Introduction
Breast cancer is the most commonly diagnosed cancer
worldwide and accounted for ~25% of new cancer cases among women in
2018 (1); it is associated with a
mortality rate of 15%, making it the most common cause of
cancer-related mortality among women (1). The prognosis of numerous patients with
breast cancer has improved considerably over the last few years
with advances in hormone therapy, chemotherapy and targeted
therapies (2). However,
triple-negative breast cancer (TNBC), the cells of which do not
express estrogen receptor, progesterone receptor or human epidermal
growth factor receptor 2, does not respond well to hormone therapy,
and no effective targeted therapy is currently available (3). In addition, TNBC is aggressive and
often metastasizes to distant sites (4). Therefore, the prognosis of patients
with TNBC is worse compared with that of patients with other
subtypes of breast cancer, and further research is needed to
identify new therapeutic targets in TNBC.
Aquaporins (AQPs) are membrane proteins that
transport water and small solutes across cell membranes, and play a
key role in fluid homeostasis (5,6). Thus
far, 13 mammalian AQPs have been identified, the first of which was
AQP1, which is highly expressed in the microvascular endothelium
(5,6). AQP1 upregulation has been reported in
colon, gastric and ovarian cancer, among others, and its
upregulation is associated with tumor cell replication, invasion,
migration and metastasis (7–9).
AQP1 upregulation has also been reported in breast
cancer, and its upregulation may be even more prominent in TNBC
(10,11). AQP1 may be an independent predictor
of prognosis in breast cancer, and it has been proposed as a novel
biomarker (12,13). However, the exact function of AQP1 in
breast cancer remains unclear. Therefore, the present study was
undertaken to investigate the effects of AQP1 knockdown on TNBC
cell proliferation, migration and invasion.
Materials and methods
Cell culture and transfection
The human breast cancer MDA-MB-231, MCF-7 and
SK-BR-3 cell lines, and the human mammary epithelial MCF-10A cell
line, were purchased from the Shanghai Institute of Biochemistry
and Cell Biology. MDA-MB-231 and MCF-7 cells were cultured in
complete DMEM supplemented with 10% FBS (both Biological
Industries), 100 U/ml penicillin and 100 µg/ml streptomycin.
SK-BR-3 and MCF-10A cells were cultured in RPMI-1640 (Biological
Industries) containing 1.5 mg/ml NaHCO3, 10% FBS, 100
U/ml penicillin and 100 µg/ml streptomycin. All cell lines were
cultured at 37°C in a 5% CO2 atmosphere.
Short hairpin RNA (shRNA) targeting AQP1 mRNA
(sh-AQP1; 5′-CCATTATGCTGGTGTATGT-3′) (GV248-AQP1) and the
corresponding negative control shRNA with a non-targeting AQP1
sequence (sh-NC; 5′-TTCTCCGAACGTGTCACGT-3′) (GV248-NC) were
designed and synthesized by Shanghai GeneChem Co., Ltd., and the
concentrations were both adjusted to 1×108 TU/ml. The
sh-AQP1 and sh-NC (MOI=20) were transfected into MDA-MB-231 cells
(3.0×103 cells per well) using polybrene (Beyotime
Institute of Biotechnology) according to the manufacturer's
protocol. The duration of transfection was 12 h at 37°C followed by
changing the fresh medium. Transfected cells were used for
subsequent experiments after 72 h.
Reverse transcription-quantitative PCR
(RT-qPCR) analysis
Total RNA was extracted from breast cancer cells
(MCF-7, MDA-MB-231 and SK-BR-3) and normal mammary epithelial cells
(MCF-10A) using TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.), and 1 µg RNA was used to generate cDNA
using the ReverTra Ace qPCR RT kit (Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions. qPCR was performed
using the SYBR Green PCR Master mix (Takara Bio, Inc.). The
following primer pairs were used: AQP1 forward,
5′-CAGCCCAAGGACAGTTCAGAG-3′ and reverse,
5′-CCATCATGGCTAAGTGCACAG-3′; and β-actin forward,
5′-TGGCACCCAGCACAATGAA-3′ and reverse,
5′-CTAAGTCATAGTCCGCCTAGAAGGA-3′. Thermocycling conditions were as
follows: Initial denaturation at 95°C for 30 sec, followed by 40
cycles of denaturation at 95°C for 5 sec, annealing at 60°C for 30
sec and extension at 72°C for 20 sec, for 40 cycles. The levels of
AQP1 mRNA were quantified relative to levels of β-actin mRNA using
the 2−ΔΔCq method (14).
Western blot analysis
Total protein was extracted from breast cancer cells
(MCF-7, MDA-MB-231 and SK-BR-3) and normal mammary epithelial cells
(MCF-10A) using RIPA lysis buffer (Beyotime Institute of
Biotechnology) and measured using Pierce BCA Protein Assay kit
(Beyotime Institute of Biotechnology). A total of 20 µg of protein
was loaded for electrophoretic separation on 10% SDS/polyacrylamide
gels and transferred onto PVDF membranes (MilliporeSigma).
Membranes were blocked with 5% skimmed milk in TBS/Tween-20 (0.1%)
buffer at room temperature for 2 h. The membranes were incubated
overnight at 4°C with anti-AQP1 antibody (cat. no. 25287; 1:100
dilution) and anti-β-actin antibody (cat. no. 8432; 1:200 dilution)
(both Santa Cruz Biotechnology, Inc.) as an internal control.
Following the primary incubation, membranes were incubated with
horseradish peroxidase-conjugated anti-mouse IgG secondary antibody
(cat. no. sc-516102; 1:5,000 dilution; Santa Cruz Biotechnology,
Inc.) at room temperature for 2 h. The immunocomplex was detected
using the ECL Plus kit (Amersham; Cytiva), and the band density was
analyzed with Image Lab 5.0 (Bio-Rad Laboratories, Inc.).
Cell proliferation assay
Cell proliferation was measured using the Cell
Counting Kit-8 (CCK-8) kit (Dojindo Molecular Technologies, Inc.)
according to the manufacturer's instructions. Untransfected
MDA-MB-231 cells (blank control), and cells transfected with sh-NC
or sh-AQP1 were seeded into 96-well culture plates at a density of
4×103 cells/per well and cultured at 24-h intervals for
4 days. Subsequently, 10 µl of CCK-8 solution was added to each
well and the cells were incubated for 1 h at 37°C. A microplate
reader (Thermo Fisher Scientific, Inc.) was used to measure the
absorbance at 450 nm.
Transwell migration and invasion
assays
In the migration assay, a total of 4×104
blank control, sh-NC or sh-AQP1 cells were seeded into the upper
chamber of a Transwell migration insert with 8-µm pores (Corning,
Inc.) containing serum-free DMEM. DMEM supplemented with 10% FBS
(600:l) was added to the lower chamber to serve as a
chemoattractant. Transwell plates were incubated for 24 h at 37°C,
and cells that migrated to the lower chamber of the insert were
fixed with 4% paraformaldehyde, stained with Giemsa (1:10 dilution;
Beijing Solarbio Science & Technology Co., Ltd.) for 15 min at
37°C and counted under a light microscope (BX-42; Olympus
Corporation; magnification, ×100).
The invasion assay was performed in the same manner,
except that 1 mg/ml Matrigel (BD Biosciences) was polymerized in
the upper chamber prior to cell seeding.
In vivo experiments
All animal handling and experimental procedures were
in compliance with and approved by the Ethics Committee of Guangxi
Medical University (Nanning, China). A total of 42 female Balb/c
nude mice (age, 5–6 weeks; weight, 15–16 g; n=14 mice/group) were
purchased from Guangxi Medical University Animal Experiment Center
and maintained under specific pathogen-free conditions at 22–25°C
and a 12-h light/dark cycle, with free access to water and sterile
food. Previously transfected blank control, sh-NC or sh-AQP1 cells
were suspended in PBS, the density was adjusted to 1×107
cells/ml and then 100 µl of the suspension was injected
subcutaneously under the breast fat pad of the nude mice. The tumor
volume was measured every 5 days using a vernier caliper and
calculated according to the following formula: (Length ×
width2)/2. The maximum tumor volume observed in the
present study was 1,102.6 mm3. After 5 weeks, the mice
were anesthetized by intraperitoneal injection with 60 µl 10%
chloral hydrate (300 mg/kg; Beyotime Institute of Biotechnology)
and sacrificed by cervical dislocation. No mice exhibited signs of
peritonitis, pain or discomfort. Death was verified by confirming
respiratory and cardiac arrest.
Statistical analysis
All data are reported as the mean ± SD. Multiple
group comparisons were performed using a one-way ANOVA followed by
Bonferroni's post hoc test. P<0.05 was considered to indicate a
statistically significant difference. All statistical analyses were
performed using SPSS 19.0 (IBM Corp.).
Results
AQP1 expression in breast cancer cell
lines
In the present study, the expression levels of AQP1
in the human breast cancer MDA-MB-231, MCF-7 and SK-BR-3 cell
lines, as well as in the normal human mammary epithelial MCF-10A
cell line, were determined by western blotting and RT-qPCR
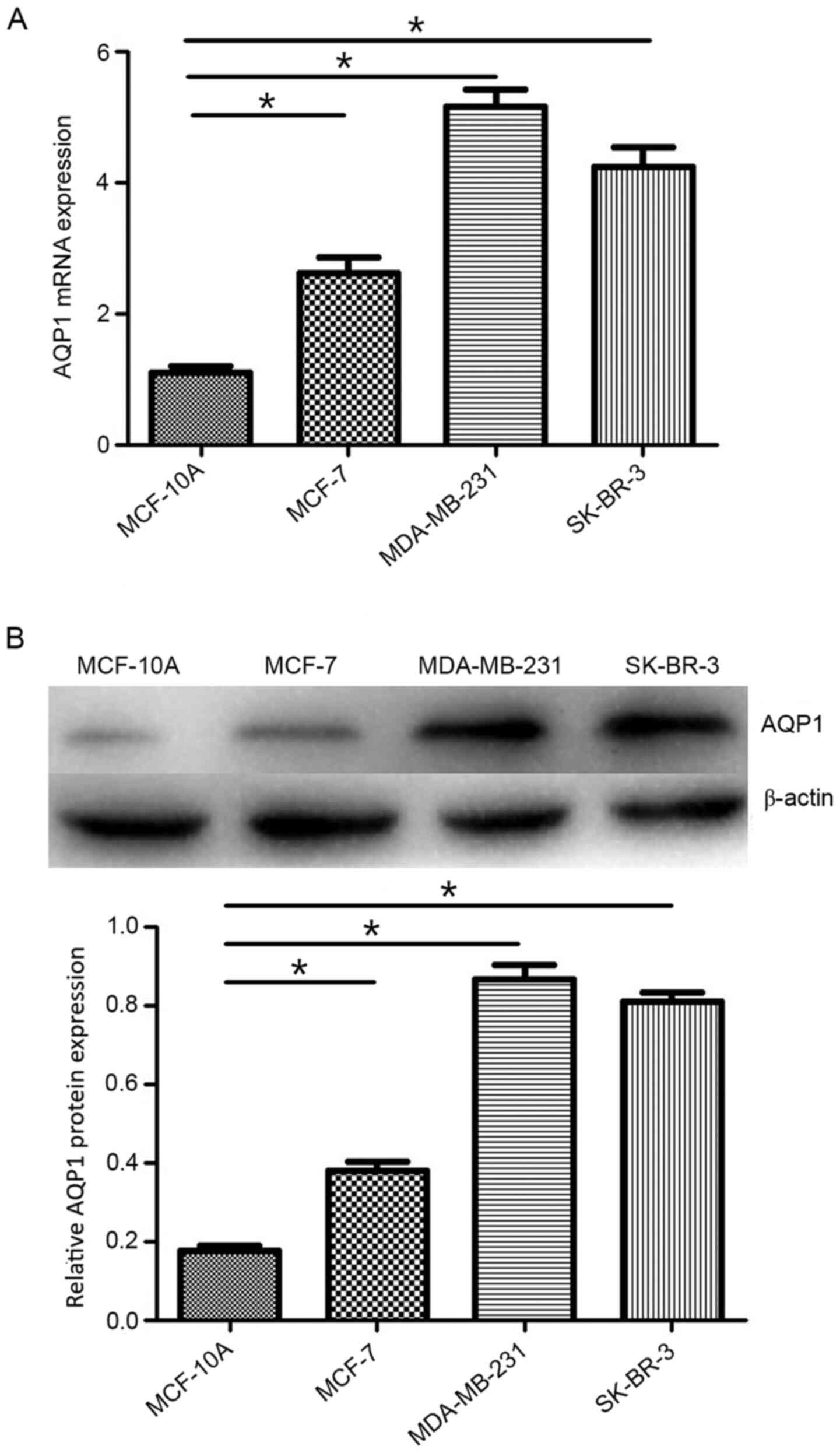
analyses. As shown in Fig. 1A, AQP1
mRNA expression was significantly higher in MDA-MB-231, MCF-7 and
SK-BR-3 cells compared with that in MCF-10A cells. Similarly,
western blotting demonstrated higher protein expression levels of
AQP1 in MDA-MB-231, MCF-7 and SK-BR-3 cells compared with those in
MCF-10A cells (Fig. 1B). The levels
of AQP1 were higher in the TNBC MDA-MB-231 cell line compared with
those in the other breast cancer cell lines. Thus, the MDA-MB-231
cell line was used to investigate the effect of AQP1 on breast
cancer cell proliferation, invasion and migration.
AQP1 knockdown inhibits TNBC cell
proliferation, invasion and migration in vitro
To assess how AQP1 affects cell proliferation in
vitro, shRNA was used to knock down AQP1 expression in
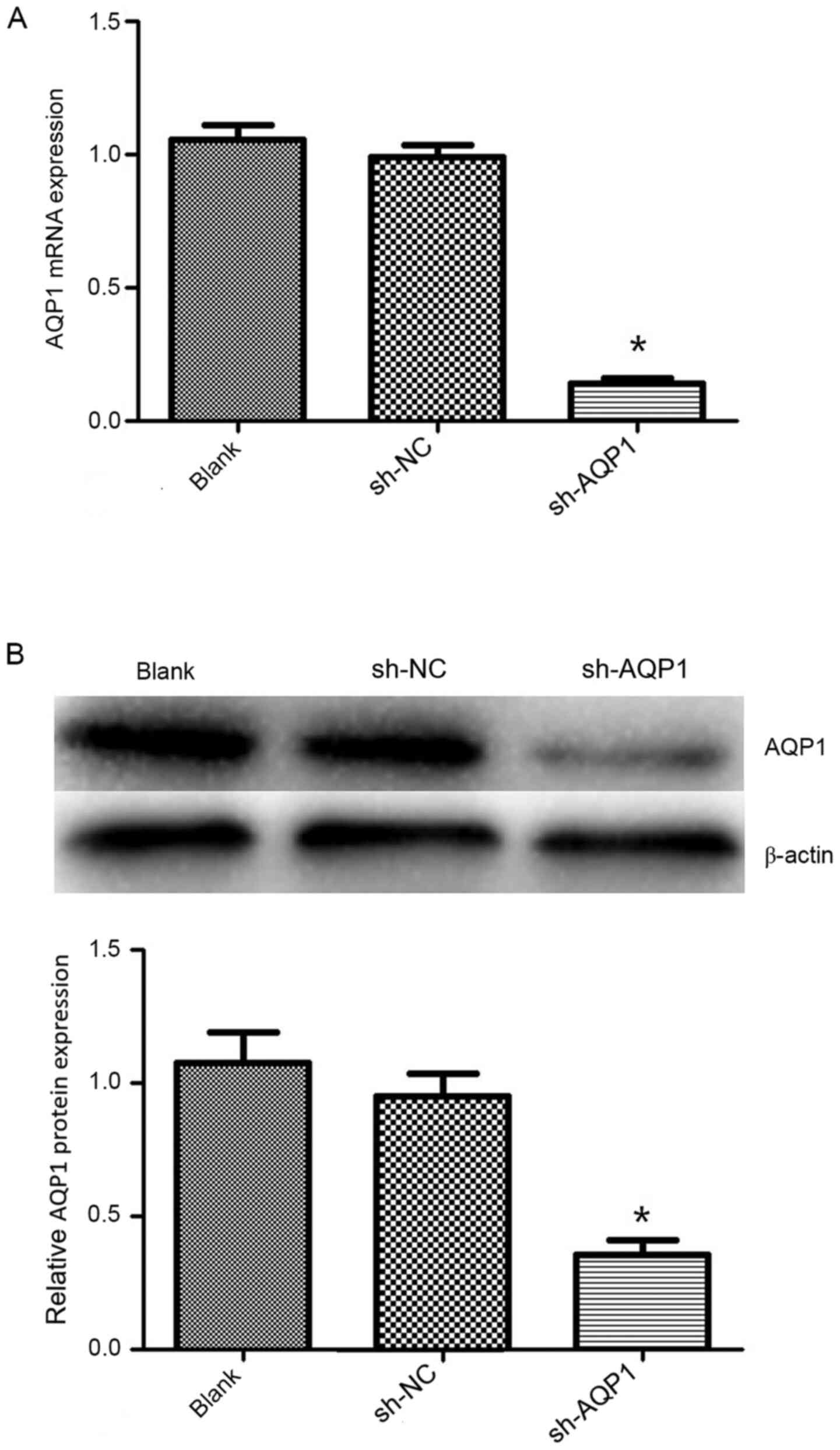
MDA-MB-231 cells. The expression of AQP1 at the mRNA (Fig. 2A) and protein (Fig. 2B) levels was significantly decreased
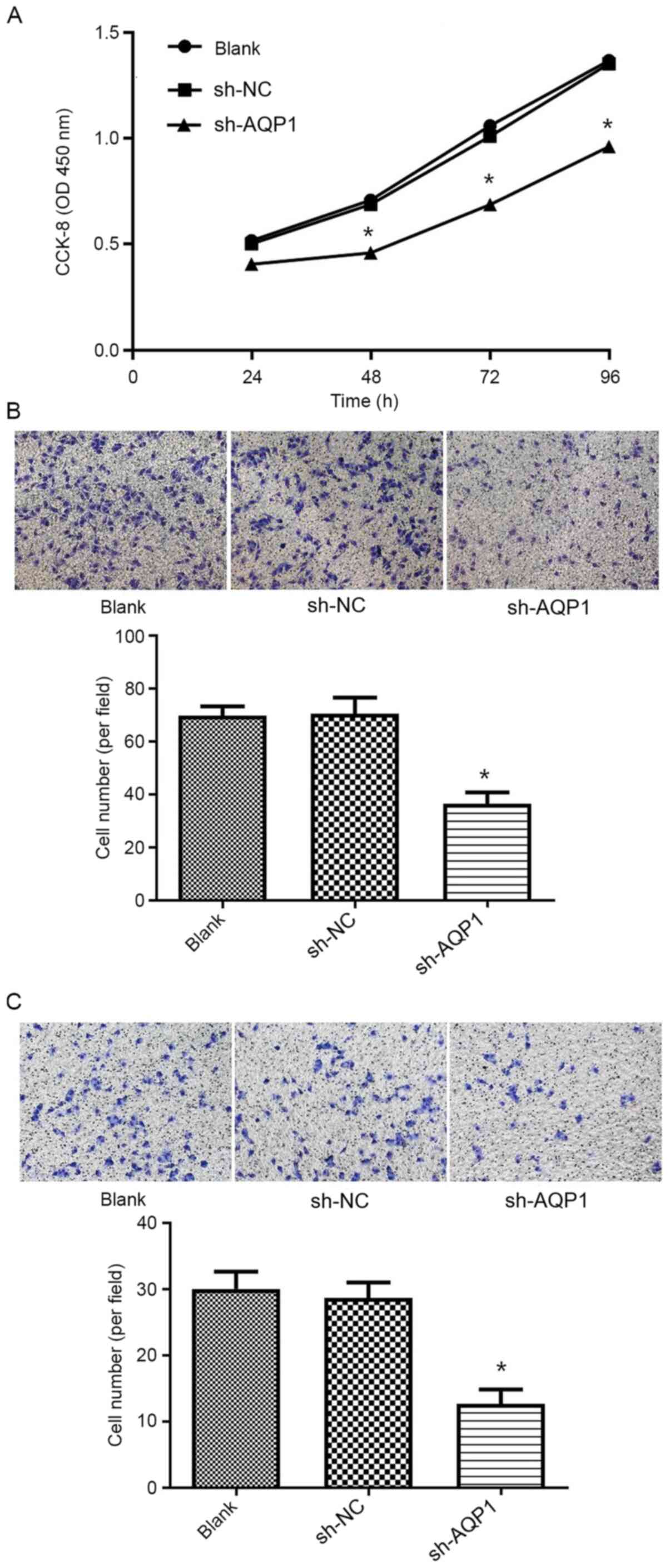
in MDA-MB-231 cells post-transfection. The CCK-8 proliferation
assay was subsequently applied to assess the effect of AQP1
knockdown on the proliferation of breast cancer cells, and it was
found that AQP1 knockdown led to a significantly lower level of
proliferation compared with that in blank control cells and cells
treated with sh-NC (Fig. 3A).
Transwell migration and invasion assays were
performed to investigate the effects of AQP1 knockdown on the
migration and invasion of breast cancer cells. Compared with that
in the blank control and sh-NC groups, a significant decrease in
the number of migrating (Fig. 3B)
and invading (Fig. 3C) MDA-MB-231
cells was apparent following transfection with sh-AQP1.
AQP1 knockdown suppresses TNBC
xenograft growth in vivo
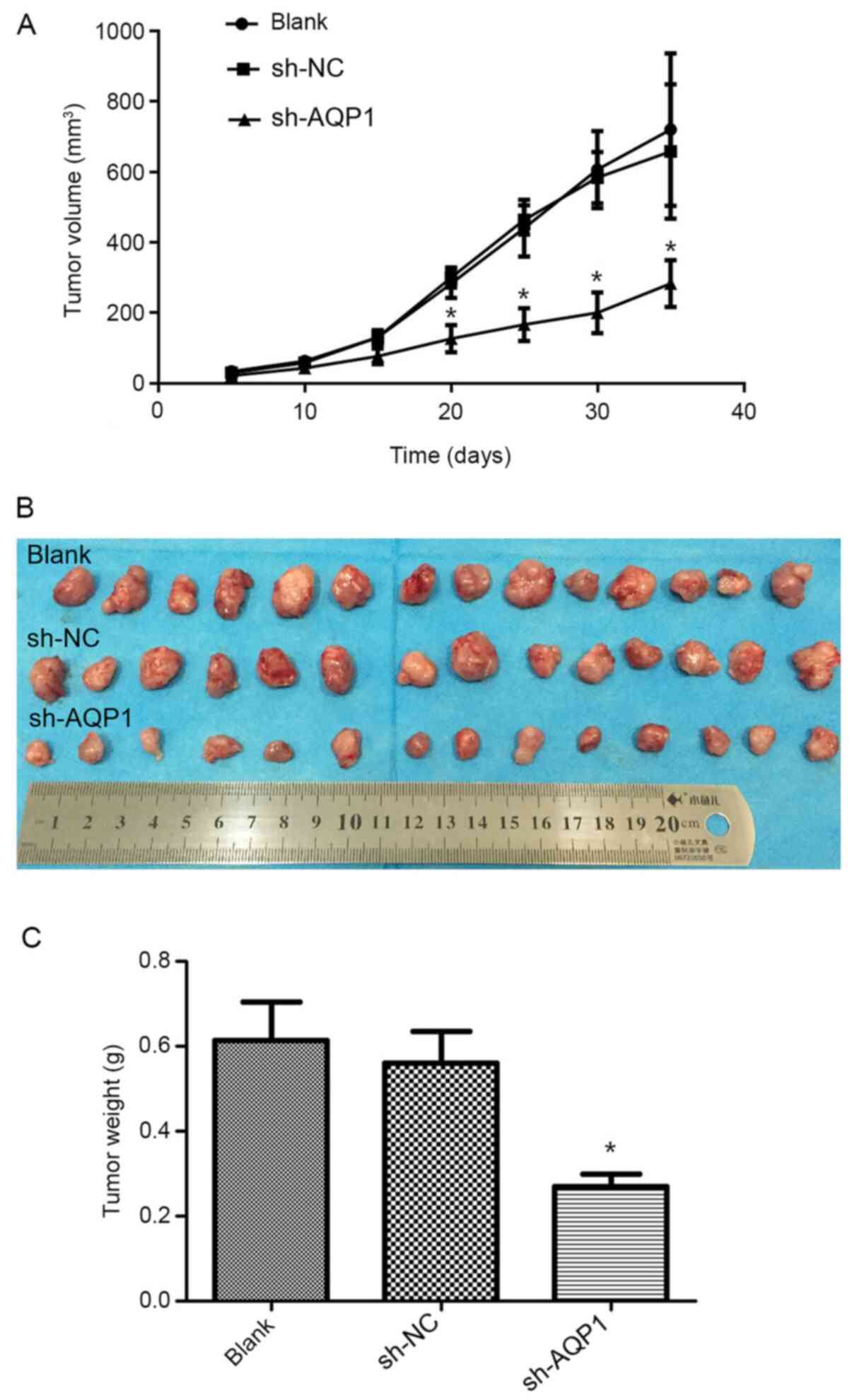
In order to confirm the potential antitumor effects
of AQP1 downregulation in vivo, female BALB/c nude mice were
subcutaneously injected with MDA-MB-231 cells that were stably
transfected with sh-AQP1. When comparing tumor sizes, the knockdown
of AQP1 was found to significantly decrease tumor growth compared
with the blank control and sh-NC groups (Fig. 4A and B). Furthermore, AQP1 knockdown
significantly decreased the weight of the xenografted tumors
(Fig. 4C).
Discussion
The findings of the present study confirmed that
AQP1 was overexpressed in breast cancer cells compared with that in
normal mammary epithelial cells, which was in agreement with
previous findings (15). It was also
confirmed that AQP1 was expressed at higher levels in the TNBC
MDA-MB-231 cell line compared with that in other types of breast
cancer cells, which was consistent with a previous study reporting
a strong association between AQP1 expression and high tumor grade
and hormone receptor negativity (11). A previous study showed that
overexpression of AQP1 promoted the proliferation and invasion of
breast cancer cells (12). The
result suggested that AQP1 could be a potential prognostic
biomarker for breast cancer. However, it is not clear whether AQP1
can be used as a potential therapeutic target in breast cancer,
particularly TNBC. Thus, to explore the antitumor potential of
downregulating AQP1, the present study investigated the effects of
AQP1 knockdown on TNBC growth in vitro and in
vivo.
It was also demonstrated that AQP1 knockdown
inhibited the proliferation of TNBC cells, which was consistent
with the results of previous studies in which AQP1 knockdown
markedly suppressed the viability and promoted the apoptosis of
ovarian cancer cells (16), and
suppressed the proliferation of lung adenocarcinoma, osteosarcoma
and glioma cells (17–19). However, AQP1 downregulation did not
alter the proliferation of B16F10 melanoma cells, although it did
alter their water permeability (20). These results suggest that AQP1 plays
different roles in different types of cancer.
Breast cancer invasiveness decreases patient
survival and doubles the mortality rate (21). The present study demonstrated that
AQP1 knockdown suppressed the migration and invasion of TNBC cells
in vitro, as previously reported for ovarian cancer, lung
adenocarcinoma, osteosarcoma and glioma cells in culture (16–19), but
not for melanoma cells (20). AQP1
overexpression may help drive tumor cell migration by importing
water to fill cellular protrusions, creating more space for actin
polymerization at the leading edge of migration (22–24).
AQP1 may also promote tumor invasion by stimulating angiogenesis
(25–27).
Regardless of the underlying mechanism, AQP1 may
serve as a potential therapeutic target in TNBC by inhibiting the
proliferation, migration and invasion of tumor cells.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81860464) and the
Natural Science Foundation of Guangxi Province (grant no.
2019GXNSFAA185038).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
HY conceived and designed the study. YJ and XL
performed the experiments. YNJ and WW analyzed the data. YJ wrote
the manuscript. YNJ and HY confirm the authenticity of all the raw
data. All the authors have read and approved the final manuscript,
and agree to be accountable for all aspects of the research.
Ethics approval and consent to
participate
All animal handling and experimental procedures were
in compliance with and approved by the Ethics Committee of the
Guangxi Medical University (Nanning, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
DeSantis CE, Ma J, Goding Sauer A, Newman
LA and Jemal A: Breast cancer statistics, 2017, racial disparity in
mortality by state. CA Cancer J Clin. 67:439–448. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Scott LC, Mobley LR, Kuo TM and Il'yasova
D: Update on triple-negative breast cancer disparities for the
United States: A population-based study from the United States
cancer statistics database, 2010 through 2014. Cancer.
125:3412–3417. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hwang KT, Kim J, Jung J, Chang JH, Chai
YJ, Oh SW, Oh S, Kim YA, Park SB and Hwang KR: Impact of breast
cancer subtypes on prognosis of women with operable invasive breast
cancer: A population-based study using SEER database. Clin Cancer
Res. 25:1970–1979. 2019.PubMed/NCBI
|
|
5
|
Verkman AS and Mitra AK: Structure and
function of aquaporin water channels. Am J Physiol Renal Physiol.
278:F13–F28. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Agre P, King LS, Yasui M, Guggino WB,
Ottersen OP, Fujiyoshi Y, Engel A and Nielsen S: Aquaporin water
channels-from atomic structure to clinical medicine. J Physiol.
542:3–16. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kang BW, Kim JG, Lee SJ, Chae YS and Jeong
JY, Yoon GS, Park SY, Kim HJ, Park JS, Choi GS and Jeong JY:
Expression of aquaporin-1, aquaporin-3, and aquaporin-5 correlates
with nodal metastasis in colon cancer. Oncology. 88:369–376. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Thapa S, Chetry M, Huang K, Peng Y, Wang
J, Wang J, Zhou Y, Shen Y, Xue Y and Ji K: Significance of
aquaporins' expression in the prognosis of gastric cancer. Biosci
Rep. 38:BSR201716872018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chetry M, Li S, Liu H, Hu X and Zhu X:
Prognostic values of aquaporins mRNA expression in human ovarian
cancer. Biosci Rep. 38:BSR201801082018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Otterbach F, Callies R, Kimmig R, Schmid
KW and Bánkfalvi A: Aquaporin 1 expression in invasive breast
carcinomas. Pathologe. 29 (Suppl 2):S357–S362. 2008.(In German).
View Article : Google Scholar
|
|
11
|
Otterbach F, Callies R, Adamzik M, Kimmig
R, Siffert W, Schmid KW and Bankfalvi A: Aquaporin 1 (AQP1)
expression is a novel characteristic feature of a particularly
aggressive subgroup of basal-like breast carcinomas. Breast Cancer
Res Treat. 120:67–76. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Qin F, Zhang H, Shao Y, Liu X, Yang L,
Huang Y, Fu L, Gu F and Ma Y: Expression of aquaporin1, a water
channel protein, in cytoplasm is negatively correlated with
prognosis of breast cancer patients. Oncotarget. 7:8143–8154. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhu L, Ma N, Wang B, Wang L, Zhou C, Yan
Y, He J and Ren Y: Significant prognostic values of aquaporin mRNA
expression in breast cancer. Cancer Manag Res. 11:1503–1515. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shi Z, Zhang T, Luo L, Zhao H, Cheng J,
Xiang J and Zhao C: Aquaporins in human breast cancer:
Identification and involvement in carcinogenesis of breast cancer.
J Surg Oncol. 106:267–272. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang Y, Fan Y, Zheng C and Zhang X:
Knockdown of AQP1 inhibits growth and invasion of human ovarian
cancer cells. Mol Med Rep. 16:5499–5504. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wei X and Dong J: Aquaporin 1 promotes the
proliferation and migration of lung cancer cell in vitro.
Oncol Rep. 34:1440–1448. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Guan Y, Chen J, Zhan Y and Lu H: Effects
of dexamethasone on C6 cell proliferation, migration and invasion
through the upregulation of AQP1. Oncol Lett. 15:7595–7602.
2018.PubMed/NCBI
|
|
19
|
Wu Z, Li S, Liu J, Shi Y, Wang J, Chen D,
Luo L, Qian Y, Huang X and Wang H: RNAi-mediated silencing of AQP1
expression inhibited the proliferation, invasion and tumorigenesis
of osteosarcoma cells. Cancer Biol Ther. 16:1332–1340. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hu J and Verkman AS: Increased migration
and metastatic potential of tumor cells expressing aquaporin water
channels. FASEB J. 20:1892–1894. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hussain SA, Ganesan R, Reynolds G, Gross
L, Stevens A, Pastorek J, Murray PG, Perunovic B, Anwar MS,
Billingham L, et al: Hypoxia-regulated carbonic anhydrase IX
expression is associated with poor survival in patients with
invasive breast cancer. Br J Cancer. 96:104–109. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Verkman AS: Aquaporin water channels and
endothelial cell function. J Anat. 200:617–627. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Verkman AS: Knock-out models reveal new
aquaporin functions. Handb Exp Pharmacol. 359–381. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Verkman AS, Hara-Chikuma M and
Papadopoulos MC: Aquaporins-new players in cancer biology. J Mol
Med (Berl). 86:523–529. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Esteva-Font C, Jin BJ and Verkman AS:
Aquaporin-1 gene deletion reduces breast tumor growth and lung
metastasis in tumor-producing MMTV-PyVT mice. FASEB J.
28:1446–1453. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Nicchia GP, Stigliano C, Sparaneo A, Rossi
A, Frigeri A and Svelto M: Inhibition of aquaporin-1 dependent
angiogenesis impairs tumour growth in a mouse model of melanoma. J
Mol Med (Berl). 91:613–623. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
El Hindy N, Bankfalvi A, Herring A,
Adamzik M, Lambertz N, Zhu Y, Siffert W, Sure U and Sandalcioglu
IE: Correlation of aquaporin-1 water channel protein expression
with tumor angiogenesis in human astrocytoma. Anticancer Res.
33:609–613. 2013.PubMed/NCBI
|