Introduction
Nasopharyngeal carcinoma (NPC) is a type of cancer
that originates from epithelial cells in the nasopharynx (1). According to GLOBOCAN 2012, 86,691 new
NPC cases and 50,831 NPC-associated deaths were reported worldwide
(2). Although NPC is relatively rare
in the rest of the world, its prevalence is relatively high in
Southern China and Southeast Asian countries (3). Epidemiological data indicate that the
NPC incidence rate has gradually decreased and the mortality rate
of NPC has significantly declined in recent years due to the
advances in diagnostic and therapeutic strategies (4). However, the 5-year survival rate of
patients with NPC remains very low due to recurrence and
metastasis, particularly in cases with late-stage disease (5). Therefore, it is necessary to identify
new and efficient methods for NPC treatment.
MicroRNAs (miRNAs/miRs) are short-chain non-coding
RNAs that are 19–24 nucleotides in length. Previous studies have
reported that miRNAs play key roles in various cellular functions,
such as proliferation, metastasis, apoptosis and chemoresistance
(6). miRNAs usually bind to the
3-untranslated region (UTR) of their target mRNAs and inhibit
translation (7). Numerous studies
have revealed that miR-223-5p is significantly downregulated in
cancer tissues and cells, suggesting its inhibitory effect on tumor
progression. For example, miR-223-5p may inhibit the progression of
non-small cell lung cancer by modulating E2F transcription factor 8
(8), suppress the malignant
phenotype of prostate cancer cells via modulating ETS transcription
factor ERG (9) and repress the
aggressiveness of bladder cancer cells (10). However, the potential regulatory role
of miR-223-5p in NPC has not been extensively investigated.
Doublecortin-like kinase 1 (DCLK1) is a kinase
highly expressed in various types of cancer (11) and has been identified as a potential
oncogene implicated in the progression of human cancers, such as
pancreatic cancer (12), lung
squamous cell carcinoma (13),
ovarian clear cell carcinoma (14),
intestinal tumors (15) and
basal-like breast cancer (16). In
addition, DCLK1 has been reported to contribute to the tumorigenic
process of colorectal cancer by downregulating miR-200c (17). However, whether DCLK1 is implicated
in NPC progression and the underlying mechanism remain to be
investigated.
The present study was undertaken to investigate
miR-223-5p and DCLK1 expression levels in NPC tissues and cells,
and to determine the effects of these factors on the viability,
migration and invasion of NPC cells. The aim was to elucidate
whether miR-223-5p can suppress NPC progression by downregulating
DCLK1 expression, in the hope of providing a novel approach to the
diagnosis and treatment of NPC.
Materials and methods
Clinical tissues
A total of 28 paired samples of NPC and adjacent
normal nasopharyngeal tissues were obtained from patients
undergoing surgery at The Third Affiliated Hospital of Soochow
University (Changzhou, China) between March 2016 and January 2019.
All the tissues were reviewed by two pathologists blinded to the
clinicopathological information and were immediately frozen in
liquid nitrogen after surgical excision. None of the patients with
NPC had received anticancer treatments prior to surgery. The
present study was approved by the Ethics Committee of The Third
Affiliated Hospital of Soochow University, and written informed
consent was obtained from each patient enrolled in the study.
Cell culture and transfection
Human NPC cell lines (6-10B, 5-8F, SUNE-1, SUNE-2
and C666-1) and a human nasopharyngeal-derived epithelial cell line
(NP69) were obtained from the Cell Bank of the Chinese Academy of
Science. Cells were cultured in DMEM (Gibco; Thermo Fisher
Scientific, Inc.) containing 12 U/l gentamicin and 100 ml/l
inactivated FBS (Gibco; Thermo Fisher Scientific, Inc.) in a
humidified atmosphere containing 5% CO2 at 37°C, and
were passaged once every 2 days.
NPC cells (5×104) were transfected with
100 nM miR-223-5p mimics (5′-UGUCAGUUUGUCAAAUACCCCA-3′), 100 nM
corresponding scrambled negative control (NC mimics;
5′-UUCUCCGAACGUGUCACGUUU-3′), 100 nM miR-223-5p inhibitor
(5′-UGGGGUAUUUGACAAACUGACA-3′), 100 nM corresponding scrambled NC
(NC inhibitor; 5′-CAGUACUUUUGUGUAGUACAA-3′), 0.2 µM small
interfering (si)RNA targeting DCLK1 (si-DCLK1;
5′-UUUCAGAGCAUACUCUCUAGC-3′), 0.2 µM scrambled siRNA NC (si-NC;
5′-UUCUCCGAACGUGUCACGUUU-3′), 2 µl pcDNA3.1 (empty vector; Ctrl) or
2 µl pcDNA3.1-DCLK1 (DCLK1) using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) at room temperature
for ~20 min. The transfected cells were cultured for 48 h and were
then used in subsequent experiments. The transfection efficiency
was confirmed by reverse transcription-quantitative (RT-q)PCR.
RT-qPCR assay
Total RNA was extracted from tissues and cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) and was reverse-transcribed into cDNA using a RT kit (Takara
Bio, Inc.) following the manufacturers protocol. Subsequently, qPCR
was performed with SYBR Select Master Mix (Thermo Fisher
Scientific, Inc.) on a ViiA™ 7 Real-Time PCR System (Thermo Fisher
Scientific, Inc.) to measure the mRNA expression levels, according
to the manufacturers instructions. The thermocycling conditions
were: 95°C for 10 min, followed by 40 cycles at 95°C for 10 sec and
55°C for 10 sec, and a final step at 72°C for 30 sec. The
2−ΔΔCq method (18) was
used to calculate the relative DCLK1 and miR-223-5p expression
levels, with GAPDH and U6 as internal controls. The primers used
were as follows: miR-223-5p forward, 5′-GCAGCGTGTATTTGACAAG-3′ and
reverse, 5′-CTCAACTGGTGTCGTGGA-3′; DCLK1 forward,
5′-CAGCGCCATCAAATACCTGC-3′ and reverse, 5′-TGGTCATCACCACTTCCACG-3′;
U6 forward, 5′-GCTTCGGCAGCACATATACTAAAAT-3′ and reverse,
5′-CGCTTCACGAATTTGCGTGTCAT-3′; and GAPDH forward,
5′-GGAGTCCACTGGCGTCTT-3′ and reverse,
5′-ATCTTGAGGCTGTTGTCATAC-3′.
Cell Counting Kit (CCK)-8 assay
After 48 h of transfection, the cells were plated
into 96-well plates at 5,000 cells per well. Then, 20 µl CCK-8
reagent (Dojindo Molecular Technologies, Inc.) was added into each
well after the cells were cultured for 0, 48, 72 and 96 h at 37°C
with 5% CO2. Subsequently, the cells were incubated at
room temperature for 4 h. The optical density of each well was
measured using a microplate reader (Bio-Rad Laboratories, Inc.) at
a wavelength of 450 nm. The cell viability was considered to be
directly proportional to the optical density of the cells in this
assay.
Transwell assay
Cell migratory and invasive abilities were analyzed
in 96-well Transwell chambers with an 8-µm pore membrane (Corning,
Inc.) following the manufacturers instructions. A total of 100 µl
cell suspension (1×105 cells/ml) was added to the upper
chamber, and 600 µl medium supplemented with 10% FBS was added to
the lower chamber. After incubation at 37°C overnight,
non-migrating cells in the upper chamber were removed with a cotton
swab, and the upper chamber was washed three times with PBS. Then,
the cells were fixed with 4% paraformaldehyde at room temperature
for 30 min and stained with 0.1% crystal violet solution at room
temperature for ~20 min. In order to observe the migrating cells
attached to the lower surface of the chamber, five fields of view
were randomly selected and the cells were counted under a light
microscope (Olympus Corporation; magnification, ×200). For the
detection of cell invasion, the upper surface of the chamber was
pre-coated with Matrigel® for 1 h at room temperature,
and the following experimental steps were the same as those
described for the detection of cell migration.
Target prediction and luciferase
reporter assay
TargetScan (http://www.targetscan.org/vert_72/) was used to
predict the binding sites between miR-223-5p and DCLK1. psiCHECK2
vectors (Promega Corporation) were used to construct
psiCHECK2-DCLK1-wild-type (WT) and psiCHECK2-DCLK1-mutant (MUT)
plasmids (containing WT and MUT 3′-UTR, respectively), according to
the manufacturers instructions. DCLK1 MUT luciferase reporter
containing MUT miR-223-5p-binding sites were produced using
GeneArt™ Site-Directed Mutagenesis System (Invitrogen; Thermo
Fisher Scientific, Inc.). Subsequently, miR-223-5p mimics were
co-transfected into 6-10B cells together with psiCHECK2-DCLK1-WT or
psiCHECK2-DCLK1-MUT using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.). After 48 h of
transfection, luciferase activity was evaluated using a
Dual-Luciferase Reporter Analysis system (Promega Corporation).
Firefly luciferase activity was normalized to Renilla
(Promega Corporation) luciferase gene activity.
Immunohistochemistry (IHC)
The tissue expression of DCLK1 was evaluated via IHC
based on the intensity and the proportion of positively stained
cells as previously described (19).
Sections were incubated with primary antibody against DCLK1
(1:1,000; cat. no. 62257; Cell Signaling Technology, Inc.)
overnight at 4°C, followed by incubation with goat anti-rabbit
secondary antibody (cat. no. ab205718; 1:2,000; Abcam) at room
temperature for 20 min. The sections were stained with
3,3-diaminobenzidine and the nuclei were counterstained with
hematoxylin for 5 min at room temperature. Images were captured
under a light microscope (Olympus Corporation; magnification,
×200).
Western blotting
Total protein was isolated from transfected cells
using RIPA lysis buffer (Beyotime Institute of Biotechnology) and
the protein concentrations were determined using a BCA kit (Thermo
Fisher Scientific, Inc.). Subsequently, 10 µg protein/lane was
separated via SDS-PAGE on 10% gels (Bio-Rad Laboratories, Inc.) and
was electrotransferred to PVDF membranes. After blocking with 5%
non-fat milk for 2 h at room temperature, the membranes were first
incubated with primary antibodies against Notch receptor 1 (Notch1;
1:1,000; cat. no. ab52627; Abcam) and GAPDH (1:1,000; cat. no.
ab8245; Abcam) overnight at 4°C, followed by incubation with
corresponding HRP-conjugated secondary antibody (1:1,000, cat. nos.
ab6789 and ab205718; Abcam) for 1 h at room temperature. Blots were
visualized with an enhanced chemiluminescent detection system (EMD
Millipore). Protein expression was measured using
Image-Pro® Plus software (version 6.0; Media
Cybernetics, Inc.).
Statistical analysis
All experiments were repeated at least 3 times. All
the data collected in the experiments were analyzed with GraphPad
Prism 7 (GraphPad Software, Inc.). Data are presented as the mean ±
SD. The association between miR-223-5p or DCLK1 expression and
clinicopathological characteristics of patients with NPC was
analyzed using χ2 tests. Both paired and unpaired
Students t-tests were used for comparisons between two groups, and
one-way ANOVA followed by Tukeys post hoc test was conducted for
comparisons among multiple groups. P<0.05 was considered to
indicate a statistically significant difference.
Results
miR-223-5p and DCLK1 expression levels
in NPC
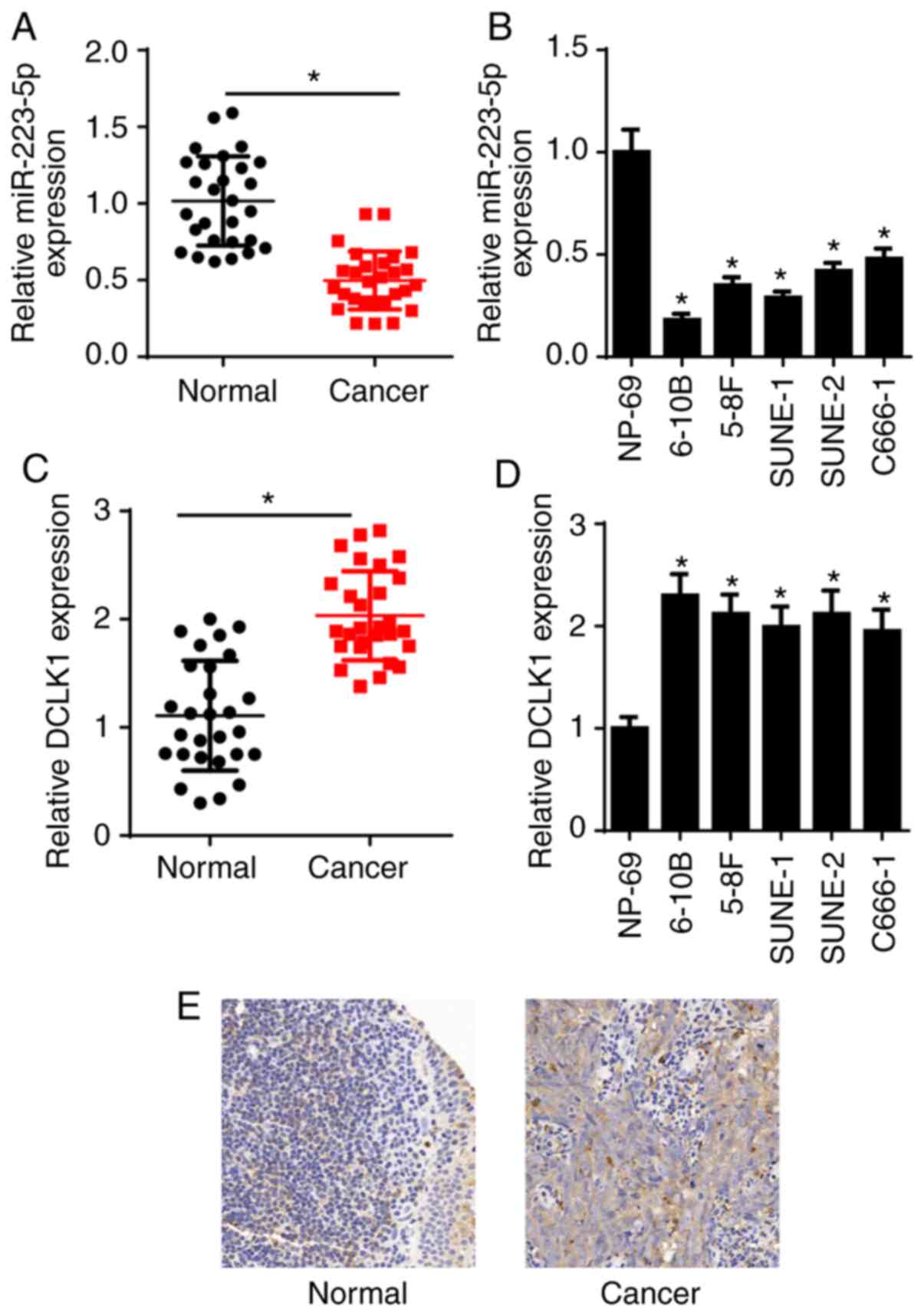
miR-223-5p expression was measured via RT-qPCR, and
the results demonstrated that miR-223-5p expression was decreased
in NPC tissues and cell lines (6-10B, 5-8F, SUNE-1, SUNE-2 and
C666-1), compared with that in adjacent normal nasopharyngeal
tissues and NP69 human nasopharyngeal-derived epithelial cells
(Fig. 1A and B). DCLK1 expression
was also examined in the present study. The RT-qPCR results
indicated that DCLK1 expression was significantly increased in NPC
tissues and cell lines (6-10B, 5-8F, SUNE-1, SUNE-2 and C666-1)
compared with that in adjacent normal nasopharyngeal tissues and
NP69 cells (Fig. 1C and D).
Moreover, the expression of DCLK1 in NPC tissues was higher
compared with that in adjacent normal nasopharyngeal tissues, as
determined via IHC (Fig. 1E). It was
observed that low miR-223-5p or high DCLK1 expression was
associated with distant metastasis and TNM stage, while there was
no significant association with sex or age in patients with NPC
(Table I). These results suggested
that miR-223-5p expression was downregulated and DCLK1 expression
was upregulated in NPC tissues and cell lines. 5-8F and 6-10B cells
were selected for subsequent experiments due to their high DCLK1
expression.
 | Table I.Association between miR-223-5p or
DCLK1 expression and clinicopathological characteristics in
nasopharyngeal carcinoma. |
Table I.
Association between miR-223-5p or
DCLK1 expression and clinicopathological characteristics in
nasopharyngeal carcinoma.
|
|
| miR-223-5p
expression | DCLK1 expression |
|---|
|
|
|
|
|
|---|
|
Characteristics | Patients, n | Low | High | P-value | Low | High | P-value |
|---|
| Sex |
|
|
| 0.625 |
|
| 0.573 |
|
Male | 17 | 9 | 8 |
| 7 | 10 |
|
|
Female | 11 | 6 | 5 |
| 5 | 6 |
|
| Age, years |
|
|
| 0.452 |
|
| 0.362 |
|
<45 | 13 | 6 | 7 |
| 5 | 8 |
|
|
≥45 | 15 | 9 | 6 |
| 7 | 8 |
|
| Distant
metastasis |
|
|
| 0.005 |
|
| 0.032 |
| No | 18 | 7 | 11 |
| 9 | 9 |
|
|
Yes | 10 | 8 | 2 |
| 3 | 7 |
|
| TNM stage |
|
|
| 0.013 |
|
| 0.011 |
| I +
II | 16 | 7 | 9 |
| 9 | 7 |
|
| III +
IV | 12 | 8 | 4 |
| 3 | 9 |
|
miR-223-5p overexpression suppresses
viability, migration and invasion of NPC cells
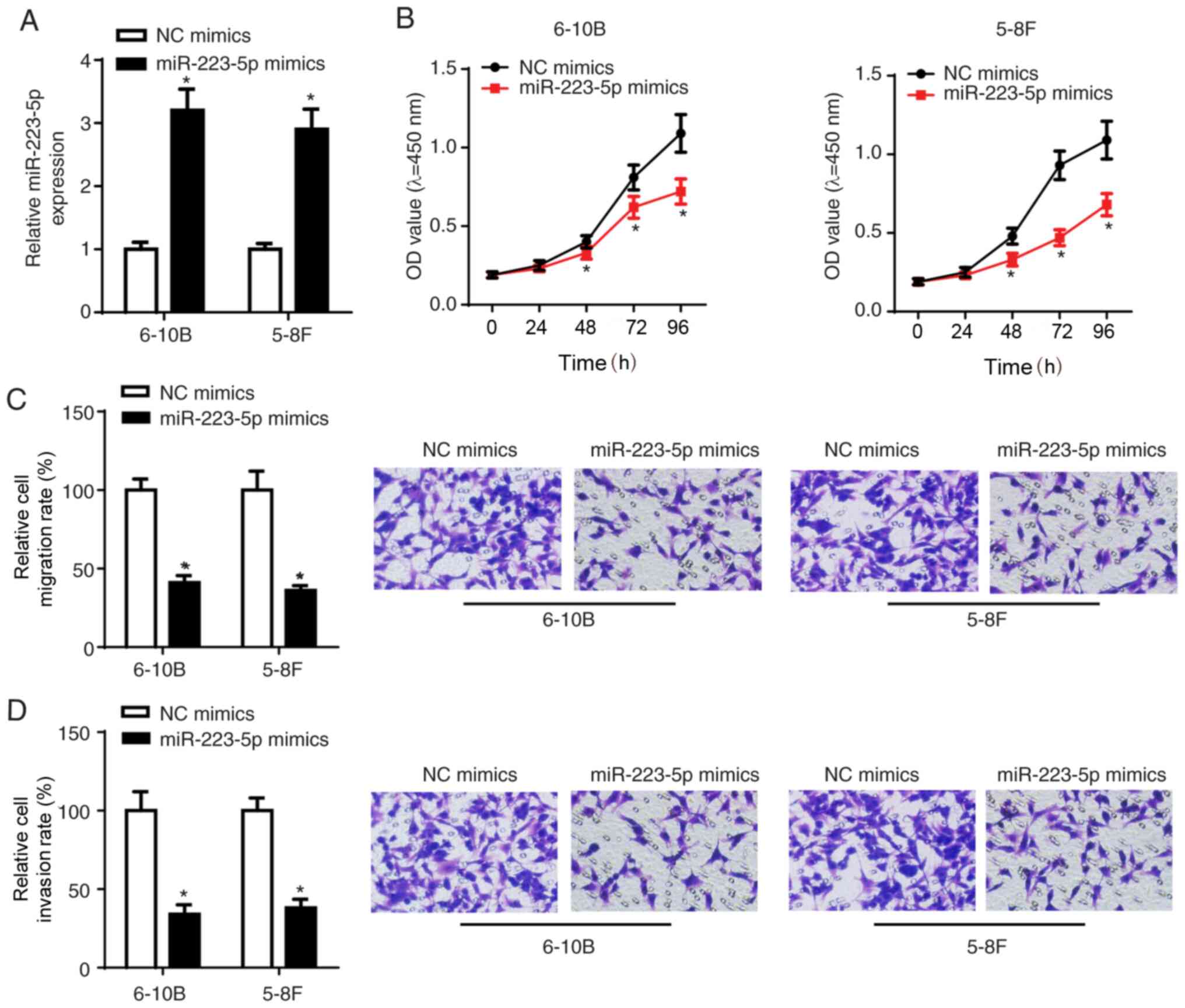
To investigate the biological role of miR-223-5p in
NPC, 5-8F and 6-10B cells were transfected with NC mimics and
miR-223-5p mimics. The RT-qPCR results demonstrated that miR-223-5p
expression was significantly elevated following transfection with
miR-223-5p mimics (Fig. 2A). The
CCK-8 and Transwell assay results indicated that cell viability,
migration and invasion were inhibited following transfection of
5-8F and 6-10B cells with miR-223-5p mimics (Fig. 2B-D), suggesting that miR-223-5p
overexpression NPC cell viability, migration and invasion.
DCLK1 silencing suppresses viability,
migration and invasion of NPC cells
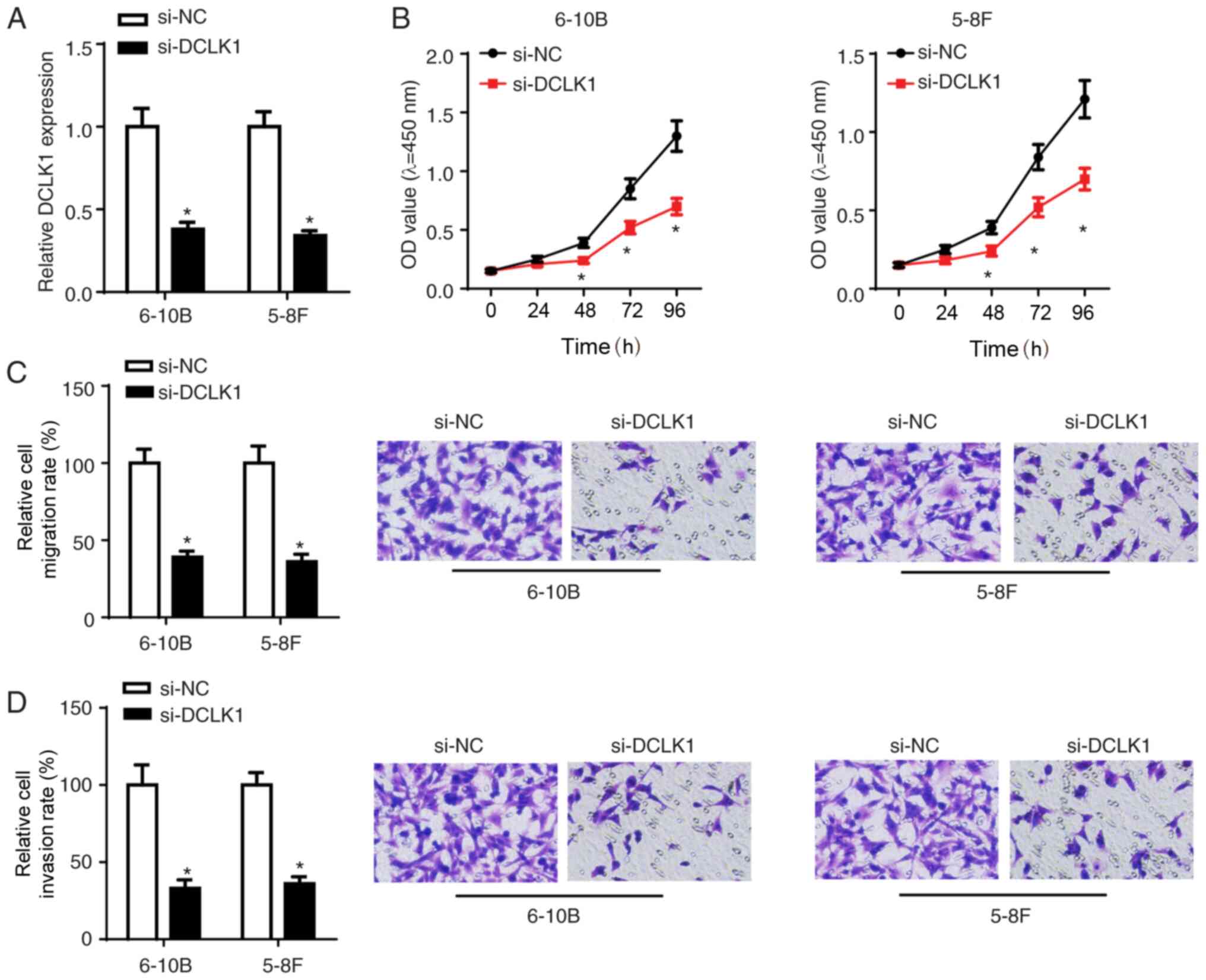
To investigate the function of DCLK1 in NPC
tumorigenesis, si-DCLK1 or si-NC were transfected into 5-8F and
6-10B cells. The transfection efficiency was confirmed via RT-qPCR
(Fig. 3A). The CCK-8 assay results
demonstrated that DCLK1 knockdown inhibited the viability of NPC
cells (Fig. 3B). Moreover, the
Transwell assay results further confirmed that DCLK1 silencing
suppressed the migration and invasion of NPC cells (Fig. 3C and D).
DCLK1 is a target of miR-223-5p
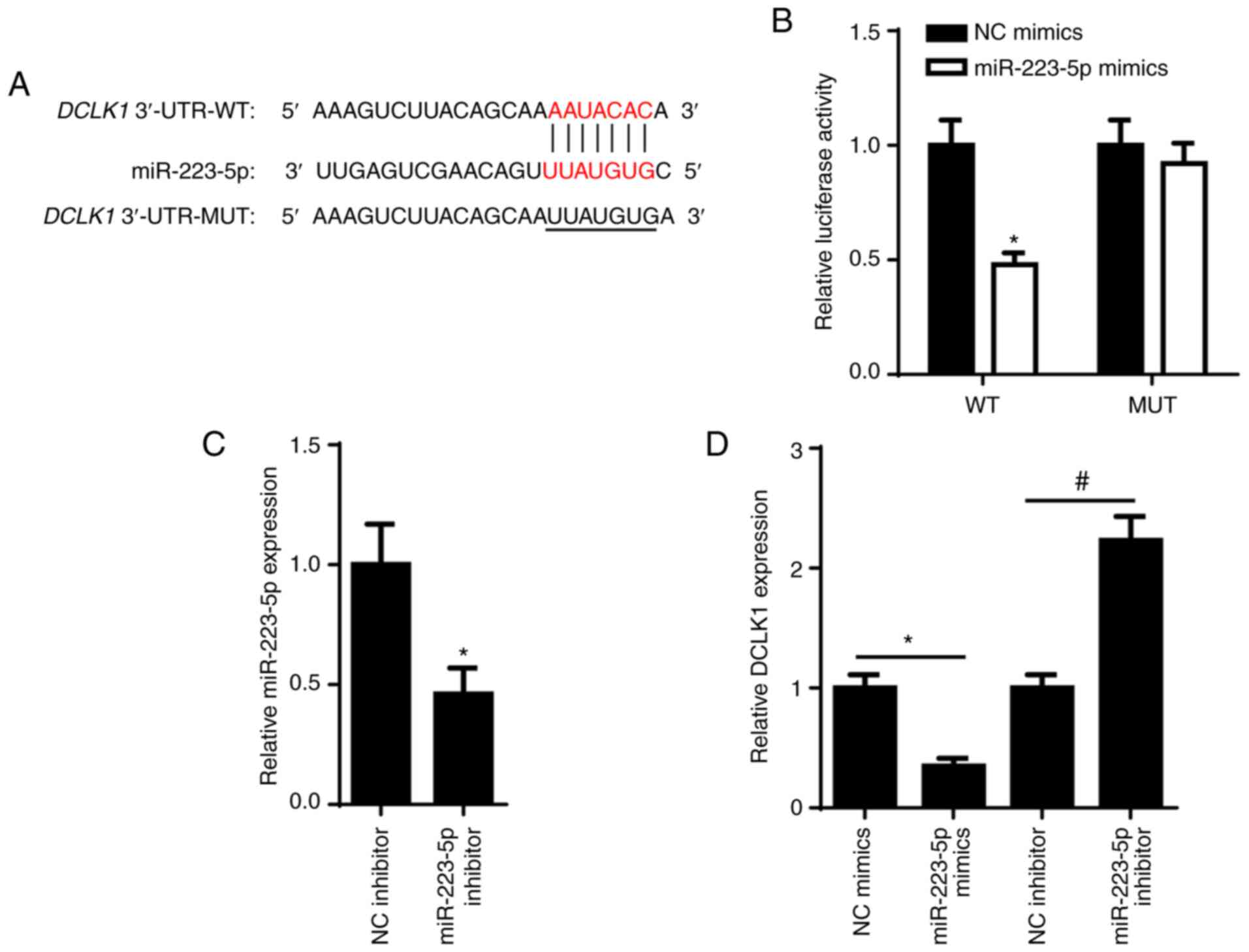
Since DCLK1 was reported to act as an oncogene and
promote tumorigenesis in different types of human cancer (20,21),
TargetScan was used to predict the binding sites between miR-223-5p
and DCLK1. The potential binding sites are shown in Fig. 4A. Luciferase reporter analysis
demonstrated that the luciferase activity was reduced in 6-10B
cells co-transfected with DCLK1-WT and miR-223-5p mimics, while
there was little change in the DCLK1-MUT group (Fig. 4B), suggesting a specific interaction
between miR-223-5p and DCLK1. RT-qPCR revealed that miR-223-5p
expression was significantly downregulated in 6-10B cells
transfected with miR-223-5p inhibitor (Fig. 4C). In addition, the effect of
miR-223-5p on DCLK1 expression was examined (Fig. 4D) and the results indicated that
miR-223-5p overexpression suppressed DCLK1 expression in 6-10B
cells, while miR-223-5p knockdown resulted in increased expression
of DCLK1.
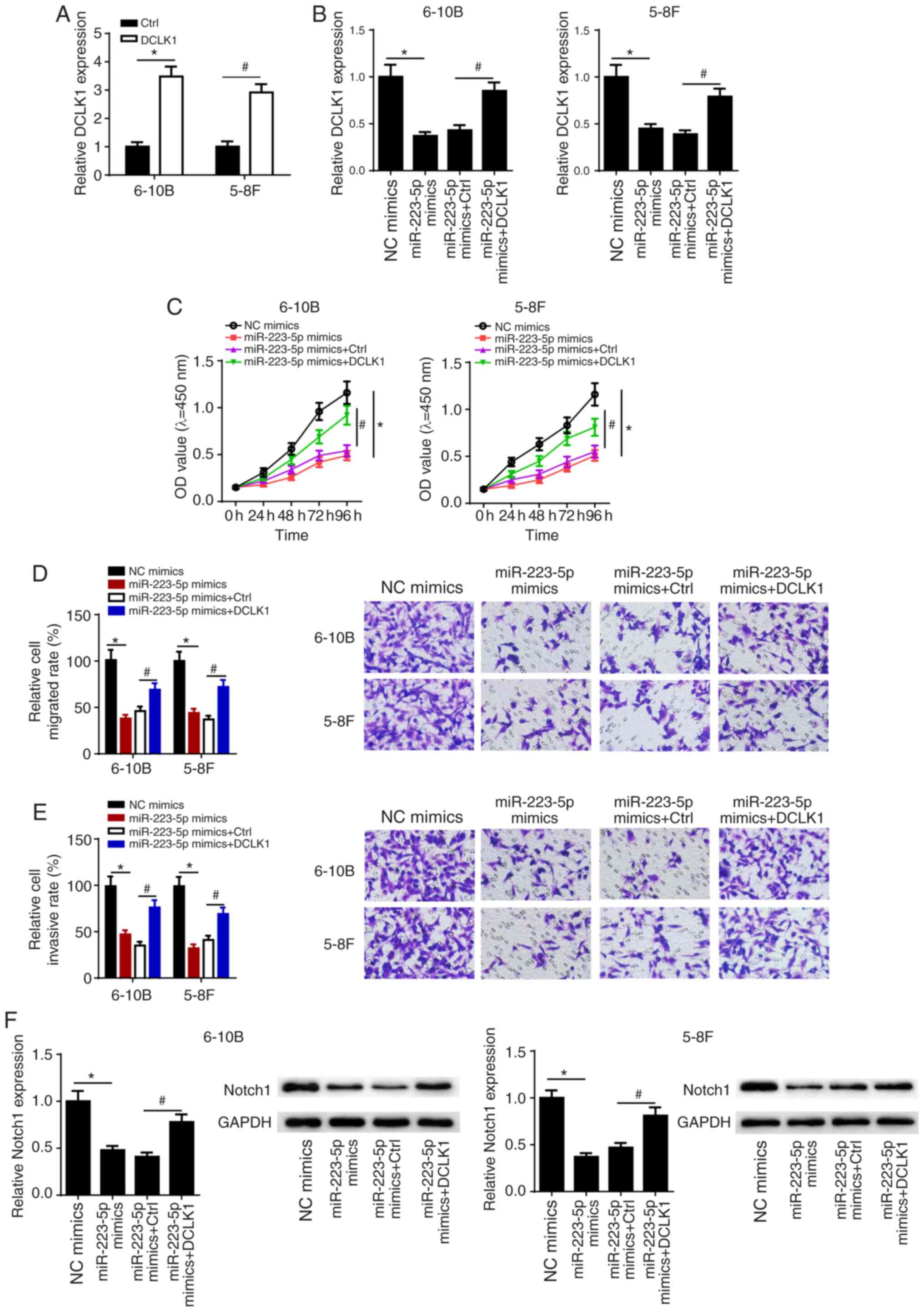
miR-223-5p overexpression suppresses
NPC cell viability, migration and invasion via the DCLK1/Notch1
signaling pathway
To examine whether miR-223-5p overexpression
suppressed cell viability, migration and invasion in NPC via DCLK1,
5-8F and 6-10B cells were transfected with NC mimics, miR-223-5p
mimics, miR-223-5p + Ctrl and miR-223-5p + DCLK1, and then DCLK1
expression, viability, migration and invasion were measured in 5-8F
and 6-10B cells. DCLK1 expression was found to be markedly
increased in NPC cells transfected with DCLK1 overexpression
plasmid (Fig. 5A). In addition, it
was found that miR-223-5p overexpression not only decreased DCLK1
expression in 5-8F and 6-10B cells, but also inhibited the
viability, migration and invasion of 5-8F and 6-10B cells. However,
DCLK1 overexpression could partially reverse the suppressive
effects of miR-223-5p overexpression on the viability, migration
and invasion of 5-8F and 6-10B cells (Fig. 5B-E).
A previous study reported that DCLK1 regulated tumor
metastasis and epithelial-to-mesenchymal transition of gastric
cancer cells via the Notch1 signaling pathway (22). Therefore, it was hypothesized that
miR-223-5p and DCLK1 may regulate NPC progression via Notch1
signaling. The results suggested that overexpression of miR-223-5p
decreased the protein expression of Notch1, which was partially
restored following overexpression of DCLK1 in 5-8F and 6-10B cells
(Fig. 5F). These results indicated
that the effects of miR-223-5p and DCLK1 on NPC progression may be
mediated via the Notch1 signaling pathway.
Discussion
Multiple factors, such as Epstein-Barr virus
infection, environmental factors and genetic susceptibility genes,
have been reported to contribute to the development of NPC
(23). Since the two major causes of
mortality among patients with NPC are recurrence and metastasis, it
is crucial to further elucidate the molecular mechanism underlying
NPC progression. As a result, additional personalized treatments
may be developed and provided to patients with NPC. The present
study demonstrated that miR-223-5p was downregulated in NPC tissues
and cells, and acted as a tumor suppressor by suppressing the
malignant behavior of NPC cells.
Abnormally expressed miRNAs may play key roles in
NPC progression by facilitating cell proliferation, invasion and
angiogenesis. For example, miR-216b was found to suppress NPC
growth by downregulating KRAS expression (24), and miR-26a inhibited NPC cell
proliferation and cell cycle by suppressing enhancer of zeste 2
polycomb repressive complex 2 subunit (25). miR-142-3p silencing may contribute to
NPC progression via modulating zinc finger E-box binding homeobox 2
(26). Moreover, miR-223 was found
to be involved in various types of cancer. For example, miR-223
expression was found to be suppressed in hepatocellular carcinoma
and facilitated Stathmin1 expression (27). miR-223 also suppressed the
progression of prostate cancer via regulating integrin subunit
α3/integrin subunit β1 signaling (28). Additionally, miR-223 suppressed cell
proliferation and migration in NPC via targeting MAF bZIP
transcription factor B (29). The
aforementioned studies indicated the important role of miR-223 in
NPC progression. miR-223-5p is the passenger strand of the miR-223
duplex (10). Consistent with all
these studies, the present study demonstrated that miR-223-5p
expression was downregulated in NPC tissues compared with that in
adjacent normal nasopharyngeal tissues. In the present study,
RT-qPCR analysis revealed low miR-223-5p expression in NPC tissues
and cells. The results of the CKK-8 and Transwell assays suggested
that miR-223-5p overexpression may suppress cell viability,
migration and invasion in NPC. Therefore, the present study
demonstrated the inverse association between miR-223-5p expression
and NPC tumor progression.
Previous studies have reported that aberrant DCLK1
expression may be closely associated with the malignant biological
properties of tumors. For example, miR-613 was found to suppress
the growth and invasion of human hepatocellular carcinoma by
inhibiting DCLK1 (30), whereas
miR-137 inhibited the malignant behavior of colon cancer via
downregulation of DCLK1 (31).
miR-424 also suppressed the viability and invasion of neuroblastoma
cells by directly modulating DCLK1 (32). In addition, high expression of DCLK1
has been demonstrated to promote the progression of human
pancreatic cancer (33). In the
present study, DCLK1 was predicted as a functional target gene of
miR-223-5p in NPC. Subsequently, DCLK1 expression was measured in
NPC tissues and cells, and it was observed that DCLK1 was highly
expressed in NPC. The effect of DCLK1 knockdown on NPC was similar
to that of miR-223-5p overexpression. Moreover, DCLK1
overexpression was able to reverse the inhibitory effect of
miR-223-5p on NPC cell viability, migration and invasion.
Furthermore, DCLK1 has been reported to be involved
in the progression of human cancers via several signaling pathways.
For example, Wang et al (34)
reported that DCLK1 facilitated progression of breast cancer via
the Wnt/β-catenin signaling pathway. Liu et al (22) indicated that DCLK1 promoted the
epithelial-to-mesenchymal transition of gastric cancer cells
through Notch1 signaling. The inhibition of the Notch1 signaling
pathway is considered as an effective target for human cancer
treatment (35). In the present
study, it was observed that overexpression of miR-223-5p decreased
the expression of Notch1, while this effect was partially reversed
by the overexpression of DCLK1. Therefore, these results suggested
that miR-223-5p and DCLK1 may regulate the tumorigenic process of
NPC via the Notch1 signaling pathway.
In conclusion, the present study demonstrated that
miR-223-5p expression was downregulated in NPC tissues and cells,
and miR-223-5p functioned as a tumor suppressor in NPC. miR-223-5p
overexpression may decrease the viability, migration and invasion
of NPC cells, and suppress tumor progression via downregulating
DCLK1. The present findings may improve our understanding of the
mechanism involved in the progression of NPC mediated by
miR-223-5p, and prompt further investigation of novel targeted
therapies based on miRNA-mRNA networks for patients with NPC.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Applied Basic
Research Program of Changzhou (grant no. CJ20200056).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors contributions
ZZ and CC designed the study. HL, JY, HX and XT
performed the experiments, analyzed the data and prepared the
figures. ZZ and CC drafted the initial manuscript and revised the
manuscript. ZZ, HL, JY, HX, XT and CC confirm the authenticity of
all the raw data. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of The Third Affiliated Hospital of Soochow University,
and written informed consent was obtained from each patient
enrolled in the study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Cao SM, Simons MJ and Qian CN: The
prevalence and prevention of nasopharyngeal carcinoma in China.
Chin J Cancer. 30:114–119. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lee VH, Lam KO, Chang AT, Lam TC, Chiang
CL, So TH, Choi CW and Lee AW: Management of nasopharyngeal
carcinoma: Is adjuvant therapy needed? J Oncol Pract. 14:594–602.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang TT, Chen ZZ, Xie P, Zhang WJ, Du MY,
Liu YT, Zhu HY and Guo YS: Isoliquiritigenin suppresses the
proliferation and induced apoptosis via miR-32/LATS2/Wnt in
nasopharyngeal carcinoma. Eur J Pharmacol. 856:1723522019.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yi SJ, Liu P, Chen BL, Ou-Yang L, Xiong WM
and Su JP: Circulating miR-31-5p may be a potential diagnostic
biomarker in nasopharyngeal carcinoma. Neoplasma. 66:825–829. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Genova P, Brunetti F, Bequignon E, Landi
F, Lizzi V, Esposito F, Charpy C, Calderaro J, Azoulay D and
deAngelis N: Solitary splenic metastasis from nasopharyngeal
carcinoma: A case report and systematic review of the literature.
World J Surg Oncol. 14:1842016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Venkatadri R, Muni T, Iyer AK, Yakisich JS
and Azad N: Role of apoptosis-related miRNAs in resveratrol-induced
breast cancer cell death. Cell Death Dis. 7:e21042016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dou L, Han K, Xiao M and Lv F: miR-223-5p
suppresses tumor growth and metastasis in non-small cell lung
cancer by targeting E2F8. Oncol Res. 27:261–268. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wei Y, Peng J, He S, Huang H, Lin L, Zhu
Q, Ye L, Li T, Zhang X, Gao Y and Zheng X: miR-223-5p targeting ERG
inhibits prostate cancer cell proliferation and migration. J
Cancer. 11:4453–4463. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sugawara S, Yamada Y, Arai T, Okato A,
Idichi T, Kato M, Koshizuka K, Ichikawa T and Seki N: Dual strands
of the miR-223 duplex (miR-223-5p and miR-223-3p) inhibit cancer
cell aggressiveness: Targeted genes are involved in bladder cancer
pathogenesis. J Hum Genet. 63:657–668. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ferguson FM, Nabet B, Raghavan S, Liu Y,
Leggett AL, Kuljanin M, Kalekar RL, Yang A, He S, Wang J, et al:
Discovery of a selective inhibitor of doublecortin like kinase 1.
Nat Chem Biol. 16:635–643. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sureban SM, May R, Qu D, Weygant N,
Chandrakesan P, Ali N, Lightfoot SA, Pantazis P, Rao CV, Postier RG
and Houchen CW: DCLK1 regulates pluripotency and angiogenic factors
via microRNA-dependent mechanisms in pancreatic cancer. PLoS One.
8:e739402013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Shan C, Fei F, Li F, Zhuang B, Zheng Y,
Wan Y and Chen J: miR-448 is a novel prognostic factor of lung
squamous cell carcinoma and regulates cells growth and metastasis
by targeting DCLK1. Biomed Pharmacother. 89:1227–1234. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liang H, Yu T, Han Y, Jiang H, Wang C, You
T, Zhao X, Shan H, Yang R, Yang L, et al: LncRNA PTAR promotes EMT
and invasion-metastasis in serous ovarian cancer by competitively
binding miR-101-3p to regulate ZEB1 expression. Mol Cancer.
17:1192018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chandrakesan P, Yao J, Qu D, May R,
Weygant N, Ge Y, Ali N, Sureban SM, Gude M, Vega K, et al: Dclk1, a
tumor stem cell marker, regulates pro-survival signaling and
self-renewal of intestinal tumor cells. Mol Cancer. 16:302017.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang J, Wang S, Zhou J and Qian Q:
miR-424-5p regulates cell proliferation, migration and invasion by
targeting doublecortin-like kinase 1 in basal-like breast cancer.
Biomed Pharmacother. 102:147–152. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mohammadi Y, Tavangar SM, Saidijam M,
Amini R, Etemadi K, Karimi Dermani F and Najafi R: DCLK1 plays an
important role in colorectal cancer tumorgenesis through the
regulation of miR-200c. Biomed Pharmacother. 103:301–307. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang Z, Yuan XG, Chen J, Luo SW, Luo ZJ
and Lu NH: Reduced expression of PTEN and increased PTEN
phosphorylation at residue Ser380 in gastric cancer tissues: A
novel mechanism of PTEN inactivation. Clin Res Hepatol
Gastroenterol. 37:72–79. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu H, Wen T, Zhou Y, Fan X, Du T, Gao T,
Li L, Liu J, Yang L, Yao J, et al: DCLK1 plays a
metastatic-promoting role in human breast cancer cells. Biomed Res
Int. 2019:10619792019.PubMed/NCBI
|
|
21
|
Liu W, Wang S, Sun Q, Yang Z, Liu M and
Tang H: DCLK1 promotes epithelial-mesenchymal transition via the
PI3K/Akt/NF-κB pathway in colorectal cancer. Int J Cancer.
142:2068–2079. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu ZQ, He WF, Wu YJ, Zhao SL, Wang L,
Ouyang YY and Tang SY: LncRNA SNHG1 promotes EMT process in gastric
cancer cells through regulation of the miR-15b/DCLK1/Notch1 axis.
BMC Gastroenterol. 20:1562020. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen YP, Chan ATC, Le QT, Blanchard P, Sun
Y and Ma J: Nasopharyngeal carcinoma. Lancet. 394:64–80. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Deng M, Tang H, Zhou Y, Zhou M, Xiong W,
Zheng Y, Ye Q, Zeng X, Liao Q, Guo X, et al: miR-216b suppresses
tumor growth and invasion by targeting KRAS in nasopharyngeal
carcinoma. J Cell Sci. 124:2997–3005. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lu J, He ML, Wang L, Chen Y, Liu X, Dong
Q, Chen YC, Peng Y, Yao KT, Kung HF and Li XP: MiR-26a inhibits
cell growth and tumorigenesis of nasopharyngeal carcinoma through
repression of EZH2. Cancer Res. 71:225–233. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li Y, He Q, Wen X, Hong X, Yang X, Tang X,
Zhang P, Lei Y, Sun Y, Zhang J, et al: EZH2-DNMT1-mediated
epigenetic silencing of miR-142-3p promotes metastasis through
targeting ZEB2 in nasopharyngeal carcinoma. Cell Death Differ.
26:1089–1106. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wong QW, Lung RW, Law PT, Lai PB, Chan KY,
To KF and Wong N: MicroRNA-223 is commonly repressed in
hepatocellular carcinoma and potentiates expression of Stathmin1.
Gastroenterology. 135:257–269. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kurozumi A, Goto Y, Matsushita R, Fukumoto
I, Kato M, Nishikawa R, Sakamoto S, Enokida H, Nakagawa M, Ichikawa
T and Seki N: Tumor-suppressive microRNA-223 inhibits cancer cell
migration and invasion by targeting ITGA3/ITGB1 signaling in
prostate cancer. Cancer Sci. 107:84–94. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yang W, Lan X, Li D, Li T and Lu S:
MiR-223 targeting MAFB suppresses proliferation and migration of
nasopharyngeal carcinoma cells. BMC Cancer. 15:4612015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang W, Zhang H, Wang L, Zhang S and Tang
M: miR-613 inhibits the growth and invasiveness of human
hepatocellular carcinoma via targeting DCLK1. Biochem Biophys Res
Commun. 473:987–992. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sakaguchi M, Hisamori S, Oshima N, Sato F,
Shimono Y and Sakai Y: miR-137 regulates the tumorigenicity of
colon cancer stem cells through the inhibition of DCLK1. Mol Cancer
Res. 14:354–362. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wan MF, Yang N, Qu NY, Pan YY, Shan YQ and
Li P: MiR-424 suppressed viability and invasion by targeting to the
DCLK1 in neuroblastoma. Eur Rev Med Pharmacol Sci. 24:5526–5533.
2020.PubMed/NCBI
|
|
33
|
Ito H, Tanaka S, Akiyama Y, Shimada S,
Adikrisna R, Matsumura S, Aihara A, Mitsunori Y, Ban D, Ochiai T,
et al: Dominant expression of DCLK1 in human pancreatic cancer stem
cells accelerates tumor invasion and metastasis. PLoS One.
11:e01465642016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang YL, Li Y, Ma YG and Wu WY: DCLK1
promotes malignant progression of breast cancer by regulating
Wnt/β-catenin signaling pathway. Eur Rev Med Pharmacol Sci.
23:9489–9498. 2019.PubMed/NCBI
|
|
35
|
Yuan X, Wu H, Xu H, Xiong H, Chu Q, Yu S,
Wu GS and Wu K: Notch signaling: An emerging therapeutic target for
cancer treatment. Cancer Lett. 369:20–27. 2015. View Article : Google Scholar : PubMed/NCBI
|