Introduction
Lung cancer is one of the most common types of
cancer worldwide (1). The incidence
and mortality rates of lung cancer in the United States in 2019 are
estimated to be 13 and 23.5%, respectively (2). The relative 5-year survival rate of
lung cancer was 19% in the United States between 2008–2014
(1). The onset of lung cancer has
been associated with genetic and epigenetic abnormalities (1). Although significant progress has been
made during the last few decades, the molecular mechanisms
underlying the development and progression of lung cancer remain
partially unknown (1). Thus,
identifying novel therapeutic targets for lung cancer is urgently
required to provide personalized therapy to patients with specific
molecular subtypes.
Autophagy is an important cellular process, where
the intracellular damaged organelles and misfolded proteins are
degraded and reused for cell metabolism, thus maintaining cellular,
tissue and organismal homeostasis (3). mTOR is a negative regulator of
autophagy, which is activated in response to both nutrients and
growth factors (4). Furthermore, AKT
links receptor tyrosine kinases to mechanistic target of rapamycin
complex 1 (mTORC1), thereby repressing autophagy in response to
insulin-like and other growth factor signals (4). Autophagy has dual roles in cancer. On
the one hand, autophagy plays a significant role in tumor
suppression; however, it also promotes cancer cell survival during
metabolic stress (5).
Increasing evidence has suggested that RNA
post-synthesis modifications control RNA folding, stability and
function (6). In total, >100
different types of post-synthesis modifications have been
identified on RNA, including transfer RNA (tRNA), ribosomal RNA
(rRNA), messenger RNA (mRNA) and microRNA (miRNA) (6). All four RNA bases, as well as the
ribose sugar, can be the targets of a variety of enzymes, such as
methyltransferase-like 3, transfer RNA methyltransferase 10 and DNA
methyltransferase like 2 (6).
Dysregulation of specific RNA modifications has been associated
with cancer (6). Recently, it has
been reported that fat mass and obesity-associated protein, a
demethylase of N6-methyladenosine (m6A) of
RNA, upregulates the Unc-51-like kinase 1 (ULK1), a critical
molecule of autophagy, thus promoting the initiation of autophagy
in HeLa and HEK293 cells (7).
However, whether other types of RNA modification-related proteins
affect autophagy remains unknown.
Methyltransferase-like 1 (METTL1) is a tRNA and
miRNA modification enzyme, which forms a complex with WD repeat
domain 4 to catalyze the 7-methylguanosine (m7G)
modification of tRNA and miRNA in mammalian cells (8,9).
Previous studies have demonstrated that METTL1 is phosphorylated at
Ser27 and inactivated by AKT and ribosomal S6 kinase (S6K)
(10), and serves a positive role in
murine embryonic stem cell proliferation (8). A study revealed that silencing METTL1
increases sensitivity of HeLa cells to 5-fluorouracil (11). Furthermore, METTL1 promotes miRNA
processing in A549 cells to suppress cell migration (9), whereas its overexpression promotes
hepatocellular carcinoma (HCC) and is associated with poor
prognosis (12).
In the present study, METTL1 expression was assessed
in LUAD tissues and its levels were associated with the prognosis
of patients with LUAD. In addition, the molecular mechanisms
underlying the effects of METTL1 were also investigated.
Materials and methods
Plasmids and small interfering
(si)RNAs
The METTL1 coding sequence (NM_005371) fused with
FLAG coding sequence at 3′-terminal was synthesized by Shanghai
ShineGene Molecular Biotech, Inc., and sub-cloned into the
pFLAG-CMV-4 vector, which was kindly provided by Dr Xianqiong Zou
(Guilin Medical University). The METTL1 siRNA sequence (si-METTL1,
5′-GGACAUCUAGGCACCUCAATT-3′) and control siRNA sequence
(si-control, 5′-UUGAGGUGCCUAGAUGUCCTT-3′) were synthesized by
Shanghai GenePharma Co., Ltd.
Cell culture and transfection
The LUAD cell lines, A549 and H1993 were purchased
from the Kunming Cell Bank of the Chinese Academy of Sciences.
Cells were cultured and transfected as previously described
(13). Briefly, cells were cultured
in RPMI-1640 medium (Thermo Fisher Scientific, Inc.) supplemented
with 10% fetal bovine serum (Thermo Fisher Scientific, Inc.) at
37°C in a humidified atmosphere of 5% CO2.
A549 and H1993 cells were transfected at 37°C for 48
h using Lipofectamine® 3000 reagent (Thermo Fisher
Scientific, Inc.), according to the manufacturer's instructions.
For the overexpression experiments, cells were transfected with 1
µg of METTL1 overexpression plasmid or pFLAG-CMV-4 empty vector.
For the METTL1 knockdown experiments, A549 and H1993 cells were
transfected with 0.25 nmol si-METTL1 or si-control. Cells were
harvested for subsequent experimentation 48 h
post-transfection.
Autophagy assays
A549 cells were transfected at 37°C for 48 h, and
then cells were harvested to detect autophagy markers LC3 and
p62/sequestosome 1 (SQSTM1) using western blot (14). Autophagy inhibitor bafilomycin A1
(cat. no. tlrl-baf1, InvivoGen) was added into culture media (final
concentration 100 nM) and incubated at 37°C for 1 h prior to
harvesting cells in autophagy inhibition experiment. GFP-LC3 stable
transformant HCC827 cells were transiently transfected with
si-METTL1 or control siRNA for 48 h. The cells were fixed with 2%
paraformaldehyde at room temperature for 10 min, followed by
washing with PBS three times. Cells were observed under a
fluorescent microscope (magnification, ×400, Zeiss Axio Imager Z2,
Carl Zeiss Microscopy, LLC), and the GFP-LC3 fluorescent puncta
(autophagosome) were counted manually (14).
Western blotting
Western blotting was performed as previously
described (15). Briefly, A549 and
H1993 cells were harvested and lysed with lysis buffer (cat. no.
R0020; Beijing Solarbio Science & Technology Co., Ltd.) on ice
for 5 min. Subsequently, the lysate was centrifuged at 13,500 × g
for 15 min at 4°C to obtain the supernatant. Protein concentration
was measured using a BCA kit (cat. no. P0012, Beyotime Institute of
Biotechnology), according to the manufacturer's instructions.
Proteins (30 µg) were separated by 12 or 15% SDS-PAGE gel,
transferred onto polyvinylidene fluoride membranes and blocked with
5% non-fat milk at room temperature for 2 h. The membranes were
incubated with the following primary antibodies: Rabbit anti-METTL1
antibody (cat. no. 14994-1; 1:2,000; ProteinTech Group, Inc.),
rabbit anti-light chain (LC) 3B polyclonal antibody (cat. no.
NB100-2220; 1:1,000; Novus Biologicals, LLC), rabbit anti-p62
polyclonal antibody (cat. no. 39749; 1:1,000), anti-AKT (cat. no.
9272; 1:1,000), anti-phosphorylated (p)-AKT (cat. no. 4060;
1:1,000), anti-S6K (cat. no. 9234; 1:1,000) and anti-p-S6K (cat.
no. 9204; 1:1,000), overnight at 4°C (all purchased from Cell
Signaling Technology, Inc). Following the primary incubation,
membranes were incubated with secondary antibodies (horseradish
peroxidase-conjugated goat anti-rabbit immunoglobulin G (IgG; cat.
no. G-21234; 1:5,000) or goat anti-mouse IgG (cat. no. G-21040;
1:5,000) at room temperature for 1 h (both purchased from Pierce;
Thermo Fisher Scientific, Inc.). Protein bands were developed using
the enhanced chemiluminescence kit (cat. no. P0018; Beyotime
Institute of Biotechnology).
Cell proliferation assay
Cell proliferation was determined via the Cell
Counting Kit-8 (CCK-8; cat. no. CK04; Dojindo Molecular
Technologies, Inc.) assay, according to the manufacturer's
instructions. Briefly, A549 cells were seeded into 96-well plates
at a density of 3×103 cells/well. Subsequently, 10 µl
CCK-8 solution was added to each well at 0, 24, 48, 72 and 96 h,
and cells were incubated at 37°C for 1 h. The optical density (OD)
values were analyzed at a wavelength of 450 nm, using a microplate
reader (EPOCH; BioTek Instruments, Inc.).
Colony formation assay
Cells were seeded into 6-well plates (600 cells/well
for overexpression in A549, and 800 cells/well for other
experiments), and 2 ml RPMI-1640 medium supplemented with 10% fetal
bovine serum (both purchased from Thermo Fisher Scientific, Inc.)
was added into each well. Cells were cultured for 10–15 days at
37°C in a humidified atmosphere of 5% CO2. Following
incubation, cells were washed three times with PBS and fixed with
4% neutral paraformaldehyde solution at room temperature for 30
min. Cells were re-washed three times with PBS and 2 ml of 1%
crystal violet solution was added into each well at room
temperature for an additional 2 h. Cells were washed three times
with PBS, the plates were dried and scanned with Epson Perfection
V370 Photo scanner. Cell colonies were counted manually.
METTL1 expression in LUAD tissues and
prognosis analysis
The METTL1 expression profiles in LUAD tissues and
normal tissues were obtained from The Cancer Genome Atlas (TCGA,
http://portal.gdc.cancer.gov) dataset
and GSE10072 dataset in the Gene Expression Omnibus (GEO) database
(https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE10072)
(16). The TCGA LUAD results were
analyzed using the online software Gene Expression Profiling
Interactive Analysis (GEPIA; http://gepia.cancer-pku.cn). The METTL1 expression
profiles in LUAD tissues and normal tissues of the GSE10072 dataset
were analyzed using Prism 5.01 software (GraphPad Software, Inc.).
METTL1 protein expression at in LUAD tissues was assessed using via
immunohistochemical staining results downloaded from The Human
Protein Atlas (https://www.proteinatlas.org). Survival analysis of
patients with LUAD was performed using the Kaplan-Meier Plotter
online software (http://kmplot.com). The log-rank test
was used to determine survival probability. In addition, the
expression of METTL1 at different stages was assessed using the
UALCAN database (http://ualcan.path.uab.edu).
TCGA dataset and Gene Set Enrichment
Analysis (GSEA)
The TCGA-LUAD gene expression dataset
(TCGA-LUAD.htseq_fpkm.tsv) was downloaded from the Xenahubs
database (https://gdc.xenahubs.net/download/TCGA-LUAD.htseq_fpkm.tsv.gz).
Tumor samples were classified into high- and low-METTL1 groups
using the median METTL1 expression value as the cut-off. GSEA was
performed using GSEA 4.0.3 software (http://www.broad.mit.edu/gsea), which was applied with
the predefined gene sets. The number of permutations was set at
1,000, and a gene set was considered significantly enriched with a
False Discovery Rate score <0.25.
Functional analysis of GSE112180
dataset
The top 250 differentially expressed genes from the
GEO dataset GSE112180 were identified by comparing the shCTRL and
shMETTL1 groups using GEO2R (https://www.ncbi.nlm.nih.gov/geo/geo2r/?acc=GSE112180).
The identified genes were uploaded onto the Database for
Annotation, Visualization and Integrated Discovery (DAVID,
http://david.ncifcrf.gov/tools.jsp).
The genes were analyzed using functional annotation tool of DAVID
to determine their association with signaling pathways. The related
pathways were graphed using the gene counts (17).
Statistical analysis
Statistical analysis was performed using GraphPad
Prism 5 software (GraphPad Software, Inc.) and SPSS 19.0 software
(IBM Corp.). All experiments were performed independently at least
three times and data are presented as the mean ± standard
deviation. Unpaired Student's t-test was used to compare
differences between two groups, while one-way ANOVA followed by
Tukey's post hoc test were used to compare differences between
multiple groups. The log-rank test was used to determine survival
probability. The χ2 test was used to assess the
association between METTL1 expression and the clinicopathological
characteristics of patients with LUAD. P<0.05 was considered to
indicate a statistically significant difference.
Results
METTL1 expression is upregulated in
LUAD tissues, and its high expression is associated with
unfavorable prognosis
The TCGA-LUAD dataset was analyzed using GEPIA. The
results demonstrated that METTL1 expression was significantly
upregulated in LUAD tissues (n=483) compared with normal lung
tissues (n=59) (P<0.05; Fig. 1A).
In addition, METTL1 expression was significantly higher in LUAD
samples at different clinical stages compared with those in normal
lung tissues (P<0.001; Fig. 1B).
The expression of METTL1 in LUAD samples was also confirmed using
data from another study (GSE10072) in the Gene Expression Omnibus
(GEO) database (16) (Fig. 1C and D). As METTL1 expression from
the TCGA-LUAD and GSE10072 datasets was at RNA level, its
expression at protein level was further assessed via
immunohistochemistry staining, using the Human Protein Atlas
database. The results demonstrated that there were five LUAD
samples with METTL1 positive staining (2 medium-, and 3
low-staining samples) among nine assessed lung cancer samples,
which confirmed that METTL1 protein expression is upregulated in
LUAD (Fig. 1E).
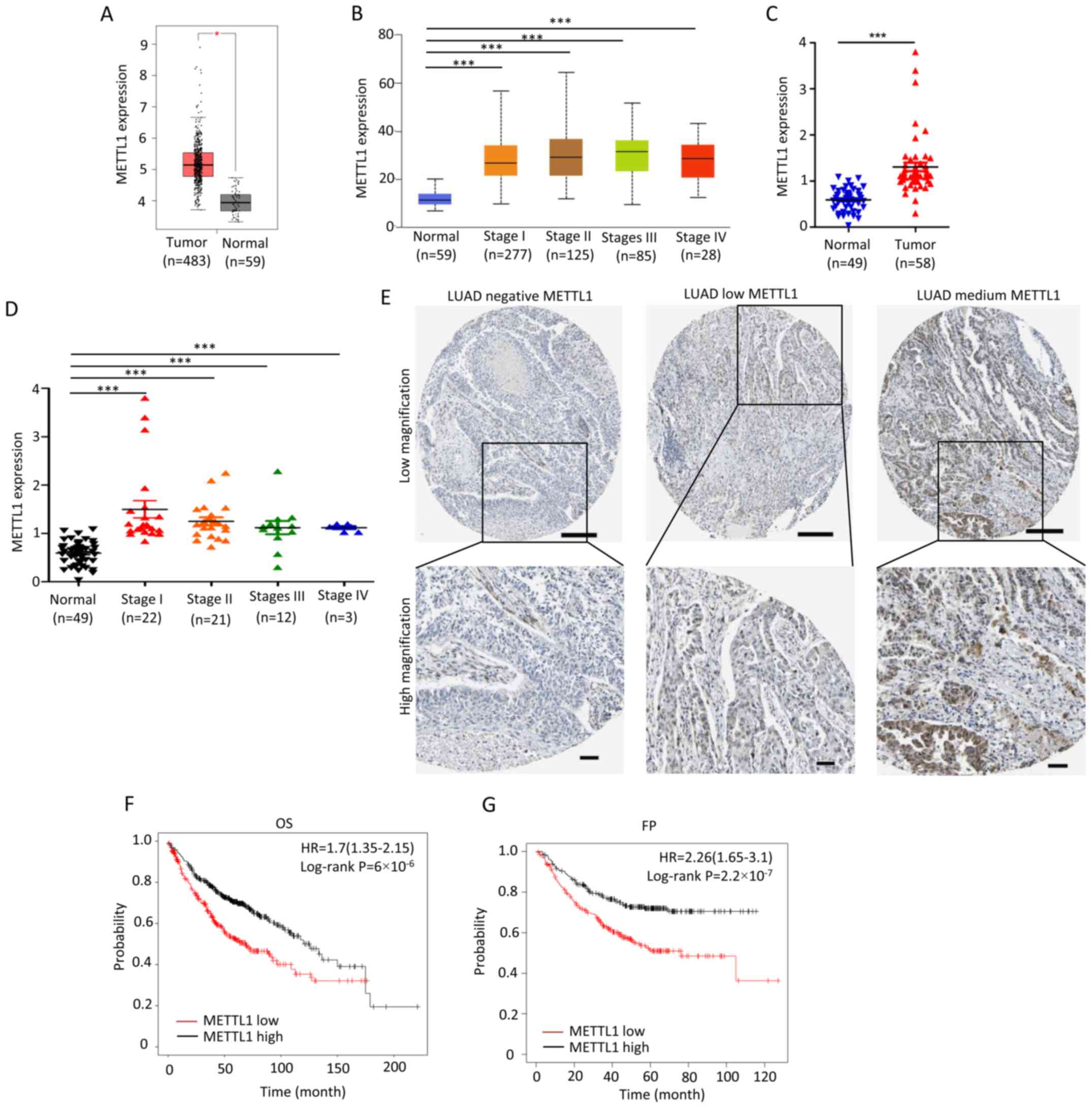 | Figure 1.METTL1 expression is upregulated in
LUAD, and its increased expression is associated with unfavorable
OS and FP. (A) METTL1 expression profile in LUAD tissues (n=483)
and normal lung tissues (n=59) from TCGA database was analyzed
using Gene Expression Profiling Interactive Analysis software.
METTL1 expression was significantly upregulated in LUAD tissues.
(B) METTL1 expression was significantly higher in different LUAD
stages compared with normal tissues. No significant differences
were observed among the different tumor stages. The METTL1
expression profile in different LUAD stages (stage I, n=277; stage
II, n=125; stage III, n=85 and stage IV, n=28) and normal lung
tissues (n=59) from TCGA database was analyzed using UALCAN
software. (C) The METTL1 expression profile in LUAD tissues (n=58)
and normal lung tissues (n=49) from the GSE10072 dataset in the GEO
database was analyzed using GraphPad Prism 5.0 software. METTL1
expression was significantly upregulated in LUAD tissues. (D)
METTL1 expression was significantly higher in different LUAD stages
compared with normal tissues. No significant differences were
observed among the different stages. The METTL1 expression profile
in different stages (stage I, n=22; stage II, n=21; stage III, n=12
and stage IV, n=3) and normal lung tissues (n=49) from the GSE10072
dataset in the GEO database was analyzed using GraphPad Prism 5.0
software. (E) METTL1 expression was upregulated in LUAD tissues.
Representative images of immunohistochemistry staining from the
Human Protein Atlas database. Scale bar at low magnification, 200
µM; high magnification, 50 µM. HPA Patients’ ID, negative: 2268,
medium: 3052, low: 2403. (F) High METTL1 expression was associated
with unfavorable OS. The OS curve of 719 patients was plotted
[cut-off value, 347; HR=1.7 (1.35–2.15), 95% CI; log-rank
P=6×10−6−05]. (G) High METTL1 expression was associated
with unfavorable FP. The FP curve of 461 patients was plotted
[cut-off value, 324; HR=2.26 (1.65–3.1), 95% CI; log-rank
P=2.2×10−7]. The online Kaplan Meier Plotter software
was used to construct the OS and FP graphs. *P<0.05,
***P<0.001. METTL1, methyltransferase-like 1; LUAD, lung
adenocarcinoma; OS, overall survival; FP, first progression; TCGA,
The Cancer Genome Atlas; GEO, Gene Expression Omnibus; HR, hazard
ratio; CI, confidence interval. |
To determine the association between METTL1
expression and the clinicopathological characteristics of patients
with LUAD, patients in the TCGA-LUAD dataset (Please note: the
value of some characteristics was absent in the original TCGA LUAD
dataset, so the number of patients for different characteristics
varies) were classified into METTL1 low or high groups, with the
median METTL1 expression value set as the cut-off. The
χ2 test was used to assess the association between
METTL1 expression and the clinicopathological characteristics of
patients with LUAD, and the results demonstrated that METTL1 was
only associated with the M stage (M1 vs. M0; P<0.01; Table I).
 | Table I.Association between METTL1 expression
and the clinicopathological characteristics of patients with lung
adenocarcinoma (n=517). |
Table I.
Association between METTL1 expression
and the clinicopathological characteristics of patients with lung
adenocarcinoma (n=517).
|
|
|
| METTL1
expression |
|
|
|---|
|
|
|
|
|
|
|
|---|
| Characteristics | Variables | Number of patients,
n | Low, n | High, n | χ2 | P-value |
|---|
| Age, years | <60 | 137 | 69 | 68 | 0.0335 | 0.8549 |
|
| ≥60 | 362 | 179 | 183 |
|
|
| Sex | Female | 277 | 140 | 137 | 0.0972 | 0.7552 |
|
| Male | 240 | 118 | 122 |
|
|
| T | T1 | 168 | 90 | 78 | 1.3390 | 0.2471 |
|
| T2/3/4 | 349 | 168 | 181 |
|
|
| N | N0 | 332 | 169 | 163 | 0.5588 | 0.4547 |
|
| N1/2/3 | 173 | 82 | 91 |
|
|
| M | M0 | 350 | 173 | 177 | 8.1040 |
0.0044a |
|
| M1 | 25 | 5 | 20 |
|
|
| Stage | Stage 1 | 276 | 142 | 134 | 3.6700 | 0.0554 |
|
| Stage 2/3/4 | 213 | 91 | 122 |
|
|
| DLCO predictive
percent | <80 | 124 | 68 | 56 | 0.8760 | 0.3493 |
|
| ≥80 | 75 | 36 | 39 |
|
|
| Bronchondilator FEV
1% | <80 | 62 | 31 | 31 | 0.0501 | 0.8229 |
|
| ≥80 | 79 | 38 | 41 |
|
|
| Location in lung
parenchyma | Central lung | 64 | 34 | 30 | 0.0023 | 0.9615 |
|
| Peripheral
lung | 127 | 67 | 60 |
|
|
| Person neoplasm
cancer status | Tumor free | 311 | 156 | 155 | 0.1945 | 0.6592 |
|
| With tumor | 109 | 52 | 57 |
|
|
| Smoking history
indicator | 1/2 | 197 | 96 | 101 | 0.1221 | 0.7268 |
|
| 3/4/5 | 308 | 155 | 153 |
|
|
| Packs of cigarettes
per year | <40 | 175 | 86 | 89 | 0.1813 | 0.6702 |
|
| ≥40 | 177 | 91 | 86 |
|
|
The clinical value of METTL1 on disease prognosis
was assessed by analyzing the overall survival (OS), first
progression (FP) and disease-free survival (DFS) of patients with
LUAD, using the Kaplan-Meier Plotter software. Patients were
divided into METTL1-high and METTL1-low groups, with the median
METTL1 mRNA value set as the cut-off. The results demonstrated that
patients in the METTL1-low group had improved OS and FP rates
compared with those in the METTL1-high group (Fig. 1F and G; P<0.0001). However, no
significant difference was observed in DFS between the two groups
(data not shown). The Cox regression model was used to perform
multivariate analysis of METTL1 expression on OS and FP. The
results demonstrated that METTL1 expression was an independent
indicator for OS, but not for FP (Table
II). In addition, AJCC stage T was an independent indicator for
OS and FP, and AJCC stage N was an independent indicator for OS
only (Table II) (18).
| Table II.Cox regression multivariate analysis
of METTL1 on OS and FP of patients with lung adenocarcinoma. |
Table II.
Cox regression multivariate analysis
of METTL1 on OS and FP of patients with lung adenocarcinoma.
|
|
Characteristics | Hazard ratio (95%
CI) | P-value |
|---|
| OS | Sex | 1.54
(0.89–2.66) | 0.1270 |
|
| Stage | 0.41
(0.15–1.11) | 0.0795 |
|
| AJCC stage T | 2.09
(1.12–3.89) |
0.0204a |
|
| AJCC stage N | 2.64
(1.03–6.75) |
0.0434a |
|
| Smoking
history | 1.96
(0.44–8.63) | 0.3700 |
|
| METTL1 | 2.07
(1.24–3.46) |
0.0056b |
| FP | Sex | 1.33
(0.71–2.48) | 0.3718 |
|
| Stage | 1.37
(0.14–13.89) | 0.7883 |
|
| AJCC stage T | 3.11
(1.30–7.44) |
0.0109a |
|
| AJCC stage N | 2.14
(0.21–21.41) | 0.5185 |
|
| Smoking
history | 1.51
(0.73–3.13) | 0.2642 |
|
| METTL1 | 1.54
(0.82–2.9) | 0.1836 |
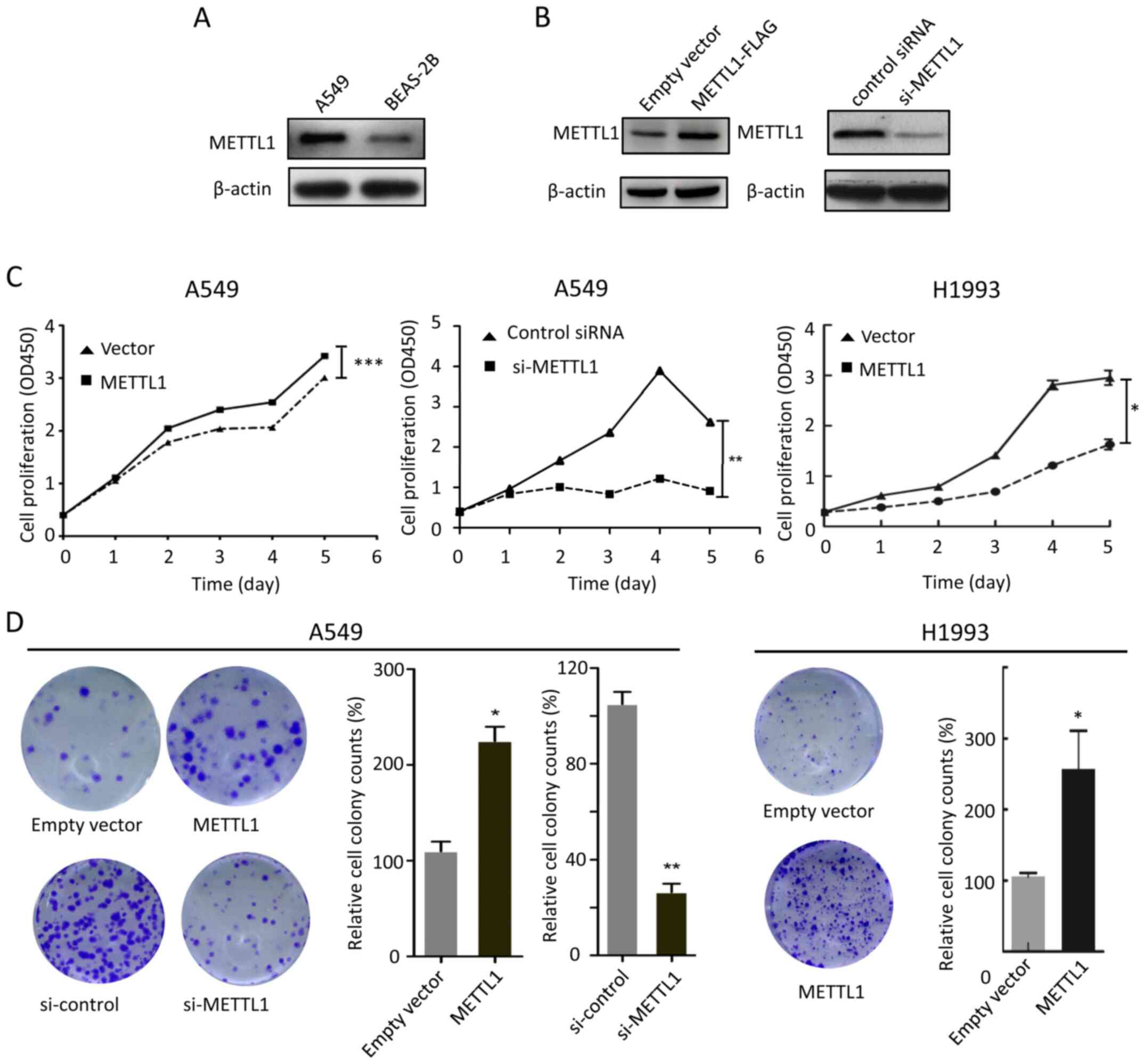
METTL1 promotes colony formation and
A549 cell proliferation
To investigate the role of METTL1 in vitro, western
blot analysis was performed to detect METTL1 protein expression in
the A549 lung cancer cell line. As presented in Fig. 2A, METTL1 was upregulated in A549
cells compared with BEAS-2B normal lung epithelial cells. Thus, the
A549 cell line was used for subsequent experiments. A549 cells were
subsequently transfected with METTL1 plasmids or si-METTL1 for
METTL1 overexpression and silencing experiments, respectively.
Western blot analysis demonstrated that METTL1 protein expression
increased and decreased in the overexpression and knockdown
experiments, respectively (Fig. 2B).
The effect of METTL1 overexpression and silencing in A549 cell
proliferation and colony formation was investigated. The results
demonstrated that A549 cell proliferation increased following
METTL1 overexpression, the effects of which were attenuated
following transfection with si-METTL1 (Fig. 2C). In addition, METTL1 overexpression
increased the colony count, which decreased in METTL1-silenced A549
cells (Fig. 2D). The effects of
METTL1 on cell proliferation and colony formation were further
confirmed in LUAD H1993 cells by overexpressing METTL1 (Fig. 2C and D).
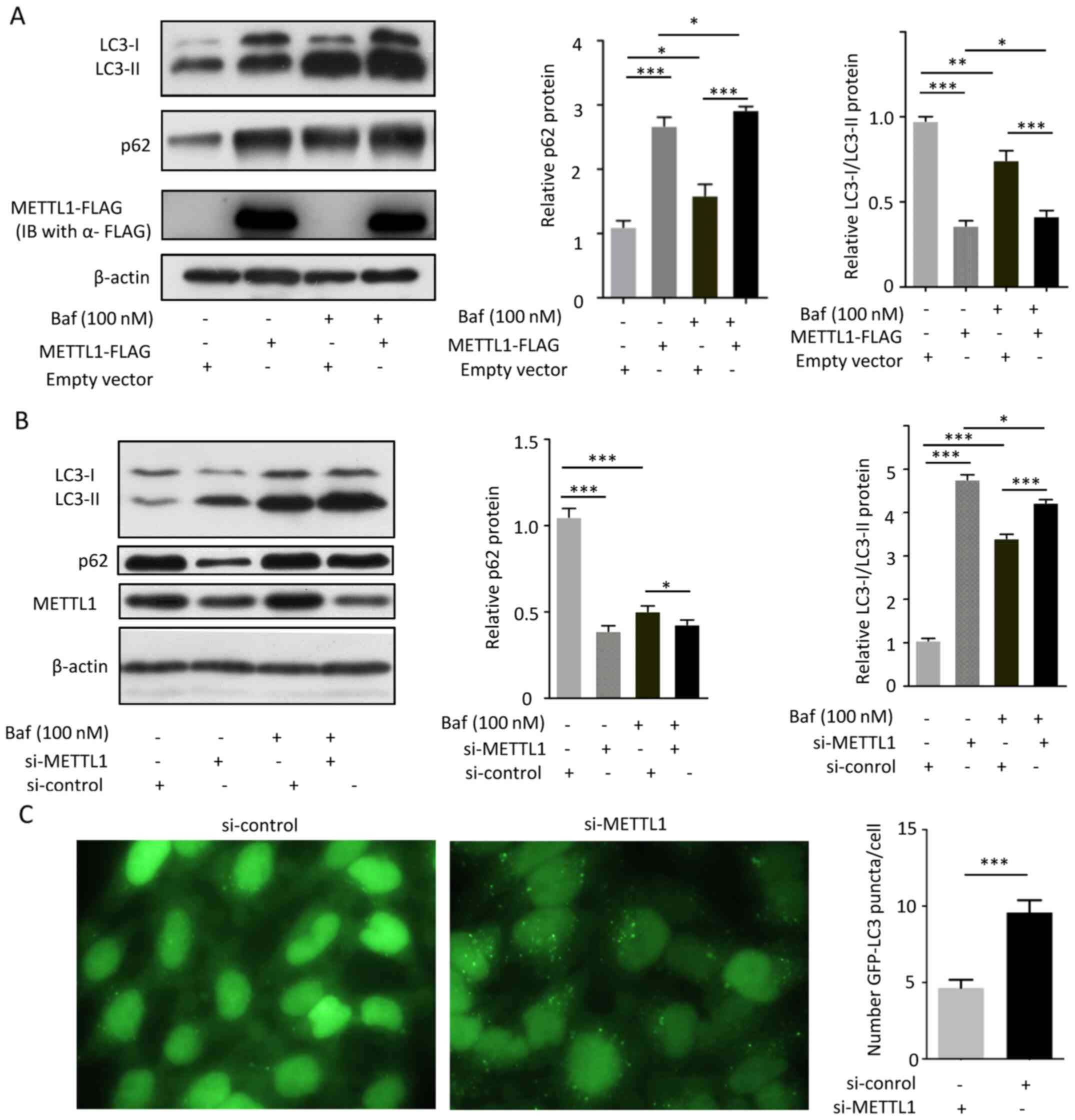
METTL1 inhibits autophagy in A549
cells
It is well-known that LC3B conversion (LC3-I to
LC3-II) and lysosomal degradation of LC3-II reflect the progression
of autophagy. In addition, it has been reported that
p62/sequestosome 1 (SQSTM1) protein is degraded by autophagy
(14). Thus, LC3B and p62 are
considered markers of autophagy (14). To assess the effects of METTL1 on
autophagy, A549 cells were transfected with METTL1 overexpression
plasmids at 37°C for 48 h and were subsequently treated with the
autophagy inhibitor, bafilomycin A1 (100 nM) at 37°C for 1 h. Cells
were subsequently harvested, cell lysates were prepared and the
protein expression levels of LC3B and p62 were determined in the
lysates via western blot analysis. The results demonstrated that
METTL1 overexpression suppressed the conversion of LC3-I to LC3-II,
turnover of LC3-II and degradation of p62/SQSTM1. Furthermore, the
inhibitory effect of METTL1 was enhanced following treatment with
bafilomycin A1 (Fig. 3A), implying
that METTL1 may inhibit autophagy. To confirm these findings, A549
cells were transfected with si-METTL1 at 37°C for 48 h and
subsequently treated with 100 nM bafilomycin A1 at 37°C for 1 h.
The protein expression levels of LC3B and p62/SQSTM1 was detected
via western blot analysis. The results demonstrated that METTL1
knockdown promoted the conversion of LC3-I to LC3-II and
degradation of p62/SQSTM1. LC3-II and p62/SQSTM1 levels accumulated
following treatment with bafilomycin A1 in the si-METTL1 group
compared with the control group (Fig.
3B), suggesting that METTL1 knockdown may enhance autophagy. To
confirm the effects of METTL1 on autophagy, HCC827 cells stably
expressing GFP-LC3 were transfected with si-METTL1 and fluorescent
puncta (autophagosome) were observed under a fluorescent microscope
and quantified. The results demonstrated that silencing METTL1
enhanced autophagy (Fig. 3). Taken
together, the overexpression and silencing experiments indicated
that METTL1 may inhibit autophagy.
METTL1 activates AKT/mTORC1
signaling
To determine the molecular mechanism underlying the
effect of METTL1 on the inhibition of autophagy, GSEA was performed
using the TCGA-LUAD dataset. The results demonstrated that the gene
sets of the phosphatidylinositol 3-kinase (PI3K)/AKT/mTOR and
MTORC1 signaling pathways were enriched in the high-METTL1
expression group (Fig. 4A and B).
Furthermore, the GSE112180 dataset was analyzed using GEO2R, and
the top 250 differentially expressed genes were identified. These
genes were analyzed using the Database for Annotation,
Visualization and Integrated Discovery (DAVID). The results
revealed that PI3K/AKT/mTOR signaling was activated by METTL1
(Fig. 4C), which was consistent with
the GSEA results. Subsequently, cellular experiments were performed
to determine the effects of METTL1 on the aforementioned signaling
pathways. A549 cells were transfected with METTL1 overexpression
plasmids or si-METTL1 37°C for 48 h. Following transfection, the
cells were collected and protein lysates were prepared and resolved
via western blot analysis. METTL1 overexpression notably increased
the expression levels of p-AKT and p-S6K, while the total protein
levels of AKT and S6K remained unchanged compared with those in the
empty vector group. Conversely, knockdown of METTL1 downregulated
p-AKT and p-S6K protein expression, while the total protein levels
of AKT and S6K remained unchanged compared with those in the
si-control group. Taken together, these results suggest that METTL1
may activate the AKT/mTORC1 signaling pathway.
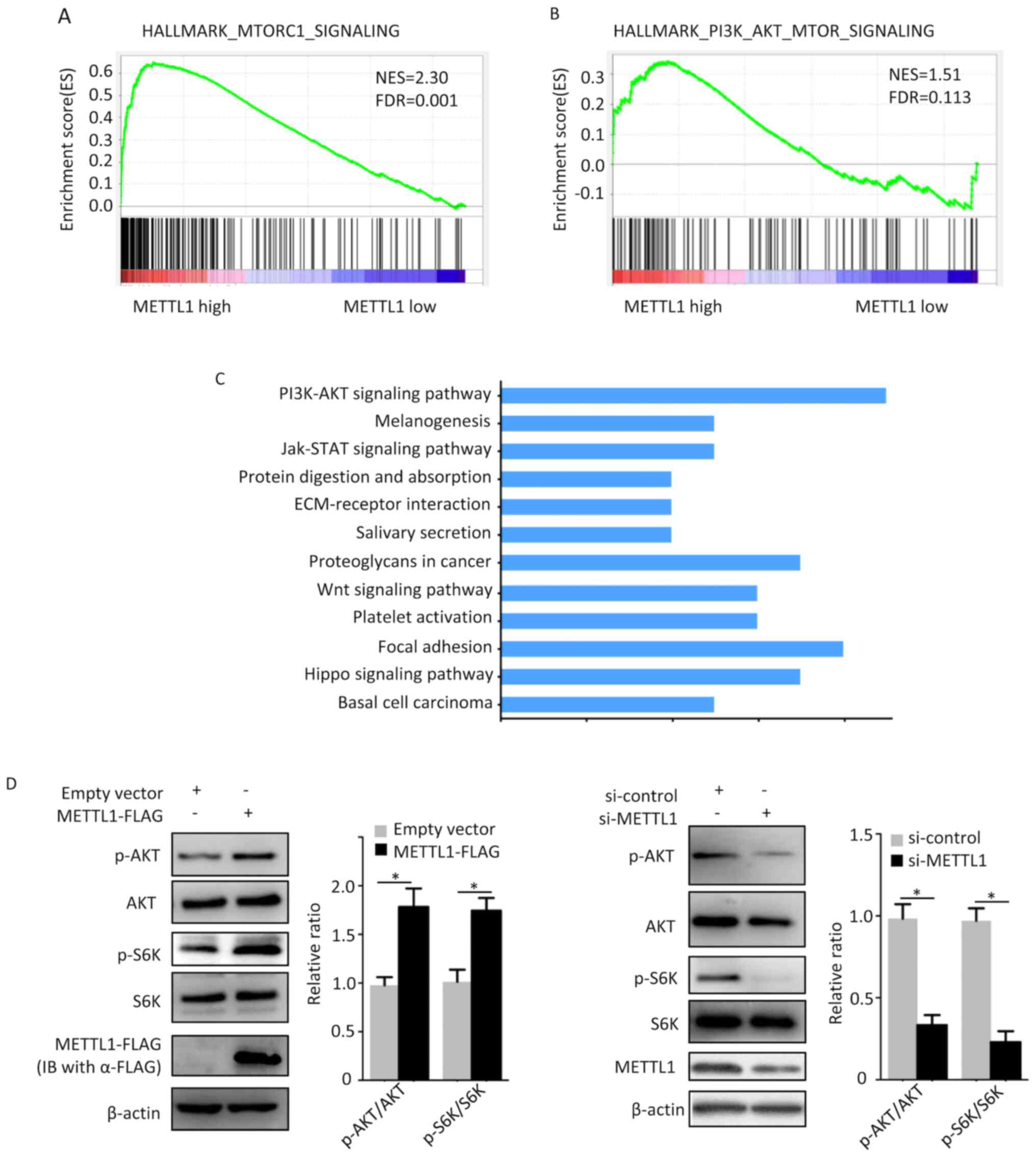 | Figure 4.METTL1 activates the AKT/mTORC1
signaling pathway. GSEA analysis of the TCGA-LUAD dataset revealed
that the gene sets (A) HALLMARK_MTORC1_SIGNALING and (B)
HALLMARK_PI3K_AKT_MTOR_SIGNALING were enriched in the high-METTL1
expression group. (C) DAVID analysis of the GSE112180 dataset
indicated that the gene cluster of the PI3K/AKT signaling pathway
was enriched in the high-METTL1 expression group (D) METTL1
overexpression increased the protein levels of p-AKT and p-S6K, the
effects of which were reversed following METTL1 silencing. Total
protein levels of AKT and S6K remained unchanged compared with the
control group. *P<0.05. METTL1, methyltransferase-like 1; p-AKT,
phosphorylated protein kinase B; mTORC1, mechanistic target of
rapamycin complex 1; GSEA, Gene Set Enrichment Analysis; TCGA, The
Cancer Genome Atlas; LUAD, lung adenocarcinoma; DAVID, Database for
Annotation, Visualization and Integrated Discovery; PI3K,
phosphatidylinositol 3-kinase; si, small interfering. |
Discussion
It has been reported that METTL1 catalyzes the
m7G modification of tRNA and miRNA (8,9).
Previous studies investigated the effect of METTL1 in different
types of cancer cells; however, their findings were controversial
(12,18). Tian et al (12) demonstrated that METTL1 is upregulated
in HCC at both the mRNA and protein levels, whereby increased
levels are associated with poor outcomes. Furthermore, METTL1 was
reported to promote HCC cell proliferation and migration in vitro,
suggesting that METTL1 acts as an oncogene in HCC. Conversely, Liu
et al (19) reported that
METTL1 is downregulated in colon cancer, thus acts as a tumor
suppressor molecule. The present study investigated METTL1
expression in two different LUAD cohorts obtained from two
databases. The results demonstrated that METTL1 was significantly
upregulated in LUAD, which was consistent with the results on HCC
(12). In addition, METTL1
expression did not significantly changed from LUAD stage I to stage
IV, indicating that METTL1 upregulation occurs early during LUAD
tumorigenesis. Taken together, these results suggest that METTL1
may be a potential diagnostic marker for LUAD. Although survival
analysis demonstrated that high METTL1 expression was associated
with unfavorable OS and FP, multivariate analysis indicated that
METTL1 was an independent indicator for OS, but not for FP.
The present study demonstrated that knockdown of
METTL1 in A549 cells inhibited cell proliferation and colony
formation, whereas METTL1 overexpression in the same cell line had
the opposite effects. These findings were in agreement with the
aforementioned study on HCC (12).
Autophagy is a lysosomal-related degradation pathway (3). In response to microenvironmental
stimuli, cells can eliminate proteins and lipids through autophagy
in order to maintain cellular homeostasis (3). Furthermore, autophagy may act as a
tumor-suppressing mechanism during tumor initiation, or as a
tumor-promoting process in established tumors (20). A study demonstrated that METTL1
exerts inhibitory effects on the phosphatase and tensin homolog
(PTEN) signaling (12). Given that
PTEN is considered a key inhibitory factor upstream mTORC1, which
acts as a suppressor of autophagy, the present study hypothesized
that METTL1 may play a role in the regulation of autophagy. Thus,
the effects of METTL1 knockdown and overexpression on autophagy
were investigated. The results demonstrated that METTL1 knockdown
and overexpression enhanced and inhibited autophagy, respectively.
To determine the molecular mechanisms underlying the effects of
METTL1, bioinformatics analysis was performed followed by
verification with in vitro experiments. The results demonstrated
that overexpression of METTL1 promoted activation of the AKT/mTORC1
signaling pathway. AKT and mTORC1 serve a pivotal role in cell
proliferation, and both proteins are hyperactivated in several
types of cancer, such as lymphoma, breast cancer and lung cancer
(21,22). As expected, the results of the
present study demonstrated that high METTL1 expression in LUAD was
associated with unfavorable prognosis. Taken together, the present
study provided cellular and mechanistic evidence to elucidate the
role of METTL1 in LUAD tumorigenesis. To the best of our knowledge,
the present study was the first to investigate the role of METTL1
in the regulation of autophagy and AKT/mTORC1 signaling. However,
the in vitro experiments were only performed in the A549 cell line,
thus, further experiments should be performed in more LUAD cell
lines to confirm the findings presented here.
The results of the present study on LUAD and those
of a previous study on HCC (12)
suggest that METTL1 may acts as an oncogene. Thus, it is of great
importance to address, in future studies, the molecular mechanism
underlying the regulation of METTL1 expression, and how changes in
METTL1 expression affect AKT/mTORC1 signaling and carcinogenesis.
Since METTL1 promotes miRNA processing (9), certain miRNA(s) may be associated with
METTL1 expression and AKT/mTORC1 signaling. Thus, it may be useful
to identify the miRNA(s) involved in this process.
In summary, the present study demonstrated that
METTL1 expression was upregulated in LUAD tissues, which was
associated with unfavorable prognosis. Furthermore, METTL1 may act
via the AKT/mTORC1 pathway in LUAD cells. Taken together, these
results suggest that METTL1 acts as an oncogene in LUAD and may be
a potential diagnostic biomarker and therapeutic target for
LUAD.
Acknowledgements
The authors would like to thank Dr Xianqiong Zou
from Guilin Medical University (Guilin, China) for providing the
pFLAG-CMV-4 plasmid.
Funding
The present study was supported in part by the
Hundred Talents Program of Guangxi (to GH), the Natural Science
Foundation of Guangxi (grant no. 2020JJA140139), the Research
Enhancement Project for Junior Faculty in Higher Education
Institutes of Guangxi (grant no. 2019KY0522), the Scientific
Research Project for Junior Faculty in Guilin Medical College
(grant no. 2018glmcy055) and the Guangxi Key Laboratory of
Molecular Medicine in Liver Injury and Repair (grant nos.
GXLIRMMKL-201802 and GXLIRMMKL-201816).
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
GH conceived the present study. CW, WW, XH, LD and
AL performed the experiments. GH and CW confirmed the authenticity
of all the raw data. GH and CW analyzed and interpreted the data.
CW and GH drafted the initial manuscript. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Herbst RS, Morgensztern D and Boshoff C:
The biology and management of non-small cell lung cancer. Nature.
553:446–454. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA Cancer J Clin. 69:7–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Levine B and Kroemer G: Biological
Functions of Autophagy Genes: A Disease Perspective. Cell.
176:11–42. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Levine B and Kroemer G: Autophagy in the
pathogenesis of disease. Cell. 132:27–42. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
White E: The role for autophagy in cancer.
J Clin Invest. 125:42–46. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Barbieri I and Kouzarides T: Role of RNA
modifications in cancer. Nat Rev Cancer. 20:303–322. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jin S, Zhang X, Miao Y, Liang P, Zhu K,
She Y, Wu Y, Liu DA, Huang J, Ren J, et al: m6A RNA
modification controls autophagy through upregulating ULK1 protein
abundance. Cell Res. 28:955–957. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lin S, Liu Q, Lelyveld VS, Choe J, Szostak
JW and Gregory RI: Mettl1/Wdr4-Mediated m7G tRNA
Methylome Is Required for Normal mRNA Translation and Embryonic
Stem Cell Self-Renewal and Differentiation. Mol Cell.
71:244–255.e5. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Pandolfini L, Barbieri I, Bannister AJ,
Hendrick A, Andrews B, Webster N, Murat P, Mach P, Brandi R, Robson
SC, et al: METTL1 Promotes let-7 MicroRNA Processing via
m7G Methylation. Mol Cell. 74:1278–1290.e9. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cartlidge RA, Knebel A, Peggie M,
Alexandrov A, Phizicky EM and Cohen P: The tRNA methylase METTL1 is
phosphorylated and inactivated by PKB and RSK in vitro and in
cells. EMBO J. 24:1696–1705. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Okamoto M, Fujiwara M, Hori M, Okada K,
Yazama F, Konishi H, Xiao Y, Qi G, Shimamoto F, Ota T, et al: tRNA
modifying enzymes, NSUN2 and METTL1, determine sensitivity to
5-fluorouracil in HeLa cells. PLoS Genet. 10:e10046392014.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tian QH, Zhang MF, Zeng JS, Luo RG, Wen Y,
Chen J, Gan LG and Xiong JP: METTL1 overexpression is correlated
with poor prognosis and promotes hepatocellular carcinoma via PTEN.
J Mol Med (Berl). 97:1535–1545. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang Q, Li A, Jin J and Huang G: Targeted
interfering DEP domain containing 1 protein induces apoptosis in
A549 lung adenocarcinoma cells through the NF-κB signaling pathway.
Onco Targets Ther. 10:4443–4454. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Klionsky DJ, Abdelmohsen K, Abe A, Abedin
MJ, Abeliovich H, Acevedo Arozena A, Adachi H, Adams CM, Adams PD,
Adeli K, et al: Guidelines for the use and interpretation of assays
for monitoring autophagy (3rd edition). Autophagy. 12:1–222. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li A, Wang Q, He G, Jin J and Huang G: DEP
domain containing 1 suppresses apoptosis via inhibition of A20
expression, which activates the nuclear factor κB signaling pathway
in HepG2 cells. Oncol Lett. 16:949–955. 2018.PubMed/NCBI
|
|
16
|
Landi MT, Dracheva T, Rotunno M, Figueroa
JD, Liu H, Dasgupta A, Mann FE, Fukuoka J, Hames M, Bergen AW, et
al: Gene expression signature of cigarette smoking and its role in
lung adenocarcinoma development and survival. PLoS One.
3:e16512008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Huang W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Amin MB, Greene FL, Edge SB, Compton CC,
Gershenwald JE, Brookland RK, Meyer L, Gress DM, Byrd DR and
Winchester DP: The Eighth Edition AJCC Cancer Staging Manual:
Continuing to build a bridge from a population-based to a more
‘personalized’ approach to cancer staging. CA Cancer J Clin.
67:93–99. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu Y, Zhang Y, Chi Q, Wang Z and Sun B:
Methyltransferase-like 1 (METTL1) served as a tumor suppressor in
colon cancer by activating 7-methyguanosine (m7G)
regulated let-7e miRNA/HMGA2 axis. Life Sci. 249:1174802020.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Marinković M, Šprung M, Buljubašić M and
Novak I: Autophagy Modulation in cancer: current knowledge on
action and therapy. Oxid Med Cell Longev. 2018:80238212018.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Manning BD and Toker A: AKT/PKB Signaling:
Navigating the Network. Cell. 169:381–405. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Saxton RA and Sabatini DM: mTOR Signaling
in Growth, Metabolism, and Disease. Cell 168: 960–976, 2017.
Erratum in: Cell. 169:361–371. 2017.
|