Introduction
Osteosarcoma (OS) is the most common primary
malignant tumor of bone and is frequently diagnosed in children and
young adults (1). Osteosarcoma is
also the most frequent cause of bone cancer-associated death in
children and adolescents (2).
Osteosarcoma mainly arises from the metaphysis of long bones, with
a predilection for the distal end of the femur and the proximal end
of the humerus (3). Common causes of
death for patients with osteosarcoma include tumor recurrence and
metastasis (4). Therefore, it is of
clinical importance to resolve the molecular mechanisms underlying
the occurrence and development of osteosarcoma to improve treatment
strategies.
Cancer stem cells (CSCs) are defined as a distinct
subpopulation of tumor cells that possess stem cell
characteristics, including unlimited self-renewal, tumor-initiating
potential and pluripotent differentiation abilities (5). Several studies haves shown that CSCs
are responsible for cancer recurrence (6,7). CSCs
are similar to normal stem cells, and their differentiation is
influenced by numerous factors, such as genetics and the
environment (8). Researchers have
shown that an increase in the stemness of cancer cells is
associated with hypoxia (9). A
previous study showed that poorly differentiated cancer cells or
CSC-like tumor cells are located in the hypoxic region of
neuroblastoma (10). In addition,
poorly differentiated pancreatic cancer cells have a significantly
increased expression of hypoxia-inducible factor (HIF)-1α protein
in the nucleus (11). Higher levels
of HIF-1α protein are observed in the stem cell-like tumor cells in
gliomas (12). Studies have also
suggested that hypoxia can affect the differentiation of tumor
cells and increase the expression of stem cell markers in tumor
cells (9–13). It has been demonstrated that hypoxia
can activate the mTOR pathway, and the increased expression of
HIF-1α may also be related to the activation of the mTOR pathway
(14). Hypoxia increases the
expression of stem cell markers in brain tumor cells (9). However, it remains unclear whether the
fate of human osteosarcoma cells is regulated by hypoxia. Does
hypoxia enhance the expression of stem cell markers in human
osteosarcoma cells? Is this associated with the mTOR signaling
pathway?
The present study was divided into two parts: In
vitro and in vivo. Firstly, in vitro, human osteosarcoma
cells were cultured under hypoxic conditions, and then changes in
the expression of CSCs markers were investigated. Secondly, in
vivo, human osteosarcoma cells were cultured under hypoxic and
normoxic conditions and subcutaneously injected into nude mice to
compare their tumorigenic ability. Finally, rapamycin was used to
block the effects of hypoxia in osteosarcoma cells to explore the
involvement of the mTOR signaling pathway in the gain of the CSCs
phenotype. The aim of the present study was to determined if
hypoxia enhanced the expression of stem cell markers in human
osteosarcoma cells, and to investigate if this was related to the
mTOR signaling pathway.
Materials and methods
Cell lines and culture
The human osteosarcoma cell line MNNG/HOS was
purchased from The Cell Bank of Type Culture Collection of The
Chinese Academy of Sciences. The cells were cultured in essential
medium MEM supplemented with 10% fetal bovine serum (both HyClone;
Cyvita) and 100 U/ml penicillin/streptomycin in vitro. The
cells were cultured at 37°C under normoxic conditions (5%
CO2, 95% air).
Tumor sphere culture
MNNG/HOS cells were plated at a density of
1×105 cells/well in ultra-low-attachment 6-well plates
(Corning Life Sciences) in MEM cell medium, supplemented with EGF
(20 ng/ml), basic fibroblast growth factor (20 ng/ml), Noggin (10
ng/ml) and leukemia inhibitory factor (1,000 U/ml). The cells were
divided into three groups: Hypoxia, rapamycin and control. Cells in
the control group were cultivated in an incubator at 37°C under
normoxic conditions (5% CO2, 95% air). Cells in the
hypoxia and rapamycin groups were cultivated in an incubator at
37°C under hypoxia (1% O2, 5% CO2 and 94%
nitrogen). Cells in rapamycin group were pretreated with rapamycin
(100 nmol/l) for 30 min before hypoxia culture, with the remaining
culture conditions the same as those of hypoxia group. After 3 days
of culture, spheres were observed and images were captured using an
inverted phase contrast microscope (Eclipse TS100/100-F; Nikon
Corporation), and those with a diameter >100 µm were counted.
The experiments were repeated at least three times.
Flow cytometry
To measure the proportion of CD133+ cells
in single cell suspension of MNNG/HOS cells from the three groups
cultured under different conditions, cells were digested with
trypsin (Sigma-Alrdich; Merck KGaA), centrifuged at room
temperature for 5 min at 200 × g, collected and washed twice with
PBS. Cells were then incubated with CD133-FITC (1:100; Miltenyi
Biotec, Inc.) at 4°C for 60 min in the dark, followed by washing
twice with combining buffer (Beyotime Institute of Biotechnology).
Finally, the labeled cells were analyzed NAVIOS flow cytometer
(Beckman Coulter, Inc.).
Reverse transcription-quantitative
(RT-qPCR)
Total RNA of MNNG/HOS cells from the three groups
was extracted using TRIzol® reagent and treated with
DNase I (both Invitrogen; Thermo Fisher Scientific, Inc.). The
purity of the extracted RNA was determined by the absorbance ratio
at 260/280 on the Nanodrop ND-1000 spectrophotometer (Thermo Fisher
Scientific, Inc.). Then, RNA was reverse transcribed using the 5X
All-In-One RT MasterMix (G492, Applied Biological Materials Inc.)
according to the manufacturer's protocol. RT-qPCR was performed on
a Light Cycler 480 system (Roche Diagnostics) using the SYBER
GreenERTM qPCR SuperMix (Invitrogen; Thermo Fisher Scientific,
Inc.) with the following parameters: Pre-incubation at 95°C for 10
min; 40 cycles amplification at 95°C for 15 sec and 60°C for 1 min.
The relative expression level of the genes was calculated using the
2−ΔΔCq method (15). The
primers used were: Oct-4, Forward: 5′-CTGAGGTGCCTGCCCTTCTA-3′ and
reverse: 5′-CCAACCAGTTGCCCCAAAC-3′; Nanog, forward:
5′-ACCTATGCCTGTGATTTGTGGG-3′ and reverse:
5′-AGAAGTGGGTTGTTTGCCTTTG-3′; GAPDH, forward:
5′-ATGACATCAAGAAGGTGGTG-3′ and reverse: 5′-CATACCAGGAAATGAGCTTG-3′.
Gene (mRNA) expression levels of Oct-4 and Nanog were normalized to
that of GAPDH transcript. Each experiment was repeated at least
three times.
In vivo xenograft experiments
In total, 15 5–6-week old (body weight 20±2 g) male
BALB/c nude mice were purchased from Shanghai Laboratory Animal
Company. The mice were kept in laminar-flow cabinets under specific
pathogen-free conditions and inspected every day. The light dark
cycle was 10 h/14 h. The mice drink water freely. The mice were
given free access to laboratory chow. The nude mice were randomized
into three groups (n=5 for each group). MNNG/HOS cells were
cultured under different conditions of hypoxia, rapamycin and
control groups as aforementioned, and then subcutaneously (right
shoulder blade) injected into nude mice (1×106 cells per
mouse). The size of the xenograft tumors was monitored once a week
after tumor inoculation. The tumor volume (V) was calculated using
the following formula: V (mm3) = 0.5 ab2. The
largest (a) and the smallest (b) superficial diameters of the tumor
were measured with a vernier caliper. The mice were sacrificed 49
days after injection of tumor cells, and the tumors were excised
and weighed. All experiments were approved by The Animal
Experimental Ethics Committee of Fujian Medical University (Fuzhou,
China). None of the mice died during the experimental process.
Western blotting
Whole-cell extracts were obtained using pre-chilled
RIPA [10 mM Tris-Cl (pH 7.4), 150 mM NaCl, 0.1% SDS, 1.0% Triton
X-100, 1.0% Na deoxycholate and 5 mM EDTA] containing protease and
phosphatase inhibitors. Cell lysates were centrifuged at 6,000 × g
for 30 min at 4°C. Then, the protein concentration of the
supernatants was determined. Determination of protein concentration
was performed with the Easy II Protein Quantitative kit (BCA;
Beijing Transgen Biotech Co., Ltd.) in accordance to the
manufacturer's protocol. The total proteins were loaded and
electrophoresed in 10% SDS-PAGE gel and blotted onto nitrocellulose
membrane (EMD Millipore). The membranes were blocked with 5%
non-fat milk in TBST buffer [10 mmol/l Tris-HCl (pH 8.0), 150
mmol/l NaCl, 0.05% Tween-20], and then were incubated overnight
with primary antibodies diluted in TBST at 4°C and subsequently
with horseradish peroxidase-labeled goat anti-rabbit IgG secondary
antibodies (1:1,000; cat. no. A0208; Beyotime Institute of
Biotechnology) for 1 h at room temperature. Primary antibodies
included: Oct-4 (1:1,000; cat. no. ab181557; Abcam), Nanog
(1:1,000; cat. no. ab109250; Abcam), β-actin (1:1,000; cat. no.
ab8227; Abcam), phosphorylated (p-)mTOR (1:1,000; cat. no. ab84400;
Abcam), mTOR (1:1,000; cat. no. ab32028; Abcam), p-P70S6K (1:1,000;
cat. no. AF5899; Beyotime Institute of Biotechnology), P70S6K
(1:1,000; cat. no. AF0258; Beyotime Institute of Biotechnology) and
HIF-1α (1:1,000; cat. no. AF7087; Beyotime Institute of
Biotechnology). The protein bands were visualized using enhanced
chemiluminescence (ECL, Thermo Fisher Scientific, Inc.) and images
were captured using a Kodak X-OMAT 2000A film processor. The
protein was analyzed by Image J software (version 1.52, National
Institutes of Health).
Statistical analysis
All statistical analyses were performed using SPSS
version 20.0 (IBM Corp) and GraphPad Prism version 6.0 (GraphPad
Software). The data are expressed as mean ± standard deviation.
One-way ANOVA followed by Dunnett's post hoc test was used to
determine the statistical significance. P<0.05 was considered to
indicate a statistically significant difference.
Results
Hypoxia increases the sphere formation
of MNNG/HOS cells
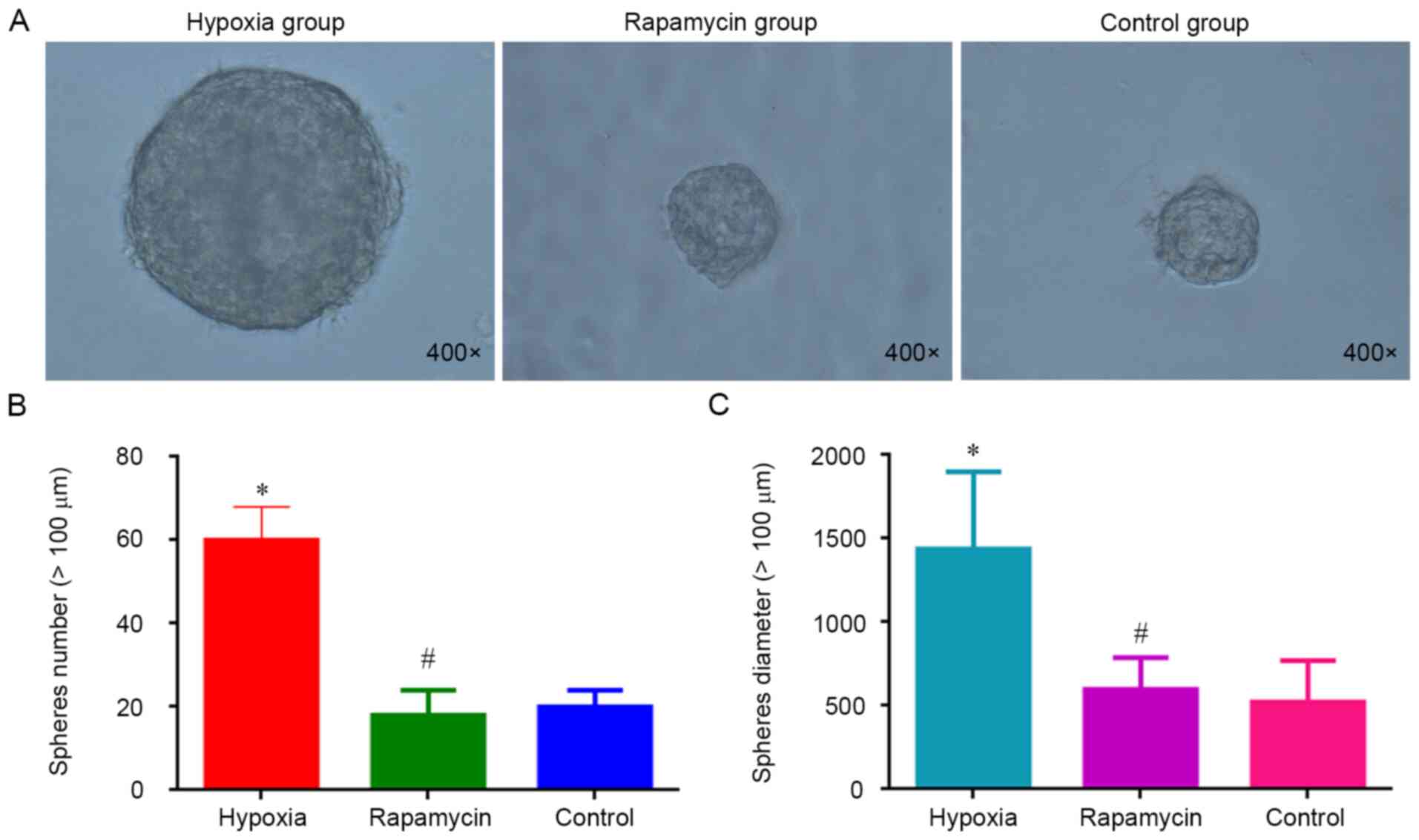
Sphere formation is considered a hallmark of CSCs
(16). To evaluate the effect of
hypoxia on the sphere formation, MNNG/HOS cells were cultured in
ultra-low-attachment plates with stem cell-specific medium under
hypoxia. It was observed that the cells cultured in hypoxia group
were significantly more efficient in forming spheres compared with
the control group (P<0.05). However, there was no difference
regarding the sphere formation ability between the rapamycin and
control groups (Fig. 1). Hypoxia
increases the sphere formation of MNNG/HOS cells and rapamycin can
inhibit this process.
Hypoxia enhances the ability of
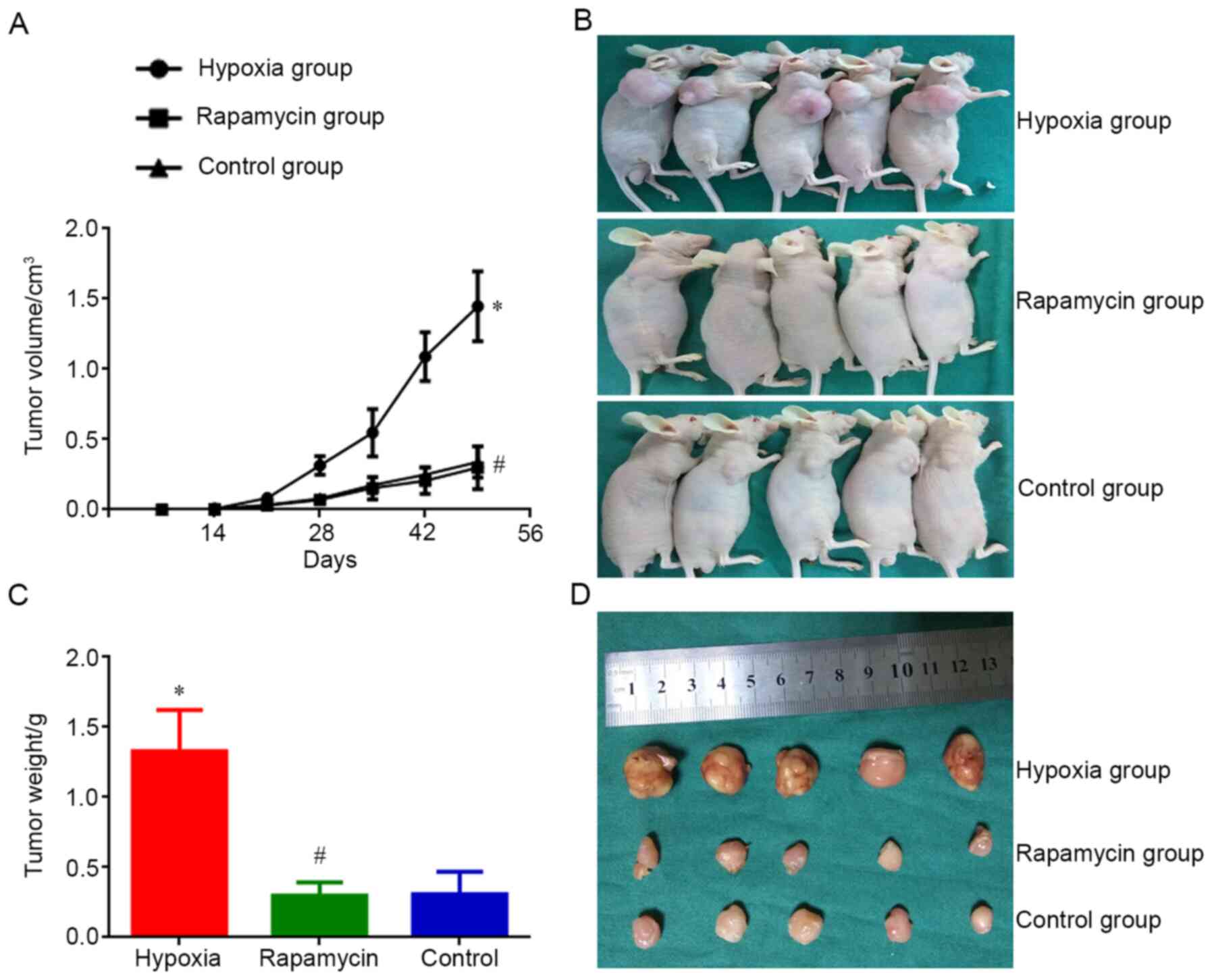
MNNG/HOS cells to form xenograft tumors in vivo
MNNG/HOS cells cultured in each group were injected
into nude mice. Tumors appeared 14 days after the injection. On day
14 after injection, the size of MNNG/HOS xenograft tumors in the
hypoxia group was significantly bigger compared with that in the
control group (P<0.05). However, the tumor size of mice in the
rapamycin group was not different compared with that in the control
group. On day 49 after injection, the size difference of xenograft
tumors was significant between the hypoxia and control groups
(P<0.05). There was also a significant difference in tumor
weight between the two groups (P<0.05). Nevertheless, the
difference regarding average volume and weight of xenograft tumors
between rapamycin and control groups was not different on day 49
after injection (Fig. 2). Hypoxia
enhanceed the ability of MNNG/HOS cells to form xenograft tumors
in vivo and rapamycin can inhibit this process.
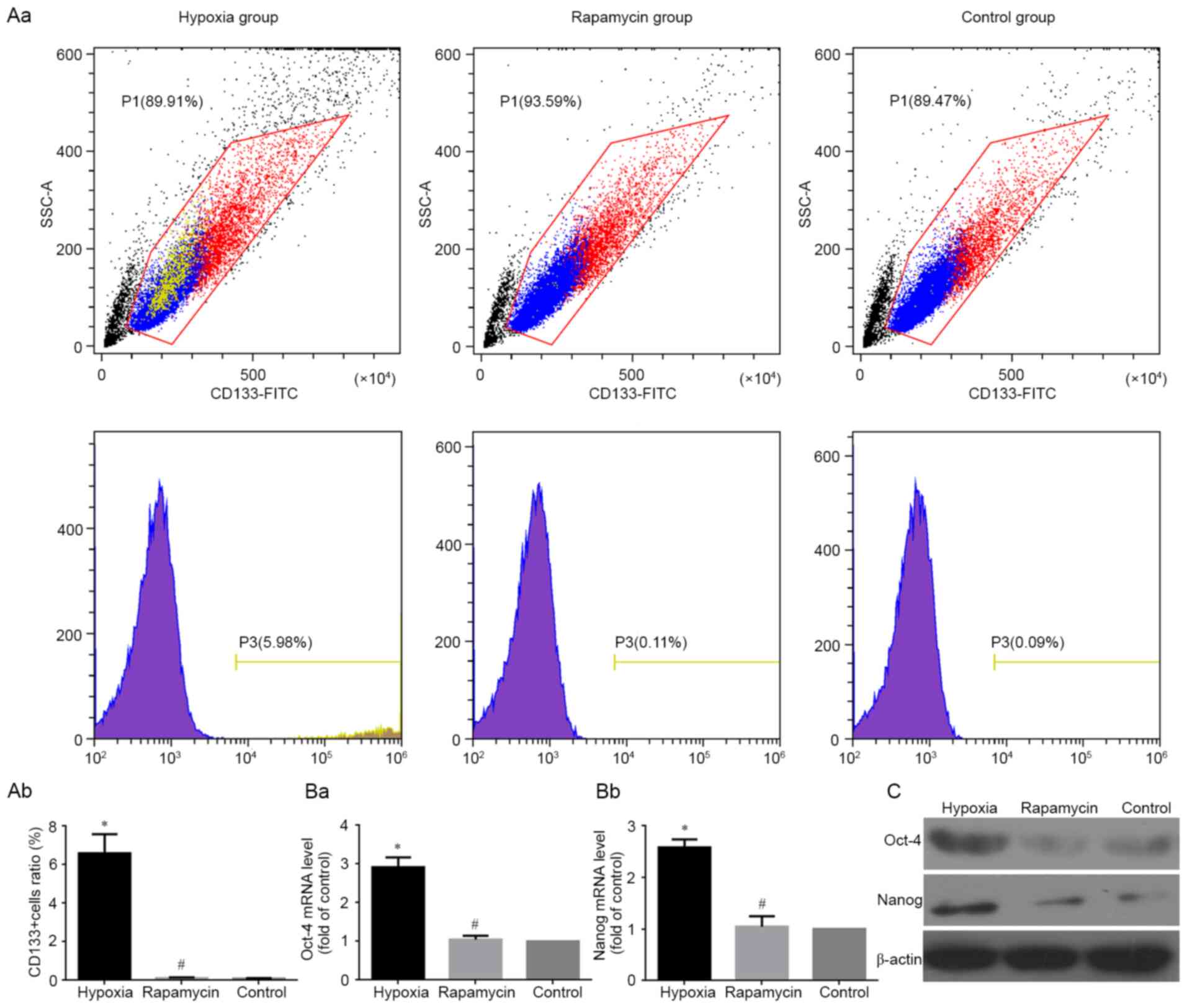
Hypoxia induces the expression of CSC
markers
The stem genes Oct4, Nanog and CD133 have been
identified as CSC markers in several types of tumors (17–19),
including human osteosarcoma (19).
Therefore, the present study detected mRNA and protein expression
of Oct4 and Nanog using RT-qPCR and western blotting, respectively,
in MNNG/HOS cells under different conditions. The results clearly
showed that the mRNA and protein levels of Oct4 and Nanog were
significantly higher in hypoxic group compared with the control
group (both P<0.05). However, there was no difference in the
expression of Oct4 and Nanog between the rapamycin and control
groups (Fig. 3Ba, Bb and C). At the
same time, the percentage of CD133+ subgroup cells were
detected using flow cytometry under different conditions. The
results showed that the percentage of CD133+ subgroup
cells was significantly increased in hypoxic group compared with
the control group (P<0.05). However, the percentage difference
of CD133+ cells between the rapamycin and control groups
were not significant (Fig. 3Aa and
Ab).
HIF-1α is involved in the effects of
hypoxia on MNNG/HOS cells
The expression levels of HIF-1α mRNA and protein in
MNNG/HOS cells cultured under different conditions were examined
using RT-qPCR and western blot analysis, respectively. The mRNA
level of HIF-1α was markedly increased in hypoxia and rapamycin
groups compared with control group (Fig.
4A). The protein expression of HIF-1α was markedly increased in
hypoxia group compared with control group (Fig. 4B and C). However, there was no
significant difference in the protein expression of HIF-1α between
the rapamycin and control groups (Fig.
4B and C). Hypoxia increased the level of HIF-1α protein
expressed by MNNG/HOS cells, and rapamycin inhibited this
process.
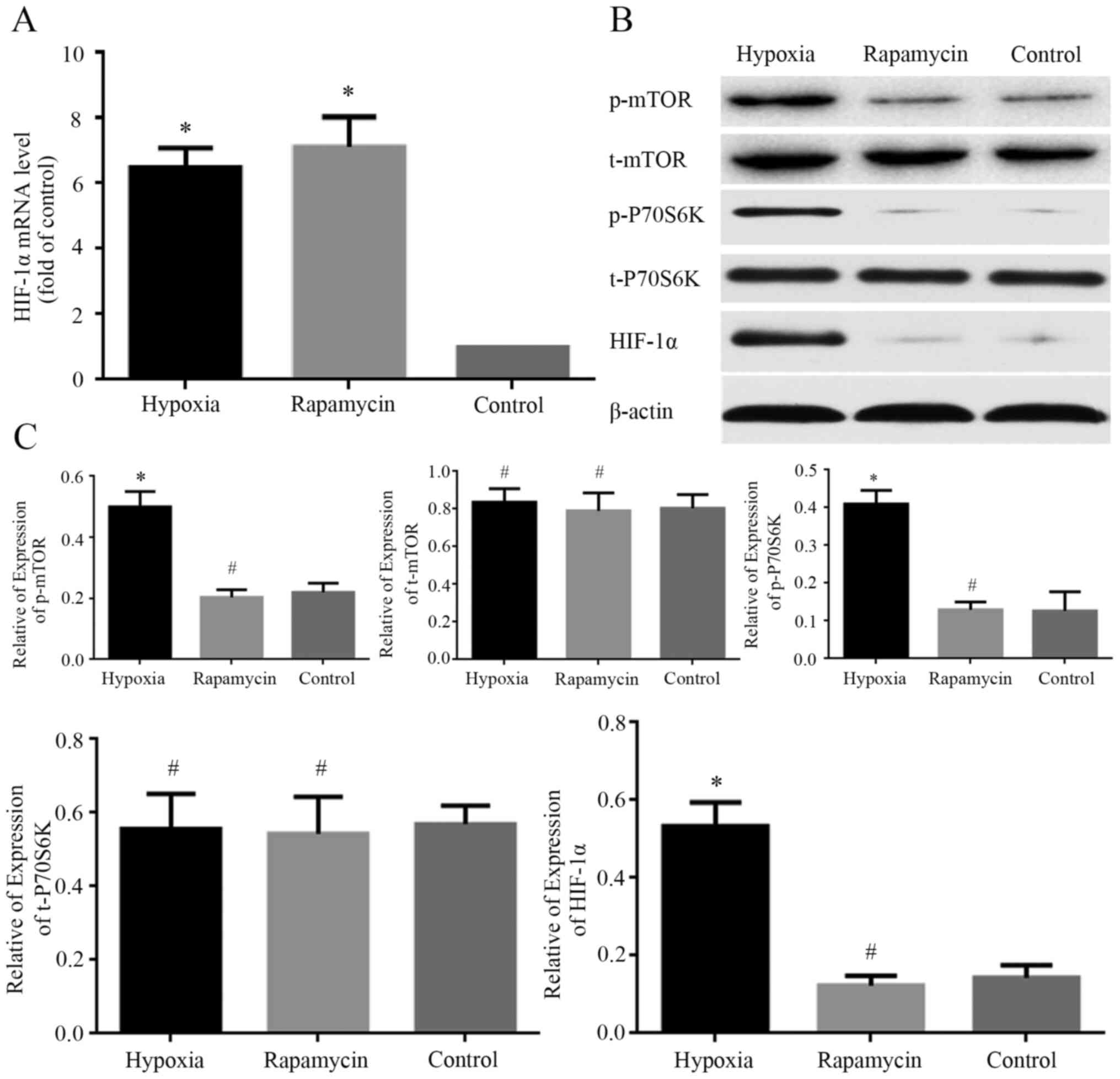 | Figure 4.Change of the expression of HIF-1α,
p-P70S6K, t-P70S6K, p-mTOR, t-mTOR. (A) mRNA level of HIF-1α
analyzed using reverse transcription-quantitative PCR. All data are
normalized to GAPDH and represented as relative expression
(relative to control). (B) Representative images showing the
expression of HIF-1α, p-mTOR, t-mTOR, p-P70S6K and t-P70S6K in
MNNG/HOS cells in different groups. (C) Quantification of
expression levels of HIF-1α, p-mTOR, t-mTOR, p-P70S6k and t-P70S6k
in MNNG/HOS cells in different groups. β-actin was used as a
loading control. *P<0.05 and #P>0.05 vs. control
group. HIF-1α, hypoxia-inducible factor 1α; p-, phosphorylated; t-,
total. |
To explore the mechanism of HIF-1α involvement in
hypoxia-induced effects on MNNG/HOS cells, the expression levels of
mTOR signaling proteins under different conditions were examined
using western blotting. The phosphorylated levels of mTOR and
P70S6K were markedly increased in the hypoxia group compared with
control group. However, there was no significant difference in the
phosphorylated expression of the two proteins between the rapamycin
and control groups (Fig. 4B and C).
Meanwhile, there was no difference in total protein expression of
mTOR and P70S6K among the three groups (Fig. 4B and C). The phosphorylated levels of
mTOR and P70S6K were markedly increased under hypoxia condition and
rapamycin can inhibit this process.
Discussion
CSCs are a small group of cancer cells that have
stem cell properties of self-renewal and differentiation (20). CSCs also play a crucial role in tumor
initiation, progression and invasion of cancer (21). CSCs have been identified in several
solid tumors, such as brain cancer (22), ovarian cancer (23) and leiomyosarcoma (24). Numerous types of malignant tumor
cells dedifferentiate into stem-like cell phenotypes under hypoxia
(25–27). However, it remains unclear whether
the human osteosarcoma cells can dedifferentiate into stem-like
cells under hypoxia. The present data reported that hypoxia induced
the stemness transformation of human osteosarcoma cells as
demonstrated by increased expression of stem cell markers,
promoting the formation of spheres and xenograft tumorigenesis. The
mTOR signaling pathway was also involved in the transformation
process via regulating activation of the HIF-1α protein.
Studies have shown that hypoxia can increase the
expression of stem cell markers in several malignant cell types,
such as glioblastoma (28), breast
cancer (29) and neuroblastoma
(25). Hypoxia, a common feature of
several solid tumors, including osteosarcoma, is associated with
tumor progression and poor prognosis (30–32). In
the present study, hypoxia led to increased expression of stem cell
markers, including Oct4, Nanog and CD133, in MNNG/HOS cells.
Moreover, hypoxia promoted the formation of spheres and xenograft
tumorigenesis of MNNG/HOS cells. These results suggested that
hypoxia can dedifferentiate MNNG/HOS cells into stem cell-like
phenotypes, which may explain the contributing role of hypoxia to
these malignant properties.
Transcriptional response to hypoxia is commonly
regulated by HIFs (33). HIFs are
composed of two subunits, HIF-α and HIF-β (34). HIF-1α is the regulatory subunit of
HIF-1, a crucial mediator of the cellular response to hypoxia
(35). Therefore, HIF-1α is a key
regulator for hypoxia tolerance of osteosarcoma (36). The present study observed an
increased level of HIF-1α under hypoxic conditions in MNNG/HOS
cells. Rapamycin can reduce HIF-1α protein expression through
promoting HIF-1α protein degradation (37). The present research also showed that
hypoxia-induced dedifferentiation of MNNG/HOS cells into stem cell
phenotypes could be inhibited rapamycin. This suggested that HIF-1α
plays an important role in the dedifferentiation of human
osteosarcoma cells into stem cell-like cells.
P70S6K1 is one of the most important effectors
downstream of the mTOR pathway. mTOR can phosphorylate P70S6K to
phosphorylate 40S ribosomal protein s6 in cells, thereby regulating
the transcription and translation of mRNA and protein synthesis
(38). Studies have shown that
hypoxia can activate the mTOR pathway, and the increased expression
of HIF-1 may also be related to the activation of the mTOR pathway
(14). In the present experiment,
under hypoxic conditions, the increased expression of p-mTOR and
p-P70S6K was expected. The present study showed that the activation
of mTOR pathway under hypoxic conditions was associated with the
expression levels of CSCs markers in osteosarcoma cells and the
formation of spheres and xenograft tumors.
Rapamycin can inhibit the activation effect of mTOR
signaling pathway caused by hypoxia, and reduce the expression
levels of mTOR pathway-related proteins (39). Rapamycin was one of the first mTOR
inhibitors used to inhibit the entire pathway (40). Rapamycin was used in the present
study to further verify the activation effect of mTOR signaling
pathway in osteosarcoma cells under hypoxia and its role in the
expression levels of CSCs markers and the formation of spheroid and
xenograft tumors. Under the action of rapamycin, the expression of
p-mTOR and p-P70S6K in osteosarcoma cells of the rapamycin group
were significantly reduced, and the mTOR signaling pathway was
inhibited, indicating that the effect of rapamycin was as expected.
Compared with the hypoxia group, osteosarcoma cells treated with
rapamycin had lower expression levels of CSCs markers, and
inhibited spheroid formation and xenograft tumor formation. These
results indicated that inhibition or deactivation of the mTOR
signaling pathway can affect osteosarcoma cell differentiation into
a stem cell-like phenotype.
In conclusion, the present study demonstrated that
hypoxia induced stemness transformation of human osteosarcoma
cells, promoting the formation of spheres and xenograft tumor.
HIF-1α activated by mTOR signaling was involved in this process and
could be a therapeutic target to inhibit the malignant
transformation of osteosarcoma cells.
Acknowledgements
The authors would like to thank Dr Guangxian Zhong
(Orthopaedic Research Institute, The First Affiliated Hospital of
Fujian Medical University, Fuzhou, China) for providing technical
assistance and useful discussions.
Funding
The present study was supported by The Scientific
Research Project for Fujian Provincial Health and Youth Health
Research Project (grant no. 2019-1-45), The Startup Fund for
Scientific Research, Fujian Medical University (grant no.
2018QH1055) and The Nature Science Foundation of Fujian Province of
China (grant no. 2017J01278).
Availability of data and materials
The data sets used and/or analyzed during the
current study are available from the corresponding author on
reasonable request.
Authors' contributions
JLL, XinwuW and XinwenW conducted the experiments
and drafted the manuscript. SLW and RKS contributed to statistical
analysis and manuscript writing. YBY and JYX participated in
performing the cell experiments. JHL conceived the present study
and helped revise the manuscript. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
The experiments were approved by the Animal Ethics
Committee of Fujian Medical University
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Huang CY, Wei PL, Wang JW, Makondi PT,
Huang MT, Chen HA and Chang YJ: Glucose-regulated protein 94
modulates the response of osteosarcoma to chemotherapy. Dis
Markers. 2019:45697182019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Urciuoli E, Giorda E, Scarsella M, Petrini
S and Peruzzi B: Osteosarcoma-derived extracellular vesicles induce
a tumor-like phenotype in normal recipient cells. J Cell Physiol.
233:6158–6172. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li Y, Zeng C, Tu M, Jiang W, Dai Z, Hu Y,
Deng Z and Xiao W: MicroRNA-200b acts as a tumor suppressor in
osteosarcoma via targeting ZEB1. Onco Targets Ther. 9:3101–3111.
2016.PubMed/NCBI
|
|
4
|
Zhang Z, Luo G, Yu C, Yu G, Jiang R and
Shi X: MicroRNA-493-5p inhibits proliferation and metastasis of
osteosarcoma cells by targeting Kruppel-like factor 5. J Cell
Physiol. 234:13525–13533. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Corrò C and Moch H: Biomarker discovery
for renal cancer stem cells. J Pathol Clin Res. 4:3–18. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wahab SMR, Islam F, Gopalan V and Lam AK:
The identifications and clinical implications of cancer stem cells
in colorectal cancer. Clin Colorectal Cancer. 16:93–102. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yi M, Li J, Chen S, Cai J, Ban Y, Peng Q,
Zhou Y, Zeng Z, Peng S, Li X, Xiong W, Li G and Xiang B: Correction
to: Emerging role of lipid metabolism alterations in Cancer stem
cells. J Exp Clin Cancer Res. 37:1552018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jiang P, Zhang Y, Zhu C, Zhang W, Mao Z
and Gao C: Fe3O4/BSA particles induce
osteogenic differentiation of mesenchymal stem cells under static
magnetic field. Acta Biomater. 46:141–150. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Prasad P, Mittal SA, Chongtham J, Mohanty
S and Srivastava T: Hypoxia-mediated epigenetic regulation of
stemness in brain tumor cells. Stem Cells. 35:1468–1478. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lin Q and Yun Z: Impact of the hypoxic
tumor microenvironment on the regulation of cancer stem cell
characteristics. Cancer Biol Ther. 9:949–956. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Couvelard A, O'Toole D, Turley H, Leek R,
Sauvanet A, Degott C, Ruszniewski P, Belghiti J, Harris AL, Gatter
K and Pezzella F: Microvascular density and hypoxia-inducible
factor pathway in pancreatic endocrine tumours: negative
correlation of microvascular density and VEGF expression with
tumour progression. Br J Cancer. 92:94–101. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li Z, Bao S, Wu Q, Wang H, Eyler C,
Sathornsumetee S, Shi Q, Cao Y, Lathia J, McLendon RE, et al:
Hypoxia-inducible factors regulate tumorigenic capacity of glioma
stem cells. Cancer Cell. 15:501–513. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kim Y, Lin Q, Glazer PM and Yun Z: Hypoxic
tumor microenvironment and cancer cell differentiation. Curr Mol
Med. 9:425–434. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yuan G, Nanduri J, Khan S, Semenza GL and
Prabhakar NR: Induction of HIF-1alpha expression by intermittent
hypoxia: Involvement of NADPH oxidase, Ca2+ signaling,
prolyl hydroxylases, and mTOR. J Cell Physiol. 217:674–685. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lin X, Sun B, Zhu D, Zhao X, Sun R, Zhang
Y, Zhang D, Dong X, Gu Q, Li Y, et al: Notch4+ cancer
stem-like cells promote the metastatic and invasive ability of
melanoma. Cancer Sci. 107:1079–1091. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fan Z, Li M, Chen X, Wang J, Liang X, Wang
H, Wang Z, Cheng B and Xia J: Prognostic value of cancer stem cell
markers in head and neck squamous cell carcinoma: a meta-analysis.
Sci Rep. 7:430082017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bradshaw A, Wickremsekera A, Tan ST, Peng
L, Davis PF and Itinteang T: Cancer stem cell hierarchy in
glioblastoma multiforme. Front Surg. 3:212016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang L, Park P, Zhang H, La Marca F and
Lin CY: Prospective identification of tumorigenic osteosarcoma
cancer stem cells in OS99-1 cells based on high aldehyde
dehydrogenase activity. Int J Cancer. 128:294–303. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhao Y, Zhao W, Lim YC and Liu T:
Salinomycin-loaded gold nanoparticles for treating cancer stem
cells by ferroptosis-induced cell death. Mol Pharm. 16:2532–2539.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zeng L, Cen Y and Chen J: Long non-coding
RNA MALAT-1 contributes to maintenance of stem cell-like phenotypes
in breast cancer cells. Oncol Lett. 15:2117–2122. 2018.PubMed/NCBI
|
|
22
|
Mukherjee S, Baidoo JNE, Sampat S, Mancuso
A, David L, Cohen LS, Zhou S and Banerjee P: Liposomal TriCurin, a
synergistic combination of curcumin, epicatechin gallate and
resveratrol, repolarizes tumor-associated microglia/macrophages,
and eliminates glioblastoma (GBM) and GBM stem cells. Molecules.
23:2012018. View Article : Google Scholar
|
|
23
|
Krishnapriya S, Sidhanth C, Manasa P,
Sneha S, Bindhya S, Nagare RP, Ramachandran B, Vishwanathan P,
Murhekar K, Shirley S, et al: Cancer stem cells contribute to
angiogenesis and lymphangiogenesis in serous adenocarcinoma of the
ovary. Angiogenesis. 22:441–455. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fourneaux B, Bourdon A, Dadone B, Lucchesi
C, Daigle SR, Richard E, Laroche-Clary A, Le Loarer F and Italiano
A: Identifying and targeting cancer stem cells in leiomyosarcoma:
Prognostic impact and role to overcome secondary resistance to
PI3K/mTOR inhibition. J Hematol Oncol. 12:112019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mohlin S, Wigerup C, Jögi A and Påhlman S:
Hypoxia, pseudohypoxia and cellular differentiation. Exp Cell Res.
356:192–196. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang P, Lan C, Xiong S, Zhao X, Shan Y, Hu
R, Wan W, Yu S, Liao B, Li G, et al: HIF1α regulates single
differentiated glioma cell dedifferentiation to stem-like cell
phenotypes with high tumorigenic potential under hypoxia.
Oncotarget. 8:28074–28092. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Maeda K, Ding Q, Yoshimitsu M, Kuwahata T,
Miyazaki Y, Tsukasa K, Hayashi T, Shinchi H, Natsugoe S and Takao
S: CD133 modulate HIF-1α expression under hypoxia in EMT phenotype
pancreatic cancer stem-like cells. Int J Mol Sci. 17:10252016.
View Article : Google Scholar
|
|
28
|
Bar EE, Lin A, Mahairaki V, Matsui W and
Eberhart CG: Hypoxia increases the expression of stem-cell markers
and promotes clonogenicity in glioblastoma neurospheres. Am J
Pathol. 177:1491–1502. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Conley SJ, Gheordunescu E, Kakarala P,
Newman B, Korkaya H, Heath AN, Clouthier SG and Wicha MS:
Antiangiogenic agents increase breast cancer stem cells via the
generation of tumor hypoxia. Proc Natl Acad Sci U S A.
109:2784–2789. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Guo Q, Lan F, Yan X, Xiao Z, Wu Y and
Zhang Q: Hypoxia exposure induced cisplatin resistance partially
via activating p53 and hypoxia inducible factor-1α in non-small
cell lung cancer A549 cells. Oncol Lett. 16:801–808.
2018.PubMed/NCBI
|
|
31
|
Yang ZL, Tian W, Wang Q, Zhao Y, Zhang YL,
Tian Y, Tang YX, Wang SJ, Liu Y, Ni QQ, et al: Oxygen-evolving
mesoporous organosilica coated prussian blue nanoplatform for
highly efficient photodynamic therapy of tumors. Adv Sci (Weinh).
5:17008472018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhao H, Wu Y, Chen Y and Liu H: Clinical
significance of hypoxia-inducible factor 1 and VEGF-A in
osteosarcoma. Int J Clin Oncol. 20:1233–1243. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Manresa MC and Taylor CT: Hypoxia
inducible factor (HIF) hydroxylases as regulators of intestinal
epithelial barrier function. Cell Mol Gastroenterol Hepatol.
3:303–315. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gorga A, Rindone G, Regueira M, Riera MF,
Pellizzari EH, Cigorraga SB, Meroni SB and Galardo MN: HIF
involvement in the regulation of rat Sertoli cell proliferation by
FSH. Biochem Biophys Res Commun. 502:508–514. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bao L, Chen Y, Lai HT, Wu SY, Wang JE,
Hatanpaa KJ, Raisanen JM, Fontenot M, Lega B, Chiang CM, et al:
Methylation of hypoxia-inducible factor (HIF)-1α by G9a/GLP
inhibits HIF-1 transcriptional activity and cell migration. Nucleic
Acids Res. 46:6576–6591. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Guo M, Cai C, Zhao G, Qiu X, Zhao H, Ma Q,
Tian L, Li X, Hu Y, Liao B, et al: Hypoxia promotes migration and
induces CXCR4 expression via HIF-1α activation in human
osteosarcoma. PLoS One. 9:e905182014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Dodd KM, Yang J, Shen MH, Sampson JR, Tee
AR and KM D: mTORC1 drives HIF-1α and VEGF-A signalling via
multiple mechanisms involving 4E-BP1, S6K1 and STAT3. Oncogene.
34:2239–2250. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Cornu M, Albert V, Hall MN and M C: mTOR
in aging, metabolism, and cancer. Curr Opin Genet Dev. 23:53–62.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Verheul HM, Salumbides B, Van Erp K,
Hammers H, Qian DZ, Sanni T, Atadja P and Pili R: Combination
strategy targeting the hypoxia inducible factor-1 alpha with
mammalian target of rapamycin and histone deacetylase inhibitors.
Clin Cancer Res. 14:3589–3597. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hou H, Miao J, Cao R, Han M, Sun Y, Liu X
and Guo L: Rapamycin ameliorates experimental autoimmune
encephalomyelitis by suppressing the mTOR-STAT3 pathway. Neurochem
Res. 42:2831–2840. 2017. View Article : Google Scholar : PubMed/NCBI
|