Introduction
Pancreatic ductal adenocarcinoma (PDAC) is the most
common tumor subtype of pancreatic cancer accounting for ~85% of
cases (1), which is observed in the
digestive system. The global incidence rate of PDAC continues to
increase annually, due in part to its extreme aggressiveness and
early metastasis characteristics (2). The 5-year survival rate of patients
with PDAC is ~5% (3). The outcomes
of immunotherapy, surgery, chemotherapy and radiotherapy for PDAC
remain unsatisfactory, and treatment side effects have resulted in
patients experiencing a lower quality of life (4). Comprehensive genomic analysis of PDAC
has demonstrated that PDAC occurrence and development is closely
associated with multiple gene mutations and signal transduction
pathways, which are involved in the mutant genes (5). In addition to the four common cancer
mutant genes, KRAS, TP53, SMAD4 and CDKN2A, it also includes some
genes that have a mutation rate of ~10%, including KDM6A, RBM10 and
MLL3 (6). These mutant genes mainly
influence PDAC progression in DNA damage repair, cell cycle
regulation, chromatin regulation and TGF-β signaling pathway
(7). Thus, understanding the
underlying molecular mechanisms of PDAC is vital for improving
current therapeutic options and developing novel therapeutic
strategies.
Recently, notable progress has been made in
microarray technology and bioinformatics methods. Unlike
traditional experimental methods, where only either a single gene
or several genes can be studied at once, large-scale research and
analysis can be performed, and mutant genes can be screened out at
the genomic level, which provides a better method for the
identification of cancer biomarkers and the study of molecular
mechanisms (8).
In the present study, three microarray datasets,
GSE15471 (9), GSE16515 (10) and GSE28735 (11) were downloaded from the Gene
Expression Omnibus (GEO) database to identify differentially
expressed genes (DEGs) in PDAC tissues and adjacent normal tissues,
and five hub genes were further screened from the DEGs using
Cytoscape software. Gene Ontology (GO) and Kyoto Encyclopedia of
Genes and Genomes (KEGG) functional and signaling pathway
enrichment analyses of the DEGs were performed, and a
protein-protein interaction (PPI) network was constructed using the
Search Tool for the Retrieval of Interacting Genes/Proteins
(STRING) database to assess PDAC development at the molecular
level.
MicroRNAs (miRNAs/miRs) are non-coding
single-stranded RNA molecules that are 20–24 nucleotides in length,
which can be paired with the 3′-untranslated region of the target
gene to regulate its expression (12). miRNAs are involved in several
cellular biological processes, including cell proliferation,
migration, apoptosis, invasion, angiogenesis and immune escape, and
also participate in the occurrence and development of cancer
through regulation of the expression of oncogenes and antioncogenes
(13). Increasing evidence suggests
that specific cancer-related miRNAs that are detected in body
fluids, such as plasma, serum, urine and saliva can serve as
biomarkers for cancer diagnosis and prognosis (14,15).
In the present study, six mesenchymal-to-epithelial
transition (MET)-targeted miRNAs (miR-1-3p, miR-23b-3p, miR-31-5p,
miR-34a-5p, miR-340-5p and miR-130a-3p) were screened using the
DIANA, TargetScan, miRDB and miRTarBase databases, and the results
demonstrated that miR-1-3p, miR-23b-3p, miR-34a-5p and miR-130a-3p
have conserved binding sites with MET. Thus, a miRNA-mRNA signaling
pathway was assessed, which affects PDAC progression, thereby
providing potential biomarkers for the early diagnosis and
treatment of patients with PDAC.
Materials and methods
Microarray datasets
A total of three microarray datasets, GSE15471,
GSE16515 and GSE28735, were downloaded from the GEO database
(http://www.ncbi.nlm.nih.gov/geo), based
on the GPL570 Affymetrix Human Genome U133 Plus 2.0 Array and the
Affymetrix Human Gene 1.0 ST Array. The GSE15471 dataset contained
36 PDAC tissues and 36 normal tissues, the GSE16515 dataset
contained 36 PDAC tissues and 16 normal tissues, and the GSE28735
dataset contained 45 PDAC tissues and 36 normal tissues. The
datasets were screened for the following criteria: i) Samples
contained PDAC tissues and normal pancreatic tissues, ii) study
type was restricted to expression profiling by array, iii) organism
was restricted to homo sapiens and iv) original data were
accessible.
GO and KEGG functional and signaling
pathway enrichment analyses of the DEGs
GEO2R is an interactive web tool that allows users
to compare two or more databases in a GEO series, and screen DEGs
under different experimental conditions (8). Thus, the GEO2R database (http://www.ncbi.nlm.nih.gov/geo/geo2r)
was used to screen the DEGs between PADC tissues and normal
pancreatic tissues. The probe sets were converted into
corresponding gene symbols based on the annotation information in
the platform. Probe sets that did not have corresponding gene
symbols or genes with more than one probe set were removed or
averaged. |Log2 fold change| >1 and adjusted
P<0.01 were considered to indicate a statistically significant
difference.
The Database for Annotation, Visualization and
Integrated Discovery (DAVID; http://david.ncifcrf.gov) was used to perform GO and
KEGG functional and signaling pathway enrichment analyses of the
DEGs. GO analysis (http://www.geneontology.org/) is used to predict the
potential functions of target genes based on molecular function
(MF), cell component (CC) and biological processes (BP) (16). KEGG (http://www.genome.jp/kegg/pathway.html) is a database
that is associated with gene pathways and improves the
understanding of metabolic pathways where genes are involved, and
the associations between pathways by linking genomic information to
a higher-level system (17).
PPI network construction
Analysis of the interactions between proteins can
help uncover the molecular mechanisms of disease occurrence and
development (18). In the present
study, the STRING database (http://string-db.org) was used to construct the PPI
network of the DEGs, and interaction with a combined score >0.4
was considered to indicate statistical significance. The Cytoscape
software (version 3.6.1) (8) was
subsequently used to visualize the PPI network.
Selection and analysis of hub
genes
A total of five hub genes were selected from the
DEGs using MCODE plug-in (version 1.5.1) within the Cytoscape
software, and BiNGO plug-in (version 3.0.4) was subsequently used
to perform biological process analyses of the hub genes. The Gene
Expression Profiling Interactive Analysis (GEPIA) database
(http://gepia.cancer-pku.cn/detail.php) was used to
assess the differences in expression of the five hub genes in PDAC
tissues and normal pancreatic tissues. The Kaplan-Meier plotter
database (http://kmplot.com) was used to perform
disease-free survival and overall survival of the hub genes in
PDAC. The Oncomine database (http://www.oncomine.com) is used to compute gene
expression signatures, clusters, gene-set modules and automatically
extract biological insights from data (19). The Oncomine database was used in the
present study to analyze the DEGs between PDAC tissues and normal
pancreatic tissues, and detect the expression levels of the hub
gene MET in different types of tumors.
Cell culture and transfection
The PDAC cell line, PANC-1, was purchased from the
Cell Bank of Type Culture Collection of the Chinese Academy of.
Cells were maintained in DMEM (HyClone; Cytiva) supplemented with
10% fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.)
and 1% penicillin/streptomycin (cat. no. SV30010; Shanghai Canspec
Scientific & Technology Co., Ltd.), at 37°C with 5%
CO2. The medium was replaced every 2 days for 2
weeks.
Small interfering (si)RNAs were subsequently used to
silence MET expression and assess the effect of MET on PDAC cells.
Cells were seeded into 6-well plates at a density of
5×105 cells/ml and cultured until they reached 60–70%
confluence. Cells were transfected at 37°C for 48 h with 5 µl MET
siRNA (siRNA-MET) and 5 µl negative control (scrambled) siRNA
(siRNA-NC) using 10 µl Lipofectamine® 2000 reagent
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's instructions. The following sequences were used:
siRNA-MET sense, 5′-CACUAACUACAUUUAUGUUUU-3′ and antisense,
5′-AACAUAAAUGUAGUUAGUGGC-3′; siRNA-NC sense,
5′-AUAUAAUUCUACUUACUUGCU-3′, and antisense,
5′-CGAGUAAGUACAGUAAUGAUU-3′ (Shanghai GenePharma Co., Ltd.).
Non-transfected cells were used as the control group. Transfection
efficiency was assessed via reverse transcription-quantitative
(RT-q)PCR analysis.
RT-qPCR
Total RNA was extracted from cells using
TRIzol® reagent (Thermo Fisher Scientific, Inc.), and
NanoDrop 1000 Spectrophotometer (Thermo Fisher Scientific, Inc.)
was used to measure the concentration and purity. Total RNA (1 µg)
was reverse transcribed into cDNA using TaqMan™ Reverse
Transcription Reagents (Applied Biosystems; Thermo Fisher
Scientific, Inc.), according to the manufacturer's instructions, in
a 20 µl volume of reaction mixture. qPCR was subsequently performed
using the SYBR® Premix Ex Taq™ kit (cat. no. RR820A;
Takara Bio, Inc.), according to the manufacturer's instructions.
The following primer sequences were used for qPCR: MET forward,
5′-CCTGCGAAGTGAAGGGTCTCC-3′ and reverse,
5′-CTGGCAGCTTTGCACCTGTTT-3′; and GAPDH forward,
5′-CTTCAAGATCATCAGCAATGC-3′ and reverse,
5′-GTTGAAGTCAGAGGAGACCACC-3′. The following thermocycling
conditions were used for qPCR: Initial denaturation at 95°C for 5
min, followed by 39 cycles of denaturation at 95°C for 30 sec,
annealing at 60°C for 30 sec and extension at 72°C for 60 sec, and
a final extension step at 72°C for 5 min. Relative expression
levels were calculated using the 2−ΔΔCq method (20) and normalized to the internal
reference gene GAPDH. All experiments were performed in
triplicate.
Cell Counting Kit-8 (CCK-8) assay
PANC-1 cells were seeded into 96-well plates at a
density of 5,000 cells/well, with each sample containing three
replicates. CCK-8 reagent (10%, Dojindo Molecular Technologies,
Inc.) was added to each well following culturing for 0, 6, 12, 18,
24, 30, 36, 42 and 48 h. Following incubation with CCK-8 for 1 h,
the results were analyzed at a wavelength of 450 nm, using a
microplate reader (Bio-Rad Laboratories, Inc.). All experiments
were performed in triplicate.
Wound healing assay
PANC-1 cells were seeded into 6-well plates
(104/well) and cultured until they reached 90%
confluence. Subsequently, the cells were serum-starved for 12 h,
the cell monolayers were scratched using 200-µl sterile pipette
tips, and the cells were continued to be cultured with serum-free
DMEM. Wound healing was observed at 0, 24 and 48 h under an
inverted light microscope (magnification, ×40; Olympus
Corporation).
Transwell assay
Cell invasion was evaluated using Transwell chambers
coated with Matrigel at 37°C for 48 h (BD Biosciences). A total of
2×105 PANC-1 cells were plated in the upper chambers of
24-well Transwell plates in serum-free medium and 200 µl cell
suspension was subsequently added. DMEM (700 µl) supplemented with
20% FBS was plated in the lower chambers. Following incubation for
48 h at 37°C, cells were fixed with 4% paraformaldehyde for 15 min
at room temperature and subsequently stained with 0.1% crystal
violet for 20 min at room temperature. The non-invading cells were
removed using a wet cotton swab, whilst the stained cells were
counted under an inverted light microscope (magnification, ×100;
Olympus Corporation).
Western blotting
PANC-1 cells were lysed using RIPA lysis buffer
(Beyotime Institute of Biotechnology) supplemented with protease
and phosphatase inhibitors (Beyotime Institute of Biotechnology).
Protein concentration was determined using the BCA protein assay
kit (cat. no. P0012S; Beyotime Institute of Biotechnology) and 40
µg protein/lane was separated by 10% SDS-PAGE. The separated
proteins were subsequently transferred onto PVDF membranes and
blocked with 5% non-fat milk for 1 h at room temperature. The
membranes were incubated with primary antibodies against rabbit
anti-GAPDH (1:1,000; cat. no. ab9485; Abcam), PI3K (1:1,000; cat.
no. 4249; Cell Signaling Technology, Inc.), phosphorylated (p)-PI3K
(1:1,000; cat. no. 17366; Cell Signaling Technology, Inc.), Akt
(1:1,000; cat. no. 4691; Cell Signaling Technology, Inc.) and p-Akt
(1:1,000; cat. no. 4060; Cell Signaling Technology, Inc.) overnight
at 4°C. Following the primary incubation, membranes were incubated
with HRP-conjugated goat anti-rabbit (1:2,000; cat. no. ab150079;
Abcam) secondary antibody at room temperature for 1 h. Protein
bands were visualized using ECL reagents (Tanon, Science and
Technology Co., Ltd.) and the intensity of each band was quantified
using Image Lab software (version 4.1; Bio-Rad Laboratories, Inc.).
All experiments were performed in triplicate.
miRNA-MET prediction
The DIANA (http://diana.imis.athena-innovation.gr), TargetScan
(http://www.targetscan.org), miRDB
(http://www.mirdb.org) and miRTarBase (http://mirtarbase.mbc.nctu.edu.tw) databases were
used to predict MET-targeted miRNAs. The results from the four
databases were intersected to obtain six common miRNAs, and
functional and signaling pathway enrichment analyses of the miRNAs
were subsequently performed. The TargetScanHuman database
(http://www.targetscan.org) was used to
analyze and predict the MET-targeted miRNAs, as well as to obtain
the sequences of fragments that contain binding sites.
Statistical analysis
Statistical analysis was performed using SPSS 20.0
software (IBM Corp.). All experiments were performed in triplicate
and data are presented as the mean ± standard deviation. One-way
ANOVA followed by Dunnett's multiple comparison test were used to
compare differences among multiple groups, while differences
between two groups were compared by Wilcoxon rank-sum test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Screening DEGs in the microarray
datasets
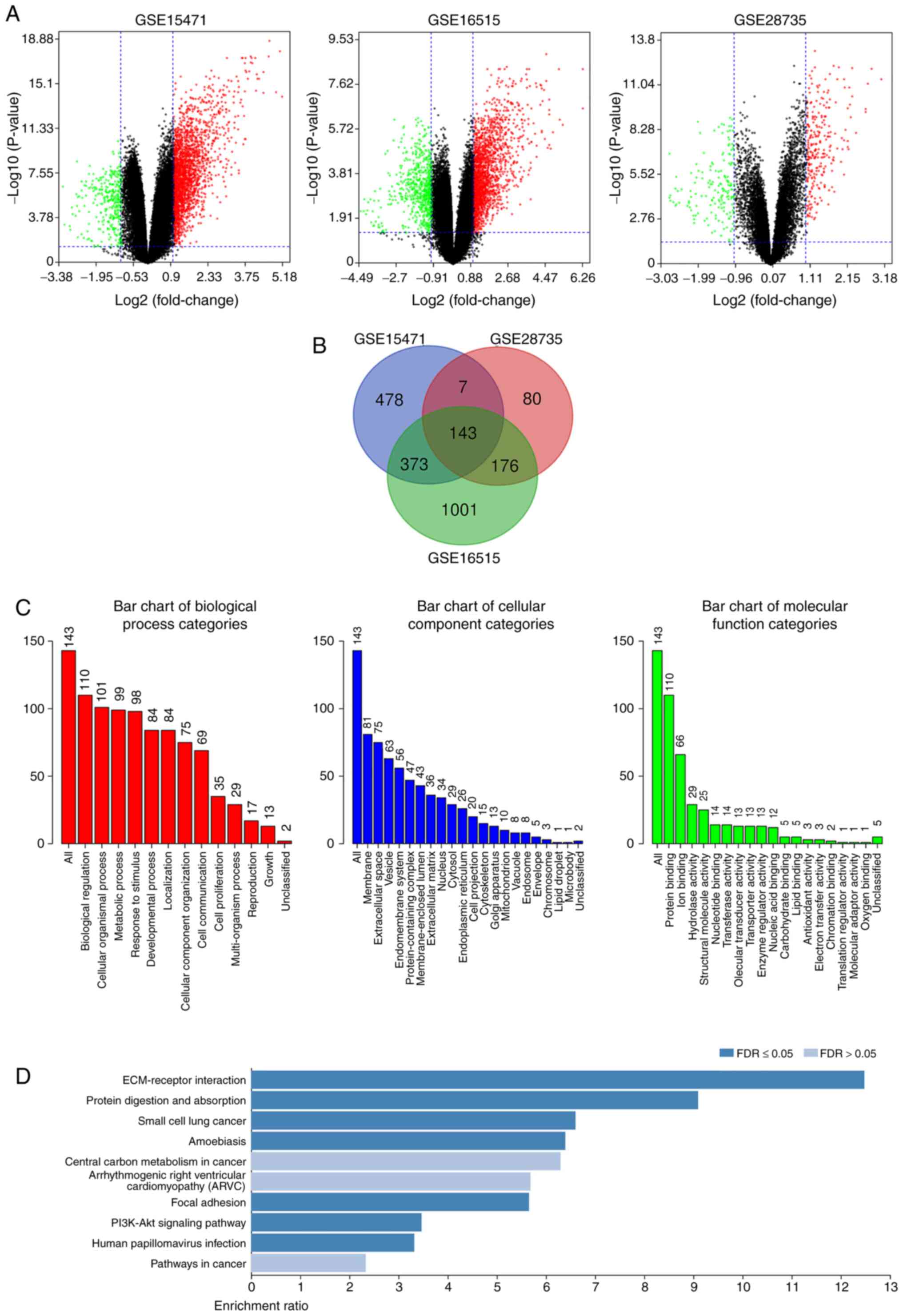
The present study analyzed the DEGs in PDAC tissues
and normal pancreatic tissues. The DEGs were obtained following
standardization of the microarray results, which demonstrated that
the GSE15471 dataset contained 1,330 upregulated genes and 66
downregulated genes, the GSE16515 dataset contained 1,801
upregulated genes and 561 downregulated genes, and the GSE28735
dataset contained 267 upregulated genes and 156 downregulated genes
(Fig. 1A). As presented in the Venn
diagram, the intersection of the three microarray datasets
contained 143 genes, composed of 132 upregulated genes and 11
downregulated genes (Fig. 1B).
Functional and signaling pathway
enrichment analyses of the DEGs
The DAVID database was used to perform GO and KEGG
functional and signaling pathway enrichment analyses of the DEGs.
GO analysis demonstrated that the DEG BP are mainly focused on cell
metabolism, intercellular communication and cell proliferation
regulation. CC are mainly concentrated in the structural
composition of extracellular matrix, cell membrane system, and the
binding with collagen and integrin. MF is mainly focused on cell
molecular transport, protein binding and enzyme activity regulation
(Fig. 1C). KEGG analysis
demonstrated that DEG signaling pathways mainly enriched
extracellular matrix receptors, protein digestion and absorption
and the PI3K-Akt signaling pathway (Fig.
1D).
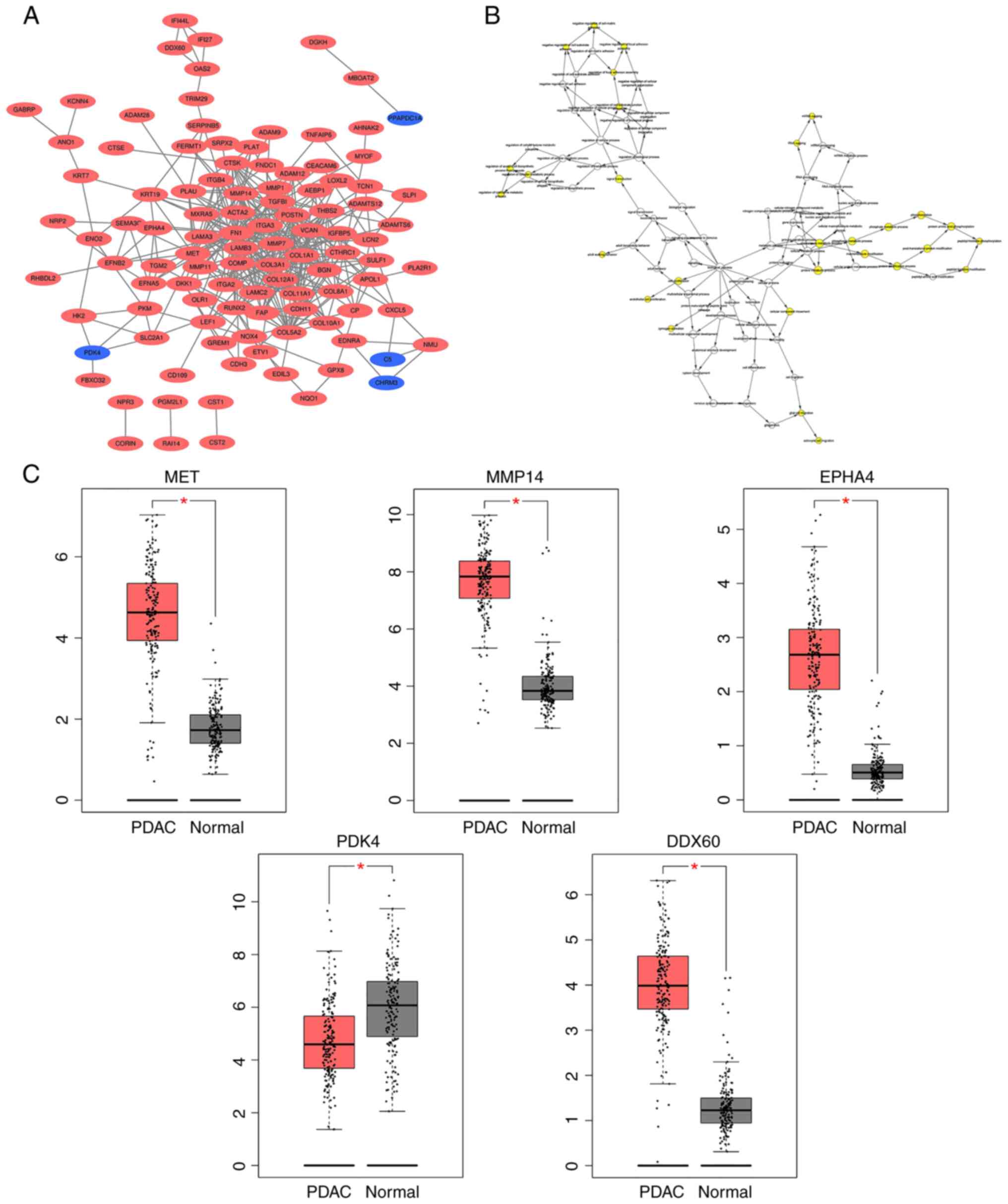
PPI network construction, and
selection and analysis of hub genes
A DEG-based PPI network was constructed using the
STRING database and visualized using Cytoscape software (Fig. 2A). MCODE was subsequently used to
screen out five hub genes [MET, matrix metalloproteinase (MMP)14,
EPHA4, pyruvate dehydrogenase kinase (PDK)4 and DDX60] from the
DEGs. PDK4 expression was downregulated, while the expression
levels of the other four hub genes were upregulated. BiNGO was
subsequently used to assess the biological processes of the hub
genes, revealing that their functions were mostly focused on cell
proliferation and migration, phosphorylation metabolism process and
matrix metalloproteinase activation (Fig. 2B). Analysis using the GEPIA database
demonstrated that the differences in the expression levels of these
five hub genes between PDAC tissues and normal pancreatic tissues
were statistically significant (all P<0.05; Fig. 2C).
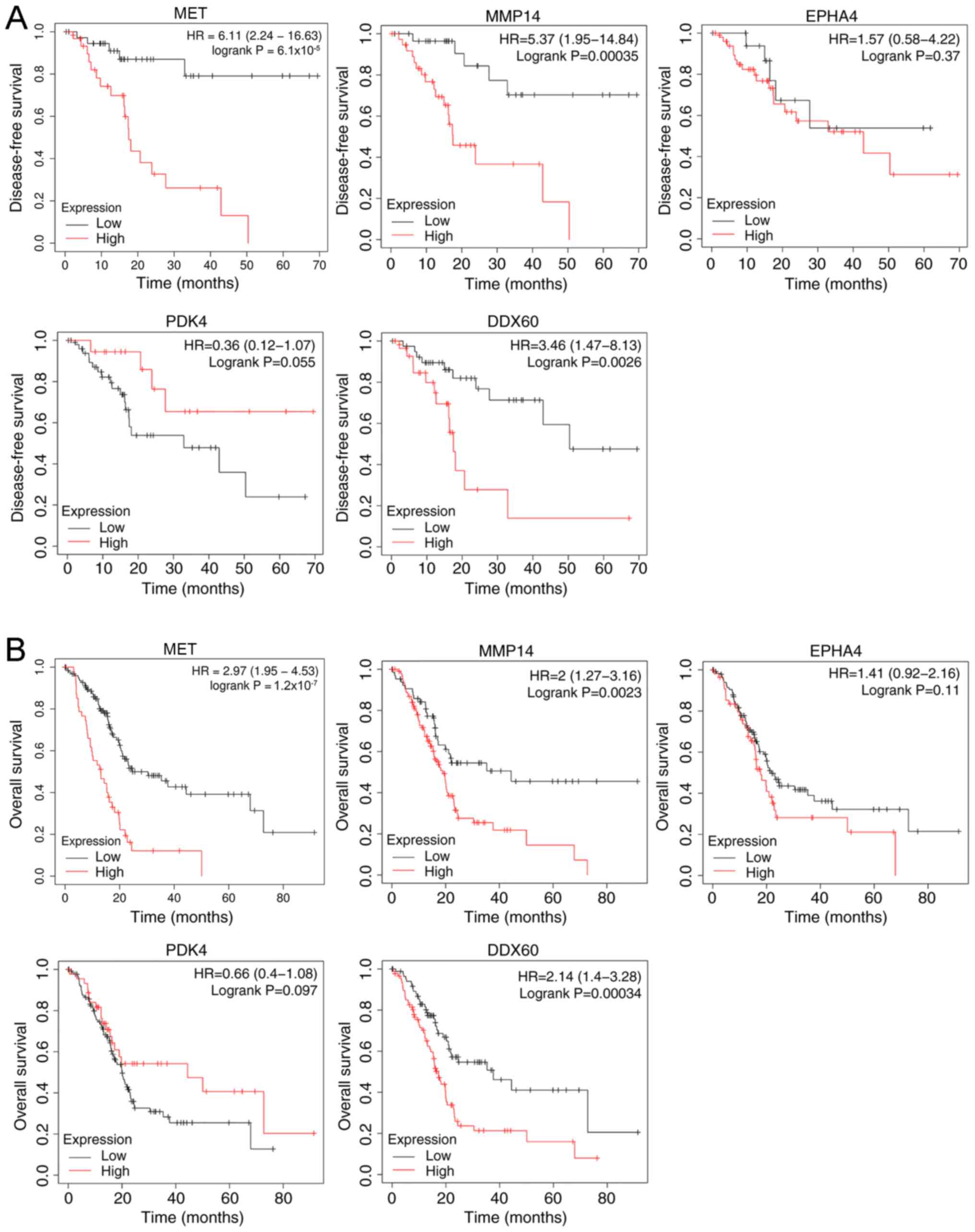
Kaplan-Meier survival analysis of the hub genes
demonstrated that both high expression levels of MET, MMP14, EPHA4
and DDX60, and low PDK4 expression resulted in a shorter
disease-free survival time (Fig. 3A)
and overall survival time (Fig. 3B)
of patients with PDAC.
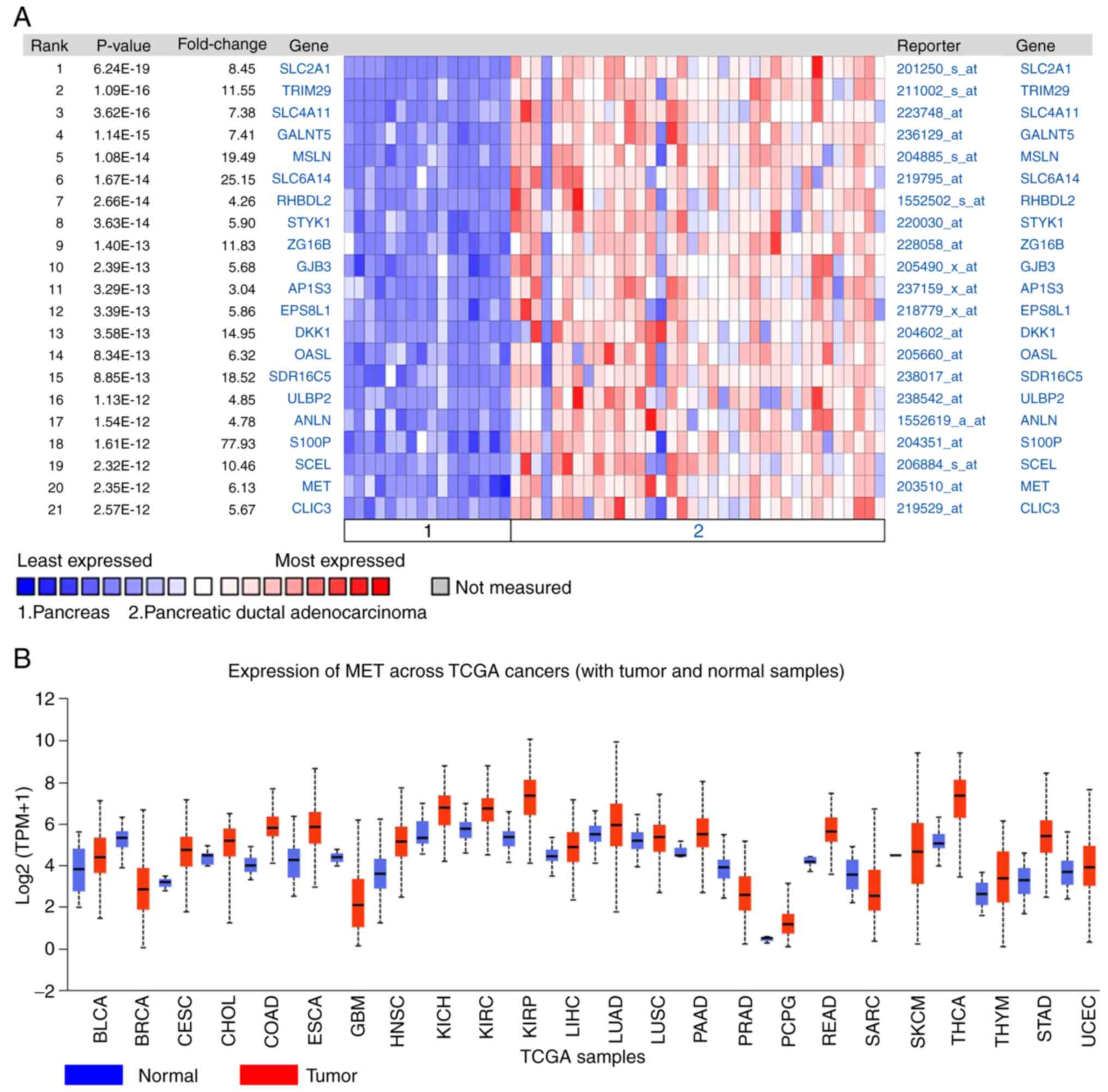
The DEGs between PDAC tissues and normal pancreatic
tissues were analyzed using the Oncomine database. The results
demonstrated that MET expression was 6.13 times higher in PDAC
tissues compared with normal tissues (Fig. 4A). The expression of MET in different
tumors was subsequently analyzed, and the results demonstrated that
MET expression was upregulated in different types of cancer,
including lung, cervical, gastric and colorectal cancer (Fig. 4B).
Silencing MET inhibits the
proliferation, migration and invasion of PDAC cells
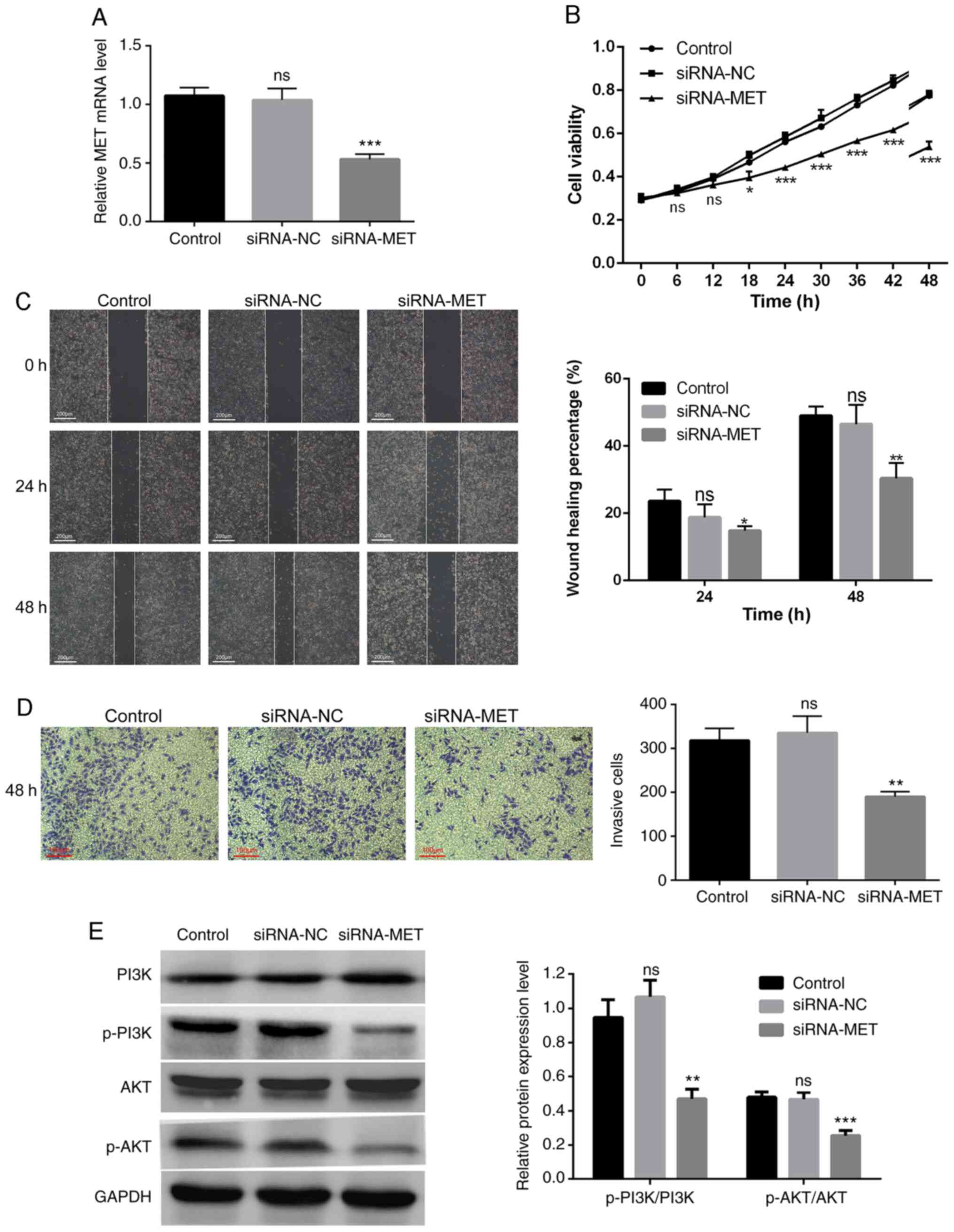
MET expression was knocked down in PANC-1 cells via
siRNA transfection (Fig. 5A). The
results of the CCK-8 assay confirmed that MET knockdown
significantly inhibited the proliferation of PANC-1 cells (Fig. 5B). The results of the wound healing
and Transwell assays demonstrated that high MET expression promoted
the migration and invasion of PDAC cells. In addition, the wound
healing rate of the siRNA-MET group significantly decreased
compared with the control group (P<0.05), whereas the
differences between the siRNA-NC group and the control group were
not statistically significant (P>0.05; Fig. 5C). The number of invasive cells in
the siRNA-MET group was significantly lower compared with the
control group (P<0.01), and the differences between the siRNA-NC
group and the control group were not statistically significant
(P>0.05; Fig. 5D).
MET promotes the progression of PDAC
by activating the PI3K/AKT signaling pathway
To further study the molecular mechanism by which
MET regulates the proliferation, migration and invasion of PDAC
cells, the protein levels of the PI3K/AKT pathway-related molecules
were assessed in PANC-1 cells transfected with siRNA-MET. The
results demonstrated that MET knockdown significantly decreased the
levels of phosphorylated PI3K and AKT, but not their total protein
levels (Fig. 5E). Taken together,
these results suggest that MET is an upstream signal factor that
modulates the PI3K/AKT pathway in PDAC, and that MET promotes the
proliferation, migration and invasion of PDAC cells through
activation of the PI3K/AKT pathway.
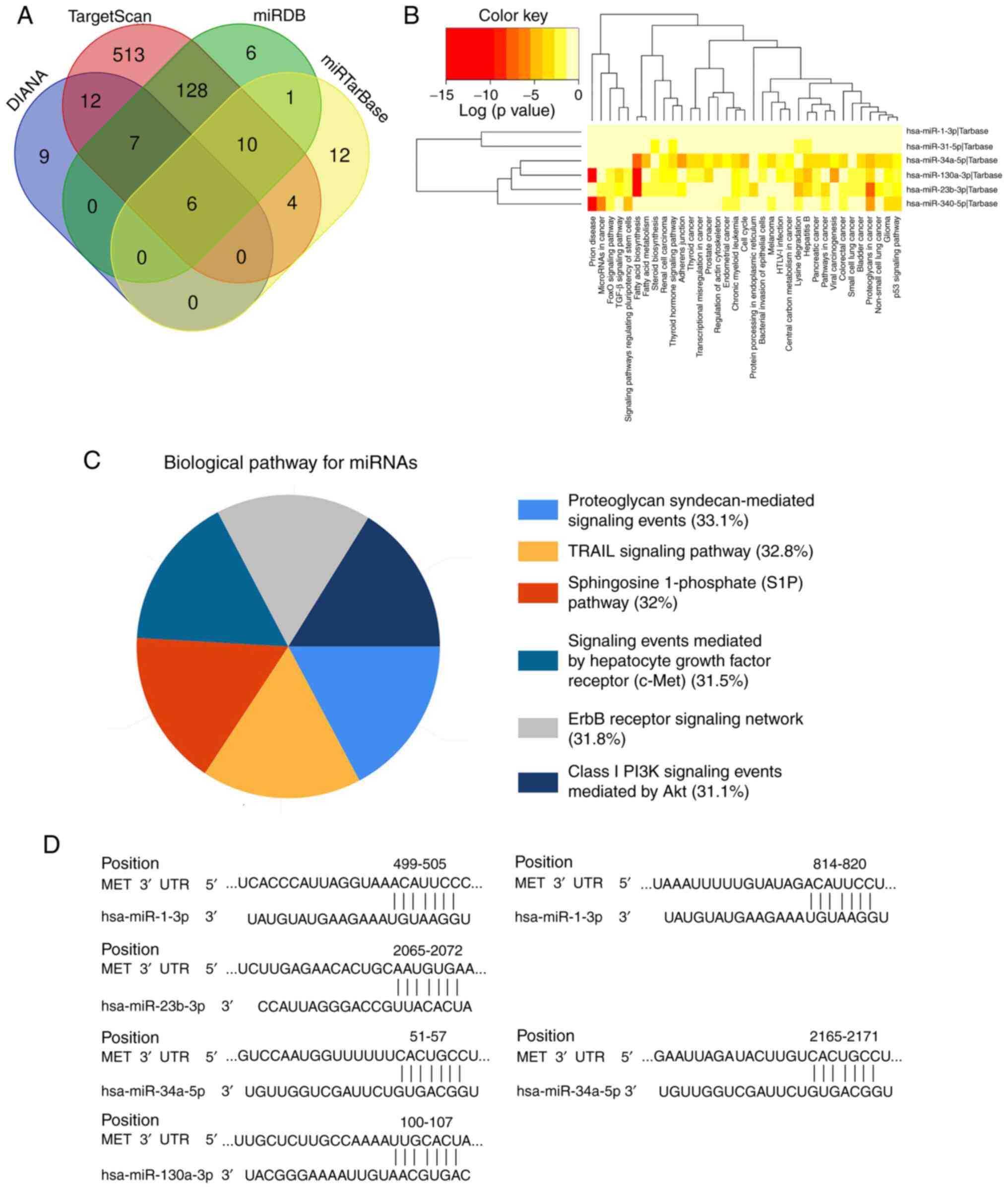
Prediction of the miRNA-MET signaling
pathway
A total of six MET-targeted miRNAs (miR-1-3p,
miR-23b-3p, miR-31-5p, miR-34a-5p, miR-340-5p and miR-130a-3p) were
screened using the DIANA, TargetScan, miRDB, and miRTarBase
databases (Fig. 6A). Functional and
signaling pathway analyses of these six miRNAs demonstrated that
their functions mainly focused on prion diseases, fatty acid
synthesis and metabolism, and they are involved in the progression
of thyroid, prostate, colorectal and pancreatic cancers. The
signaling pathways mainly include the TRAIL-mediated external
apoptosis signaling pathway, sphingosine-1-phosphate signaling
pathway, c-MET/hepatocyte growth factor (HGF) signaling pathway,
PI3K/Akt signaling pathway and ErbB receptor signaling pathway
(Fig. 6B and C). A total of four
miRNAs (miR-1-3p, miR-23b-3p, miR-34a-5p and miR-130a-3p) were
demonstrated to have conserved binding sites with MET, using the
TargetScanHuman database (Fig. 6D).
Thus, it is suggested that these four miRNAs can act as upstream
regulators of the MET/PI3K/AKT pathway, and serve as potential
biomarkers and therapeutic targets for patients with PDAC.
Discussion
Currently, there are still significant challenges
facing the early diagnosis of PDAC. A lack of understanding of the
molecular mechanism in PDAC progression and effective treatments
for PDAC facilitates a poor prognosis in patients. Thus, the
identification of signal factors that play a key regulatory role in
the occurrence and development of PDAC is of great importance, and
they may become pivotal early diagnostic biomarkers and targets in
the treatment of PDAC.
Microarray technology can be used for studying gene
transcription and epigenetic changes, and it is an incredibly
effective method for identifying disease biomarkers (21). In the present study, microarray
technology and bioinformatics methods were used to analyze three
PDAC-related microarray datasets, which were downloaded from the
GEO database, and 143 DEGs between PDAC tissues and normal tissue
were screened out, including 132 upregulated genes and 11
downregulated genes. GO and KEGG functional and signaling pathway
enrichment analyses were performed, and it was discovered that
DEG's functions are mainly focused on the extracellular matrix
(ECM), protein digestion and absorption, and the regulation of
cellular metabolic processes. ECM is a network structure that is
composed of collagen, glycoprotein and proteoglycan, and provides
biomechanics for the regulation of cell behavior, while maintaining
organs' morphology and integrity (22). ECM is in dynamic equilibrium under
the influence of extracellular proteases and their inhibitors;
however, when a tumor grows, the dynamic balance is broken, which
triggers pathological ECM remodeling, and results in reduced
adhesion of cells to the ECM (23).
This is conducive to cancer cells invading nearby organs and blood
vessels, thereby promoting cancer invasion and metastasis (24). This also explains the reason for
PDAC's high invasiveness (23).
A total of five hub genes (MET, MMP14, EPHA4, PDK4
and DDX60) were screened out from the DEGs, PDK4 was downregulated
and the other four genes were upregulated. MET is a potential
therapeutic target for different types of cancer, and it can
promote cell proliferation, migration and epithelial-to-mesenchymal
transition (25). The binding of MET
with its ligand, HGF, is a key driver in the development of cancer
(26). MMP14 is a type of MMP, which
is upregulated in different types of cancer, including colorectal,
lung and nasopharyngeal cancer (27–29).
MMP14 promotes angiogenesis, inflammation, invasion and metastasis
of cancer cells, and severely affects the prognosis of patients
(30). EPHA4 plays a significant
role in neurodegenerative diseases, including Alzheimer's disease
and amyotrophic lateral sclerosis (31). Previous studies have demonstrated
that EPHA4 is upregulated in PDAC and promotes tumor growth
(32–34). Its high expression is also associated
with poor prognosis of patients with gastric cancer and breast
cancer (35). PDK plays a vital role
in the regulation of cell metabolism and mitochondrial function
(36). PDK includes four isoenzymes
(PDK1, 2, 3, and 4); PDK4 provides a tri-carbon substrate
(pyruvate, lactic acid and alanine) for gluconeogenesis during
fasting to maintain blood glucose levels (37). RIG-I-mediated type I interferon (IFN)
and nuclease-mediated viral RNA degradation are of great importance
for the innate antiviral immune response in vivo (38). DDX60 is an IFN-induced cytoplasmic
helicase that participates in viral RNA degradation, performs a
vital role in the innate immune response and is dependent on the
RIG-I antiviral pathway (39). A
previous study has demonstrated that DDX60 is highly expressed in
oral cancer and is closely associated with tumor progression and
poor prognosis (40). The results of
the Oncomine and GEPIA analyses in the present study demonstrated
that MET, MMP14, EPHA4 and DDX60 were highly expressed in PDAC,
while PDK4 was expressed at low levels. The impact of hub genes on
the survival time of patients with PDAC was subsequently analyzed
and the results revealed that abnormal expression of these five hub
genes was associated with poor disease-free survival time and
overall survival time. Taken together, these results suggest that
the five hub genes play a significant role in the progression of
PDAC. Furthermore, through a series of biological experiments, it
was confirmed that the hub gene, MET, can promote the
proliferation, migration and invasion of PDAC cells via the
PI3K/AKT signaling pathway.
miRNAs, as major regulators of several biological
and pathological processes, including growth, development,
metabolism, infection, immunity, cell death, messenger signaling,
DNA repair and self-renewal, have exhibited carcinogenic or
anticancer activity in different types of human tumors (41,42).
Increasing evidence suggests that several miRNAs are crucial for
cancer development as they regulate various biological processes in
cancer cells, including proliferation, differentiation, apoptosis,
cell cycle, migration and invasion, which has led to miRNAs
becoming potential diagnostic biomarkers and therapeutic targets
for different types of cancer (43).
In the present study, six MET-targeted miRNAs were screened out
using several online databases, and four (miR-1-3p, miR-23b-3p,
miR-34a-5p and miR-130a-3p) of them were demonstrated to have
conserved binding sites with MET. Previous studies have
demonstrated that miR-1-3p expression is downregulated in different
types of cancer, including colon cancer, esophageal squamous cell
carcinoma and bladder cancer, and that the overexpression of
miR-1-3p can inhibit the growth of these tumors (44–46). Li
et al (47) demonstrated that
miR-1-3p can inhibit prostate cancer cell proliferation by
regulating the cell cycle-related genes, E2F5 and PFTK1. miR-23b-3p
is considered a tumor suppressor that affects the carcinogenesis
and aggressiveness of liver cancer, and it can be used as a
predictive biomarker and therapeutic target for liver cancer
(48). Ge and Li (49) have demonstrated that long non-coding
RNA SNHG17 can promote glioma cell proliferation, migration and
invasion by regulating the miR-23b-3p/ZHX1 axis. Studies have
reported that miR-34a-5p is involved in the occurrence and
development of cancer through regulation of the Notch, PI3K/Akt,
SIRT1/HIF-1α and Wnt/β-Catenin signaling pathways (50–52). As
a tumor suppressor, miR-34a-5p is able to inhibit the growth of
glioma and esophageal squamous cell carcinoma (53,54).
miR-130a-3p is a key regulator of human cancer, and it can inhibit
the proliferation, migration and invasion of various cancer cells,
such as liver cancer, breast cancer and nasopharyngeal carcinoma
(55–57). Notably, Dai et al (58) have also demonstrated that miR-130a-3p
promotes the growth of gastric cancer. Several studies have
reported that abnormal miRNA expression is closely associated with
the occurrence and development of PDAC, and that miRNAs can be used
as early detection biomarkers for potential treatment and
prognostic monitoring targets (59,60).
However, the miRNAs that play a regulatory role in the development
of PDAC and their molecular mechanisms are rarely reported. The
present study assessed four miRNAs (miR-1-3p, miR-23b-3p,
miR-34a-5p and miR-130a-3p) that can potentially be involved in
PDAC development; however, further studies are required to
determine their molecular mechanisms.
MET is a receptor for HGF, which is encoded by the
proto-oncogene, c-Met. MET is a tyrosine kinase receptor that is
mainly distributed in the membranes of epithelial-derived or
endothelium-derived cells (61). MET
is upregulated in different types of cancer, including breast
cancer, colorectal cancer, glioma and non-small cell lung cancer
(62–64). MET has the ability to help cells
adapt to adverse environment, promote tumor angiogenesis, enhance
tumor cell survival, and their metastatic and invasive abilities
(65). This is consistent with the
experimental results of the present study. When HGF ligand binds to
a MET receptor, MET is activated and induces intracellular tyrosine
residue phosphorylation, activating the downstream RAS/ERK/MAPK,
PI3K/Akt, Wnt/β-catenin and STAT signaling pathways (64). These pathways can drive
proliferation, migration, invasion, angiogenesis and
epithelial-to-mesenchymal transition in a variety of tumors
(66–68). According to the functional and
signaling pathway enrichment analysis of DEGs and miRNAs in the
present study, they were all involved in the PI3K/Akt signaling
pathway. The PI3K/Akt signaling pathway plays a crucial regulatory
role in different types of cancer, including PDAC (69). In the present study, western blot
analysis demonstrated that MET activated the PI3K/AKT pathway in
PDAC cells. Based on this study's experimental results and
bioinformatics analysis, it can be speculated that under the
modulation of miRNA, MET promotes PDAC progression through the
PI3K/AKT signaling pathway. In addition, four alternative
MET-targeted miRNAs, miR-1-3p, miR-23b-3p, miR-34a-5p and
miR-130a-3p are provided in the present study.
Future studies will perform dual-luciferase reporter
assays to verify the targeting association between these four
miRNAs and MET. In addition, the expression levels of the miRNAs
will be restrained in PANC-1 cells via transfection, and cell
proliferation, migration and invasion will be assessed.
Furthermore, prospective studies will perform western blot analysis
to detect the expression levels of PI3K/AKT pathway-related
proteins. Collectively, these experiments will help identify novel
diagnostic biomarkers and therapeutic targets for patients with
PDAC.
In conclusion, the present study aimed to identify
DEGs and molecular mechanisms that affect the progression of PDAC
to discover potential diagnostic biomarkers and therapeutic targets
for patients with PDAC. A total of 143 DEGs were screened out, five
hub genes and four miRNAs were further identified, and the
cancer-promoting effect of MET in PDAC was proven. According to the
functional and signaling pathway enrichment analysis of DEGs and
miRNAs, it can be speculated that these four miRNAs affect PDAC
progression by targeting MET via the PI3K/AKT signaling pathway.
However, further biological experiments must be performed to
confirm this speculation. Notably, the present study identified a
novel mechanism that affects PDAC progression, and provided four
potential miRNAs that can serve as promising biomarkers for the
early diagnosis and treatment of patients with PDAC.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Zhejiang
Provincial Natural Science Foundation of China (grant no.
LGF19H290004).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author upon reasonable
request.
Authors' contributions
LCY, XHJ, SSY, WW and ZGT contributed to
experimental design, data acquisition, and drafting and editing the
initial manuscript. LW, LLZ, FX, TJ and LY analyzed the data and
assisted in the experiments. All authors have read and approved the
final version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ilic M and Ilic I: Epidemiology of
pancreatic cancer. World J Gastroenterol. 22:9694–9705. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lin QJ, Yang F, Jin C and Fu DL: Current
status and progress of pancreatic cancer in China. World J
Gastroenterol. 21:7988–8003. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhou B, Xu JW, Cheng YG, Gao JY, Hu SY,
Wang L and Zhan HX: Early detection of pancreatic cancer: Where are
we now and where are we going? Int J Cancer. 141:231–241. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Garrido-Laguna I and Hidalgo M: Pancreatic
cancer: From state-of-the-art treatments to promising novel
therapies. Nat Rev Clin Oncol. 12:319–334. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Waddell N, Pajic M, Patch AM, Chang DK,
Kassahn KS, Bailey P, Johns AL, Miller D, Nones K, Quek K, et al:
Whole genomes redefine the mutational landscape of pancreatic
cancer. Nature. 518:495–501. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dreyer SB, Chang DK, Bailey P and Biankin
AV: Pancreatic cancer genomes: Implications for clinical management
and therapeutic development. Clin Cancer Res. 23:1638–1646. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bailey P, Chang DK, Nones K, Johns AL,
Patch AM, Gingras MC, Miller DK, Christ AN, Bruxner TJ, Quinn MC,
et al: Genomic analyses identify molecular subtypes of pancreatic
cancer. Nature. 531:47–52. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li L, Lei Q, Zhang S, Kong L and Qin B:
Screening and identification of key biomarkers in hepatocellular
carcinoma: Evidence from bioinformatic analysis. Oncol Rep.
38:2607–2618. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Badea L, Herlea V, Dima SO, Dumitrascu T
and Popescu I: Combined gene expression analysis of whole-tissue
and microdissected pancreatic ductal adenocarcinoma identifies
genes specifically overexpressed in tumor epithelia.
Hepatogastroenterology. 55:2016–2027. 2008.PubMed/NCBI
|
|
10
|
Pei H, Li L, Fridley BL, Jenkins GD,
Kalari KR, Lingle W, Petersen G, Lou Z and Wang L: FKBP51 affects
cancer cell response to chemotherapy by negatively regulating Akt.
Cancer Cell. 16:259–266. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang G, Schetter A, He P, Funamizu N,
Gaedcke J, Ghadimi BM, Ried T, Hassan R, Yfantis HG, Lee DH, et al:
DPEP1 inhibits tumor cell invasiveness, enhances chemosensitivity
and predicts clinical outcome in pancreatic ductal adenocarcinoma.
PLoS One. 7:e315072012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tian T, Wang J and Zhou X: A review:
microRNA detection methods. Org Biomol Chem. 13:2226–2238. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hayes J, Peruzzi PP and Lawler S:
MicroRNAs in cancer: Biomarkers, functions and therapy. Trends Mol
Med. 20:460–469. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mishra S, Yadav T and Rani V: Exploring
miRNA based approaches in cancer diagnostics and therapeutics. Crit
Rev Oncol Hematol. 98:12–23. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Qu K, Zhang X, Lin T, Liu T, Wang Z, Liu
S, Zhou L, Wei J, Chang H, Li K, et al: Circulating miRNA-21-5p as
a diagnostic biomarker for pancreatic cancer: Evidence from
comprehensive miRNA expression profiling analysis and clinical
validation. Sci Rep. 7:16922017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ashburner M, Ball CA, Blake JA, Botstein
D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT,
et al: Gene ontology: Tool for the unification of biology. The gene
ontology consortium. Nat Genet. 25:25–29. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kanehisa M, Furumichi M, Tanabe M, Sato Y
and Morishima K: KEGG: New perspectives on genomes, pathways,
diseases and drugs. Nucleic Acids Res. 45(D1): D353–D361. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rabbani G, Baig MH, Ahmad K and Choi I:
Protein-protein Interactions and their role in various diseases and
their prediction techniques. Curr Protein Pept Sci. 19:948–957.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rhodes DR, Yu J, Shanker K, Deshpande N,
Varambally R, Ghosh D, Barrette T, Pandey A and Chinnaiyan AM:
ONCOMINE: A cancer microarray database and integrated data-mining
platform. Neoplasia. 6:1–6. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kulasingam V and Diamandis EP: Strategies
for discovering novel cancer biomarkers through utilization of
emerging technologies. Nat Clin Pract Oncol. 5:588–599. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Vogel V: Unraveling the mechanobiology of
extracellular matrix. Annu Rev Physiol. 80:353–387. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tian C, Clauser KR, Öhlund D, Rickelt S,
Huang Y, Gupta M, Mani DR, Carr SA, Tuveson DA and Hynes RO:
Proteomic analyses of ECM during pancreatic ductal adenocarcinoma
progression reveal different contributions by tumor and stromal
cells. Proc Natl Acad Sci USA. 116:19609–19618. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yuzhalin AE, Lim SY, Kutikhin AG and
Gordon-Weeks AN: Dynamic matrisome: ECM remodeling factors
licensing cancer progression and metastasis. Biochim Biophys Acta
Rev Cancer. 1870:207–228. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Moosavi F, Giovannetti E, Saso L and
Firuzi O: HGF/MET pathway aberrations as diagnostic, prognostic,
and predictive biomarkers in human cancers. Crit Rev Clin Lab Sci.
56:533–566. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Matsumoto K, Umitsu M, De Silva DM, Roy A
and Bottaro DP: Hepatocyte growth factor/MET in cancer progression
and biomarker discovery. Cancer Sci. 108:296–307. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cui G, Cai F, Ding Z and Gao L: MMP14
predicts a poor prognosis in patients with colorectal cancer. Hum
Pathol. 83:36–42. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Stawowczyk M, Wellenstein MD, Lee SB,
Yomtoubian S, Durrans A, Choi H, Narula N, Altorki NK, Gao D and
Mittal V: Matrix metalloproteinase 14 promotes lung cancer by
cleavage of heparin-binding EGF-like growth factor. Neoplasia.
19:55–64. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yan T, Lin Z, Jiang J, Lu S, Chen M, Que
H, He X, Que G, Mao J, Xiao J and Zheng Q: MMP14 regulates cell
migration and invasion through epithelial-mesenchymal transition in
nasopharyngeal carcinoma. Am J Transl Res. 7:950–958.
2015.PubMed/NCBI
|
|
30
|
Gonzalez-Molina J, Gramolelli S, Liao Z,
Carlson JW, Ojala PM and Lehti K: MMP14 in sarcoma: A regulator of
tumor microenvironment communication in connective tissues. Cells.
8:9912019. View Article : Google Scholar
|
|
31
|
Vargas LM, Cerpa W, Muñoz FJ, Zanlungo S
and Alvarez AR: Amyloid-β oligomers synaptotoxicity: The emerging
role of EphA4/c-Abl signaling in Alzheimer's disease. Biochim
Biophys Acta Mol Basis Dis. 1864:1148–1159. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu C, Huang H, Wang C, Kong Y and Zhang
H: Involvement of ephrin receptor A4 in pancreatic cancer cell
motility and invasion. Oncol Lett. 7:2165–2169. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Iiizumi M, Hosokawa M, Takehara A, Chung
S, Nakamura T, Katagiri T, Eguchi H, Ohigashi H, Ishikawa O,
Nakamura Y and Nakagawa H: EphA4 receptor, overexpressed in
pancreatic ductal adenocarcinoma, promotes cancer cell growth.
Cancer Sci. 97:1211–1216. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Giaginis C, Tsourouflis G,
Zizi-Serbetzoglou A, Kouraklis G, Chatzopoulou E, Dimakopoulou K
and Theocharis SE: Clinical significance of ephrin (eph)-A1, -A2,
-a4, -a5 and -a7 receptors in pancreatic ductal adenocarcinoma.
Pathol Oncol Res. 16:267–276. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Takano H, Nakamura T, Tsuchikawa T,
Kushibiki T, Hontani K, Inoko K, Takahashi M, Sato S, Abe H,
Takeuchi S, et al: Inhibition of Eph receptor A4 by
2,5-dimethylpyrrolyl benzoic acid suppresses human pancreatic
cancer growing orthotopically in nude mice. Oncotarget.
6:41063–41076. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Michelakis ED, Gurtu V, Webster L, Barnes
G, Watson G, Howard L, Cupitt J, Paterson I, Thompson RB, Chow K,
et al: Inhibition of pyruvate dehydrogenase kinase improves
pulmonary arterial hypertension in genetically susceptible
patients. Sci Transl Med. 9:eaao45832017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Leem J and Lee IK: Mechanisms of vascular
calcification: The pivotal role of pyruvate dehydrogenase kinase 4.
Endocrinol Metab (Seoul). 31:52–61. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chan YK and Gack MU: RIG-I-like receptor
regulation in virus infection and immunity. Curr Opin Virol.
12:7–14. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Oshiumi H, Miyashita M, Okamoto M, Morioka
Y, Okabe M, Matsumoto M and Seya T: DDX60 is involved in
RIG-I-dependent and independent antiviral responses, and its
function is attenuated by virus-induced EGFR Activation. Cell Rep.
11:1193–1207. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Fu TY, Wu CN, Sie HC, Cheng JT, Lin YS,
Liou HH, Tseng YK, Shu CW, Tsai KW, Yen LM, et al: Subsite-specific
association of DEAD box RNA helicase DDX60 with the development and
prognosis of oral squamous cell carcinoma. Oncotarget.
7:85097–85108. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kabekkodu SP, Shukla V, Varghese VK,
D'Souza J, Chakrabarty S and Satyamoorthy K: Clustered miRNAs and
their role in biological functions and diseases. Biol Rev Camb
Philos Soc. 93:1955–1986. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Romano G and Kwong LN: Diagnostic and
therapeutic applications of miRNA-based strategies to cancer
immunotherapy. Cancer Metastasis Rev. 37:45–53. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Liang B, Li Y and Wang T: A three miRNAs
signature predicts survival in cervical cancer using bioinformatics
analysis. Sci Rep. 7:56242017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wang JY, Huang JC, Chen G and Wei DM:
Expression level and potential target pathways of miR-1-3p in
colorectal carcinoma based on 645 cases from 9 microarray datasets.
Mol Med Rep. 17:5013–5020. 2018.PubMed/NCBI
|
|
45
|
Sang C, Chao C, Wang M, Zhang Y, Luo G and
Zhang X: Identification and validation of hub microRNAs
dysregulated in esophageal squamous cell carcinoma. Aging (Albany
NY). 12:9807–9824. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wang W, Shen F and Wang C, Lu W, Wei J,
Shang A and Wang C: MiR-1-3p inhibits the proliferation and
invasion of bladder cancer cells by suppressing CCL2 expression.
Tumour Biol. 39:10104283176983832017.PubMed/NCBI
|
|
47
|
Li SM, Wu HL, Yu X, Tang K, Wang SG, Ye ZQ
and Hu J: The putative tumour suppressor miR-1-3p modulates
prostate cancer cell aggressiveness by repressing E2F5 and PFTK1. J
Exp Clin Cancer Res. 37:2192018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
He RQ, Wu PR, Xiang XL, Yang X, Liang HW,
Qiu XH, Yang LH, Peng ZG and Chen G: Downregulated miR-23b-3p
expression acts as a predictor of hepatocellular carcinoma
progression: A study based on public data and RT-qPCR verification.
Int J Mol Med. 41:2813–2831. 2018.PubMed/NCBI
|
|
49
|
Ge BH and Li GC: Long non-coding RNA
SNHG17 promotes proliferation, migration and invasion of glioma
cells by regulating the miR-23b-3p/ZHX1 axis. J Gene Med.
22:e32472020. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Misso G, Di Martino MT, De Rosa G, Farooqi
AA, Lombardi A, Campani V, Zarone MR, Gullà A, Tagliaferri P,
Tassone P and Caraglia M: Mir-34: A new weapon against cancer? Mol
Ther Nucleic Acids. 3:e1942014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Li YY, Tao YW, Gao S, Li P, Zheng JM,
Zhang SE, Liang J and Zhang Y: Cancer-associated fibroblasts
contribute to oral cancer cells proliferation and metastasis via
exosome-mediated paracrine miR-34a-5p. EBioMedicine. 36:209–220.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Xu H, Zhang Y, Qi L, Ding L, Jiang H and
Yu H: NFIX circular RNA promotes glioma progression by regulating
miR-34a-5p via notch signaling pathway. Front Mol Neurosci.
11:2252018. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Ma S, Fu T, Zhao S and Gao M:
MicroRNA-34a-5p suppresses tumorigenesis and progression of glioma
and potentiates Temozolomide-induced cytotoxicity for glioma cells
by targeting HMGA2. Eur J Pharmacol. 852:42–50. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Wang X, Zhao Y, Lu Q, Fei X, Lu C, Li C
and Chen H: MiR-34a-5p inhibits proliferation, migration, invasion
and epithelial-mesenchymal transition in esophageal squamous cell
carcinoma by targeting LEF1 and inactivation of the Hippo-YAP1/TAZ
signaling pathway. J Cancer. 11:3072–3081. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Wang G, Popovic B, Tao J and Jiang A:
Overexpression of COX7RP promotes tumor growth and metastasis by
inducing ROS production in hepatocellular carcinoma cells. Am J
Cancer Res. 10:1366–1383. 2020.PubMed/NCBI
|
|
56
|
Kong X, Zhang J, Li J, Shao J and Fang L:
MiR-130a-3p inhibits migration and invasion by regulating RAB5B in
human breast cancer stem cell-like cells. Biochem Biophys Res
Commun. 501:486–493. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Chen X, Yue B, Zhang C, Qi M, Qiu J, Wang
Y and Chen J: MiR-130a-3p inhibits the viability, proliferation,
invasion, and cell cycle, and promotes apoptosis of nasopharyngeal
carcinoma cells by suppressing BACH2 expression. Biosci Rep.
37:BSR201605762017. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Dai X, Guo X, Liu J, Cheng A, Peng X, Zha
L and Wang Z: Circular RNA circGRAMD1B inhibits gastric cancer
progression by sponging miR-130a-3p and regulating PTEN and p21
expression. Aging (Albany NY). 11:9689–9708. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Madhavan B, Yue S, Galli U, Rana S, Gross
W, Müller M, Giese NA, Kalthoff H, Becker T, Büchler MW and Zöller
M: Combined evaluation of a panel of protein and miRNA
serum-exosome biomarkers for pancreatic cancer diagnosis increases
sensitivity and specificity. Int J Cancer. 136:2616–2627. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Rawat M, Kadian K, Gupta Y, Kumar A, Chain
PSG, Kovbasnjuk O, Kumar S and Parasher G: MicroRNA in pancreatic
cancer: From biology to therapeutic potential. Genes (Basel).
10:7522019. View Article : Google Scholar
|
|
61
|
Bouattour M, Raymond E, Qin S, Cheng AL,
Stammberger U, Locatelli G and Faivre S: Recent developments of
c-Met as a therapeutic target in hepatocellular carcinoma.
Hepatology. 67:1132–1149. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Christensen JG, Burrows J and Salgia R:
c-Met as a target for human cancer and characterization of
inhibitors for therapeutic intervention. Cancer Lett. 225:1–26.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Heist RS, Shim HS, Gingipally S,
Mino-Kenudson M, Le L, Gainor JF, Zheng Z, Aryee M, Xia J, Jia P,
et al: MET exon 14 skipping in non-small cell lung cancer.
Oncologist. 21:481–486. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Raghav K, Bailey AM, Loree JM, Kopetz S,
Holla V, Yap TA, Wang F, Chen K, Salgia R and Hong D: Untying the
gordion knot of targeting MET in cancer. Cancer Treat Rev.
66:95–103. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Comoglio PM, Trusolino L and Boccaccio C:
Known and novel roles of the MET oncogene in cancer: A coherent
approach to targeted therapy. Nat Rev Cancer. 18:341–358. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Organ SL and Tsao MS: An overview of the
c-MET signaling pathway. Ther Adv Med Oncol. 3 (1 Suppl):S7–S19.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Drilon A, Cappuzzo F, Ou SI and Camidge
DR: Targeting MET in lung cancer: Will expectations finally be MET?
J Thorac Oncol. 12:15–26. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Deying W, Feng G, Shumei L, Hui Z, Ming L
and Hongqing W: CAF-derived HGF promotes cell proliferation and
drug resistance by up-regulating the c-Met/PI3K/Akt and GRP78
signalling in ovarian cancer cells. Biosci Rep. 37:BSR201604702017.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Khan KH, Yap TA, Yan L and Cunningham D:
Targeting the PI3K-AKT-mTOR signaling network in cancer. Chin J
Cancer. 32:253–265. 2013. View Article : Google Scholar : PubMed/NCBI
|