Introduction
Head and neck squamous cell carcinoma (HNSCC) is the
sixth most common malignancy worldwide (1). There are ~650,000 new cases of HNSCC
diagnosed every year, with >350,000 associated deaths occurring
in a year (2). Several risk factors
involved in the development of HNSCC have been identified,
including alcohol, tobacco use and human papillomavirus (HPV)
infection, the latter of which is a major independent risk factor
for HNSCC (3). Compared with
HPV-positive HNSCC, HPV-negative HNSCC is more aggressive and is
associated with a higher rate of drug resistance (4). Similarly, patients with HPV-negative
HNSCC demonstrate lower overall survival rates than those with
HPV-positive HNSCC (5). The
identification of novel and specific biomarkers for HPV-negative
HNSCC may therefore help to improve the understanding of the
mechanism that underlies the progression of HPV-negative HNSCC, as
well as to improve the diagnosis and treatment of the disease.
With the progression of high-throughput sequencing
technology and bioinformatics methods, additional oncogenes
involved in the development of HNSCC have been identified (6,7).
Weighted gene co-expression network analysis (WGCNA) is a
bioinformatics method that involves calculating the associations
between gene co-expression modules and clinical traits using a
weighted soft threshold (8).
Recently, with the use of WGCNA, a series of biomarkers for HNSCC
have been identified. For example, Song et al (9) identified 16 genes involved in the
immune and inflammatory response that promoted the development of
HNSCC. Similarly, using WGCNA, Zhang et al (10) identified 12 genes associated with
perineural invasion in patients with HNSCC. However, the molecular
mechanism of each HNSCC subtype is different, and the feasibility
and specificity of the majority of biomarkers obtained from
previous studies are limited.
The current study combined WGCNA, differential
expression gene (DEG) analysis and experimental verification to
identify specific biomarkers for HPV-negative HNSCC.
Materials and methods
Clinical specimens
A total of 62 pairs of primary HNSCC tissues and
adjacent non-tumor tissues (1-2 cm distance from the tumor tissues)
were collected from patients at The Affiliated Hospital of Guizhou
Medical University (Guiyang, China). None of the enrolled patients
underwent treatment (such as chemotherapy or radiation) prior to
tissue collection. HPV infection status had been determined prior
to study commencement by performing in situ hybridization
(ISH), as previously described (11), and P16 staining (Fig. S1). HPV-positive HNSCC was diagnosed
based on ISH or P16 (also named p16INK4a) positive results, while
HPV-negative HNSCC was defined based on negative ISH and P16
results. A total of 35 patients with HPV-positive HNSCC (26 tissues
were ISH and P16 positive, 9 tissues were ISH positive only and
none of the tissues were P16 positive only) and 27 patients with
HPV-negative HNSCC were enrolled in the present study. The basic
clinical characteristics (American Joint Committee on Cancer)
(12) of the patients with
HPV-negative and HPV-positive HNSCC are presented in Table I. The current study was approved by
the Human Trait Ethics Committee of Guizhou Medical University and
performed in accordance with the Declaration of Helsinki. All
patients who provided samples signed written informed consent.
 | Table I.Basic clinical characteristics of
patients with HPV-negative (n=27) and HPV-positive (n=35)
HNSCC. |
Table I.
Basic clinical characteristics of
patients with HPV-negative (n=27) and HPV-positive (n=35)
HNSCC.
| Clinicopathological
feature | HPV-negative
HNSCC | HPV-positive
HNSCC | P-value |
|---|
| Mean age ± SD,
years | 71.4±6.7 | 72.4±7.3 | >0.05 |
| Sex, n (%) |
|
| >0.05 |
|
Male | 12 (44.4) | 17 (48.5) |
|
|
Female | 15 (55.6) | 18 (51.5) |
|
| Tumor grade
(Broders), n (%) |
|
| >0.05 |
| Grade
I | 12 (44.4) | 16 (45.7) |
|
| Grade
II | 13 (48.1) | 16 (45.7) |
|
| Grade
III | 2
(7.5) | 3
(9.6) |
|
| Metastasis at
diagnosis, n (%) |
|
| >0.05 |
|
Yes | 2
(7.5) | 4
(11.4) |
|
| No | 25 (92.5) | 31 (88.6) |
|
| Tumor subsite, n
(%) |
|
| >0.05 |
| Oral
tongue | 10 (37.0) | 13 (37.1) |
|
| Buccal
mucosa | 2
(7.4) | 2
(5.7) |
|
|
Larynx | 7
(26.0) | 9
(25.7) |
|
| Oral
cavity | 4
(14.8) | 5
(14.2) |
|
| Floor
of mouth | 4
(14.8) | 6
(17.1) |
|
Data processing
For constructing the WGCNA, the gene expression
profiles of 54 HPV-negative HNSCC tissues (ISH and P16 negative
stain), along with their corresponding clinical characteristics
(Table SI), were downloaded from
The Cancer Genome Atlas (TCGA) database (https://portal.gdc.cancer.gov/). After normalization,
outliers were assessed using the hierarchical cluster algorithm
with a cut-off of 140. A fragments per kilobase of transcript per
million mapped reads (FPKM) value of <0.5 was used as the
threshold for removing the genes with low expression. This was
necessary as a gene must be expressed at a minimal level before it
is likely to be translated into a protein or be considered
biologically important (13).
Consequently, 15,703 genes expressed in the 54 HPV-negative HNSCC
tissues, along with the corresponding clinical characteristics
data, were used in the WGCNA. For DEGs analysis, the gene
expression prolife in TCGA and Gene Expression Omnibus datasets
[GSE117973 (14), GSE85446 (15) and GSE112026 (16); https://www.ncbi.nlm.nih.gov/gds] containing 103
HPV-positive HNSCC and 165 HPV-negative HNSCC was downloaded. After
merge and batch normalization using the sva package (version: 3.11;
Bioconductor; https://www.bioconductor.org/packages/release/bioc/html/sva.htl),
the merge gene expression profile were used for DEGs analysis. |Log
fold-change (FC)| >0.5 and adjusted P<0.05 were set as
cut-off values to consider genes differentially expressed between
HPV-positive and HPV-negative HNSCC.
WGCNA
The WGCNA R package (version 1.69; https://cran.r-project.org/web/packages/WGCNA/index.html)
was used to conduct the WGCNA. All gene pairs were analyzed using
Pearson's correlation analysis, the results of which were used to
construct a matrix of similarity. Subsequently, to produce a
scale-free co-expression network, the matrix of similarity was
constructed using a soft power of β=7. The adjacency matrix was
then translated into a topological overlap matrix (TOM).
Furthermore, median linkage hierarchical clustering was analyzed
using the TOM-based dissimilarity measure with a minimum size of
50.
Identification of clinically
significant modules
Following WGCNA, the Pearson's correlation analyses
between various module eigengenes and the clinical characteristics
of patients were assessed. These traits included node (N) stage,
tumor (T) stage, tumor grade, recurrence, perineural invasion,
tobacco use history and alcohol consumption history. Modules were
considered clinically significant if they were correlated with two
clinical characteristics (r>0.3 and P<0.05). In the
clinically significant modules, gene significance (GS) was
quantified using associations between the individual genes and the
clinical characteristics of interest, along with the module
membership (MM), which was itself determined using the correlation
between the module eigengenes and the gene expression profiles. If
P<0.05 for the correlation between GS and MM in the clinically
significant modules, the modules were subjected to further
analysis.
Construction of a protein-protein
interaction (PPI) network
The online Search Tool for the Retrieval of
Interacting Genes/Proteins (STRING; http://string-db.org) was used to construct the PPI
network. Nodes without connections were removed. The minimum
required interaction score was set at 0.7 (high confidence). Node
and edge information was exported into a text file, which was
subsequently imported into Cytoscape software (version 6.1;
http://cytoscape.org/). In the PPI network, nodes
represented the protein information encoded by the gene of
interest. Lines indicated the interactions between two proteins.
The degree score was calculated using the Cytohub plug-in (version
1.0; http://github.com/cytoscape/appstore) and was set as
the criterion used to calculate the interaction between a single
protein and other proteins. Finally, proteins encoded by genes with
a degree score in the top 10% were visualized using Cytoscape, and
the genes coding for proteins with a degree score in the top 1%
were considered candidate hub genes.
Pearson's correlation analysis between
the candidate hub genes and tumor grade
Pearson's correlation analysis between the
expression of candidate hub genes and tumor grade was performed
using SPSS version 20.0 (IBM Corp.). P<0.05 was set as the
cut-off indicating a significant correlation. The genes
significantly correlated with tumor grade were considered hub
genes.
Immunohistochemical (IHC)
staining
All 27 HPV-negative and 35 HPV-positive HNSCC
tissues were fixed in 4% paraformaldehyde (Boster Biological
Technology) at room temperature. Subsequently, they were embedded
into paraffin and cut into 4-µm-thick sections. After heating at
65°C, the sections were deparaffinized using xylene and rehydrated
in a graded ethanol series (100, 80, 60 and 40%). After conducting
antigen retrieval using sodium citrate (0.01 mol/l; Boster
Biological Technology), endogenous peroxidase activity was blocked
using 3% H2O2. Subsequently, 5% bovine serum
albumin (Wuhan Servicebio Technology Co., Ltd.) was added at room
temperature for 1 h. The sections were then incubated for 16 h at
4°C with the following primary antibodies: p16INK4a (1:200; cat.
no. A11651; ABclonal Biotech Co., Ltd.), Serrate RNA effector
molecule (SRRT; 1:200; cat. no. A8219; ABclonal Biotech Co., Ltd.),
checkpoint kinase 2 (CHEK2; 1:200; cat. no. A19543; ABclonal
Biotech Co., Ltd.), small nuclear ribonucleoprotein polypeptide E
(SNRPE; 1:200; cat. no. A5488; ABclonal Biotech Co., Ltd.),
proteasome 26S subunit ATPase 2 (PSMC2; 1:400; cat. no. 14905-1-AP;
ProteinTech Group, Inc.), origin recognition complex subunit 5
(ORC5; 1:400; cat no. 11542-1-AP, ProteinTech Group, Inc.), S100
calcium binding protein A7 (S100A7; 1:200; cat no. 13061-1-AP,
ProteinTech Group Inc.) and keratinocyte differentiation associated
protein (KRTDAP; 1:1,000; cat. no. ab204583; Abcam). Subsequently,
the sections were incubated for 2 h at room temperature with
anti-mouse (cat. no. BM3895) and anti-rabbit (cat. no. BM3894)
horseradish peroxidase-conjugated goat secondary antibodies (both
1:500; Boster Biological Technology). Samples were then stained
with 3,3′-diaminobenzidine and hematoxylin at room temperature for
1 min, after which a light orthophoto microscope (magnification,
×200 and ×400) was used to obtain images. Finally, the protein
expression levels of the target genes were evaluated based on the
sum of the intensity score (0, no staining; 1, weakly positive; 2,
moderately positive; and 3, strongly positive) and the score for
the proportion of positive cells (0, <1; 1, 1–33; 2, 34–66; and
3, 67–100%) using Image-Pro Plus software (version 6.0; Media
Cybernetics, Inc.). Total scores of 0–2, 3–4 and 5–6 indicated low,
moderate and high expression, respectively. The difference in the
protein expression of tissues was determined using Fisher's exact
test combined with Bonferroni's correction test based on their
expression levels. P<0.05 was considered to indicate a
statistically significant difference.
Verification of the diagnostic value
of hub genes
The diagnostic value of hub genes for HPV-negative
and HPV-positive HNSCC was analyzed using a receiver operating
characteristic (ROC) curve analysis using the IHC-based protein
levels. The protein expression scores of the HNSCC and normal
tissues were imported into SPSS version 20.0 (IBM Corp.), after
which ROC curve analysis was performed. An area under the curve
(AUC) value of >0.7 was considered to indicate a high diagnostic
value.
Results
WGCNA
The gene expression profiles of 54 HPV-negative
HNSCC tissues (with negative ISH and P16 staining) were downloaded
from the TCGA database, as well as corresponding clinical
characteristic data. The sample dendrogram revealed that there were
no outliers; similarly, the trait heatmap showed that clinical data
of most patients in TCGA database were completely documented
(Fig. 1). Therefore, after removing
the genes with low expression, the gene expression profiles of all
54 HPV-negative HNSCC tissues were used in the WGCNA. When the soft
power β was set as 7, the scale independence of the topology
network reached >0.85 (Fig. 2A)
with the mean connectivity close to 0 (Fig. 2B). Therefore, a soft power of β=7 was
selected as the soft threshold for performing subsequent analyses.
The results revealed that when β=7, the topological overlap matrix
was able to meet the scale-free topology criterion with
R2=0.87 (Fig. 2C). The
results identified 27 gene co-expression modules (paleturquoise,
cyan, salmon, sienna3, violet, royalblue, saddlebrown, darkmagenta,
brown, black, purple, lightyellow, darkolivegreen, grey60, yellow,
darkgreen, skyblue, lightcyan, lightgreen, tan, darkturquoise,
darkorange, blue, green, steelblue, darkgrey and midnightblue),
while the genes that were not co-expressed were clustered in the
grey module (Fig. 2D). The 27 gene
co-expression modules were then used for further analysis.
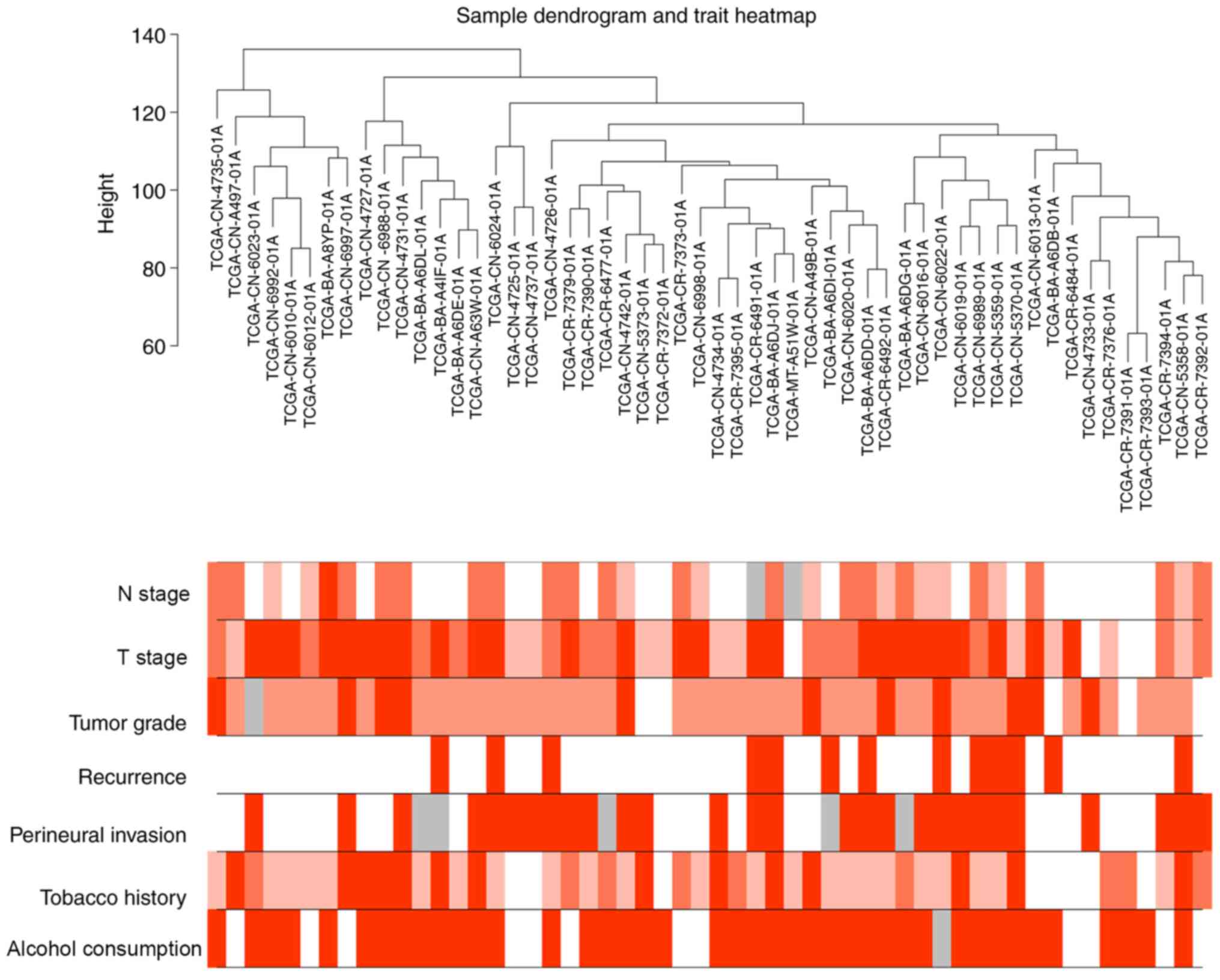 | Figure 1.Sample dendrogram of 54 human
papillomavirus-negative head and neck squamous cell carcinoma
tissues and the corresponding clinical characteristics of patients,
including N stage, T stage, tumor grade, recurrence, perineural
invasion, tobacco use history and alcohol consumption history. Red,
white and grey indicate high, low and missing values, respectively.
N, node; T, tumor. |
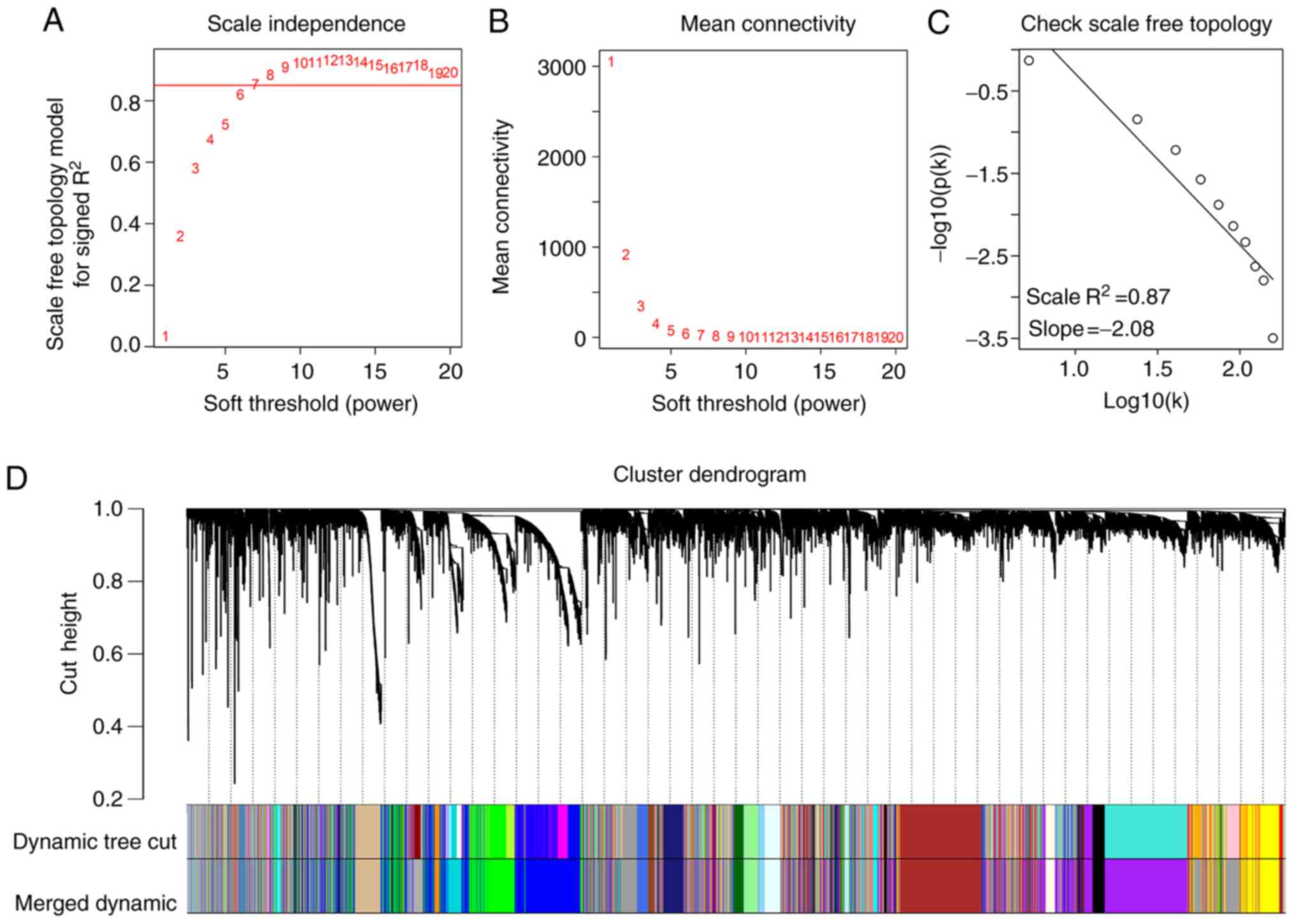 | Figure 2.Weighted gene co-expression network
analysis. (A) Analysis of the scale-free fit index for various β
soft-thresholding powers. (B) Analysis of the mean connectivity for
various soft-thresholding powers. (C) Scale-free topology when β=7.
(D) Dendrogram of all genes clustered based on a dissimilarity
measure (1-topological overlap matrix). There were 27 gene
co-expression modules in total (paleturquoise, cyan, salmon,
sienna3, violet, royalblue, saddlebrown, darkmagenta, brown, black,
purple, lightyellow, darkolivegreen, grey60, yellow, darkgreen,
skyblue, lightcyan, lightgreen, tan, darkturquoise, darkorange,
blue, green, steelblue, darkgrey and midnightblue). Genes without
co-expression were clustered into the grey module. |
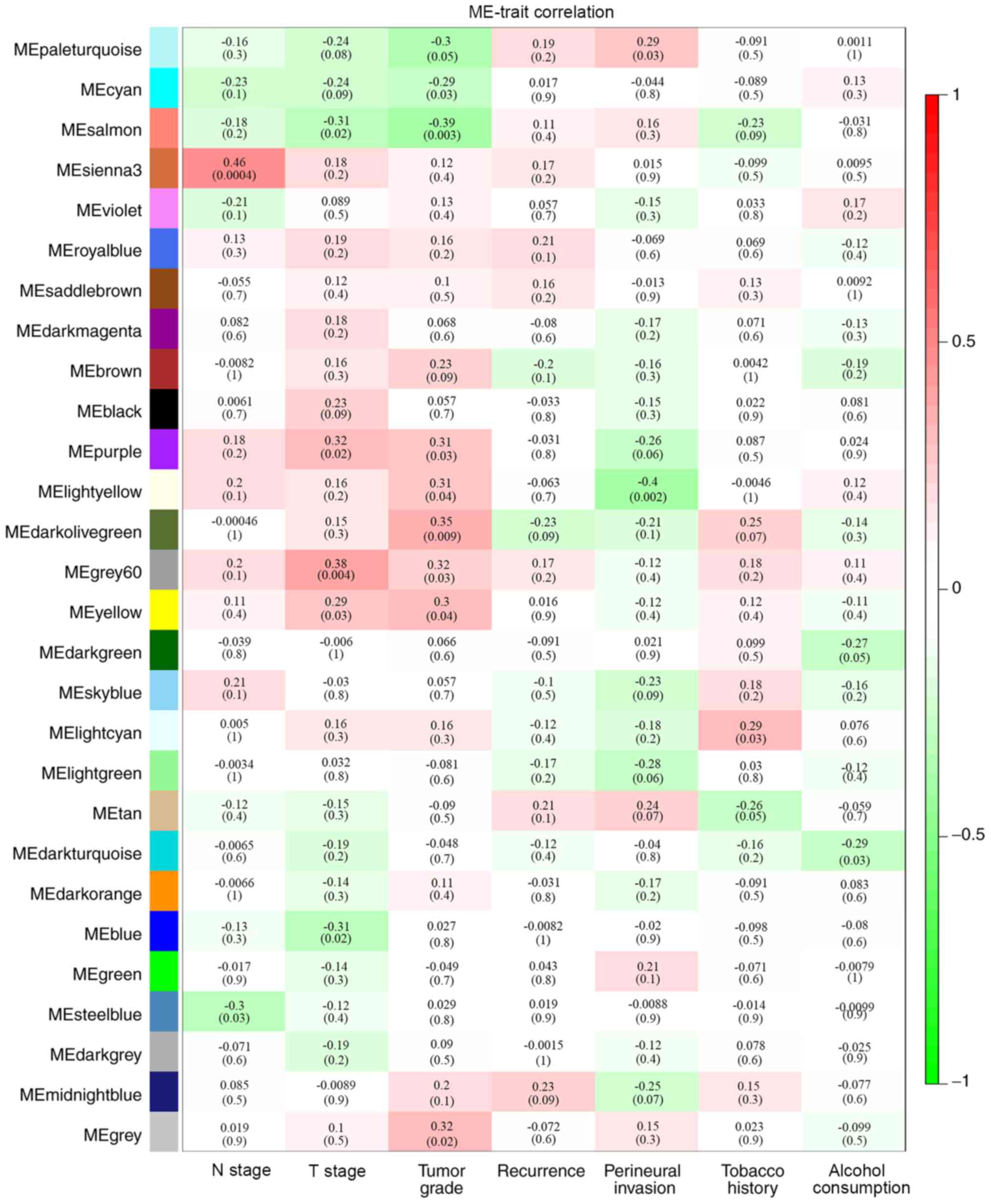
Identification of clinically
significant modules
By analyzing the correlation between the gene
co-expression modules and clinical characteristics, it was
determined that four gene co-expression modules were simultaneously
and significantly correlated with two clinical characteristics in
patients with HPV-negative HNSCC. The gene of the grey60 module was
positively correlated with T stage (R=0.38; P=0.004) and tumor
grade (R=0.32; P=0.03). Genes of the lightyellow module were
positively correlated with tumor grade (R=0.31; P=0.04) and
negatively correlated with perineural invasion (R=−0.4; P=0.002).
Additionally, genes in the purple module were positively correlated
with T stage (R=0.32; P=0.02) and tumor grade (R=0.31; P=0.03),
while genes of the salmon module were negatively correlated with T
stage (R=−0.31; P=0.02) and tumor stage (R=−0.39; P=0.003)
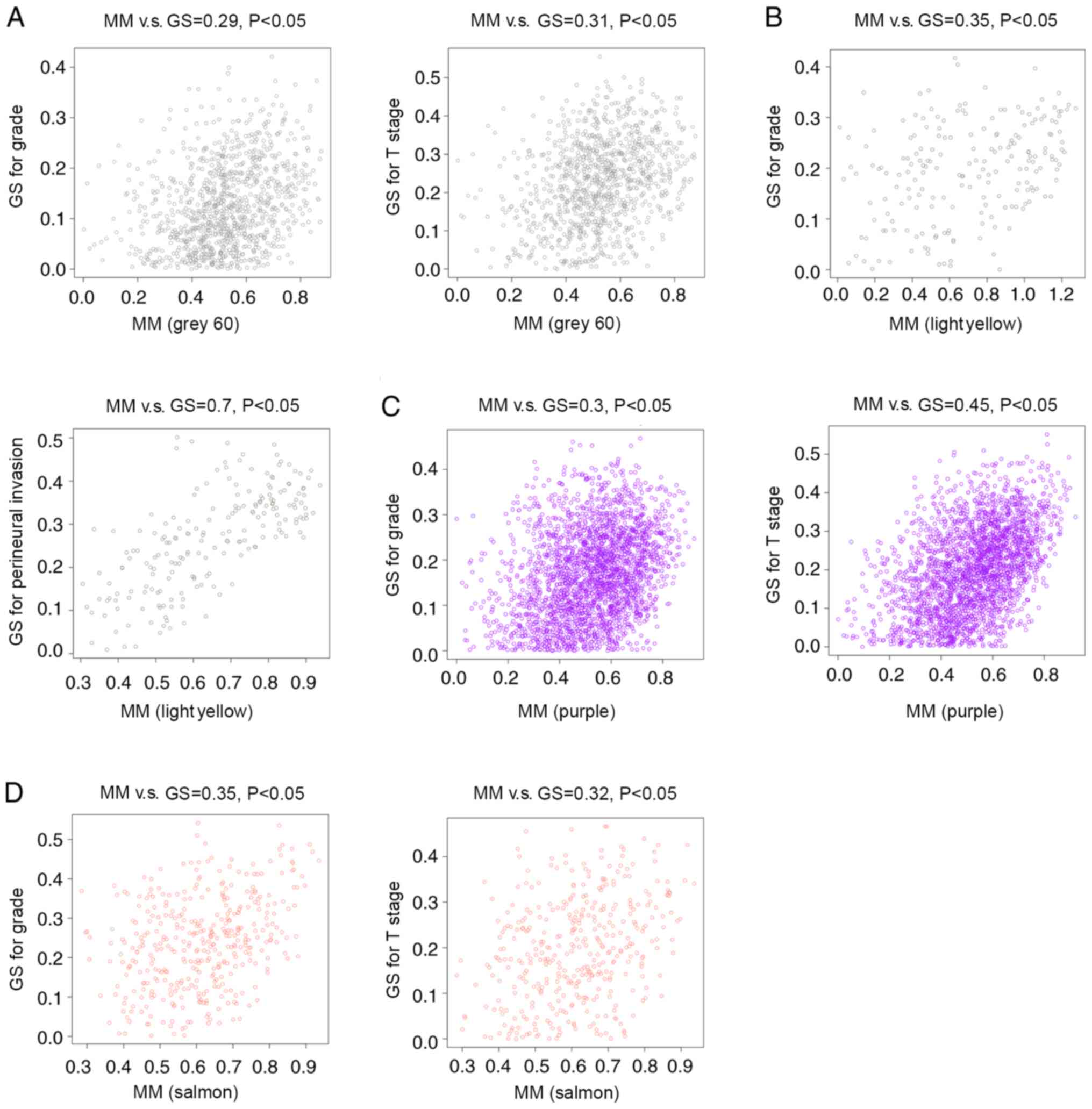
(Fig. 3). Correlations between GS
and MM were subsequently calculated in the four aforementioned
modules. The MM of the grey60 module was correlated with the GS for
tumor grade (correlation=0.29; P<0.05) and T stage
(correlation=0.31; P<0.05) (Fig.
4A). The MM of the lightyellow module was correlated with the
GS for tumor grade (correlation=0.35; P<0.05) and perineural
invasion (correlation=0.7; P<0.05) (Fig. 4B). The MM of the purple module was
correlated with the GS for tumor grade (correlation=0.3; P<0.05)
and T stage (correlation=0.45; P<0.05) (Fig. 4C), while the MM of the salmon module
was correlated with the GS for tumor grade (correlation=0.35;
P<0.05) and T stage (correlation=0.32; P<0.05) (Fig. 4D). Therefore, these four modules
(grey60, lightyellow, purple and salmon modules) were set as key
gene modules and were analyzed further.
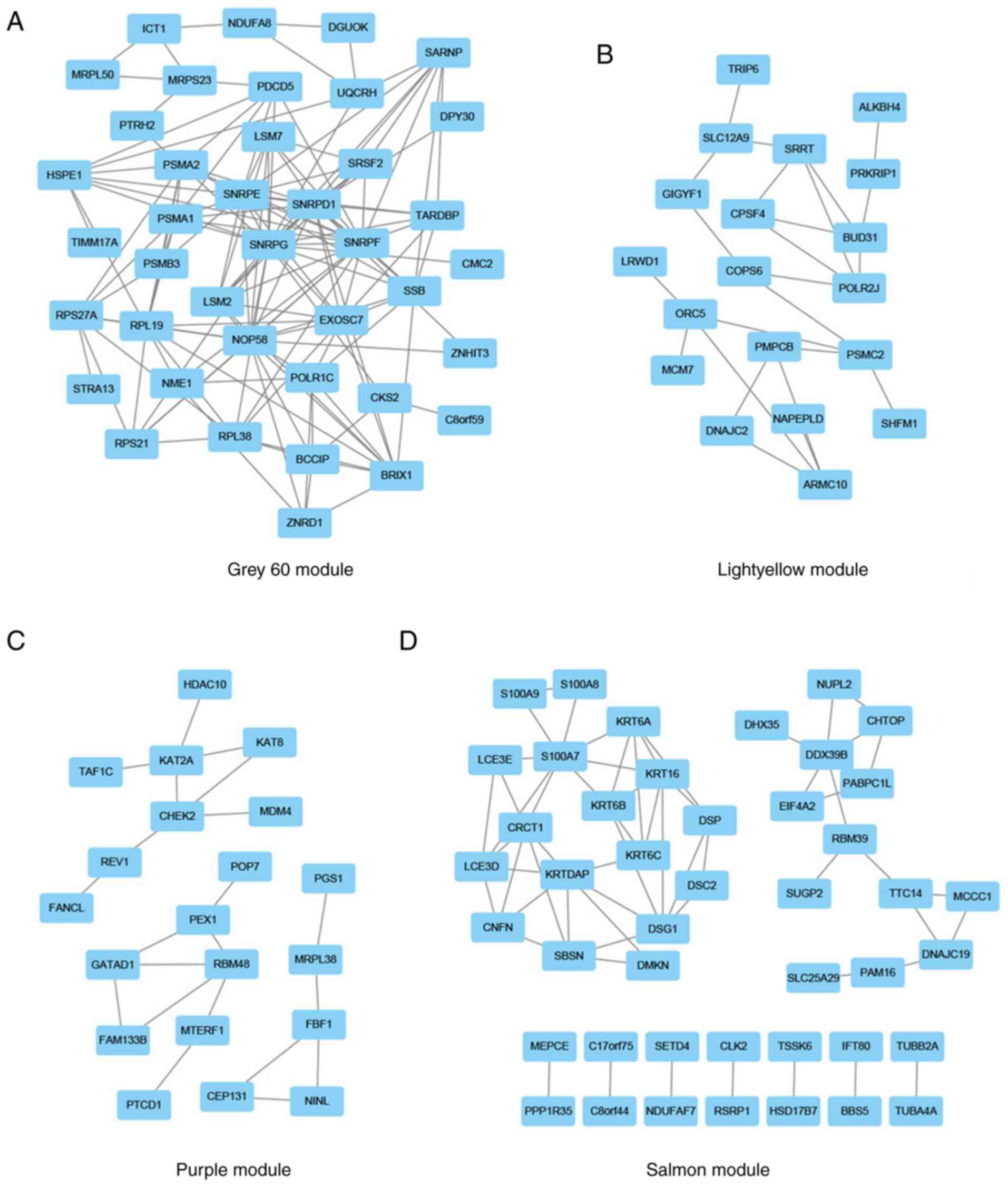
Construction of the PPI network and
selection of candidate hub genes
The lightyellow, grey60, purple and salmon modules
included 201, 997, 2,538 and 389 genes, respectively. After
importing these genes into the STRING database and removing the
nodes without connections, genes with a degree score in the top 10%
in the grey60 (Fig. 5A), lightyellow
(Fig. 5B), purple (Fig. 5C) and salmon (Fig. 5D) modules were visualized using
Cytoscape. The genes with a degree score in the top 1% in each
module are presented in Table II.
The resultant 17 genes were considered candidate hub genes and may
serve key roles in the development of HPV-negative HNSCC.
| Table II.Genes with a degree score in the top
1% in each module. |
Table II.
Genes with a degree score in the top
1% in each module.
| Module | Gene | Degree score |
|---|
| Grey60 module | SNRPG | 20 |
|
| SNRPF | 18 |
|
| SNRPE | 16 |
|
| SNRPD1 | 16 |
|
| NOP58 | 16 |
| Salmon module | S100A7 | 8 |
|
| KRTDAP | 7 |
| Purple module | DDX39B | 5 |
|
| CHEK2 | 4 |
|
| KAT2A | 4 |
| Lightyellow
module | ORC5 | 4 |
|
| PSMC2 | 4 |
|
| SRRT | 4 |
|
| BUD31 | 4 |
|
| POLR2J | 4 |
|
| ARMC10 | 4 |
|
| RBM48 | 4 |
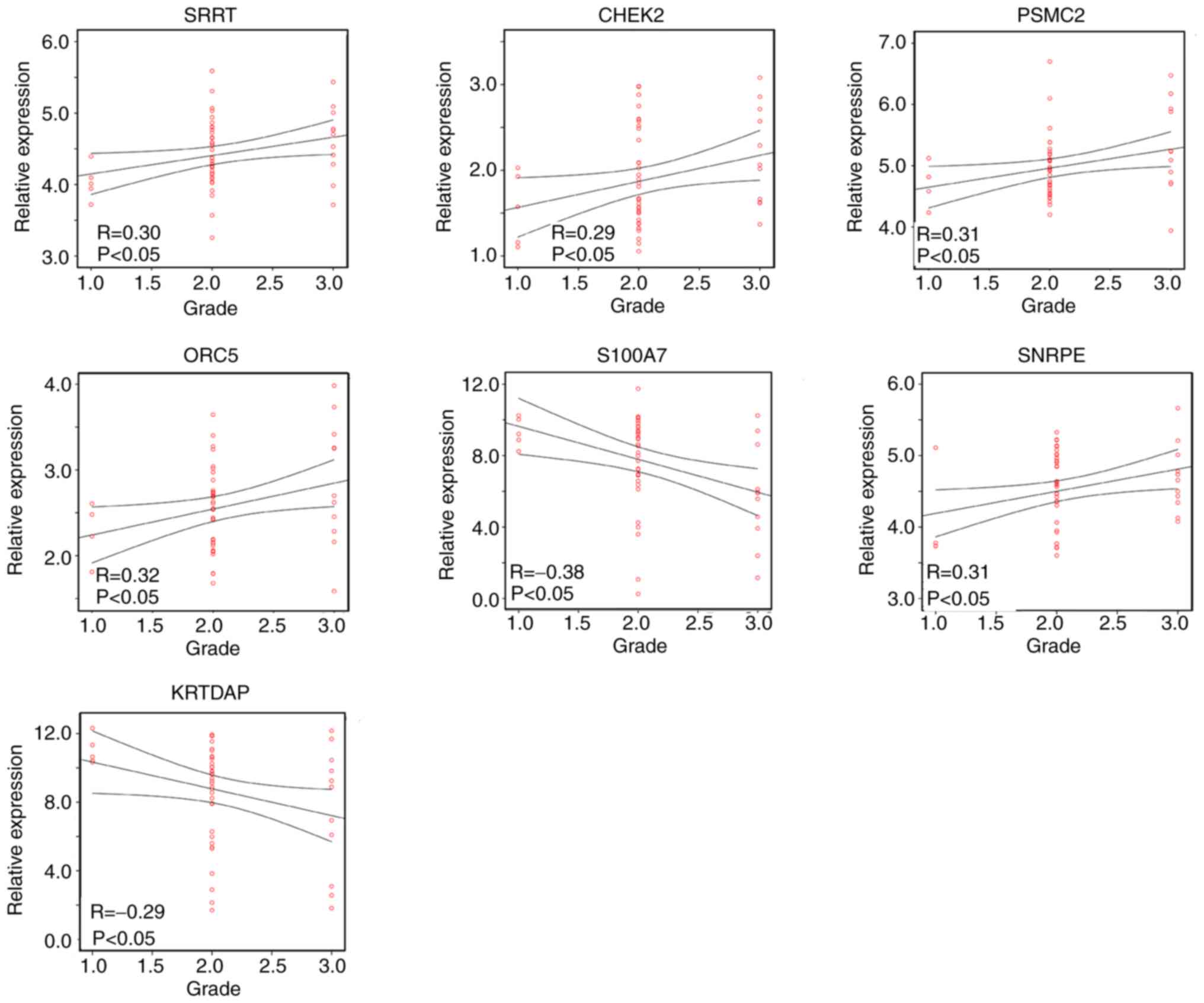
Pearson's correlation analysis between
the candidate hub genes and tumor grade
As the four gene modules were all correlated with
tumor grade, the correlation between candidate hub gene expression
and the tumor grade of HPV-negative HNSCC was further analyzed
using Pearson's correlation analysis. The results revealed that
SRRT (R=0.30; P<0.05), CHEK2 (R=0.29; P<0.05), SNRPE (R=0.31;
P<0.05), PSMC2 (R=0.31; P<0.05) and ORC5 (R=0.32; P<0.05)
were positively correlated with the tumor grade of HPV-negative
HNSCC tissue, while S100A7 (R=−0.38; P<0.05) and KRTDAP
(R=−0.29; P<0.05) were negatively correlated with tumor grade
(Fig. 6). Therefore, SRRT, CHEK2,
SNRPE, PSMC2, ORC5, S100A7 and KRTDAP were identified as hub genes
in HPV-negative HNSCC.
Identification of the expression
levels of hub genes in HPV-negative and HPV-positive HNSCC
To analyze the expression levels of hub genes in
HPV-negative and HPV-positive HNSCC, the merged gene data
expression profile of 103 HPV-positive HNSCC and 165 HPV-negative
HNSCC samples in TCGA and GEO databases (GSE117973, GSE85446 and
GSE112026) was used to perform DEGs analysis. The results revealed
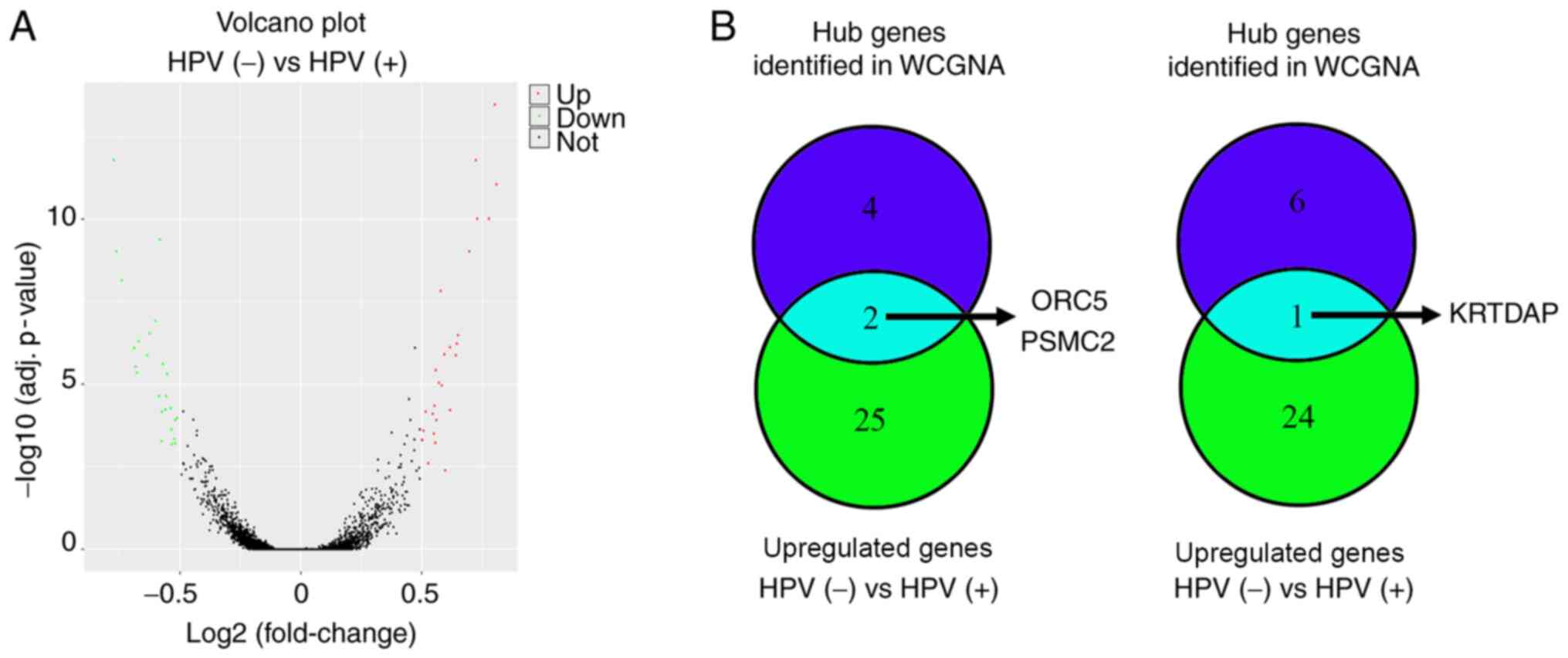
that there were 27 upregulated genes and 25 downregulated genes in
HPV-negative HNSCC compared with in HPV-positive HNSCC (Fig. 7A; Table
III). Through an intersection analysis, it was found that the
hub genes ORC5 and PSMC2 were upregulated in HPV-negative HNSCC
tissues compared with in HPV-positive HNSCC tissues, while KRTDAP
was downregulated (Fig. 7B).
| Table III.Detailed information of genes
differently expressed in HPV-negative and HPV-positive head and
neck squamous cell carcinoma according to the different expression
analysis. |
Table III.
Detailed information of genes
differently expressed in HPV-negative and HPV-positive head and
neck squamous cell carcinoma according to the different expression
analysis.
| Gene | Log
(fold-change) |
|---|
| DSG1 | 0.814689016 |
| SPRR2G | 0.807843405 |
| KLK5 | 0.78303004 |
| KRT1 | 0.735022158 |
| PSMC2 | 0.729090901 |
| MMP10 | 0.701782465 |
| ORC5 | 0.655262803 |
| SPRR2B | 0.650839232 |
| LCE3E | 0.646742549 |
| DSC1 | 0.622678824 |
| KLK7 | 0.622523674 |
| DEFB103B | 0.602302594 |
| PTHLH | 0.598655999 |
| MMP13 | 0.588604647 |
| MMP3 | 0.584023444 |
| FAM25A | 0.576520859 |
| ASPRV1 | 0.566257594 |
| LCE3D | 0.564179626 |
| CCNA1 | 0.561757208 |
| KLK8 | 0.558113758 |
| CPA4 | 0.555893249 |
| IGFL1 | 0.551152103 |
| CDSN | 0.532585037 |
| KLK6 | 0.52189567 |
| CDA | 0.512697702 |
| KRT75 | 0.507135684 |
| WFDC12 | 0.506385839 |
| CYP4X1 | −0.50678768 |
| SYNGR3 | −0.514548202 |
| RIBC2 | −0.514701295 |
| RNF212 | −0.517145557 |
| ABCA3 | −0.528349222 |
| TMPRSS2 | −0.530093548 |
| TMSB15A | −0.53215901 |
| KRTDAP | −0.54853466 |
| MEI1 | −0.551064139 |
| GABRP | −0.554796624 |
| CDKN2C | −0.565098291 |
| YBX2 | −0.569455778 |
| TCP11 | −0.570473276 |
| KRT19 | −0.578420257 |
| SMC1B | −0.580791027 |
| PODXL2 | −0.59651466 |
| PLAC8 | −0.620386125 |
| FAM3B | −0.630548652 |
| STAG3 | −0.665598925 |
| TAF7L | −0.672735588 |
| ZNF541 | −0.677964463 |
| KCNS1 | −0.683860665 |
| SYCP2 | −0.735795983 |
| NEFH | −0.757761211 |
| CDKN2A | −0.768176467 |
Verification of the protein expression
levels of hub genes in HPV-negative and HPV-positive HNSCC
tissues
To verify the bioinformatics results, IHC staining
of 27 HPV-negative and 35 HPV-positive HNSCC tissues was performed,
alongside their corresponding adjacent tissues. The results
revealed that the protein expression levels of SRRT, CHEK2 and
SNRPE were increased in HPV-negative HNSCC and HPV-positive HNSCC
tissues compared with in their adjacent counterparts; however,
there was no significant difference between HPV-negative and
HPV-positive HNSCC tissues. S100A7 expression was not significantly
different among HPV-negative HNSCC, HPV-positive HNSCC and adjacent
tissues. Furthermore, it was demonstrated that PSMC2 and ORC5
expression was significantly increased, while KRTDAP expression was
significantly decreased in HPV-negative HNSCC tissues compared with
in adjacent and HPV-positive HNSCC tissues (Table IV; Figs.
8 and S2).
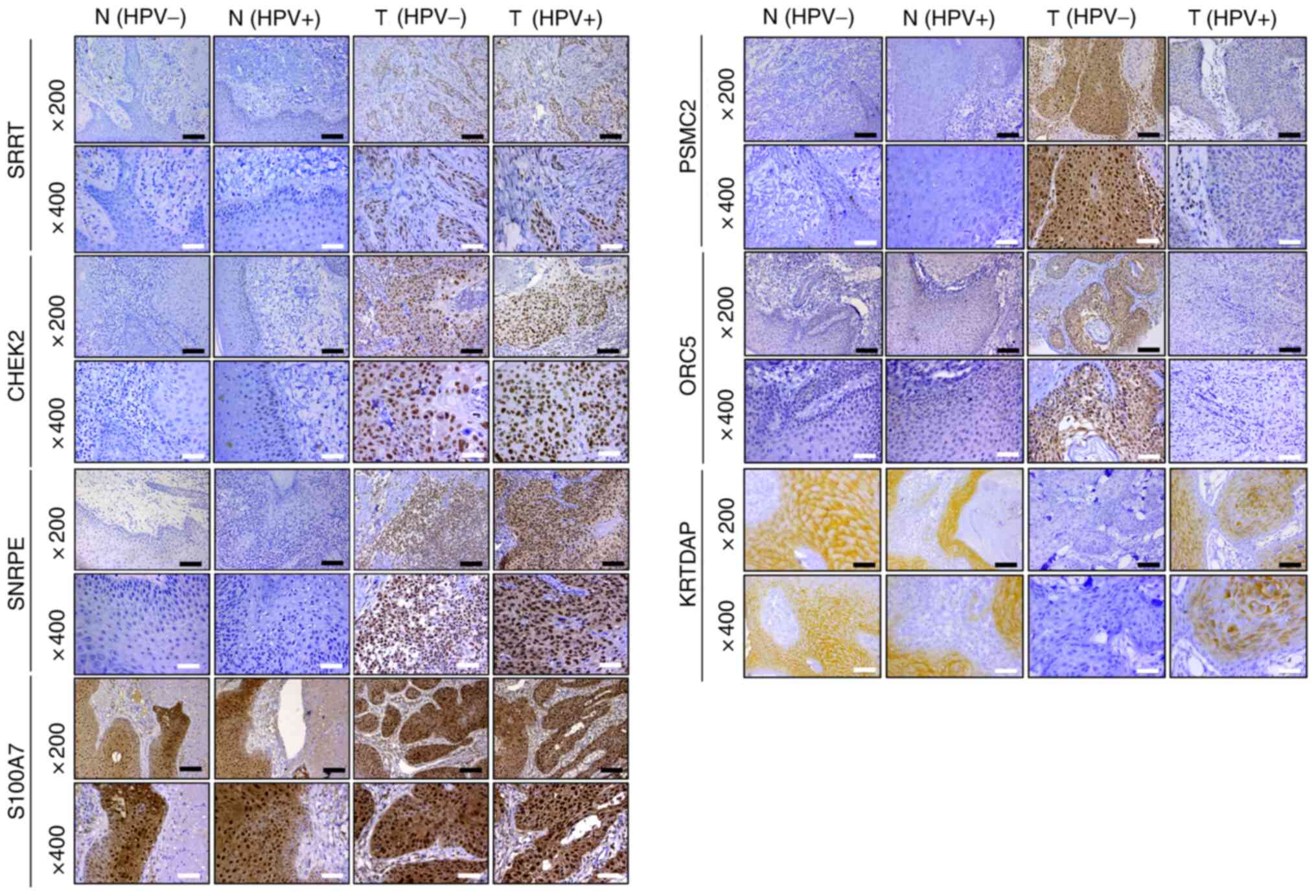 | Figure 8.Immunohistochemistry was used to
detect the protein expression levels of SRRT, CHEK2, SNRPE, PSMC2,
ORC5, S100A7 and KRTDAP in HPV-negative HNSCC, HPV-positive HNSCC
and adjacent tissues (magnification, ×200 and ×400). Black scale
bar, 100 µm; white scale bar, 50 µm. SRRT, serrate RNA effector
molecule; CHEK2, checkpoint kinase 2; SNRPE, small nuclear
ribonucleoprotein polypeptide E; PSMC2, proteasome 26S subunit
ATPase 2; ORC5, origin recognition complex subunit 5; S100A7, S100
calcium binding protein A7; KRTDAP, keratinocyte differentiation
associated protein; HPV, human papillomavirus; HNSCC, head and neck
squamous cell carcinoma; N, normal tissues; T, tumor tissues. |
| Table IV.Detailed immunohistochemical scoring
of SRRT, CHEK2, SNRPE, PSMC2, ORC5, S100A7 and KRTDAP expression in
HPV− HNSCC (n=27), HPV+ HNSCC (n=35) and
corresponding adjacent tissues. |
Table IV.
Detailed immunohistochemical scoring
of SRRT, CHEK2, SNRPE, PSMC2, ORC5, S100A7 and KRTDAP expression in
HPV− HNSCC (n=27), HPV+ HNSCC (n=35) and
corresponding adjacent tissues.
| Gene | Tissues | Low expression
(0–2), n | Medium expression
(3–4), n | High expression
(5–6), n |
|---|
| SRRT | Tumor
(HPV+)a | 5 | 4 | 26 |
|
| Adjacent
(HPV+) | 27 | 6 | 2 |
|
| Tumor
(HPV−)b | 2 | 3 | 22 |
|
| Adjacent
(HPV−) | 18 | 6 | 3 |
| CHEK2 | Tumor
(HPV+)a | 3 | 3 | 29 |
|
| Adjacent
(HPV+) | 28 | 4 | 3 |
|
| Tumor
(HPV−)b | 5 | 4 | 18 |
|
| Adjacent
(HPV−) | 18 | 7 | 2 |
| PSMC2 | Tumor
(HPV+) | 15 | 13 | 7 |
|
| Adjacent
(HPV+) | 22 | 7 | 6 |
|
| Tumor
(HPV−)b,c | 2 | 3 | 22 |
|
| Adjacent
(HPV−) | 20 | 5 | 2 |
| OCR5 | Tumor
(HPV+) | 19 | 10 | 6 |
|
| Adjacent
(HPV+) | 24 | 9 | 2 |
|
| Tumor
(HPV−)b,c | 2 | 7 | 18 |
|
| Adjacent
(HPV−) | 19 | 5 | 3 |
| S100A7 | Tumor
(HPV+) | 6 | 5 | 24 |
|
| Adjacent
(HPV+) | 4 | 4 | 27 |
|
| Tumor
(HPV−) | 2 | 4 | 21 |
|
| Adjacent
(HPV−) | 4 | 5 | 18 |
| SNRPE | Tumor
(HPV+)a | 3 | 4 | 28 |
|
| Adjacent
(HPV+) | 26 | 3 | 6 |
|
| Tumor
(HPV−)b | 4 | 2 | 21 |
|
| Adjacent
(HPV−) | 17 | 6 | 4 |
| KRTDAP | Tumor
(HPV+) | 9 | 6 | 20 |
|
| Adjacent
(HPV+) | 8 | 8 | 19 |
|
| Tumor
(HPV−)b,c | 20 | 4 | 3 |
|
| Adjacent
(HPV−) | 4 | 7 | 16 |
Verification of the diagnostic value
of hub genes for HPV-negative and HPV-positive HNSCC
Using protein level data obtained by IHC staining,
ROC curve analysis was performed. The results revealed that SRRT,
CHEK2, SNRPE, PSMC2, ORC5 and KRTDAP exhibited high diagnostic
value for distinguishing between HPV-negative HNSCC and adjacent
tissues, while S100A7 did not exert this effect (Fig. 9A). However, only SRRT, CHEK2 and
SNRPE had high diagnostic value for distinguishing between
HPV-positive HNSCC and adjacent tissues (Fig. 9B). Thus, SRRT, CHEK2 and SNRPE may be
used as common biomarkers for both HPV-negative and HPV-positive
HNSCC, while PSMC2, ORC5 and KRTDAP may be specific biomarkers for
HPV-negative HNSCC only.
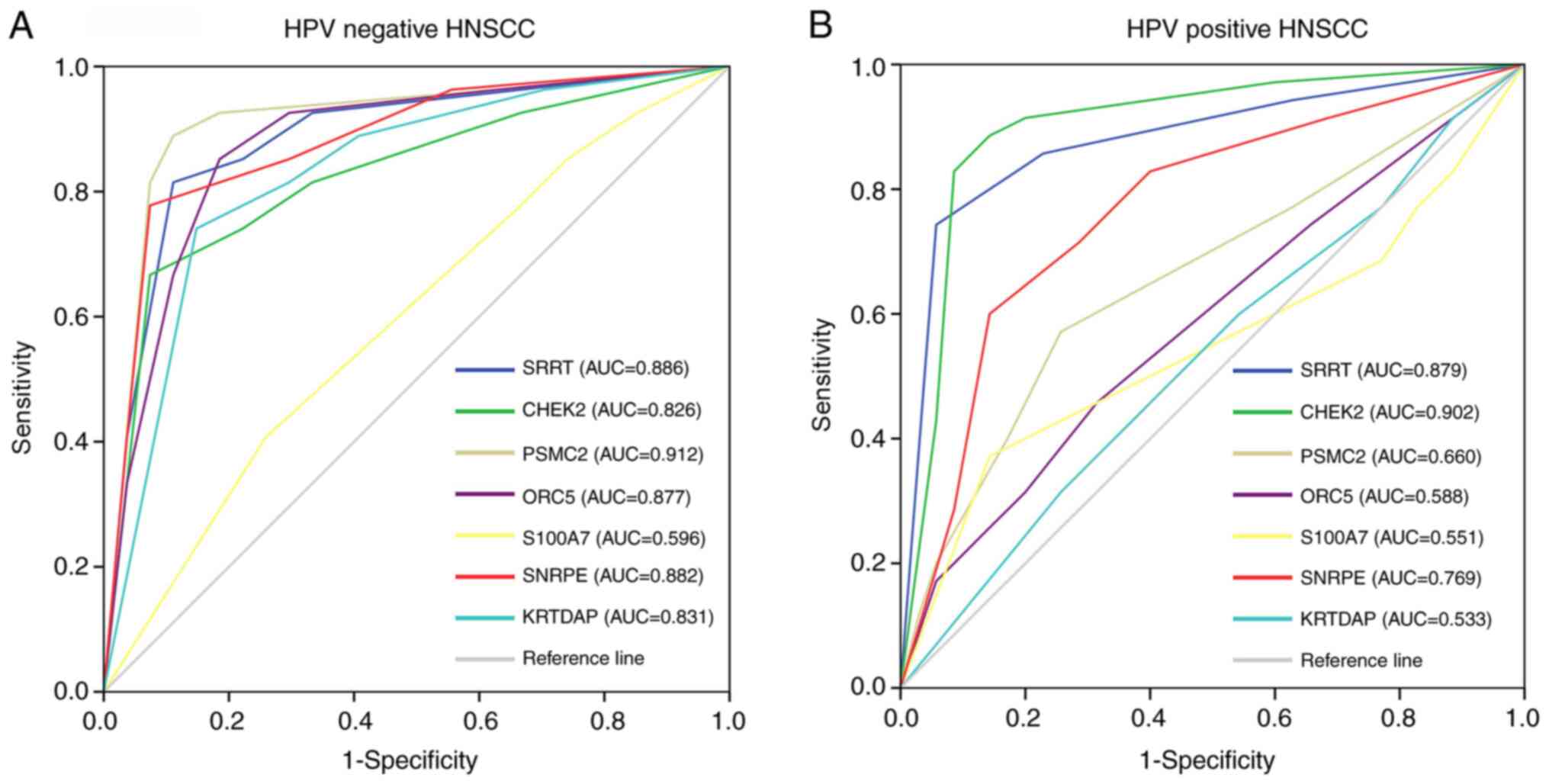 | Figure 9.Receiver operating characteristic
curve analysis was used to determine the diagnostic value of SRRT,
CHEK2, SNRPE, PSMC2, ORC5, S100A7 and KRTDAP for (A) HPV-negative
and (B) HPV-positive HNSCC based on protein level data obtained by
immunohistochemical staining. AUC >0.7 indicated genes with high
diagnostic value for distinguishing between HNSCC and adjacent
tissues. SRRT, serrate RNA effector molecule; CHEK2, checkpoint
kinase 2; SNRPE, small nuclear ribonucleoprotein polypeptide E;
PSMC2, proteasome 26S subunit ATPase 2; ORC5, origin recognition
complex subunit 5; S100A7, S100 calcium binding protein A7; KRTDAP,
keratinocyte differentiation associated protein; HPV, human
papillomavirus; HNSCC, head and neck squamous cell carcinoma; AUC,
area under the curve. |
Discussion
There are numerous risk factors involved in the
pathogenesis of HNSCC, including HPV infection. Although the
majority of HNSCC cases involve HPV infection, certain patients
develop HNSCC without it (17,18). The
molecular mechanisms underlying these two subtypes are different
(19). Furthermore, HPV-negative
HNSCC is more aggressive than HPV-positive HNSCC (20,21).
Therefore, the identification of novel and specific biomarkers for
HPV-negative HNSCC may improve the understanding of the specific
molecular mechanism associated with HPV-negative HNSCC, which may
improve the diagnosis and treatment of patients.
The current study identified four gene co-expression
modules associated with clinical characteristics of patients via
WGCNA. The results revealed 17 genes in these gene co-expression
modules that had a high degree score (top 1% of genes in each
module) in the PPI network. Additionally, seven of these genes
(SRRT, CHEK2, SNRPE, PSMC2, ORC5, S100A7 and KRTDAP) were
correlated with tumor grade in HPV-negative HNSCC. DEG analysis
demonstrated that PSMC2 and ORC5 expression was higher in
HPV-negative HNSCC compared with in HPV-positive HNSCC, while
KRTDAP expression was lower. Furthermore, using IHC staining and
ROC curve analysis, it was revealed that SRRT, CHEK2, SNRPE, PSMC2,
ORC5 and KRTDAP were differentially expressed in HPV-negative HNSCC
tissues compared with in adjacent tissues and had high diagnostic
values for distinguishing between HPV-negative HNSCC tissues and
adjacent tissues. However, after performing additional IHC staining
and ROC curve analysis, it was demonstrated that SRRT, CHEK2 and
SNRPE were differentially expressed in HPV-positive HNSCC tissues
compared with in adjacent tissues, exhibiting a high diagnostic
value for distinguishing between HPV-positive HNSCC and adjacent
tissues. Thus, while SRRT, CHEK2 and SNRPE may be used as common
biomarkers for both HPV-negative and HPV-positive HNSCC, PSMC2,
ORC5 and KRTDAP may be specific to HPV-negative HNSCC only.
SRRT (also named Ars2) was first isolated from a
hamster cell line and serves a key role in sodium arsenite
resistance (22). Furthermore, it
has been demonstrated that SRRT is involved in the biosynthesis of
certain microRNAs (23). However,
research on the role of SRRT in the development of cancer is
conflicting. It has been demonstrated that SRRT serves as an
oncogene in glioblastoma and promotes the proliferation and
migration of LN-229 and U87 glioblastoma cells via the MAPK/ERK
signaling pathway (24).
Additionally, SRRT expression is upregulated in cholangiocarcinoma
and associated with poor outcomes (25). However, pediatric acute lymphoblastic
leukemia involving low SRRT expression has poor chemotherapy
outcomes (26). CHEK2 was first
reported as an important breast cancer susceptibility gene; it
serves a key role in regulating cell apoptosis, cell cycle and DNA
repair (27). It increases the
stability of P53 and induces cell cycle arrest in the G1
phase (28). However, CHEK2
mutations are very common in several types of cancer, including
breast cancer (29), colorectal
cancer (30) and oral squamous cell
carcinoma (31). Additionally, CHEK2
promotes the progression of various types of cancer, including
hepatocellular carcinoma (32) and
colorectal cancer (33). SNRPE is a
member of a large family of polypeptides that are conserved in
eukaryotes and archaebacteria (34).
Previous studies have revealed that it is involved in RNA
processing and mRNA degradation (35,36).
Furthermore, a previous study has revealed that SNRPE is highly
expressed in high-grade prostate cancer, promoting cell
proliferation (37). In the present
study, it was demonstrated that SRRT, CHEK2 and SNRPE were in
clinically significant gene modules, had high degree scores in the
PPI network, were associated with tumor grade and were highly
expressed in HPV-negative and HPV-positive HNSCC tissues.
Furthermore, it was revealed that SRRT, CHEK2 and SNRPE had a high
diagnostic value in distinguishing HNSCC and adjacent tissues.
Therefore, to the best of our knowledge, the present study was the
first to demonstrate that SRRT, CHEK2 and SNRPE may be used as
common biomarkers for both HPV-negative and HPV-positive HNSCC.
Targeting SRRT, CHEK2 and SNRPE may aid the therapy of patients
with HPV-negative and HPV-positive HNSCC. However, the mechanism of
SRRT, CHEK2 and SNRPE in the development of HNSCC requires further
study.
PSMC2 is a novel gene on chromosome 7q22.1-q22.3
that encodes a member of the 19S proteasome (38). Recently, PSMC2 was revealed as an
oncogene in several types of cancer, including pancreatic cancer
(39) and osteosarcoma (40). Similarly, high PSMC2 expression
predicts a poor prognosis in patients with osteosarcoma (41). ORC5 is a key member of the origin
recognition complex that binds to replication origins, initiating
replication itself (42).
Additionally, it induces large-scale chromatin decondensation and
regulates the cell cycle and proliferation (43). KRTDAP acts as a soluble regulator of
keratinocyte differentiation and serves a key role in embryonic
skin morphogenesis (44). Research
has revealed that KRTDAP mRNA is upregulated in HPV-positive HNSCC
tissues compared with in adjacent tissues (45). In the present study, it was
demonstrated that PSMC2 and ORC5 expression was significantly
increased in HPV-negative HNSCC tissues compared with in
corresponding adjacent and HPV-positive HNSCC tissues.
Additionally, KRTDAP was significantly decreased in HPV-negative
HNSCC tissues compared with in corresponding adjacent and
HPV-positive HNSCC tissues. Similarly, it was revealed that PSMC2,
ORC5 and KRTDAP had a high diagnostic value in distinguishing
HPV-negative HNSCC and adjacent tissues. However, it could not
distinguish between HPV-positive HNSCC and adjacent tissues.
Therefore, it was hypothesized that PSMC2, ORC5 and KRTDAP may be
used as specific biomarkers for patients with HPV-negative HNSCC
only. It is well known that HPV-negative HNSCC is more aggressive
than HPV-positive HNSCC, so the downregulation of KRTDAP and
upregulation of PSMC2 and ORC5 may be key for HPV-negative HNSCC
cells to become aggressive. Targeting ORC5, PSMC and KRTDAP may
therefore help the individualized treatment of patients with
HPV-negative HNSCC. However, additional experiments should be
performed to verify this hypothesis.
In conclusion, based on WGCNA, PPI network,
Pearson's correlation, DEG, IHC and ROC curve analyses, SRRT, CHEK2
and SNRPE were revealed to be common biomarkers for both
HPV-negative and HPV-positive HNSCC, while PSMC2, ORC5 and KRTDAP
may be potential specific biomarkers for HPV-negative HNSCC. The
results may improve the understanding of HNSCC, as well as
providing potential biomarkers and targets for the diagnosis and
treatment of patients with this disease.
Supplementary Material
Supporting Data
Supporting Data
Acknowledgements
Not applicable.
Funding
The current study was supported by Guizhou Big Data
Health Management Industry Technology Innovation Strategic Alliance
[grant no. Qianketong (2016)193].
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request, and the GSE117973, GSE85446 and GSE112026 datasets
analyzed during the current study are available in the Gene
Expression Omnibus repository (https://www.ncbi.nlm.nih.gov/gds).
Authors' contributions
YS, YC and ZZ were responsible for the experimental
design, analysis and interpretation of data. DR, YY and BW
performed the experiments. YS and YC were responsible for
confirming the authenticity of the data. All authors took part in
drafting the article and revising it critically for important
intellectual content, agreed to be accountable for all aspects of
the work, and read and approved the final manuscript.
Ethics approval and consent to
participate
The current study was approved by the Human Trait
Ethics Committee of Guizhou Medical University (Guiyang, China) and
performed in accordance with the Declaration of Helsinki. All
patients from which samples were obtained provided written informed
consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Johnson DE, Burtness B, Leemans CR, Lui
VWY, Bauman JE and Grandis JR: Head and neck squamous cell
carcinoma. Nat Rev Dis Primers. 6:922020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dhull AK, Atri R, Dhankhar R, Chauhan AK
and Kaushal V: Major risk factors in head and neck cancer: A
retrospective analysis of 12-year experiences. World J Oncol.
9:80–84. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Plzák J, Bouček J, Bandúrová V, Kolář M,
Hradilová M, Szabo P, Lacina L, Chovanec M and Smetana K Jr: The
head and neck squamous cell carcinoma microenvironment as a
potential target for cancer therapy. Cancers (Basel). 11:4402019.
View Article : Google Scholar
|
|
4
|
Sabatini ME and Chiocca S: Human
papillomavirus as a driver of head and neck cancers. Br J Cancer.
122:306–314. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Viros Porcuna D, Pollan Guisasola C, Viña
Soria C, Cirauqui Cirauqui B, Pardo Muñoz L, Collurá F and Mesia
Nin R: Transoral robotic surgery for squamous cell carcinoma of the
oropharynx in a primarily human papillomavirus-negative patient
population. Clin Transl Oncol. 22:1303–1311. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shen Y, Liu J, Zhang L, Dong S, Zhang J,
Liu Y, Zhou H and Dong W: Identification of potential biomarkers
and survival analysis for head and neck squamous cell carcinoma
using bioinformatics strategy: A study based on TCGA and GEO
datasets. Biomed Res Int. 2019:73760342019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yang B, Fu L, Xu S, Xiao J, Li Z and Liu
Y: A nomogram based on a gene signature for predicting the
prognosis of patients with head and neck squamous cell carcinoma.
Int J Biol Markers. 34:309–317. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Langfelder P and Horvath S: WGCNA: An R
package for weighted correlation network analysis. BMC
Bioinformatics. 9:5592008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Song Y, Pan Y and Liu J: The relevance
between the immune response-related gene module and clinical traits
in head and neck squamous cell carcinoma. Cancer Manag Res.
11:7455–7472. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang Z, Liu R, Jin R, Fan Y, Li T, Shuai
Y, Li X, Wang X and Luo J: Integrating clinical and genetic
analysis of perineural invasion in head and neck squamous cell
carcinoma. Front Oncol. 9:4342019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chi J, Preeshagul IR, Sheikh-Fayyaz S,
Teckie S, Kohn N, Ziemba Y, Laser A, Frank D, Ghaly M, Kamdar D, et
al: Evaluating of HPV-DNA ISH as an adjunct to p16 testing in
oropharyngeal cancer. Future Sci OA. 6:FSO6062020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lydiatt WM, Patel SG, O'Sullivan B,
Brandwein MS, Ridge JA, Migliacci JC, Loomis AM and Shah JP: Head
and neck cancers-major changes in the American joint committee on
cancer eighth edition cancer staging manual. CA Cancer J Clin.
67:122–137. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bi N, Sun Y, Lei S, Zeng Z, Zhang Y, Sun C
and Yu C: Identification of 40S ribosomal protein S8 as a novel
biomarker for alcohol-associated hepatocellular carcinoma using
weighted gene co-expression network analysis. Oncol Rep.
44:611–627. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Feng B, Shen Y, Pastor Hostench X, Bieg M,
Plath M, Ishaque N, Eils R, Freier K, Weichert W, Zaoui K and Hess
J: Integrative analysis of multi-omics data identified EGFR and
PTGS2 as key nodes in a gene regulatory network related to immune
phenotypes in head and neck cancer. Clin Cancer Res. 26:3616–3628.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mes SW, Te Beest D, Poli T, Rossi S,
Scheckenbach K, van Wieringen WN, Brink A, Bertani N, Lanfranco D,
Silini EM, et al: Prognostic modeling of oral cancer by gene
profiles and clinicopathological co-variables. Oncotarget.
8:59312–59323. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kelley DZ, Flam EL, Izumchenko E, Danilova
LV, Wulf HA, Guo T, Singman DA, Afsari B, Skaist AM, Considine M,
et al: Integrated analysis of whole-genome ChIP-Seq and RNA-Seq
data of primary head and neck tumor samples associates HPV
integration sites with open chromatin marks. Cancer Res.
77:6538–6550. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jethwa AR and Khariwala SS:
Tobacco-related carcinogenesis in head and neck cancer. Cancer
Metastasis Rev. 36:411–423. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Paver EC, Currie AM, Gupta R and Dahlstrom
JE: Human papilloma virus related squamous cell carcinomas of the
head and neck: Diagnosis, clinical implications and detection of
HPV. Pathology. 52:179–191. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Koneva LA, Zhang Y, Virani S, Hall PB,
McHugh JB, Chepeha DB, Wolf GT, Carey TE, Rozek LS and Sartor MA:
HPV Integration in HNSCC correlates with survival outcomes, immune
response signatures, and candidate drivers. Mol Cancer Res.
16:90–102. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Qian X, Nguyen DT, Dong Y, Sinikovic B,
Kaufmann AM, Myers JN, Albers AE and Graviss EA: Prognostic score
predicts survival in HPV-negative head and neck squamous cell
cancer patients. Int J Biol Sci. 15:1336–1344. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Vossen DM, Verhagen CVM, van der Heijden
M, Essers PBM, Bartelink H, Verheij M, Wessels LFA, van den Brekel
MWM and Vens C: Genetic factors associated with a poor outcome in
head and neck cancer patients receiving definitive
chemoradiotherapy. Cancers (Basel). 11:4452019. View Article : Google Scholar
|
|
22
|
Elahi S, Egan SM, Holling GA, Kandefer RL,
Nemeth MJ and Olejniczak SH: The RNA binding protein Ars2 supports
hematopoiesis at multiple levels. Exp Hematol. 64:45–58.e9. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gruber JJ, Olejniczak SH, Yong J, La Rocca
G, Dreyfuss G and Thompson CB: Ars2 promotes proper
replication-dependent histone mRNA 3′ end formation. Mol Cell.
45:87–98. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ke XX, Pang Y, Chen K, Zhang D, Wang F,
Zhu S, Mao J, Hu X, Zhang G and Cui H: Knockdown of arsenic
resistance protein 2 inhibits human glioblastoma cell proliferation
through the MAPK/ERK pathway. Oncol Rep. 40:3313–3322.
2018.PubMed/NCBI
|
|
25
|
He Q, Cai L, Shuai L, Li D, Wang C, Liu Y,
Li X, Li Z and Wang S: Ars2 is overexpressed in human
cholangiocarcinomas and its depletion increases PTEN and PDCD4 by
decreasing microRNA-21. Mol Carcinog. 52:286–296. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cui L, Gao C, Zhang RD, Jiao Y, Li WJ,
Zhao XX, Liu SG, Yue ZX, Zheng HY, Deng GR, et al: Low expressions
of ARS2 and CASP8AP2 predict relapse and poor prognosis in
pediatric acute lymphoblastic leukemia patients treated on China
CCLG-ALL 2008 protocol. Leuk Res. 39:115–123. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ansari N, Shahrabi S, Khosravi A, Shirzad
R and Rezaeean H: Prognostic significance of CHEK2 mutation in
progression of breast cancer. Lab Med. 50:e36–e41. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Luo Q, Guo H, Kuang P, Cui H, Deng H, Liu
H, Lu Y, Wei Q, Chen L, Fang J, et al: Sodium fluoride arrests
renal G2/M phase cell-cycle progression by activating
ATM-Chk2-P53/Cdc25C signaling pathway in mice. Cell Physiol
Biochem. 51:2421–2433. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Apostolou P and Papasotiriou I: Current
perspectives on CHEK2 mutations in breast cancer. Breast Cancer
(Dove Med Press). 9:331–335. 2017.PubMed/NCBI
|
|
30
|
Wang W, Guo M, Xia X, Zhang C, Zeng Y and
Wu S: XRRA1 targets ATM/CHK1/2-mediated DNA repair in colorectal
cancer. Biomed Res Int. 2017:57189682017.PubMed/NCBI
|
|
31
|
Yoon AJ, Shen J, Santella RM, Zegarelli
DJ, Chen R and Weinstein IB: Activated checkpoint kinase 2
expression and risk for oral squamous cell carcinoma. Cancer
Epidemiol Biomarkers Prev. 16:2768–2772. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Carloni V, Lulli M, Madiai S, Mello T,
Hall A, Luong TV, Pinzani M, Rombouts K and Galli A: CHK2
overexpression and mislocalisation within mitotic structures
enhances chromosomal instability and hepatocellular carcinoma
progression. Gut. 67:348–361. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Advani SJ, Camargo MF, Seguin L, Mielgo A,
Anand S, Hicks AM, Aguilera J, Franovic A, Weis SM and Cheresh DA:
Kinase-independent role for CRAF-driving tumour radioresistance via
CHK2. Nat Commun. 6:81542015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Quidville V, Alsafadi S, Goubar A, Commo
F, Scott V, Pioche-Durieu C, Girault I, Baconnais S, Le Cam E,
Lazar V, et al: Targeting the deregulated spliceosome core
machinery in cancer cells triggers mTOR blockade and autophagy.
Cancer Res. 73:2247–2258. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Valles I, Pajares MJ, Segura V, Guruceaga
E, Gomez-Roman J, Blanco D, Tamura A, Montuenga LM and Pio R:
Identification of novel deregulated RNA metabolism-related genes in
non-small cell lung cancer. PLoS One. 7:e420862012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chen T, Zhang B, Ziegenhals T, Prusty AB,
Fröhler S, Grimm C, Hu Y, Schaefke B, Fang L, Zhang M, et al: A
missense mutation in SNRPE linked to non-syndromal microcephaly
interferes with U snRNP assembly and pre-mRNA splicing. PLoS Genet.
15:e10084602019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Anchi T, Tamura K, Furihata M, Satake H,
Sakoda H, Kawada C, Kamei M, Shimamoto T, Fukuhara H, Fukata S, et
al: SNRPE is involved in cell proliferation and progression of
high-grade prostate cancer through the regulation of androgen
receptor expression. Oncol Lett. 3:264–268. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
He J, Xing J, Yang X, Zhang C, Zhang Y,
Wang H, Xu X, Wang H, Cao Y, Xu H, et al: Silencing of proteasome
26S subunit ATPase 2 regulates colorectal cancer cell
proliferation, apoptosis, and migration. Chemotherapy. 64:146–154.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Qin J, Wang W, An F, Huang W and Ding J:
PSMC2 is up-regulated in pancreatic cancer and promotes cancer cell
proliferation and inhibits apoptosis. J Cancer. 10:4939–4946. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Song M, Wang Y, Zhang Z and Wang S: PSMC2
is up-regulated in osteosarcoma and regulates osteosarcoma cell
proliferation, apoptosis and migration. Oncotarget. 8:933–953.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Li GW and Yan X: Lower miR-630 expression
predicts poor prognosis of osteosarcoma and promotes cell
proliferation, migration and invasion by targeting PSMC2. Eur Rev
Med Pharmacol Sci. 23:1915–1925. 2019.PubMed/NCBI
|
|
42
|
Shibata E and Dutta A: A human cancer cell
line initiates DNA replication normally in the absence of ORC5 and
ORC2 proteins. J Biol Chem. 295:16949–16959. 2020. View Article : Google Scholar
|
|
43
|
Giri S, Chakraborty A, Sathyan KM,
Prasanth KV and Prasanth SG: Orc5 induces large-scale chromatin
decondensation in a GCN5-dependent manner. J Cell Sci. 129:417–429.
2016. View Article : Google Scholar
|
|
44
|
Oomizu S, Sahuc F, Asahina K, Inamatsu M,
Matsuzaki T, Sasaki M, Obara M and Yoshizato K: Kdap, a novel gene
associated with the stratification of the epithelium. Gene.
256:19–27. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Thibodeau BJ, Geddes TJ, Fortier LE, Ahmed
S, Pruetz BL, Wobb J, Chen P, Wilson GD and Akervall JA: Gene
expression characterization of HPV positive head and neck cancer to
predict response to chemoradiation. Head Neck Pathol. 9:345–353.
2015. View Article : Google Scholar : PubMed/NCBI
|