels in different subtypes of gastric cancer (32). Although Amodeo et al (33) demonstrated the involvement of SLIT1
in glioblastoma cell migration, the influence of SLIT1 on
glioblastoma is not yet fully understood.
The present study aimed to investigate the
underlying molecular mechanism of glioblastoma by assessing the
regulatory impact of miR-640 and its target genes on the
progression of glioblastoma cells. The results demonstrated that
miR-640 exerted a cancer-promoting effect on glioblastoma cells by
targeting SLIT1. Taken together, these results provide novel
insight to help decrease the mortality rate of patients with
glioblastoma.
Materials and methods
Microarray analysis
A total of two mRNA microarray datasets [GSE90886
(34) and GSE104291 (35)] were obtained from the National Center
for Biotechnology Information (https://www.ncbi.nlm.nih.gov/). The data were
subjected to microarray analysis using limma 3.26.8 (36), with P<0.05. Similarly, the
GSE103229 dataset (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE103229)
was downloaded from the Gene Expression Omnibus (GEO) database
(https://www.ncbi.nlm.nih.gov/gds), and
microarray analysis of the miRNAs was performed using limma 3.26.8,
with P<0.05 and log-fold change (logFC)>0. The data were
subsequently uploaded onto Metascape (https://metascape.org/gp/index.html#/main/step1),
UALCAN (http://ualcan.path.uab.edu/index.html) for The Cancer
Genome Atlas analysis, ENCORI Pan-Cancer Analysis Platform, ENCORI
Pan-Cancer Analysis Platform (http://starbase.sysu.edu.cn/panCancer.php), miRDB
(http://mirdb.org) and Venny 2.1.0 (https://bioinfogp.cnb.csic.es/tools/venny/) for gene
enrichment and gene analyses.
Tissue acquisition and cell
culture
The present study was approved by the Ethics
Committee of Puai Hospital (Wuhan, China) and written informed
consent was provided by all patients prior to the study start. A
total of 32 pairs of glioblastoma tissues and adjacent normal
tissues were collected from Puai Hospital between June 2017 and
January 2020. The clinical characteristics of the patients with
glioblastoma are presented in Table
I.
 | Table I.Clinicopathological characteristics
of patients with glioblastoma (n=32). |
Table I.
Clinicopathological characteristics
of patients with glioblastoma (n=32).
|
Characteristics | Number of patients,
n | Percentage, % |
|---|
| Age, years |
|
≤50 | 17 | 53.1 |
|
>50 | 15 | 46.9 |
| Sex |
|
Male | 19 | 59.4 |
|
Female | 13 | 40.6 |
| Tumor origin |
|
Primary | 21 | 65.6 |
|
Secondary | 11 | 34.4 |
| KPS value |
|
≤70 | 20 | 62.5 |
|
>70 | 12 | 37.5 |
| Location |
|
Frontal | 5 | 15.6 |
|
Temporal | 11 | 34.4 |
|
Parietal | 9 | 28.1 |
|
Other | 7 | 21.9 |
The glioblastoma cell lines (A172, U251, H4 and
SHG44) and normal astrocyte cell line (HA) were purchased from the
BeNa Culture Collection. H4, A172, SHG44 and HA cells were
maintained in DMEM (cat. no. E600004; Sangon Biotech Co., Ltd.)
supplemented with 10% fetal bovine serum (FBS) and 100 U/ml
streptomycin (both Gibco; Thermo Fisher Scientific, Inc.), at 37°C
in 5% CO2. U251 cells were maintained in MEM-EBSS medium
(cat. no. E600020; Sangon Biotech Co., Ltd.) supplemented with 10%
FBS and 100 U/ml streptomycin, at 37°C in 5% CO2.
Cell transfection
miR-640 inhibitor, miR-640 mimic and their
corresponding negative controls (mimic-NC and inhibitor-NC) were
purchased from Guangzhou RiboBio Co., Ltd. SLIT1 overexpression
plasmid and empty vector were purchased from GeneCopoeia, Inc. All
sequences are listed in Table SI.
A172 and U251 cells were transfected with 50 nM inhibitor-NC, 50 nM
miR-640 inhibitor, 50 nM mimic-NC, 50 nM miR-640 mimic and 2.5 µg
SLIT1 overexpression plasmid or empty vector using
Lipofectamine® 2000 reagent (cat. no. 11668019; Thermo
Fisher Scientific, Inc.) at room temperature, according to the
manufacturer's instructions. Following incubation for 2 days at
37°C, transfection efficiency was assessed via reverse
transcription-quantitative (RT-q)PCR analysis. Due to similar
transfection efficiencies of inhibitor-NC, mimic-NC or empty vector
(Fig. S1), cells in the NC group
were co-transfected with inhibitor-NC and mimic-NC. Subsequent
experiments were performed 48 h after transfection.
RT-qPCR
Total RNA from cells and tissues was extracted using
TRIzol® reagent (cat. no. 15596026; Thermo Fisher
Scientific, Inc.) and reverse transcribed into cDNA. miRNA was
reverse-transcribed using the mirVana™ qRT-PCR miRNA Detection kit
(cat. no. AM1558, Invitrogen; Thermo Fisher Scientific, Inc.),
according to the manufacturer's instructions. The reaction
condition was as follows: 37°C for 30 min and 95°C for 10 min. mRNA
was reverse-transcribed using SuperScript™ III First-Strand
Synthesis SuperMix (cat. no. 11752050; Thermo Fisher Scientific,
Inc.). The reaction condition was as follows: 25°C for 10 min, 50°C
for 30 min and 85°C for 5 min. qPCR was subsequently performed
using the SYBR Premix Ex Taq (cat. no. RR420A; Takara Biotechnology
Co., Ltd.) using the StepOnePlus™ Real-Time PCR System (cat. no.
4376600; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. The reaction condition was as follows:
95°C for 30 sec, followed by 40 cycles at 95°C for 5 sec and 60°C
for 30 sec. The primer sequences used for qPCR are listed in
Table II. Relative miRNA and mRNA
expression levels were calculated using the 2−ΔΔCq
method (37). U6 and β-actin were
used as the internal controls for miRNA and mRNA, respectively.
| Table II.Primer sequences used for
quantitative PCR. |
Table II.
Primer sequences used for
quantitative PCR.
| Gene | Primer sequences
(5′-3′) |
|---|
| miR-640 | Forward:
GTGACCCTGGGCAAGTTCCT |
|
| Reverse:
CCCCAAGGCAACCGTAGAGG |
| SLIT1 | Forward:
CTGCTCCCCGGATATGAACC |
|
| Reverse:
TAGCATGCACTCACACCTGG |
| U6 | Forward:
CTCGCTTCGGCAGCACA |
|
| Reverse:
AACGCTTCACGAATTTGCGT |
| β-actin | Forward:
TCGTGCGTGACATTAAGGAG |
|
| Reverse:
GTCAGGCAGCTCGTAGCTCT |
Cell Counting Kit-8 (CCK-8) assay
The CCK-8 assay was performed to assess cell
viability. A172 and U251 cell suspensions (100 µl) in a logarithmic
growth phase were seeded into 96-well plates at a density of 2,000
cells/well. Following incubation overnight at 37°C, cells were
transfected with miR-640 mimic, miR-640 inhibitor and corresponding
NCs, or SLIT1 overexpression plasmid and corresponding empty vector
for 0, 24, 48 and 72 h at 37°C. CCK-8 solution (10 µl; cat. no.
E606335; Sangon Biotech Co., Ltd.) was added to each well,
according to the manufacturer's instructions. Following incubation
for 2 h, cell viability was analyzed at a wavelength of 450 nm,
using a microplate reader.
BrdU ELISA assay
The BrdU ELISA assay was performed to assess cell
proliferation, using the BrdU Cell Proliferation ELISA kit (cat.
no. ab126556; Abcam). Transfected cells were seeded into 96-well
plates at a density of 3,000 cells/well and incubated for 2 days in
DMEM for A172 cells and MEM-EBSS medium for U251 cells at 37°C.
Subsequently, cells were incubated with 100 µl 1X BrdU reagent
(included in the kit) for 2 h at 37°C. The medium was discarded and
200 µl/well fixing solution (included in the kit) was added to the
cells, which were incubated for 30 min at 25°C. Cells were washed
three times with PBS, prior to incubation with 100 µl/well
anti-BrdU monoclonal detector antibody (1:1,000; included in the
kit) for 1 h at 25°C. Subsequently, 100 µl/well goat anti-mouse IgG
conjugate (1:2,000; included in the kit) was added and cells were
incubated for 30 min at 25°C. Cells were subsequently incubated
with 100 µl/well TMB peroxidase substrate for 30 min in the dark at
25°C and analyzed at a wavelength of 450 nm, using a microplate
reader.
Cell adhesion assay
For the cell adhesion assay, a 96-well plate was
pre-coated with 10 µg/ml collagen I (cat. no. C7661; Sigma-Aldrich;
Merck KGaA) and blocked with 1% BSA (Beijing Solarbio Science &
Technology Co., Ltd.) overnight at 4°C. The transfected cells were
suspended in serum-free medium and seeded into the pre-coated
96-well plate at a density of 1×105 cells/well. After 1
h, the adherent cells were fixed with 4% paraformaldehyde for 15
min at 25°C, while the non-adherent cells were removed using PBS.
The fixed cells were subsequently stained with 2.5% crystal violet
for 20 min at 25°C in methanol and measured at a wavelength of 450
nm, using a microplate reader.
Caspase-3 activity assay
Caspase-3 activity was measured to assess cell
apoptosis, as it is highly expressed during cell apoptosis
(38). The Caspase-3 Assay kit (cat.
no. ab39401; Abcam) was used to measure caspase-3 activity.
Following cell transfection for 48 h, 1×106 cells were
collected and incubated with 50 µl cell lysis buffer for 15 min at
25°C. Protein concentration was measured and adjusted to 50–200 µg
protein per 50 µl reaction buffer. Cell lysates were subsequently
incubated with 50 µl reaction buffer supplemented with 10 mM DTT
and 5 µl 4 mM DEVD-p-NA substrate at 25°C for 30 min. Caspase-3
activity was measured at a wavelength of 405 nm, using a microplate
reader.
Dual-luciferase reporter assay
A Dual-luciferase reporter system was used to assess
the luciferase activity of cells. miRDB predicted two binding sites
of SLIT1 3′-untranslated region (UTR) or the area to which miR-640
could bind. These two binding sites (2442–2448 and 2945–2952) of
SLIT1 3′-UTR were mutated using the QuikChange XL Site-Directed
Mutagenesis kit (cat. no. 200522; Agilent Technologies, Inc.). The
wild-type SLIT1 3′-UTR and mutant SLIT1 3′-UTR were inserted into
the pEZX-MT05 reporter plasmids (GeneCopoeia, Inc.) and
subsequently co-transfected into A172 and U251 cells with miR-640
mimic using Lipofectamine® 2000 reagent (cat. no.
11668019; Thermo Fisher Scientific, Inc.). Following incubation for
48 h at 37°C, luciferase activity was detected using the
Secrete-Pair Dual Luminescence Assay kit (cat. no. LF031;
GeneCopoeia, Inc.). The activity of Gaussia Luciferase was
normalized to the activity of secreted alkaline phosphatase.
RNA pull-down assay
The RNA pull-down assay was performed to verify the
association between SLIT1 and miR-640. Briefly, 0.5 ml of 25 mM
Tris-HCl, 0.05% NP-40, 70 mM KCl, 2.5 mM EDTA, 80 U/ml RNase
inhibitor and 1X protease inhibitor cocktail (cat. no. 539136;
Sigma-Aldrich; Merck KGaA) were used to incubate 3×106
A172 and U251 cells for 20 min on ice. The mixture was centrifuged
at 1,400 × g at 4°C for 20 min and the supernatant lysate was
collected. Subsequently, 10 nM biotin-coupled miR-22-3p
(bio-miR-22-3p) or biotin-coupled NC (bio-NC) (Guangzhou RiboBio
Co., Ltd.) was added to the supernatant lysate and incubated for 2
h at 25°C. Following incubation, 100 µl streptavidin magnetic beads
(cat. no. 88816; Thermo Fisher Scientific, Inc.) were added to the
supernatant lysate. After 4 h, the beads were washed with PBS and
the expression of pull-down RNA was assessed via RT-qPCR
analysis.
Western blotting
Total protein was extracted from cells (A172, H4,
U251, SHG44 and HA) and tissues (normal and glioblastoma tissues)
using RIPA buffer (Keygene N.V.), according to the manufacturer's
instructions. Total protein was separated via SDS-PAGE, transferred
onto cellulose nitrate membranes (Bio-Rad Laboratories, Inc.) and
blocked with 5% non-fat milk in TBST buffer (TBS containing 0.1%
Tween-20) for 1 h at room temperature. The membranes were incubated
with primary antibodies against GAPDH (cat. no. ab8245; 1:2,500;
Abcam) and SLIT1 (cat. no. ab151724; 1:2,000; Abcam) overnight at
4°C. Membranes were washed three times with TBST buffer. Following
the primary incubation, membranes were incubated with goat
anti-rabbit IgG (cat. no. ab97051) and goat anti-mouse IgG (cat.
no. ab175783) secondary antibodies labeled with HRP (both 1:10,000;
Abcam) at room temperature for 1 h. Protein blots were subsequently
washed with TBST buffer and visualized using ECL-Plus reagent (cat.
no. 345818; EMD Millipore).
Statistical analysis
All experiments were performed in triplicate and
data are presented as the mean ± standard deviation. Data were
analyzed using SPSS 19.0 software (IBM Corp.). Log-rank test was
performed for prognosis analysis. A paired or unpaired two-tailed
Student's t-test was used to compare differences between two groups
of paired tissues or unpaired cells, respectively, while one-way
ANOVA followed by Dunnett's or Tukey's post-hoc test was used to
compare differences among multiple groups. Spearman's correlation
analysis was used for analyzing the correlation between SLIT1 and
miR-640 expression. P<0.05 was considered to indicate a
statistically significant difference.
Results
SLIT1 and miR-640 are key regulators
of glioblastoma
The GSE90886 and GSE104291 datasets were downloaded
from the GEO database to identify the key gene in glioblastoma.
Venny 2.1.0 identified 362 common differentially expressed genes
(DEGs) that overlapped between the two mRNAs microarray (Fig. 1A). The 362 DEGs were subsequently
uploaded onto Metascape for biological processes analysis. The
central nervous system neuron development, which involves eight
DEGs, was selected as the key biological process as glioblastoma is
associated with central nervous system neuron development (39) (Fig.
1B). Based on The Cancer Genome Atlas data (https://www.cancer.gov/about-nci/organization/ccg/research/structural-genomics/tcga)
on glioblastoma multiforme, low SLIT1 expression and high NRP1
expression were observed in glioblastoma multiforme (Fig. 1C). Due to its association with poor
prognosis, SLIT1 was investigated in the present study (Fig. 1D). The GSE103229 dataset was used to
identify the differentially expressed miRNAs, while miRDB was used
to predict the miRNAs binding to SLIT1. hsa-miR-640 was identified
as the key miRNA in glioblastoma (Fig.
1E), so its role was further investigated in the present
study.
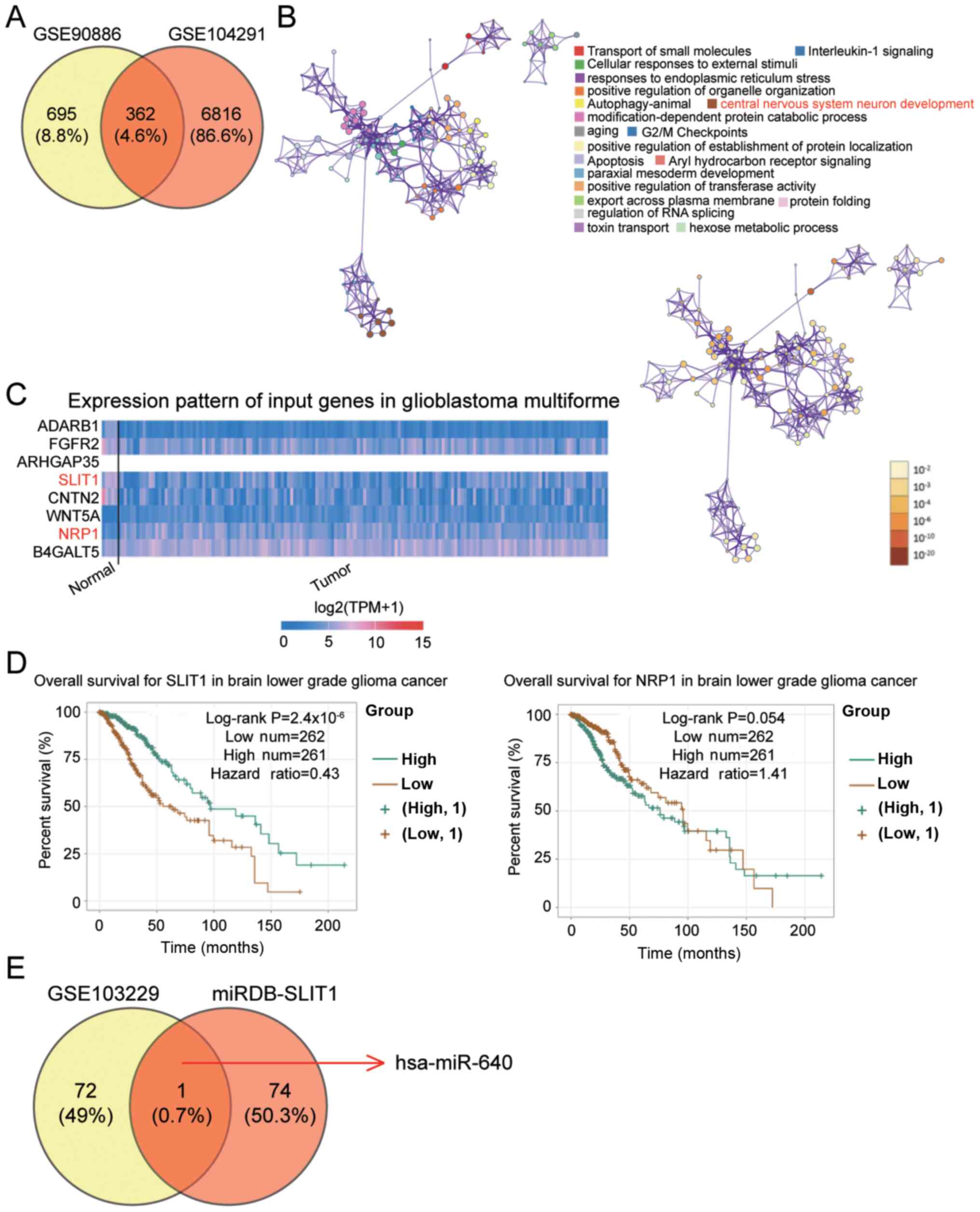 | Figure 1.SLIT1 and miR-640 are associated with
glioblastoma. (A) A total of 362 DEGs were identified to overlap
between the GSE90886 and GSE104291 datasets using Venny 2.1.0. (B)
The central nervous system neuron development involving eight DEGs
(ADARB1, FGFR2, ARHGAP35, SLIT1, CNTN2, WNT5A, NRP1 and B4GALT5)
was selected as the key biological process using Metascape. (C)
SLIT1 and NRP1 were abnormally expressed in glioblastoma multiforme
based on The Cancer Genome Atlas analysis. (D) Low SLIT1 expression
was significantly associated with poor prognosis of brain lower
grade glioma cancer using the ENCORI Pan-Cancer Analysis platform.
(E) miR-640 was identified as the key miRNA that could bind to the
SLIT1 3′-UTR, and was associated with glioblastoma. The GSE103229
dataset was the miRNA expression profile. miRDB was used to predict
the miRNAs binding to mRNA 3′-UTR. SLIT1, Slit guidance ligand 1;
miR, microRNA; UTR, untranslated region. |
miR-640 promotes the malignant
phenotype of glioblastoma cells
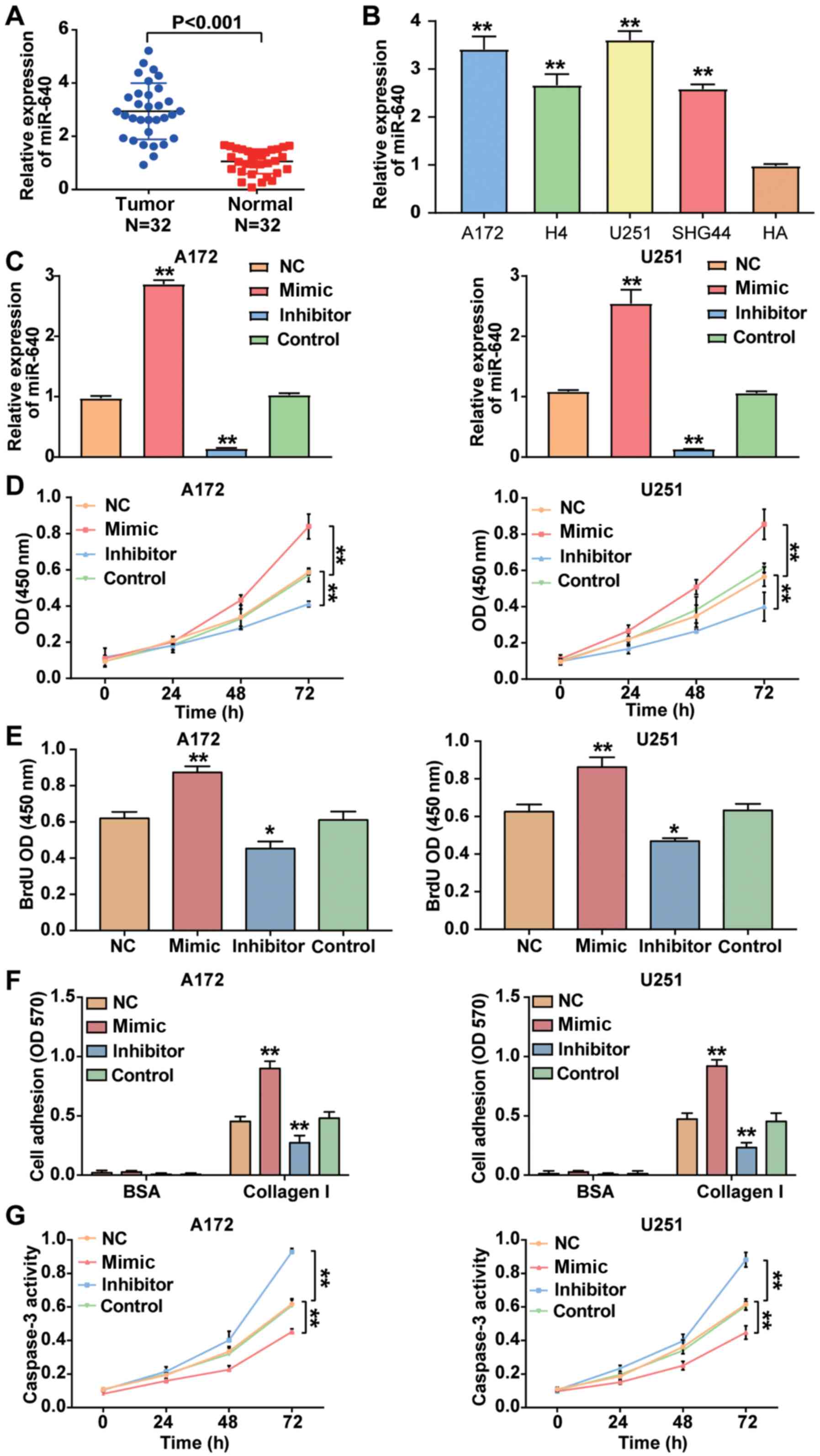
RT-qPCR analysis was performed to detect miR-640
expression in glioblastoma tissues and cells. The result
demonstrated that miRNA expression was higher in glioblastoma
tissues by 2.78-fold compared with normal tissues (P<0.001;
Fig. 2A). Similarly, miR-640
expression was significantly higher in the glioblastoma cell lines
(A172, H4, U251 and SHG44) compared with the normal astrocyte HA
cell line (P<0.001; Fig. 2B).
Given that miR-640 expression was higher in A172 and U251 cells
compared with the other glioblastoma cell lines, the A172 and U251
cell lines were selected for subsequent cytological experiments.
Following transfection of A172 and U251 cells with miR-640 mimic,
miR-640 inhibitor and plasmids, the results demonstrated that
miR-640 expression increased by ~2.5-fold in A172 and U251 cells
transfected with miR-640 mimic, while miR-640 expression decreased
by ~85% in A172 and U251 cells transfected with miR-640 inhibitor
(P<0.001; Fig. 2C). The results
of the CCK-8 assay demonstrated that overexpression of miR-640
enhanced cell viability, the effects of which were reversed
following downregulation of miR-640 (P<0.001; Fig. 2D).
The results of the BrdU ELISA assay demonstrated
that overexpression of miR-640 promoted A172 and U251 cell
proliferation, the effects of which were reversed following
downregulation of miR-640 (Fig. 2E).
The cell adhesion assay demonstrated that the cell adhesive ability
was enhanced by almost 2-fold in A172 and U251 cells transfected
with miR-640 mimic, while the cell adhesive ability was impaired by
~50% following transfection with miR-640 inhibitor (P<0.001;
Fig. 2F). Caspase-3 activity assay
was measured to assess cell apoptosis, as caspase-3 activity is
enhanced during cell apoptosis (37). The results demonstrated that
caspase-3 activity was decreased in glioblastoma cells transfected
with miR-640 mimic, while it was elevated in glioblastoma cells
transfected with miR-640 inhibitor compared with the control group
(P<0.001; Fig. 2G). Taken
together, these results suggest that miR-640 acts as an oncogene in
glioblastoma cells.
Association between miR-640 and
SLIT1
miRDB analysis predicted that SLIT1 3′-UTR contained
two binding sites to which miR-640 could bind (Fig. 3A). The two binding sites in SLIT1
3′-UTR were co-mutated and subsequently inserted into the
luciferase reporter plasmids. The results of the dual-luciferase
reporter assay demonstrated that fluorescence intensity decreased
>50% in A172 and U251 cells co-transfected with wild-type SLIT1
3′-UTR and miR-640 mimic (P<0.001), whereas the targeting effect
of miR-640 declined when the two binding sites of SLIT1 3′-UTR were
mutated (Fig. 3B). The results of
the RNA pull-down assay demonstrated that SLIT1 mRNA expression was
enriched in A172 and U251 cells transfected with biotin-coupled
miR-640 (P<0.001; Fig. 3C).
Furthermore, RT-qPCR analysis revealed that SLIT1 mRNA expression
was downregulated by 55% in glioblastoma tissue samples
(P<0.001; Fig. 3D), and SLIT1
expression was negatively correlated with miR-640 expression in
glioblastoma tissues (Fig. 3E).
Western blot analysis demonstrated that SLIT1 expression was
downregulated in glioblastoma tissues compared with normal tissues
(P<0.05; Fig. 3F). Similarly,
RT-qPCR (P<0.001) and western blot (P<0.05) analyses
indicated that SILT1 expression decreased in the glioblastoma cell
lines (A172, H4, U251 and SHG44) compared with the normal astrocyte
cell line (HA) (Fig. 3G and H).
Collectively, these results suggest that SLIT1 is directly
suppressed by miR-640 as a target downstream in glioblastoma.
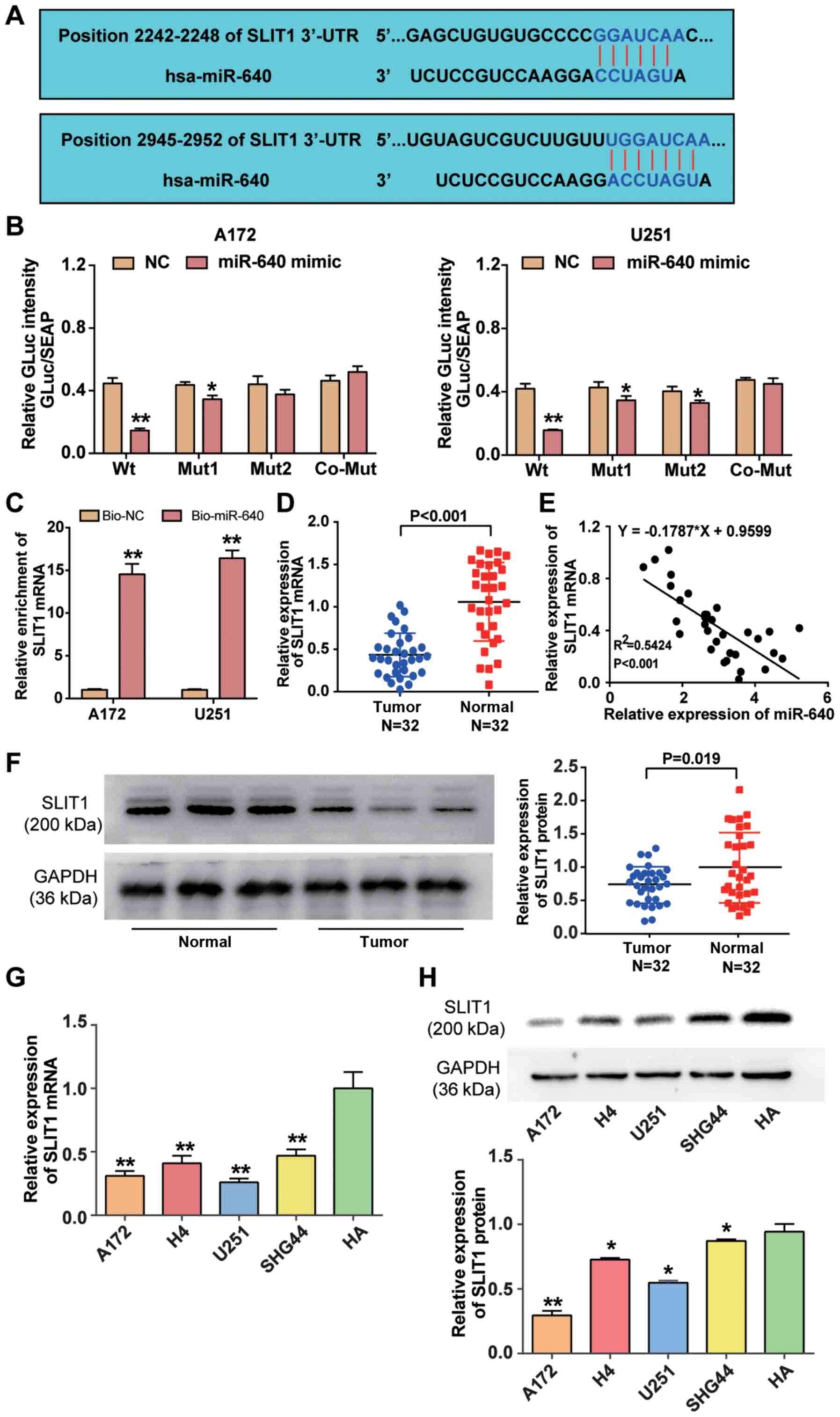 | Figure 3.miR-640 directly binds to SLIT1
3′-UTR. (A) The potential binding site between miR-640 and SLIT1
was predicted using miRDB. (B) The potential binding site between
miR-640 and 3′-UTR of SLIT1 was determined via the Dual-luciferase
reporter assay. (C) The RNA pull-down assay was performed to detect
SLIT1 enrichment in A172 and U251 cells. (D) RT-qPCR analysis was
performed to detect SLIT1 expression in glioblastoma tissues and
normal tissues. (E) Spearman's correlation analysis was performed
to determine the association between miR-640 and SLIT1 expression
levels. (F) Western blot analysis was performed to detect SLIT1
protein expression in glioblastoma tissues and normal tissues. (G)
RT-qPCR analysis was performed to detect SLIT1 mRNA expression in
the glioblastoma cell lines (A172, H4, U251 and SHG44) and normal
astrocyte HA cells. **P<0.001 vs. HA. (H) The expression level
of SLIT1 protein in glioblastoma cell lines (A172, H4, U251 and
SHG44) and normal astrocyte cell line HA was measured using a
western blot system. Data are presented as the mean ± standard
deviation (n=3). *P<0.05 vs. co-transfection with Wt and miR-640
mimic or HA; **P<0.001 vs. co-transfection with Wt and miR-640
mimic, Bio-NC or HA. miR, microRNA; SLIT1, Slit guidance ligand 1;
UTR, untranslated region; Wt, wild-type; Mut, mutant; Bio-NC,
biotin-coupled negative control; RT-qPCR, reverse
transcription-quantitative PCR. |
miR-640 contributes to the development
of glioblastoma by inhibiting SLIT1
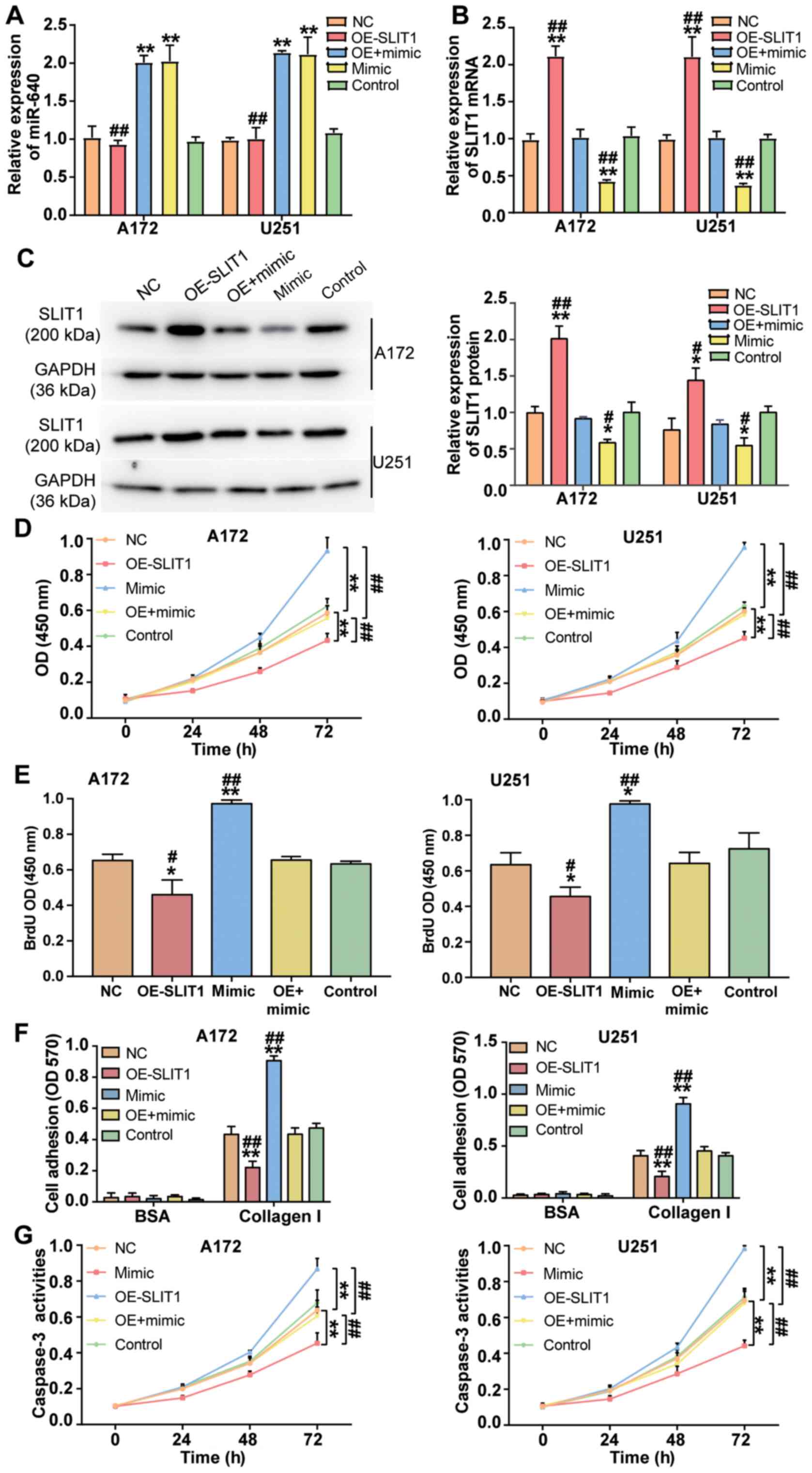
To investigate the effect of miR-640 on glioblastoma
cells, miR-640 mimic and SLIT1 overexpression plasmid were
transfected or co-transfected into A172 and U251 cells. RT-qPCR
analysis demonstrated that transfection with SLIT1 overexpression
plasmid did not affect miR-640 expression (P<0.001; Fig. 4A). Conversely, SLIT1 overexpression
plasmid enhanced SLIT1 expression by 2.1-fold, while transfection
with miR-640 mimic decreased SLIT1 expression by ~60% in both
glioblastoma cell lines (P<0.001; Fig. 4B). Western blot analysis demonstrated
that transfection with SLIT1 overexpression plasmid increased SLIT1
protein expression, while overexpression of miR-640 decreased SLIT1
protein expression in A172 and U251 cells (P<0.05; Fig. 4C). These results indicate that A172
and U251 cells were successfully transfected or co-transfected with
miR-640 mimic and SLIT1 overexpression plasmid.
Several cytological experiments were performed in
transfected A172 and U251 cells. The results of the CCK-8 assay
demonstrated that the promotive effect of miR-640 mimic on cell
viability was attenuated following transfection with SLIT1
overexpression plasmid in A172 and U251 cells (P<0.001; Fig. 4D). Similarly, the results of the BrdU
ELISA assay demonstrated that the promotive effect of miR-640 mimic
on cell proliferation was relieved following transfection with
SLIT1 overexpression plasmid (P<0.05; Fig. 4E). The cell adhesive ability in the
co-transfection group was impaired by ~50% compared with the
miR-640 mimic group (P<0.001; Fig.
4F). Notably, the inhibitory effect of miR-640 mimic on
caspase-3 activity was attenuated in both A172 and U251 cells
following transfection with SLIT1 overexpression plasmid
(P<0.001; Fig. 4G). Taken
together, these results suggest that the enhanced function of
miR-640 in the progression of glioblastoma cells is repressed
following overexpression of SLIT1.
Discussion
The results of the present study demonstrated high
miR-640 expression and the low SLIT1 expression in glioblastoma
tissues and cells. Furthermore, miR-640 was demonstrated to enhance
cell proliferation and adhesion, but impaired apoptosis in
glioblastoma cells. In addition, SLIT1 exhibited a suppressive role
in glioblastoma cells, and miR-640 was demonstrated to bind to
SLIT1 3′-UTR, thereby suppressing SLIT1 expression. Overexpression
of SLIT1 attenuated the promoting effect of miR-640 on glioblastoma
cells.
The effect of miR-640 on different types of human
cancer has been extensively studied. For example, by performing
miRNA microarray analysis, Li et al (23) demonstrated that miR-640 expression is
decreased in paclitaxel-resistant ovarian carcinoma samples, and
that high miR-640 expression yields a more favorable prognosis
(23). Furthermore, miR-640
expression has been reported to be downregulated in hepatocellular
carcinoma (HCC) tissues, which contributes to the development of
HCC cells (24). Lodes et al
(25) used a pan-human miRNA
microarray to assess miRNA expression in five forms of human
cancers, including prostate carcinoma, colon carcinoma, ovarian
carcinoma, breast carcinoma and lung carcinoma. The results
demonstrated that miR-640 is overexpressed in serum from patients
with prostate cancer. Taken together, these findings suggest that
miR-640 plays different roles in different types of cancer. In
glioblastoma, miR-640 was upregulated using microarray analysis
(40). Consistent with this finding,
the results of the microarray and RT-qPCR analyses performed in the
present study demonstrated that miR-640 expression was upregulated
in glioblastoma. In addition, the CCK-8, BrdU (41,42) and
cell adhesion (43) assays were
performed to determine the molecular mechanism of miR-640 in
glioblastoma cells. The results of these experiments demonstrated
that overexpression of miR-640 promoted the viability,
proliferation and adhesion of glioblastoma cells, while
interference with miR-640 inhibited the proliferation and adhesion
of these cells. Caspases, which are key regulators of mammalian
apoptosis, are activated following cell injury and are responsible
for regulating apoptosis (44).
Caspase-3 is the main mediator of apoptosis, and it is responsible
for the proteolysis of several downstream key proteins associated
with apoptosis (45). To determine
the molecular mechanism of miR-640-induced apoptosis in
glioblastoma multiforme cells, caspase-3 activity was analyzed. The
results demonstrated that by interfering with A172 and U251 cells,
miR-640 could promote the activity of caspase-3; however, these
effects were reversed following overexpression of miR-640.
Therefore, it was concluded that miR-640 contributed to cell
proliferation and adhesion, and impaired apoptosis in glioblastoma
cells.
Previous studies have demonstrated that SLIT1
participates in intracellular metabolism and nerve cell elongation,
branching and functional regeneration through its receptor, Robo
(30,46,47). Kim
et al (32) reported that
SLIT1 is methylated in different types of human cancer, and in 11
gastric cancer cell lines. In addition, SLIT1 methylation occurs in
patients with early gastric cancer and advanced gastric cancer
(32). Similar to gastric cancer,
the SLIT1 promoter region is methylated in glioma tumor cells
(48). Another study on glioblastoma
demonstrated that SLIT1 improves the As2O3 resistance of
glioblastoma cells (31). Based on
previous findings (32,33), it was speculated that SLIT3 may not
only be downregulated but also be a tumor suppressor in
glioblastoma. In the present study, microarray and RT-qPCR analyses
proved that SLIT1 expression was downregulated in glioblastoma.
Based on the results of the cytological experiments, overexpression
of SLIT1 inhibited cell viability, proliferation and adhesion, and
promoted apoptosis of glioblastoma cells. Additionally, SLIT1 was
identified as a target gene of miR-640, suggesting that
overexpression of SLIT1 may attenuate the promotive effect of
miR-640 on glioblastoma. Thus, the present study revealed that
miR-640 may act as a tumor promoter in glioblastoma cells by
targeting SLIT1.
The present study is not without limitations as it
fails to explain the potential signaling pathways affected by SLIT1
in the development of glioblastoma. Thus, prospective studies will
focus on investigating the effect of miR-640 on the malignant
behavior of tumors in animals.
In conclusion, the results of the present study
demonstrated that miR-640 contributed to the progression of
glioblastoma cells by directly targeting SLIT1. Taken together,
these results suggest that the miR-640/SLIT1 axis may have the
potential to provide novel targets for the treatment and diagnosis
of glioblastoma.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
CL performed the experiments and the data analysis.
ZL and SD conceived and designed the study. YC and XC contributed
to the acquisition of the data. NL, ZL and JC contributed to the
analysis and interpretation of data. JC and SD wrote the
manuscript. NL, XC and YC reviewed and edited the manuscript. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Puai Hospital (approval no. KY-2019-04702; Wuhan,
China) and written informed consent was provided by all patients
prior to the study start.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Korja M, Raj R, Seppä K, Luostarinen T,
Malila N, Seppälä M, Mäenpää H and Pitkäniemi J: Glioblastoma
survival is improving despite increasing incidence rates: A
nationwide study between 2000 and 2013 in Finland. Neuro Oncol.
21:370–379. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Alexander BM and Cloughesy TF: Adult
glioblastoma. J Clin Oncol. 35:2402–2409. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kim H, Leiby BE and Shi W: Too little, too
soon: Short-course radiotherapy in elderly patients with
glioblastoma. J Clin Oncol. 34:2191–2192. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cloughesy TF, Mochizuki AY, Orpilla JR,
Hugo W, Lee AH, Davidson TB, Wang AC, Ellingson BM, Rytlewski JA,
Sanders CM, et al: Neoadjuvant anti-PD-1 immunotherapy promotes a
survival benefit with intratumoral and systemic immune responses in
recurrent glioblastoma. Nat Med. 25:477–486. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sukumar UK, Bose RJC, Malhotra M, Babikir
HA, Afjei R, Robinson E, Zeng Y, Chang E, Habte F, Sinclair R, et
al: Intranasal delivery of targeted polyfunctional gold-iron oxide
nanoparticles loaded with therapeutic microRNAs for combined
theranostic multimodality imaging and presensitization of
glioblastoma to temozolomide. Biomaterials. 218:1193422019.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ho KH, Cheng CH, Chou CM, Chen PH, Liu AJ,
Lin CW, Shih CM and Chen KC: miR-140 targeting CTSB signaling
suppresses the mesenchymal transition and enhances temozolomide
cytotoxicity in glioblastoma multiforme. Pharmacol Res.
147:1043902019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cloughesy T, Finocchiaro G, Belda-Iniesta
C, Recht L, Brandes AA, Pineda E, Mikkelsen T, Chinot OL, Balana C,
Macdonald DR, et al: Randomized, double-blind, placebo-controlled,
multicenter phase II study of onartuzumab plus bevacizumab versus
placebo plus bevacizumab in patients with recurrent glioblastoma:
Efficacy, safety, and hepatocyte growth factor and
O(6)-Methylguanine-DNA methyltransferase biomarker analyses. J Clin
Oncol. 35:343–351. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Goswami S, Walle T, Cornish AE, Basu S,
Anandhan S, Fernandez I, Vence L, Blando J, Zhao H, Yadav SS, et
al: Immune profiling of human tumors identifies CD73 as a
combinatorial target in glioblastoma. Nat Med. 26:39–46. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lee RC, Feinbaum RL and Ambros V: The
C. elegans heterochronic gene lin-4 encodes small RNAs with
antisense complementarity to lin-14. Cell. 75:843–854. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Citron F, Segatto I, Vinciguerra GLR,
Musco L, Russo F, Mungo G, D'Andrea S, Mattevi MC, Perin T,
Schiappacassi M, et al: Downregulation of miR-223 expression is an
early event during mammary transformation and confers resistance to
CDK4/6 inhibitors in luminal breast cancer. Cancer Res.
80:1064–1077. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Carotenuto P, Hedayat S, Fassan M,
Cardinale V, Lampis A, Guzzardo V, Vicentini C, Scarpa A, Cascione
L, Costantini D, et al: Modulation of biliary cancer
chemo-resistance through microRNA-mediated rewiring of the
expansion of CD133+ cells. Hepatology. 72:982–996. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Huang T, Wan X, Alvarez AA, James CD, Song
X, Yang Y, Sastry N, Nakano I, Sulman EP, Hu B and Cheng SY: MIR93
(microRNA-93) regulates tumorigenicity and therapy response of
glioblastoma by targeting autophagy. Autophagy. 15:1100–1111. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Han M, Wang S, Fritah S, Wang X, Zhou W,
Yang N, Ni S, Huang B, Chen A, Li G, et al: Interfering with long
non-coding RNA MIR22HG processing inhibits glioblastoma progression
through suppression of Wnt/β-catenin signalling. Brain.
143:512–530. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yang CH, Wang Y, Sims M, Cai C and Pfeffer
LM: MicroRNA-1 suppresses glioblastoma in preclinical models by
targeting fibronectin. Cancer Lett. 465:59–67. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jiang J, Wang X and Lu J: PWRN1 suppressed
cancer cell proliferation and migration in glioblastoma by
inversely regulating hsa-miR-21-5p. Cancer Manag Res. 12:5313–5322.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hu X, Yan P, Feng J and Zhang F:
Expression of microRNA-210 and the prognosis in glioma patients: A
meta-analysis. Biomark Med. 14:795–805. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xia W, Zhu J, Tang Y, Wang X, Wei X, Zheng
X, Hou M and Li S: PD-L1 inhibitor regulates the miR-33a-5p/PTEN
signaling pathway and can be targeted to sensitize glioblastomas to
radiation. Front Oncol. 10:8212020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen Q, Gao J, Zhao Y and Hou R: Long
non-coding RNA LBX2-AS1 enhances glioma proliferation through
downregulating microRNA-491-5p. Cancer Cell Int. 20:4112020.
View Article : Google Scholar
|
|
20
|
Pan CM, Chan KH, Chen CH, Jan CI, Liu MC,
Lin CM, Cho DY, Tsai WC, Chu YT, Cheng CH, et al: MicroRNA-7
targets T-Box2 to inhibit epithelial-mesenchymal transition and
invasiveness in glioblastoma multiforme. Cancer Lett. 493:133–142.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lin H, Zuo D, He J, Ji T, Wang J and Jiang
T: Long noncoding RNA WEE2-AS1 plays an oncogenic role in
glioblastoma by functioning as a molecular sponge for
microRNA-520f-3p. Oncol Res. Aug 24–2020.doi:
10.3727/096504020X15982623243955 (Epub ahead of print). View Article : Google Scholar
|
|
22
|
Harel S, Sanchez-Gonzalez V, Echavarria R,
Mayaki D and Hussain SN: Roles of miR-640 and Zinc finger protein
91 (ZFP91) in angiopoietin-1-induced in vitro angiogenesis. Cells.
9:16022020. View Article : Google Scholar
|
|
23
|
Li X, Lu Y, Chen Y, Lu W and Xie X:
MicroRNA profile of paclitaxel-resistant serous ovarian carcinoma
based on formalin-fixed paraffin-embedded samples. BMC Cancer.
13:2162013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhai Z, Fu Q, Liu C, Zhang X, Jia P, Xia
P, Liu P, Liao S, Qin T and Zhang H: Emerging roles of
hsa-circ-0046600 targeting The miR-640/HIF-1α signalling pathway in
the progression of HCC. Onco Targets Ther. 12:9291–9302. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lodes MJ, Caraballo M, Suciu D, Munro S,
Kumar A and Anderson B: Detection of cancer with serum miRNAs on an
oligonucleotide microarray. PLoS One. 4:e62292009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tong M, Jun T, Nie Y, Hao J and Fan D: The
role of the Slit/Robo signaling pathway. J Cancer. 10:2694–2705.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Blockus H and Chedotal A: Slit-Robo
signaling. Development. 143:3037–3044. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ma L and Tessier-Lavigne M: Dual
branch-promoting and branch-repelling actions of Slit/Robo
signaling on peripheral and central branches of developing sensory
axons. J Neurosci. 27:6843–6851. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Marlow R, Strickland P, Lee JS, Wu X,
Pebenito M, Binnewies M, Le EK, Moran A, Macias H, Cardiff RD, et
al: SLITs suppress tumor growth in vivo by silencing Sdf1/Cxcr4
within breast epithelium. Cancer Res. 68:7819–7827. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Jaworski A, Tom I, Tong RK, Gildea HK,
Koch AW, Gonzalez LC and Tessier-Lavigne M: Operational redundancy
in axon guidance through the multifunctional receptor Robo3 and its
ligand NELL2. Science. 350:961–965. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shuai W, Wu J, Chen S, Liu R, Ye Z, Kuang
C, Fu X, Wang G, Li Y, Peng Q, et al: SUV39H2 promotes colorectal
cancer proliferation and metastasis via tri-methylation of the
SLIT1 promoter. Cancer Lett. 422:56–69. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kim M, Kim JH, Baek SJ, Kim SY and Kim YS:
Specific expression and methylation of SLIT1, SLIT2, SLIT3, and
miR-218 in gastric cancer subtypes. Int J Oncol. 48:2497–2507.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Amodeo V, A D, Betts J, Bartesaghi S,
Zhang Y, Richard-Londt A, Ellis M, Roshani R, Vouri M, Galavotti S,
et al: A PML/Slit axis controls physiological cell migration and
cancer invasion in the CNS. Cell Rep. 20:411–426. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Long H, Liang C, Zhang X, Fang L, Wang G,
Qi S, Huo H and Song Y: Prediction and analysis of key genes in
glioblastoma based on bioinformatics. Biomed Res Int.
2017:76531012017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sciuscio D, Diserens AC, van Dommelen K,
Martinet D, Jones G, Janzer RC, Pollo C, Hamou MF, Kaina B, Stupp
R, et al: Extent and patterns of MGMT promoter methylation in
glioblastoma- and respective glioblastoma-derived spheres. Clin
Cancer Res. 17:255–266. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ritchie ME, Phipson B, Wu D, Hu Y, Law CW,
Shi W and Smyth GK: Limma powers differential expression analyses
for RNA-sequencing and microarray studies. Nucleic Acids Res.
43:e472015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Porter AG and Jänicke RU: Emerging roles
of caspase-3 in apoptosis. Cell Death Differ. 6:99–104. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Pomeroy SL: Neural development and the
ontogeny of central nervous system tumors. Neuron Glia Biol.
1:127–133. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Nawaz Z, Patil V, Thinagararjan S, Rao SA,
Hegde AS, Arivazhagan A, Santosh V and Somasundaram K: Impact of
somatic copy number alterations on the glioblastoma miRNome:
miR-4484 is a genomically deleted tumour suppressor. Mol Oncol.
11:927–944. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Piao XY, Li W, Li Z, Zhang N, Fang H,
Zahid D and Qu Q: Forced FoxO1:S249V expression
suppressed glioma cell proliferation through G2/M cell cycle
arrests and increased apoptosis. Neurol Res. 41:189–198. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Liang HX, Sun LB and Liu NJ: Neferine
inhibits proliferation, migration and invasion of U251 glioma cells
by down-regulation of miR-10b. Biomed Pharmacother. 109:1032–1040.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Russo MA, Paolillo M, Sanchez-Hernandez Y,
Curti D, Ciusani E, Serra M, Colombo L and Schinelli S: A
small-molecule RGD-integrin antagonist inhibits cell adhesion, cell
migration and induces anoikis in glioblastoma cells. Int J Oncol.
42:83–92. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Li Y, Cai T, Zhang W, Zhu W and Lv S:
Effects of Saikosaponin D on apoptosis in human U87 glioblastoma
cells. Mol Med Rep. 16:1459–1464. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Mazumder S, Plesca D and Almasan A:
Caspase-3 activation is a critical determinant of genotoxic
stress-induced apoptosis. Methods Mol Biol. 414:13–21.
2008.PubMed/NCBI
|
|
46
|
Leyva-Diaz E, del Toro D, Menal MJ,
Cambray S, Susín R, Tessier-Lavigne M, Klein R, Egea J and
López-Bendito G: FLRT3 is a Robo1-interacting protein that
determines Netrin-1 attraction in developing axons. Curr Biol.
24:494–508. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Kaneko N, Herranz-Pérez V, Otsuka T, Sano
H, Ohno N, Omata T, Nguyen HB, Thai TQ, Nambu A, Kawaguchi Y, et
al: New neurons use Slit-Robo signaling to migrate through the
glial meshwork and approach a lesion for functional regeneration.
Sci Adv. 4:eaav06182018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Dickinson RE, Dallol A, Bieche I, Krex D,
Morton D, Maher ER and Latif F: Epigenetic inactivation of SLIT3
and SLIT1 genes in human cancers. Br J Cancer. 91:2071–2078. 2004.
View Article : Google Scholar : PubMed/NCBI
|