Introduction
Ovarian cancer is the second leading cause of female
reproductive cancer death. In 2018, ovarian cancer affected
approximately 295,414 women worldwide, with a predilection for
postmenopausal women (1). Ovarian
cancer carries a poor prognosis, with a 5-year survival rate of
only 30% and deaths occurring within just 2 years of diagnosis.
This low survival rate can be attributed to the late diagnosis of
the disease. A significant proportion of women are diagnosed with
ovarian cancer at advanced stages, and despite initial complete
responses to chemotherapy, 70% of patients develop recurrence
within 18 months (2). Current
treatment methods for ovarian cancer are surgery and cytotoxic
chemotherapy. When complete surgical debulking seems difficult for
the patient, pre- or postoperative chemotherapy may be used as an
additional therapeutic strategy. For the past 40 years, cytotoxic
chemotherapy based on platinum- and taxane- group drugs such as
carboplatin, cisplatin, and paclitaxel (PTX), have been used for
first-line chemotherapy. However, despite several clinical trials,
regimens containing platinum have not shown improvements for the
past 20 years largely due to drug toxicity and resistance. In fact,
of the 80% of patients with good initial responses to standard
treatments, 70% experience disease recurrence (1). In addition to the problem of
drug-resistance, drug-refractory patients exhibit the worst
prognosis for treatment (2).
Patients whose disease progress despite drug treatment have been
managed with pegylated doxorubicin or topotecan, both of which have
low response rates. Therefore, a novel strategy identified through
molecular characterization is required to overcome drug-resistance
and to offer new treatment options for patients with advanced and
relapsed ovarian cancer.
PTX is a taxane-based, microtubule-stabilizing drug
that has been approved by the Food and Drug Administration (FDA)
for the treatment of various types of solid cancers, including
those of the breast, lung, and ovary (3). PTX inhibits normal dissociation of α-
and β-tubulin subunits, thereby increasing the percentage of
tubulin subunits that polymerize into microtubules (4). As a result, PTX reduces the
concentration of free-tubulin subunits necessary for microtubule
assembly, causing the mitotic arrest of cancer cells. Despite the
efficacy of PTX in killing cancer cells, resistance remains a major
obstacle to overcome. There are numerous molecular mechanisms by
which cancers evade the cytotoxic effect of PTX. These mechanisms
include, but are not limited to, altering microtubule dynamics and
changing tubulin subtypes, compositions, and posttranslational
modifications (5).
Histone deacetylase 6 (HDAC6) is a special enzyme
with a unique structure and various functions. It possesses two
deacetylase domains and is involved in various
deacetylase-dependent and -independent cytoplasmic cellular
processes, such as inflammation, angiogenesis, transcription,
protein degradation, cell survival, and cell motility (6–8). HDAC6
targets various substrates, including α-tubulin, cortactin, HSP90,
Tat, Ku70, β-catenin, peroxiredoxins, and survivin (8). The overexpression of HDAC6 induces
tumorigenesis in mammalian cells by hypoacetylating α-tubulin and
promoting the formation of immune synapses, cell migration, and
chemotaxis. As a result, extensive studies have been conducted to
develop HDAC6-selective inhibitors to target tumorigenesis and
metastasis. One of the HDAC6-specific inhibitors, ACY-1215
(ricolinostat), is under a phase1b clinical trial for relapsed or
refractory multiple myeloma (9).
Moreover, HDAC6 inhibition through ACY-1215 was proven to be
effective in various cancers, including inflammatory and metastatic
breast cancers, melanoma, glioblastoma, prostate cancer, colorectal
cancer, multiple myeloma, and lymphoma (9–16).
ACY-241 or citarinostat, like ACY-1215, inhibits HDAC6 with 13 to
18-fold reduced potency against HDAC1, 2, and 3 isozymes (17). However, as a second generation
inhibitor, ACY-241 carries improved solubility compared to its
structurally related inhibitor ACY-1215 and is orally available.
Currently, it is under investigation in a clinical trial
(NCT02886006) for multiple myeloma. Apart from ACY-1215 and
ACY-241, A452 is a potent hydroxamic acid based HDAC6-selective
small-molecule inhibitor under preclinical studies (18,19).
A452 carries a γ-lactam core that selectively inhibits HDAC6
catalytic activity and successfully suppresses colorectal cancer
cell growth. However, further research is required to evaluate its
efficacy and mechanisms in other cancers, such as in epithelial
ovarian carcinomas (EOC).
Approximately 85–90% of malignant ovarian cancer
subtypes are EOCs. They are classified histologically into four
types: high- and low-grade serous carcinomas account for 75% of
EOCs, whereas clear cell carcinomas, mucinous carcinomas,
endometrioid carcinomas comprise 9, 3 and 8% of EOCs, respectively.
EOCs are known to carry mutually exclusive ARID1A and
TP53 mutations (20).
ARID1A, a component of the SWI/SNF complex, is mutated in
more than 50% of ovarian clear cell carcinomas and 30% of ovarian
endometrioid carcinomas. These ARID1A-mutated ovarian
cancers fail to inhibit HDAC6 activity, thus destabilizing p53 by
removing its lysine 120 acetylation (K120Ac) (21). This increased HDAC6 activity promotes
tumor growth. Therefore, using a selective HDAC6 inhibitor, such as
ACY-1215, in ARID1A-inactivated ovarian cancer cells induces
apoptosis by modulating p53 stability.
In this study, we conducted further experiments
using two different HDAC6-selective inhibitors, ACY-241 and A452,
to confirm the anticancer effect of HDAC6 inhibition on ARID1A-null
ovarian cancer TOV-21G cells with a wild type TP53. Furthermore, as
both HDAC6 and PTX alter cellular α-tubulin dynamics, the
combination treatment of PTX and HDAC6 inhibitors synergistically
enhances apoptosis in TOV-21G cancer cells. This may be a novel
therapeutic strategy in overcoming the limitations of PTX.
Materials and methods
Ovarian cancer cell lines and
culture
The human ovarian clear cell carcinoma cell,
TOV-21G, was purchased from American Type Culture Collection
(ATCC). It was cultured in Dulbecco's modified Eagle's medium
(Sigma-Aldrich; Merck KGaA) containing 10% fetal bovine serum
(HyClone; GE Healthcare), 100 U/ml penicillin and 100 µg/ml
streptomycin (Gibco; Thermo Fisher Scientific, Inc.) in a 5%
CO2 and 37°C incubator.
Cell growth and viability assay
As described elsewhere (14), to measure cell growth and viability,
the cells were seeded in triplicates at a density of
3×103 cells in 130 µl of medium in 96-well plates.
Following 24 h of incubation, drugs were added to each well, then
incubated for 24, 48 and 72 h each. Following the drug treatment,
13 µl of CCK-8 reagent (WST-8; CCK-8 kit, CK04; Dojindo Molecular
Technologies, Inc.) was added into each well, and the plate was
incubated at 37°C for 3 h. Then, absorbance was read at 450 nm
using microplate reader (Tecan Group, Ltd.). The results are
presented as percentages relative to the control wells. All
half-maximal growth inhibition concentration (GI50) and
half-maximal inhibitory concentration (IC50) values were
calculated using GraphPad Prism ver. 7.0 software.
Colony formation assay
To examine colony forming ability, the cells were
seeded at 6-well plates at a density of 1×103 cells in 2
ml of medium. Following 24 h of incubation, drugs were treated in
each wells, and the plate was incubated at 37°C for 14 days. The
medium with drugs were renewed every 3 to 4 days. The cells were
stained with 0.7 ml 0.05% crystal violet solution (Sigma-Aldrich;
Merck KGaA) for 30 min and the colonies were counted using
open-source software OpenCFU (22).
The experiment was carried out in triplicates. One-way ANOVA with
Bonferroni's post-hoc test was carried out using GraphPad Prism 7.0
software (GraphPad Software).
Drug combination analysis
To determine drug combination effects, constant
ratio of PTX to ACY-241 and PTX to A452 were used. Cells were
seeded in triplicates in 96-well plates at the density of
3×103 cells. The plate was incubated overnight before
adding drugs of diluted concentrations prior to use. The cells were
incubated with the drugs for 72 h at 37°C. A CCK-8 assay was
performed to determine cell viability. Drug synergy was determined
according to the combination index (CI) method described by Chou
(23); CI<1 implies synergism,
CI=1 implies additivity, CI>1 implies antagonism. CI values were
calculated using CalcuSyn software version 2.11 (Biosoft). Two-way
ANOVA with Bonferroni's post-hoc test for both combination was
conducted using GraphPad Prism 7.0 software (GraphPad
Software).
Wound healing assay
To determine wound closure capability, the cells
were seeded in 6-well plates at density of 1×106 cells
in triplicates. The cells were incubated in DMEM culture medium
containing 10% fetal bovine serum for 24 h. After 24-h incubation
at 37°C, the wound was scratched using 1,000 pipet tip. Then, the
cells were washed to remove cell debris and were treated with drugs
for 24 h. The cells were removed from the incubator and were washed
with 1 ml of medium and photographed. The wound was measured using
ImageJ software. One-way ANOVA with Bonferroni's post-hoc test was
carried out using GraphPad Prism 7.0 software (GraphPad
Software).
Western blot analysis
TOV-21G cells were seeded in 6-well plates at
density of 5×105 cells/well and incubated for overnight
in 37°C before drug treatment. After drug treatment, cells were
incubated for 24 h. Cells were lysed and extracted with 100 µl
NP-40 lysis buffer [0.5% NP-40, 50 mM Tris-HCl (pH 7.4), 120 mM
NaCl, 25 mM NaF, 25 mM glycerol phosphate, 1 mM EDTA, 5 mM EGTA, 1
mM PMSF and 1 mM bezamidine] and sonicated twice for 3 sec at 20%
amplitude. Bradford protein assay was conducted to measure protein
concentrations. Cell lysates containing 60–80 µg proteins were
loaded onto 7.5–12% SDS-PAGE. Then, proteins were transferred to
nitrocellulose membrane at 100 V for 100 min. Membranes were
blocked with 5% skim milk at room temperature and were incubated
for overnight with primary antibodies at 4°C. Primary antibodies
were diluted using following concentrations: α-tubulin (1:1,000),
acetyl-α-tubulin (1:2000), GAPDH (1:10,000), PARP (1:1,000),
Pro-caspase-3 (1:1,000), cleaved caspase-3 (1:500), Pro-caspase-8
(1:500), Pro-caspase-9 (1:500), Bak (1:1,000), Bax (1:500), Bcl-xL
(1:1,000), Bcl-2 (1:500), p53 (1:1,000), p53K120Ac (1:500),
p53K381Ac (1:1,000), and p-p53ser15 (1:500). Then membranes were
washed with 0.1% Tween-20/PBS and incubated with anti-rabbit/mouse
secondary antibody coupled to HRP at room temperature for 3 h.
Protein bands were detected with ECL western blotting solution
(Thermo Scientific Pierce).
Apoptosis assay
Apoptosis assay was performed with Annexin
V/propidium iodide (PI) double staining. Initially, cells were
seeded in 100 Φ plates at densities of
1.2×106−1.5×106 cells. Cells were incubated
for overnight in 37°C and treated with drugs accordingly. Drug
treated cells were incubated for 48 h, washed with ice-cold PBS and
detached with trypsin-EDTA. Cells were collected after
centrifugation at 1,000 rpm for 3 min, then resuspended with
400–700 µl binding buffer. Cells were stained with 5 µl PI and 0.7
µl Annexin V-fluorescein isothiocyanate (FITC) in dark for 15 min.
The cells were diluted with 400 µl of 1X binding buffer (Annexin
V-FITC Apoptosis Detection Kit; BD556547; BD Pharmingen). Finally,
the stained cells were analyzed using a flow cytometer and BD
FACSDiva software version 7 (both from BD Biosciences). One-way
ANOVA with Bonferroni's post-hoc test was carried out using
GraphPad Prism 7.0 software (GraphPad Software).
Statistical analysis
All data in this research are presented as means ±
standard deviation of or more than three independent experiments.
One-way or two-way ANOVA followed by a Bonferroni's multiple
comparison post-hoc analysis was conducted using GraphPad Prism
software 7.0 (GraphPad Software). P-value <0.05 was considered
statistically significant for all data presented.
Results
PTX and the two HDAC inhibitors,
ACY-241 and A452, reduce cell growth and cell viability
PTX is a taxane-based chemotherapeutic drug known to
inhibit the cell growth of various tumors (3). Similarly, ACY-241 and A452, both
HDAC6-selective inhibitors, are known to suppress cell growth and
decrease cell viability in solid tumors (17), multiple myeloma (24,25) and
colorectal cancer (18,19). To examine the anticancer effects of
PTX and the two HDAC6 inhibitors in ovarian cancer cells, we
measured the cell growth and viability of TOV-21G cells treated
with PTX, ACY-241 and A452. PTX effectively reduced the cell growth
and viability of TOV-21G in both a time- and dose- dependent manner
(Fig. 1A and B). When the
GI50 and IC50 were calculated, PTX inhibited
cell growth at 28.74 nM and reduced cell viability at 33.61 nM
(Table I). On the other hand,
compared to PTX, the HDAC6 inhibitor ACY-241 inhibited cell
viability and growth at higher concentrations with a
GI50 value of 8.89 µM and an IC50 of 9.13 µM
(Fig. 1C and D). Compared to
ACY-241, A452 had 12 times lower GI50 and 11 times lower
IC50 in TOV-21G (Fig. 1E and
F). The GI50 value of A452 was 0.69 µM
IC50 value of A452 was 0.8 µM. PTX was the most
effective drug among the three drugs, and A452 was more effective
in reducing cell viability than ACY-241.
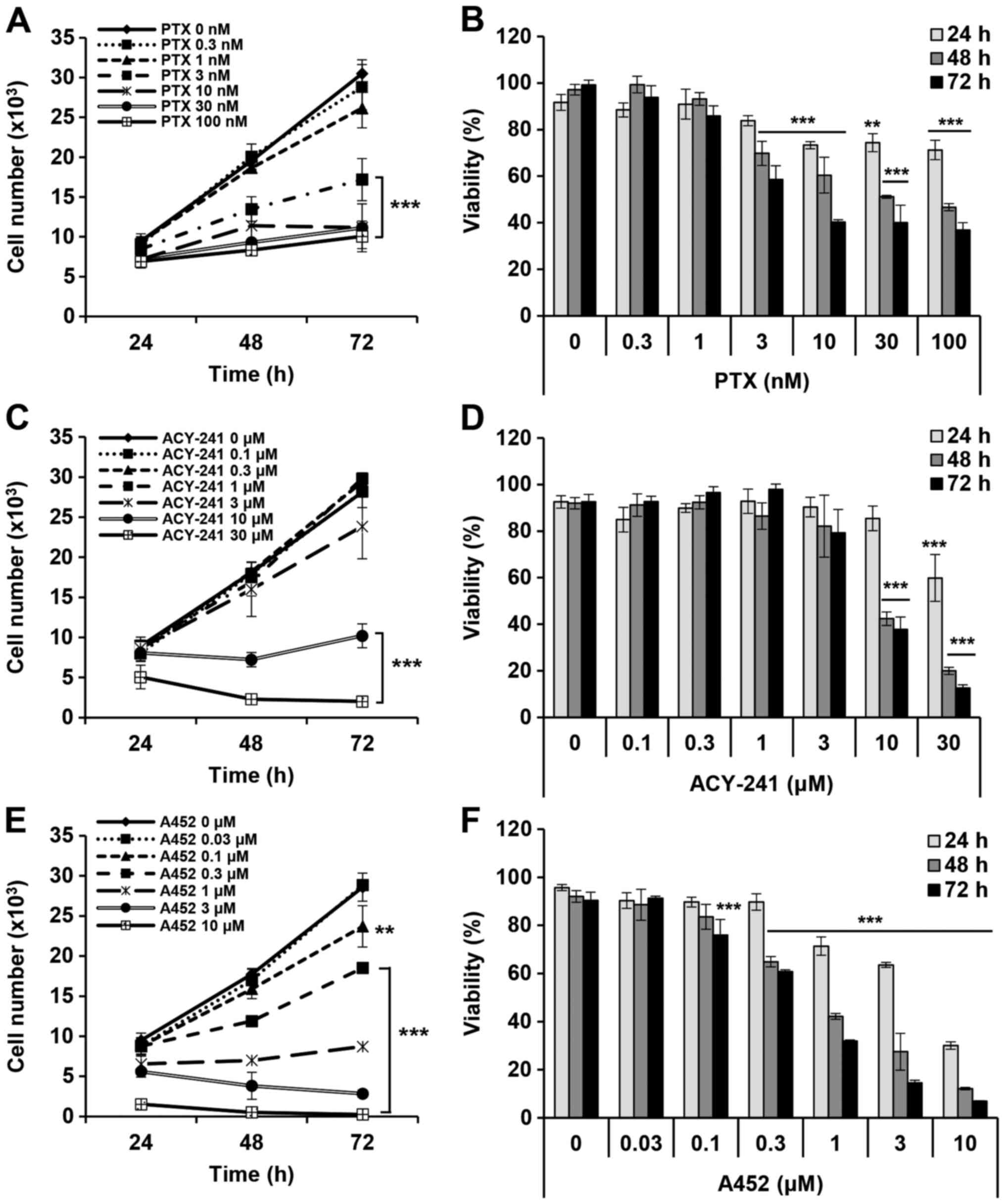 | Figure 1.PTX, ACY-241 and A452 individually
suppress the growth and viability of the AT-rich interaction domain
1A-null, p53-wild-type TOV-21G ovarian cancer cell line. (A and B)
PTX (0, 0.1, 1, 3, 10, 30 and 100 nM), (C and D) ACY-241 (0, 0.1,
0.3, 1, 3, 10 and 30 µM) and (E and F) A452 (0, 0.1, 0.3, 1, 3, 10
and 30 µM) were used for treatment of TOV-21G cells for 24, 48 and
72 h at the respective concentrations. (A, C and E) Cell growth and
(B, D and F) cell viability were measured using Cell Counting Kit-8
assays in 96-well plates. All three drugs inhibited the growth and
decreased the viability of TOV-21G cells. Cell counts were
estimated indirectly from a standard curve derived from absorbance
of known cell numbers. The absorbance at each concentration was
normalized to that of the negative control within the corresponding
time interval. Data are presented as the mean ± SD (n=3).
**P<0.01 and ***P<0.001 vs. the DMSO control (one-way ANOVA
with Bonferroni's post hoc test; the significance levels apply to
all samples under a line). PTX, paclitaxel. |
 | Table I.Drug inhibition concentrations of
TOV-21G cells treated with PTX and histone deacetylase 6-selective
inhibitors. |
Table I.
Drug inhibition concentrations of
TOV-21G cells treated with PTX and histone deacetylase 6-selective
inhibitors.
|
| Drug (48 h) |
|---|
|
|
|
|---|
| Variable | PTX, nM | ACY-241, µM | A452, µM |
|---|
|
GI50 | 28.74 | 8.89 | 0.69 |
|
IC50 | 33.61 | 9.13 | 0.80 |
Combination treatment of PTX and
HDAC6-selective inhibitors synergistically decrease cell viability
and inhibit cell growth in ovarian cancer cells
Next, we employed the combination therapy of PTX and
ACY-241 or PTX and A452 on TOV-21G and analyzed its effects on cell
viability and cell growth using Cell Counting Kit (CCK)-8 assays.
Synergy was evaluated by calculating CI values based on the Chou
and Talalay method (23). The
combination treatment of PTX and ACY-241 at a dose ratio of 1:1,000
resulted in CI values <1, with PTX and ACY-241 concentrations of
3, 4, 5 and 6 nM and 3, 4, 5 and 6 µM, respectively (Fig. 2A). On the other hand, the combination
treatment of PTX and A452 at a dose ratio of 1:400 resulted in CI
values less than 1 from concentrations of 0.25 nM PTX and 0.1 µM
A452 up to 8 nM PTX and 3.2 µM A452 (Fig. 2B). As single treatment of A452 is
more effective than ACY-241 in inhibiting cell growth and reducing
cell viability in TOV-21G, PTX synergized with A452 at
substantially lower doses than it did with ACY-241. The long-term
combination treatment of PTX and both HDAC6 inhibitors also
significantly inhibited the colony-forming abilities of TOV-21G
(Fig. 2C and D). Moreover, as HDAC6
inhibition is known to suppress migration and metastasis in various
cancers (26,27), wound healing assays were conducted to
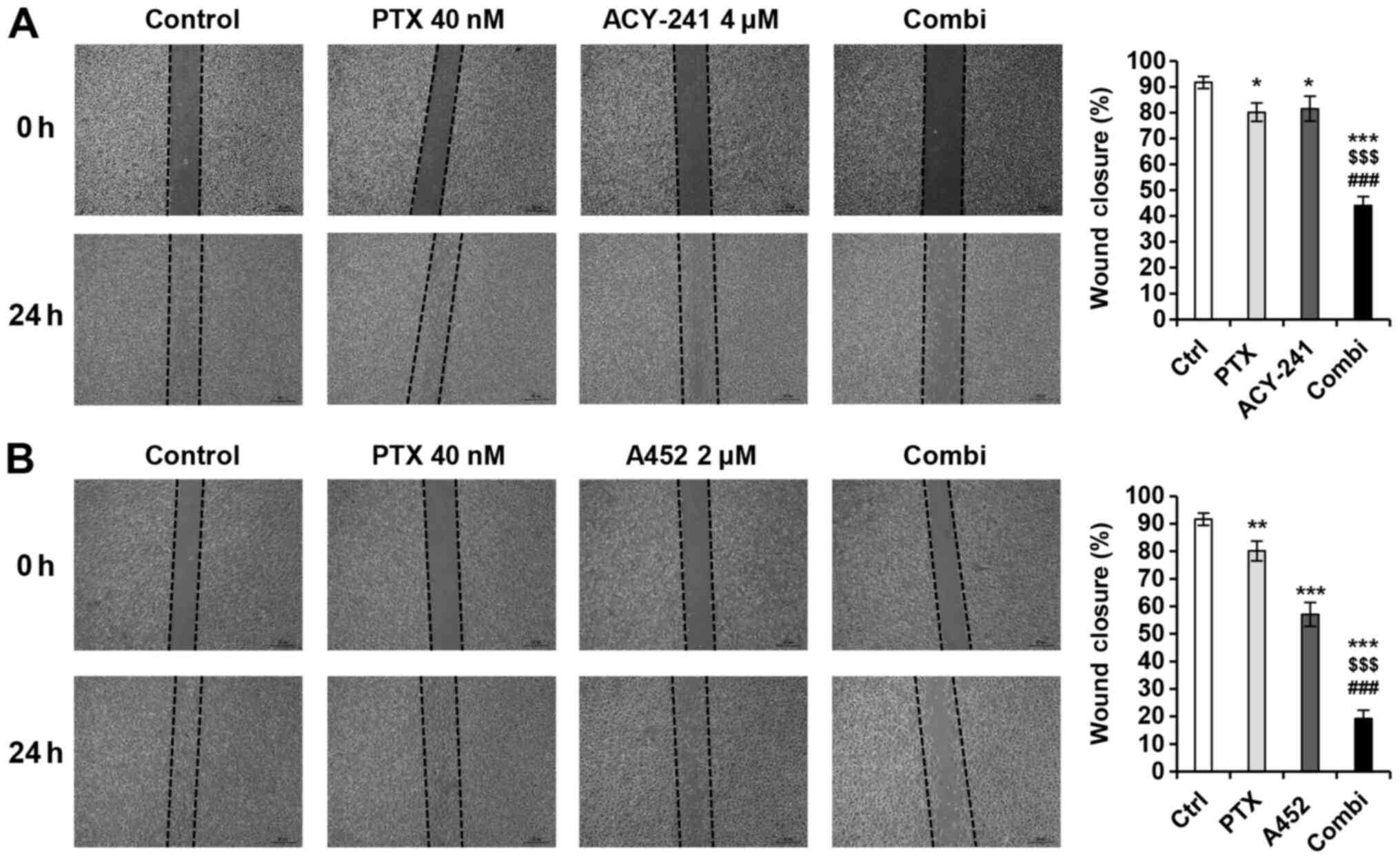
examine the effects of PTX and the two HDAC6-selective inhibitors
(Fig. 3A and B). Both drug
combinations synergistically suppressed TOV-21G migration and
proliferation.
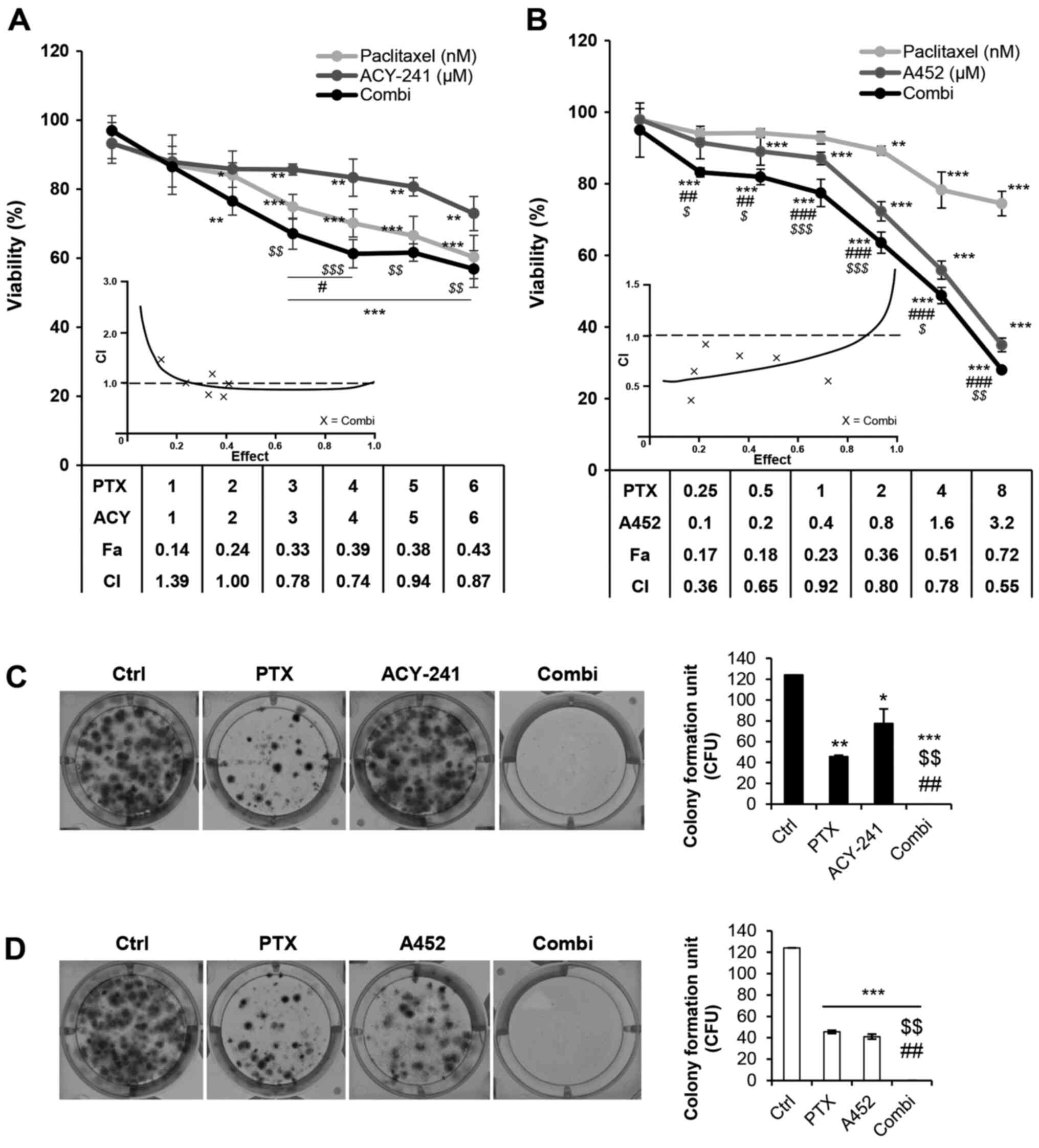 | Figure 2.PTX and HDAC6-selective inhibitors
synergistically decrease cell viability and proliferation in the
short and long term. (A) TOV-21G cells were treated with 0.1% DMSO
control, PTX, ACY-241 or a combination of these agents for 72 h.
(B) TOV-21G cells were treated with 0.1% DMSO control, PTX, A452 or
a combination of these agents for 72 h. PTX and both
HDAC6-selective inhibitors synergistically reduced cell viability.
(C) Combination treatment of PTX and ACY-241 synergistically
inhibited the colony formation of TOV-21G cells. (D) Combination
treatment of PTX and A452 synergistically inhibited the colony
formation of TOV-21G cells. The drug concentrations used for colony
formation assays were as follows: 0.1% DMSO, 4 nM PTX, 0.2 µM
ACY-241 and 0.1 µM A452. Data are presented as the mean ± SD (n=3).
*P<0.05, **P<0.01 and ***P<0.001 vs. DMSO control;
$P<0.05, $$P<0.01 and
$$$P<0.001 vs. PTX; #P<0.05,
##P<0.01 and ###P<0.001 vs. ACY-241 or
A452, using a two-way ANOVA test for (A) and (B), and one-way ANOVA
tests were used for (C) and (D). ACY, ACY-241; Combi, combination
treatment; Fa, fraction affected; CI, combination index; PTX,
paclitaxel; CFU, colony-forming unit; Ctrl, control; HDAC6, histone
deacetylase 6. |
PTX and HDAC6-selective inhibitors
synergistically result in apoptosis in ovarian cancer cells
To further confirm the synergy of PTX and
HDAC6-selective inhibitors, we performed a western blot analysis to
assess drug synergy at the protein level. Several apoptotic
proteins were activated on the combination treatment of PTX with
ACY-241 (Fig. 4A). Poly(ADP-ribose)
polymerase (PARP), which is involved in DNA repair, genomic
stability, and programmed cell death, was synergistically cleaved
on combination treatment, resulting in apoptosis. As a PARP
regulating protein, cleaved caspase-3 was also significantly
increased upon combination treatment. On the other hand, Bcl-2
family related pro-apoptotic proteins such as Bak and Bax did not
show significant changes in expression levels. However, Bcl-2
family-related anti-apoptotic proteins such as Bcl-xL and Bcl-2,
decreased synergistically after the combination treatment of PTX
and ACY-241, thus supporting the synergism between the two drugs.
Similarly, when PTX and A452 were treated together, both cleaved
PARP and cleaved caspase-3 increased synergistically (Fig. 4B). However, in this combination,
pro-apoptotic proteins Bak and Bax synergistically increased, and
anti-apoptotic proteins Bcl-xL and Bcl-2 synergistically decreased.
These results further suggest that the combination of PTX and A452
has greater synergy than that of PTX and ACY-241.
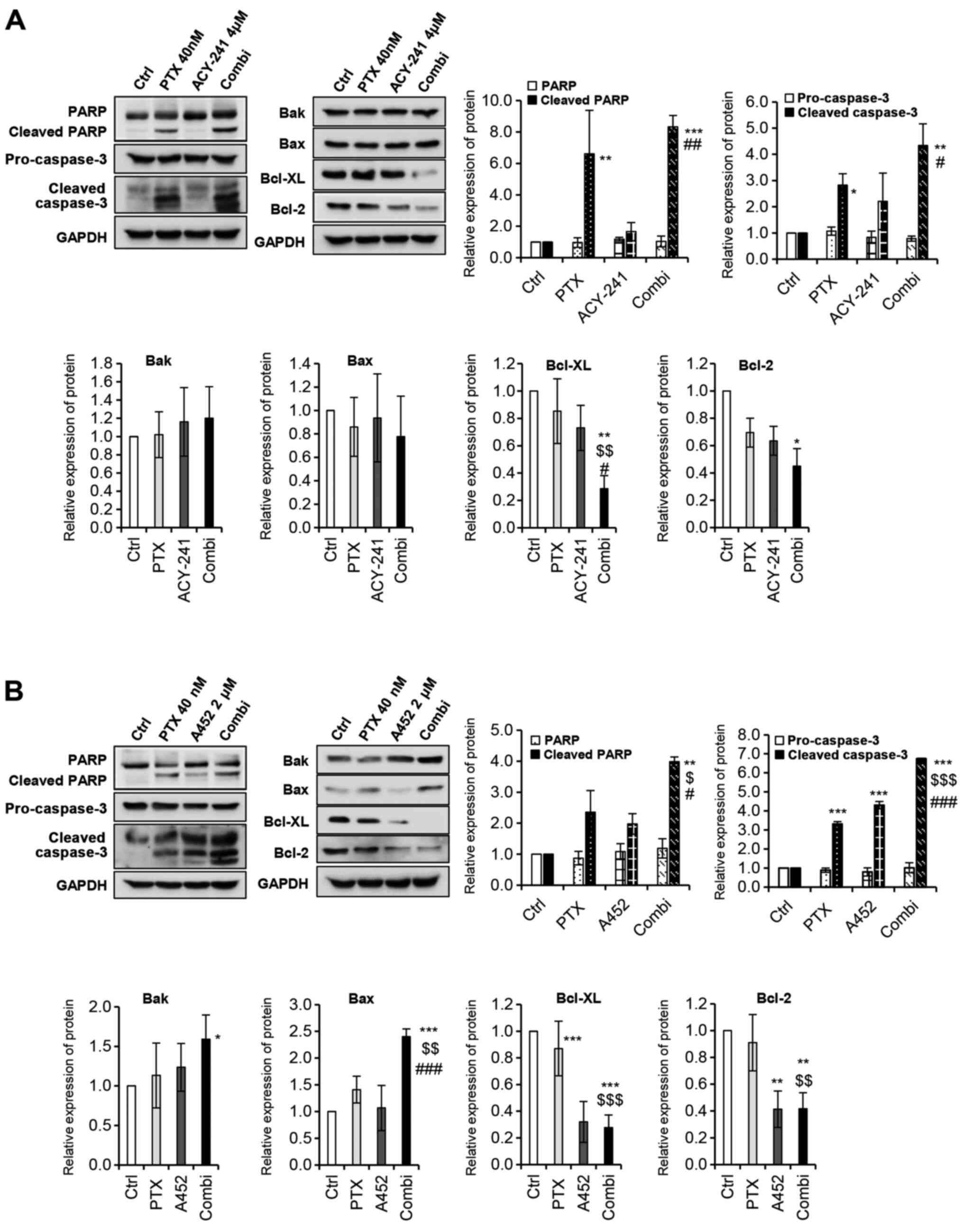 | Figure 4.PTX and histone deacetylase
6-selective inhibitors synergistically induce and reduce
pro-/anti-apoptotic markers. (A) Combination treatment of PTX (40
nM) and ACY-241 (4 µM) significantly increased pro-apoptotic
markers while decreasing anti-apoptotic markers. (B) Combination
treatment of PTX (40 nM) and A452 (2 µM) significantly induced
pro-apoptotic markers while reducing anti-apoptotic markers.
Western blot analysis was performed using the indicated antibodies.
The protein expression levels were semi-quantified relative to
α-tub or GAPDH. The levels in the 0.1% DMSO group were set as 1.
GAPDH was used as the loading control. Data are presented as the
mean ± SD (n=3). *P<0.05, **P<0.01 and ***P<0.001 vs. DMSO
control; $P<0.05, $$P<0.01 and
$$$P<0.001 vs. PTX; #P<0.05,
##P<0.01 and ###P<0.001 vs. ACY-241 or
A452 (one-way ANOVA). Bak, Bcl-2 homologous antagonist/killer; Bcl,
B-cell lymphoma; Bcl-xL, Bcl-extra large protein; Cas-3, Caspase-3;
Ctrl, control; Combi, combination treatment; PARP, poly(ADP-ribose)
polymerase; PTX, paclitaxel. |
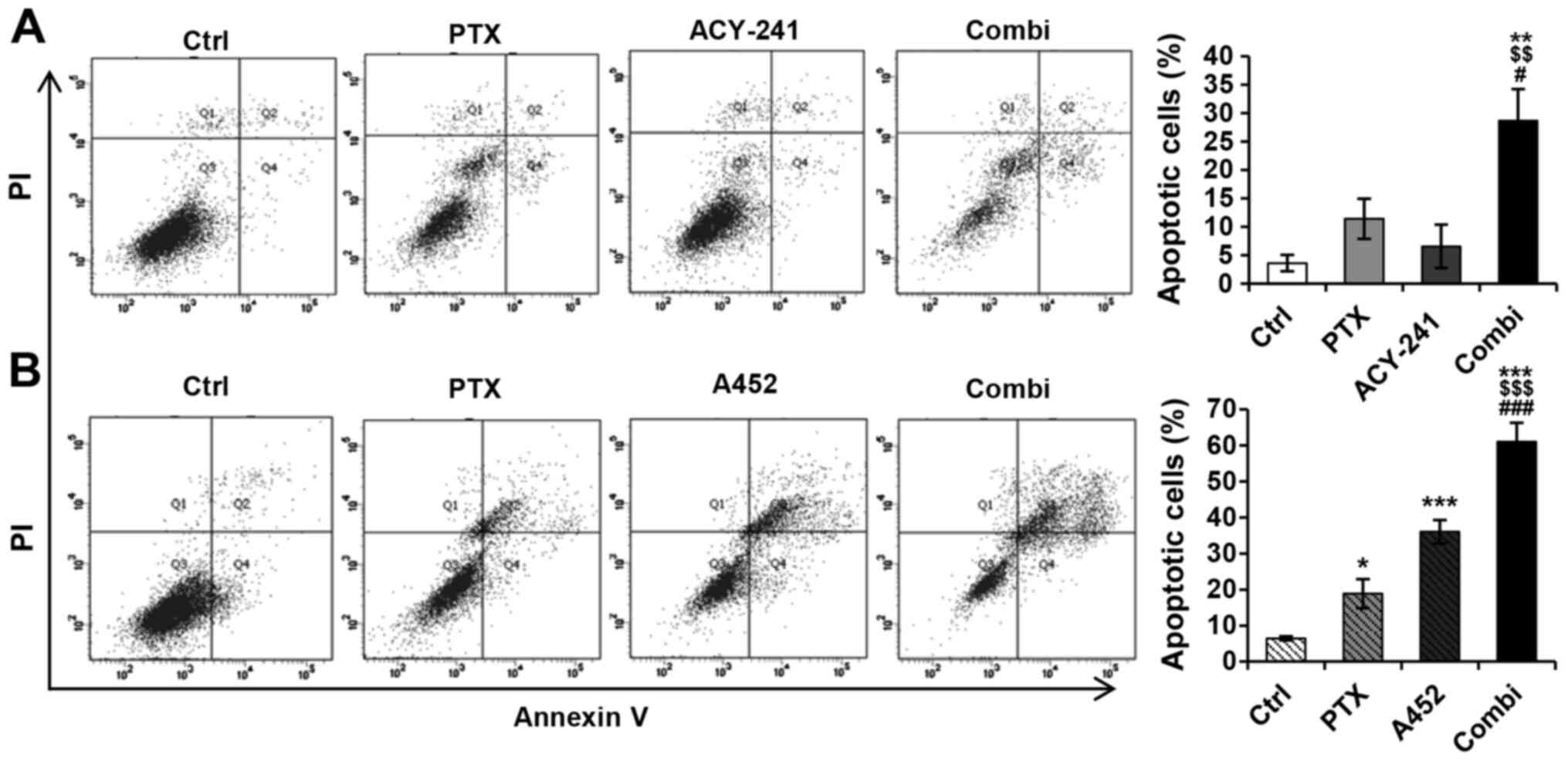
To further examine the apoptotic effect of the HDAC6
inhibitors and PTX, we performed an apoptosis assay using a flow
cytometry analysis system (Fig. 5A and
B). The percentage of apoptotic cells increased synergistically
in both drug combinations, as seen in the bar graph. The
combination treatment of PTX and ACY-241 increased the percentage
of apoptotic cells to 30%, whereas the control (dimethyl sulfoxide
[DMSO] 0.1%), PTX, and ACY-241 remained at 4, 11 and 7%,
respectively (Fig. 5A). Furthermore,
the combination treatment of PTX and A452 increased the percentage
of cell apoptosis to 61%. The percentages of cell apoptosis in the
control group (DMSO 0.1%) and in monotherapies of PTX and A452
remained at 6, 19 and 36%, respectively (Fig. 5B). Corresponding to the apoptotic
protein levels, A452 and PTX resulted in greater apoptosis than
ACY-241 and PTX. These findings indicate that the combination
treatment of PTX and HDAC6-selective inhibitors synergistically
induces apoptosis in ovarian cancer cells.
Combination treatment of PTX and
HDAC6-selective inhibitors increases p53 stability
synergistically
As HDAC6 inhibition is known to increase the lysine
acetylation of p53 (21,28), we performed a western blot analysis
to determine whether co-treatment of PTX and HDAC6 inhibitors would
affect p53 modification levels. Acetylated-α-tubulin was used as a
positive control to confirm the presence of PTX and HDAC6
inhibitors (Fig. 6A and B). The
single treatment of PTX did not increase p53 lysine (K120, K381)
acetylation levels in both combinations (Fig. 6A and B). However, co-treatment of PTX
and both HDAC6 inhibitors significantly increased acetylation,
compared to the individual monotherapy of the three drugs. Thus, it
shows that the combination treatment of PTX and ACY-241 or PTX and
A452 increases p53 stability synergistically, inducing the
apoptosis of ovarian cancer cells. Despite the increase in p53
stability, however, p53 levels did not correlate with the increase
in protein stability levels. Such trend can be explained by the
phosphorylation of p53 at serine 15, which seems to play a greater
role in determining protein levels of p53 directly. In fact, the
phosphorylation levels of p53 correlate positively with p53 protein
levels, as PTX monotherapy increases phosphorylated p53 levels to a
greater degree than the HDAC6 inhibitors either in combination or
as monotherapy. PTX treatment and HDAC6 inhibition may alter p53
levels post-transcriptionally and induce apoptosis in TOV-21G.
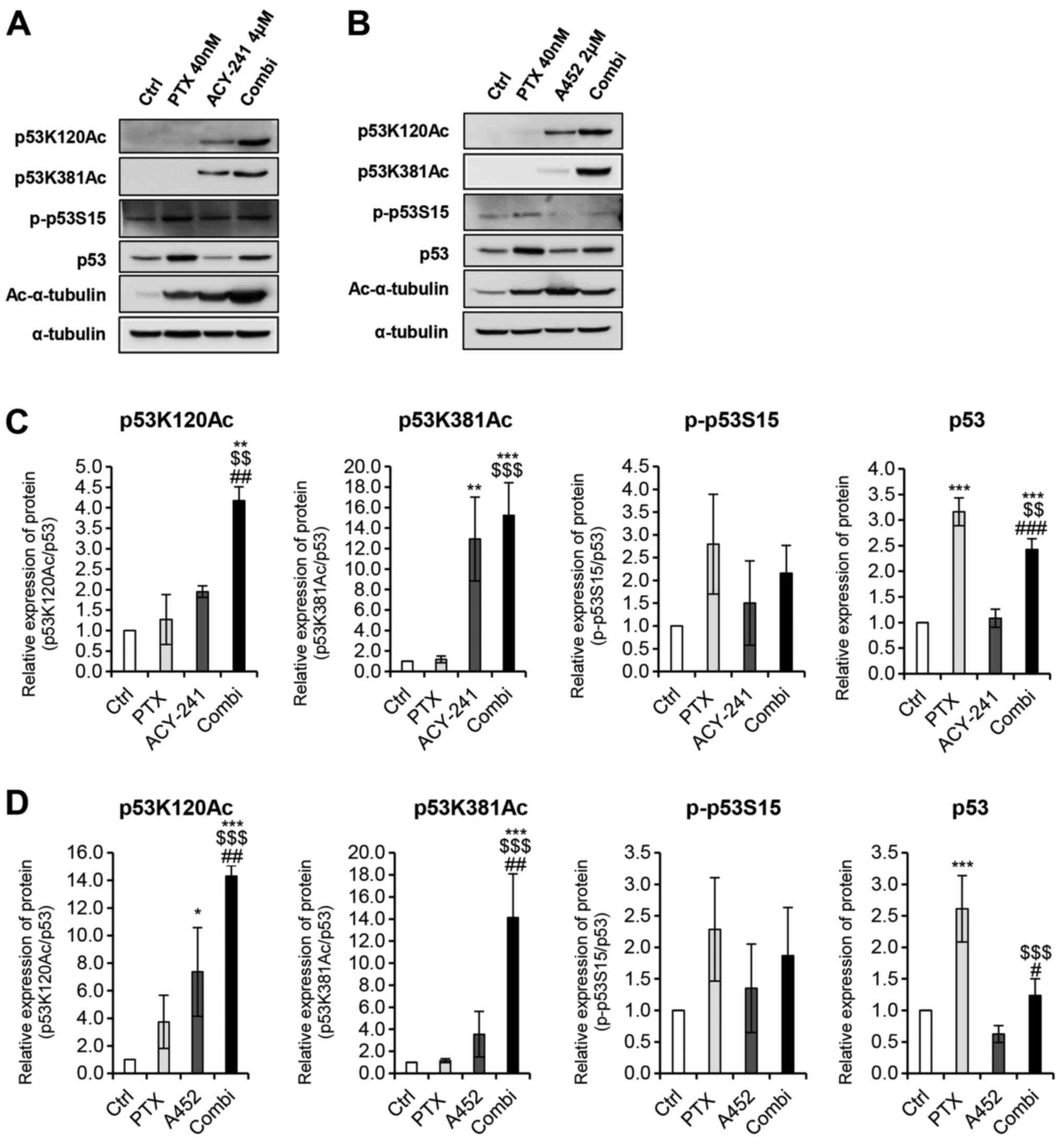 | Figure 6.Combined treatment of histone
deacetylase 6-selective inhibitors and anticancer agent PTX
synergistically alters lysine acetylation modifications of p53.
Western blot analysis was performed using the indicated antibodies.
(A) Combination treatment of PTX (40 nM) and ACY-241 (4 µM)
synergistically altered the acetylation modification of p53. (B)
Combination treatment of PTX (40 nM) and A452 (2 µM)
synergistically altered acetylation modification of p53. (C)
Relative protein levels of acetylated p53 are shown for combination
treatment of PTX (40 nM) and ACY-241 (4 µM). (D) Relative protein
levels of acetylated p53 are shown for combination treatment of PTX
(40 nM) and A452 (2 µM). p53 protein expression was semi-quantified
relative to α-tub; acetylated and phosphorylated p53 proteins were
semi-quantified relative to total p53 levels. The levels in the
0.1% DMSO group were set as 1. α-tub was used as a loading control.
Data are presented as the mean ± SD (n=3). *P<0.05, **P<0.01
and ***P<0.001 vs. DMSO control; $$P<0.01 and
$$$P<0.001 vs. PTX; #P<0.05,
##P<0.01 and ###P<0.001 vs. ACY-241 or
A452, using a one-way ANOVA test. α-tub, α-tubulin; Ac-α-tub,
acetylated-α-tubulin; Ctrl, control; Combi, combination treatment;
PTX, paclitaxel; p53K120Ac, p53 lysine 120 acetylation; p53K381Ac,
p53 lysine 381 acetylation; p-p53S15, p53 serine 15
phosphorylation. |
Discussion
Since its discovery, PTX has been used widely to
treat various types of solid tumors (3), including ovarian cancer (2). Monotherapies and combination therapies
using PTX in ovarian cancer are being used to treat EOCs and are
under various clinical studies. For example, PTX was added to
cisplatin in the first-line treatment of EOCs, and PTX combined
with carboplatin has been the standard treatment for more than 20
years (2,29). In addition to the trials in
combination with platinum-based drugs, PTX with bevacizumab in
recurrent and platinum-sensitive ovarian cancer is under a phase
III clinical trial (30). However,
incomplete clinical responses exist, and advanced ovarian cancer
remains incurable. Thus, there is a need to develop improved,
tolerable chemotherapeutic methods using novel drugs to treat
EOCs.
HDACs are essential epigenetic enzymes that regulate
various cellular pathways. HDAC inhibitors have been used in
multiple studies as alternatives to treat cancer; these induce cell
death, cell cycle arrest, and senescence in tumor cells (31,32).
Pan-HDAC inhibiting agents, such as butyrate, romidepsin, and
vorinostat are under various stages of clinical investigation to
treat various cancers. To date, vorinostat and romidepsin have been
approved by the FDA to treat cutaneous T-cell lymphoma and
peripheral T-cell lymphoma (32).
Despite the various ongoing studies, however, pan-HDAC inhibitors
elicit side effects because they lack the specificity for
individual HDAC isoforms. Therefore, more tolerable and selective
HDAC inhibitors are required, among which HDAC6-selective
inhibitors stand as potent anticancer agents.
As a first-in-class selective and potent HDAC6
inhibitor, ACY-1215 is currently being tested in various clinical
trials and is proven to have antitumor effects in various cancers
in monotherapy or in combination with other agents (9–16).
Besides ACY-1215, which has been explored widely for its clinical
efficacy, other novel HDAC6-selective inhibitors, ACY-241 and A452,
need further extensive research. Until now, it is known that
ACY-241 (citarinostat) synergizes with pomalidomide in multiple
myeloma (24), with PTX in solid
tumor models (17), and with
anti-PD-L1 antibody in multiple myeloma (25). In addition, previous studies with
A452 indicate its potency as an anticancer agent for non-Hodgkin
lymphoma (33), multiple myeloma
(34), immunomodulatory
drugs-resistant multiple myeloma (35), colorectal cancers (19,28), and
glioblastoma (36) in
vitro.
Previous studies have shown that the combination
therapy of HDAC6-selective inhibitor ACY-241 and PTX increased the
anticancer drug efficiency of conventionally used PTX in ovarian
cancer (17). In this study, we have
additionally confirmed the antitumor synergistic effect of PTX and
HDAC6 inhibition using both ACY-241 and a novel inhibitor, A452.
The co-treatment of PTX and the two HDAC6-selective inhibitors in
ARID1A-null ovarian cancer cells synergistically increased
apoptosis (Figs. 4 and 5) and decreased cell growth and viability
(Fig. 2). ARID1A deficient
ovarian cancers fail to inhibit HDAC6 activity, which normally
deacetylates α-tubulin that is crucial for the induction of
microtubule disassembly, leading to tumorigenesis. Thus,
ARID1A-mutant or -null cells are sensitive to HDAC6 inhibition.
Therefore, when PTX, which also inhibits α-tubulin polymerization,
is taken in combination with HDAC6-selective inhibitors, both
α-tubulin polymerization and dissociation is inhibited, leading to
cell death. The deacetylation of tubulin substrates also regulate
microtubule-dependent cell motility, so the inhibition of HDAC6
prevents cell movement and migration (8). Nonetheless, to wholly state the
effectiveness of HDAC6-selective inhibitors and PTX in
ARID1A-mutant ovarian cancers, further studies using other ovarian
cancer cell lines, such as OVISE (clear cell carcinoma, ARID1A
mutant, and wild-type TP53), are required. Using both HDAC6
inhibitors and PTX may also impede metastasis in ovarian cancer
cells. However, metastasis-related proteins matrix
metalloproteinase (MMP)-2 and MMP-9 were not detected (data not
shown), suggesting that the two drug combination might prevail in
inhibiting proliferation over migration. Although more studies are
required to conclusively state the synergistic effect of the two
drugs in ovarian cancer metastasis, the inhibition of cell
proliferation seems to be evident (Fig.
3). Further studies are required to assess the effects of PTX
and HDAC6 inhibition in ovarian cancer cell migration.
Existing studies have demonstrated the synergistic
hyperacetylation of α-tubulin by PTX and ACY-241 (17). In this study, we demonstrated that
A452 also hyperacetylates α-tubulin when treated together with PTX
(Fig. 6). The research data show
that regardless of the type of HDAC6-selective inhibitor used,
HDAC6 inhibition and PTX synergistically regulate α-tubulin in
ARID1A-mutated ovarian cells. In addition to α-tubulin regulation,
we found that p53 acetylation modifications at lysine 120 and 381
were synergistically increased on combination treatment with PTX
using either ACY-241 or A452 (Fig.
6). Although previous studies have shown that PTX and pan-HDAC
inhibition using SAHA, ST2785, and ST3595 can also induce the
synergistic hyperacetylation of p53 (37), we have shown for the first time that
a combination of PTX and HDAC6-selective inhibition increases p53
lysine acetylation. The stabilization of p53 in TOV-21G, which
endogenously carries a wild-type p53, resulted in tumor growth
suppression. However, extended studies using ARID1A-mutant ovarian
cancer cells with wild-type p53 are necessary to strongly confirm
that synergistic tumor growth inhibition involves p53.
Taken together, our data demonstrate the synergistic
anticancer activity of PTX and HDAC6-selective inhibitors. As
mentioned earlier, HDAC6-selective inhibitors have shown high
anticancer potential in different cancers, ranging from blood
cancers to solid cancers. In this study, we show that
HDAC6-selective inhibition through ACY-241 and A452 is capable of
being used in combination with the conventionally used
chemotherapeutic drug, PTX, in vitro. Although we did not
use clinical samples in our study, our results suggest a
proof-of-concept or the preclinical therapeutic possibility of
using ACY-241 and A452 with PTX in treating ovarian cancer by
activating p53 and inducing apoptosis. Further research is
inevitable to elucidate the exact molecular mechanisms behind such
synergism in ovarian cancers. Collectively, this study provides
beneficial information for clinical trials of combination therapy
using HDAC6-selective inhibitors, not only in ovarian cancers but
also in other solid tumors.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Basic Science
Research Program through the National Research Foundation of Korea
funded by the Ministry of Education, Science and Technology (grant
nos. 2018R1A6A1A03023718 and 2019R1I1A1A01058601).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JY and SHK were involved in the general design of
the study. JY, DHL and GWK performed the experiments. JY, YHJ, SYK,
SWL, JP and SHK analyzed the data. JY, YHJ, DHL, GWK, SYK, SWL, JP
and SHK have assessed the authenticity of all raw data. JY wrote
the initial draft of the manuscript, JY and SHK extensively edited
the manuscript, and SHK supervised the work. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Dion L, Carton I, Jaillard S, et al: The
Landscape and Therapeutic Implications of Molecular Profiles in
Epithelial Ovarian Cancer. J Clin Med. 9:22392020. View Article : Google Scholar
|
|
2
|
Jayson GC, Kohn EC, Kitchener HC and
Ledermann JA: Ovarian cancer. Lancet. 384:1376–1388. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Weaver BA: How Taxol/paclitaxel kills
cancer cells. Mol Biol Cell. 25:2677–2681. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Schiff PB, Fant J and Horwitz SB:
Promotion of microtubule assembly in vitro by taxol. Nature.
277:665–667. 1979. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Orr GA, Verdier-Pinard P, McDaid H and
Horwitz SB: Mechanisms of Taxol resistance related to microtubules.
Oncogene. 22:7280–7295. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Boyault C, Sadoul K, Pabion M and Khochbin
S: HDAC6, at the crossroads between cytoskeleton and cell signaling
by acetylation and ubiquitination. Oncogene. 26:5468–5476. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cao J, Lv W, Wang L, Xu J, Yuan P, Huang
S, He Z and Hu J: Ricolinostat (ACY-1215) suppresses proliferation
and promotes apoptosis in esophageal squamous cell carcinoma via
miR-30d/PI3K/AKT/mTOR and ERK pathways. Cell Death Dis. 9:8172018.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li Y, Shin D and Kwon SH: Histone
deacetylase 6 plays a role as a distinct regulator of diverse
cellular processes. FEBS J. 280:775–793. 2013.PubMed/NCBI
|
|
9
|
Yee AJ, Bensinger WI, Supko JG, Voorhees
PM, Berdeja JG, Richardson PG, Libby EN, Wallace EE, Birrer NE,
Burke JN, et al: Ricolinostat plus lenalidomide, and dexamethasone
in relapsed or refractory multiple myeloma: A multicentre phase 1b
trial. Lancet Oncol. 17:1569–1578. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Putcha P, Yu J, Rodriguez-Barrueco R,
Saucedo-Cuevas L, Villagrasa P, Murga-Penas E, Quayle SN, Yang M,
Castro V, Llobet-Navas D, et al: HDAC6 activity is a non-oncogene
addiction hub for inflammatory breast cancers. Breast Cancer Res.
17:1492015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Peng U, Wang Z, Pei S, Ou Y, Hu P, Liu W
and Song J: ACY-1215 accelerates vemurafenib induced cell death of
BRAF-mutant melanoma cells via induction of ER stress and
inhibition of ERK activation. Oncol Rep. 37:1270–1276. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li S, Liu X, Chen X, Zhang L and Wang X:
Histone deacetylase 6 promotes growth of glioblastoma through
inhibition of SMAD2 signaling. Tumour Biol. 36:9661–9665. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Corno C, Arrighetti N, Ciusani E, Corna E,
Carenini N, Zaffaroni N, Gatti L and Perego P: Synergistic
interaction of histone deacetylase 6- and MEK-inhibitors in
castration-resistant prostate cancer cells. Front Cell Dev Biol.
8:6102020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lee DH, Won HR, Ryu HW, Han JM and Kwon
SH: The HDAC6 inhibitor ACY 1215 enhances the anticancer activity
of oxaliplatin in colorectal cancer cells. Int J Oncol. 53:844–854.
2018.PubMed/NCBI
|
|
15
|
Santo L, Hideshima T, Kung AL, Tseng JC,
Tamang D, Yang M, Jarpe M, van Duzer JH, Mazitschek R, Ogier WC, et
al: Preclinical activity, pharmacodynamic, and pharmacokinetic
properties of a selective HDAC6 inhibitor, ACY-1215, in combination
with bortezomib in multiple myeloma. Blood. 119:2579–2589. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Amengual JE, Johannet P, Lombardo M, Zullo
K, Hoehn D, Bhagat G, Scotto L, Jirau-Serrano X, Radeski D, Heinen
J, et al: Dual targeting of protein degradation pathways with the
selective HDAC6 inhibitor ACY-1215 and bortezomib is synergistic in
lymphoma. Clin Cancer Res. 21:4663–4675. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Huang P, Almeciga-Pinto I, Jarpe M, van
Duzer JH, Mazitschek R, Yang M, Jones SS and Quayle SN: Selective
HDAC inhibition by ACY-241 enhances the activity of paclitaxel in
solid tumor models. Oncotarget. 8:2694–2707. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ryu HW, Shin DH, Lee DH, Won HR and Kwon
SH: A potent hydroxamic acid-based, small-molecule inhibitor A452
preferentially inhibits HDAC6 activity and induces cytotoxicity
toward cancer cells irrespective of p53 status. Carcinogenesis.
39:72–83. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Won HR, Ryu HW, Shin DH, Yeon SK, Lee DH
and Kwon SH: A452, an HDAC6-selective inhibitor, synergistically
enhances the anticancer activity of chemotherapeutic agents in
colorectal cancer cells. Mol Carcinog. 57:1383–1395. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bitler BG, Wu S, Park PH, Hai Y, Aird KM,
Wang Y, Zhai Y, Kossenkov AV, Vara-Ailor A, Rauscher FJ III, et al:
ARID1A-mutated ovarian cancers depend on HDAC6 activity. Nat Cell
Biol. 19:962–973. 2017. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Reed SM and Quelle DE: p53 acetylation:
Regulation and consequences. Cancers (Basel). 7:30–69. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Geissmann Q: OpenCFU, a new free and
open-source software to count cell colonies and other circular
objects. PLoS One. 8:e540722013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chou TC: Drug combination studies and
their synergy quantification using the Chou-Talalay method. Cancer
Res. 70:440–446. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
North BJ, Almeciga-Pinto I, Tamang D, Yang
M, Jones SS and Quayle SN: Enhancement of pomalidomide anti-tumor
response with ACY-241, a selective HDAC6 inhibitor. PLoS One.
12:e01735072017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ray A, Das DS, Song Y, Hideshima T, Tai
YT, Chauhan D and Anderson KC: Combination of a novel HDAC6
inhibitor ACY-241 and anti-PD-L1 antibody enhances anti-tumor
immunity and cytotoxicity in multiple myeloma. Leukemia.
32:843–846. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kanno K, Kanno S, Nitta H, Uesugi N, Sugai
T, Masuda T, Wakabayashi G and Maesawa C: Overexpression of histone
deacetylase 6 contributes to accelerated migration and invasion
activity of hepatocellular carcinoma cells. Oncol Rep. 28:867–873.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cho HY, Lee SW, Jeon YH, Lee DH, Kim GW,
Yoo J, Kim SY and Kwon SH: Combination of ACY-241 and JQ1
synergistically suppresses metastasis of HNSCC via regulation of
MMP-2 and MMP-9. Int J Mol Sci. 21:68732020. View Article : Google Scholar
|
|
28
|
Ryu HW, Shin DH, Lee DH, Choi J, Han G,
Lee KY and Kwon SH: HDAC6 deacetylates p53 at lysines 381/382 and
differentially coordinates p53-induced apoptosis. Cancer Lett.
391:162–171. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cong J, Liu R, Hou J, Wang X, Jiang H and
Wang J: Therapeutic effect of bevacizumab combined with paclitaxel
and carboplatin on recurrent ovarian cancer. J BUON. 24:1003–1008.
2019.PubMed/NCBI
|
|
30
|
Coleman RL, Brady MF, Herzog TJ, Sabbatini
P, Armstrong DK, Walker JL, Kim BG, Fujiwara K, Tewari KS, O'Malley
DM, et al: Bevacizumab and paclitaxel-carboplatin chemotherapy and
secondary cytoreduction in recurrent, platinum-sensitive ovarian
cancer (NRG Oncology/Gynecologic Oncology Group study GOG-0213): A
multicentre, open-label, randomised, phase 3 trial. Lancet Oncol.
18:779–791. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Falkenberg KJ and Johnstone RW: Histone
deacetylases and their inhibitors in cancer, neurological diseases
and immune disorders. Nat Rev Drug Discov. 13:673–691. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Mottamal M, Zheng S, Huang TL and Wang G:
Histone deacetylase inhibitors in clinical studies as templates for
new anticancer agents. Molecules. 20:3898–3941. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lee DH, Kim GW and Kwon SH: The
HDAC6-selective inhibitor is effective against non-Hodgkin lymphoma
and synergizes with ibrutinib in follicular lymphoma. Mol Carcinog.
58:944–956. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Won HR, Lee DH, Yeon SK, Ryu HW, Kim GW
and Kwon SH: HDAC6 selective inhibitor synergistically enhances the
anticancer activity of immunomodulatory drugs in multiple myeloma.
Int J Oncol. 55:499–512. 2019.PubMed/NCBI
|
|
35
|
Kim GW, Yoo J, Won HR, Yeon SK, Lee SW,
Lee DH, Jeon YH and Kwon SH: A452, HDAC6-selective inhibitor
synergistically enhances the anticancer activity of
immunomodulatory drugs in IMiDs-resistant multiple myeloma. Leuk
Res. 95:1063982020. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kim GW, Lee DH, Yeon SK, Jeon YH, Yoo J,
Lee SW and Kwon SH: Temozolomide-resistant glioblastoma depends on
HDAC6 activity through regulation of DNA mismatch repair.
Anticancer Res. 39:6731–6741. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zuco V, De Cesare M, Cincinelli R, Nannei
R, Pisano C, Zaffaroni N and Zunino F: Synergistic antitumor
effects of novel HDAC inhibitors and paclitaxel in vitro and in
vivo. PLoS One. 6:e290852011. View Article : Google Scholar : PubMed/NCBI
|