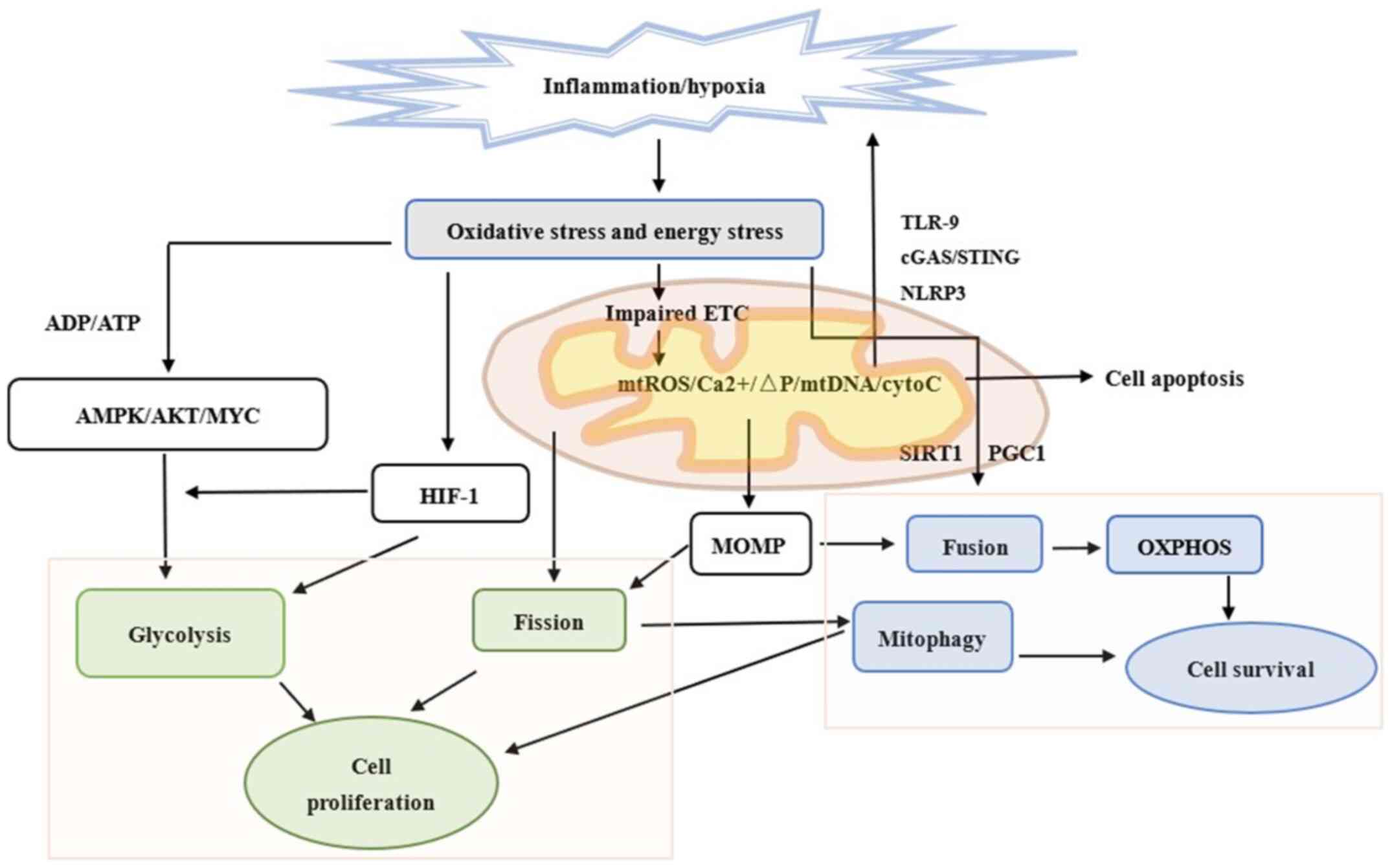
Chronic atrophic gastritis (CAG) is considered a
common risk factor for the development of gastric cancer (GC).
Endoscopic imaging and biopsy are crucial for early detection and
diagnosis of GC (1). Image-enhanced
endoscopy combined with biopsy, according to the Sydney protocol
and regular endoscopic surveillance, are recommended for patients
with extensive CAG or intestinal metaplasia (2). A visible lesion may be treated by
endoscopic mucosal resection or endoscopic submucosal dissection.
However, when a lesion is invisible, regular endoscopic
surveillance is required for high-risk patients. The interval
between Helicobacter pylori eradication and cancer occurrence may
vary from several months to >10 years (3). Surveillance endoscopy is one of the
methods enabling the early diagnosis of GC (4). Once an existing lesion is identified,
it can be treated in a timely manner. Interval cancer can occur due
to missed lesions or to a newly developed lesion during
surveillance (5). Thus, it is
essential to identify a molecular biological marker for the
detection of invisible lesions at the organelle level.
Recently, diverse pathophysiological functions of
mitochondria have been reported, including mitochondrial dynamics
(6), metabolic reprogramming
(7), mitochondria-released
damage-associated molecular patterns and NLRP3 inflammasome
activation (8), mitochondrial DNA
(mtDNA), autophagy and mitophagy (9), mitochondrial outer membrane
permeabilization (10) and
mitochondrial aging (11). In
addition, mitochondrial (mt)DNA mutations, deletions and impaired
DNA replication are the most common causes of mitochondrial
dysfunction (12). mtDNA sensing via
STING signaling participates in inflammation and cancer (12,13). The
effects of mitochondrial dynamics on carcinogenesis and cancer
progression have also been reported, highlighting the potential use
of mitochondrial biomarkers in cancer detection and prognosis, as
well as the potential targeting of mitochondrial dynamics for
treating cancer (14). However,
there is still a paucity of research associated with GC.
The present review summarizes the role of
mitochondrial dynamics and energy metabolism reprogramming in GC to
identify potential indicators for biologically complemented
endoscopy and further promote translating discoveries of molecular
biology. Thus, fission and glycolysis from mitochondria may be
useful in detecting GC. If an electron microscope can be installed
on the endoscopy system, the mitochondrial dynamics may be
observable during the early stages of GC. Furthermore, when fission
is increased and fusion is decreased, further precision biopsy of
the targeted tissue should be performed to detect metabolic
activity. The combination of both approaches may enable early
diagnosis and provide a novel treatment strategy. However, further
investigation is required.
Mitochondria are responsible for energy supply and
are involved in several biological processes, including cell death
and proliferation (6). Mitochondria
constantly maintain a dynamic shape, which may change in response
to cellular bioenergetic demands, such as nutrient status, which is
defined as mitochondrial dynamics (12). The mitochondrial morphology is a
result of the interplay between rapid fusion and fission events
(15). The key components mediating
these processes belong to the dynamin family of GTPases that
utilize GTP hydrolysis to drive mechanical work on biological
membranes (16). Mitofusin proteins,
Mfn1 and Mfn2, are involved in the fusion of the outer
mitochondrial membrane, while GTPase optic atrophy 1 mediates the
fusion of the inner mitochondrial membrane (17). Mitochondrial fission is mediated by
the GTPase dynamin-related protein 1 (Drp1) following its
recruitment by the membrane-anchored proteins, namely mitochondrial
fission factor and fission protein 1 (Fis1) (18). Commonly, the mitochondrial
fission/fusion machinery is involved in generating new
mitochondria, and eliminating old, damaged and non-repairable
mitochondria (6). Mitochondrial
fission plays an important role in mitochondrial proliferation,
mitochondrial distribution during cell division and the removal of
damaged mitochondria via mitophagy (19). Unopposed mitochondrial fission causes
mitochondrial fragmentation, which is generally associated with
metabolic dysfunction and several diseases, such as degenerative
diseases and cancer (20). It has
been reported that impaired mitochondrial fission is associated
with mitochondrial elongation (21).
In addition, unopposed fusion results in a hyperfused network and
serves to counteract metabolic insults, preserve cellular integrity
and protect against autophagy (20).
It was previously reported that impaired mitochondrial fusion may
promote fission-induced mitochondrial fragmentation (21). Thus, the maintenance of mitochondrial
fission/fusion balance plays a key role in cell cycle progression
(6). The dynamics is critical for
the effects of fission/fusion on morphology regulation, content
exchange, and the maintenance of mtDNA and mitochondrial oxidative
phosphorylation (OXPHOS) activity (22,23).
Fission isolates depolarized mitochondria, while it
coordinates the downregulation of fusion mediators to prevent
network reintegration, thereby facilitating mitophagy, mainly via
interactions between Parkin, Bcl-2/adenovirus E1B 19 kDa protein
interacting protein 3 (BNIP3) and Drp1 (24). Increasing Drp1 results in excessive
mitochondrial fragmentation and deficiencies, decreases
mitochondrial motility and shortens mitochondrial length (25), which may be further enhanced in
hypoxia (26). Fission can also be
triggered by stress stimuli, such as nutrient deprivation, DNA
damage, inflammation and mitochondrial membrane depolarization
(27). Given that
mitochondria-associated membranes related to the endoplasmic
reticulum at specific regions can facilitate calcium
(Ca2+) flux into the mitochondria and further control
the homeostasis and metabolism of Ca2+, close coupling
of these organelles increases mitochondrial Ca2+ levels,
thus initiating apoptosis (28). It
has also been reported that enhanced fission attenuates adherence
to inhibit Ca2+ overload in mitochondria and apoptosis
(29). In terms of mitophagy, this
process maintains cellular health by selectively enclosing damaged
and depolarized mitochondria in autophagic vacuoles for
lysosome-mediated elimination (30).
Mitophagy degrades dysfunctional mitochondria and further
attenuates reactive oxygen species (ROS) generation, which in turn
promotes cell survival and protects against cell death (31). Increasing evidence suggest that
several modulators of mitophagy are deregulated in human cancer,
including Parkinson protein 2 E3 ubiquitin protein ligase, FUN14
domain containing 1, BNIP3 and BNIP3L (32,33). In
addition, a study revealed that impaired mitophagy can enhance the
aggressiveness in GC cells under hypoxia by activating the
mtROS/hypoxia-inducible factor (HIF)-1α interplay (34). Mitophagy may also be enhanced by
overexpression of Opa-interacting protein 5, thus plays an
important role in cell survival and death in docetaxel-treated GC
cells (35). Another study
demonstrated that Drp1 expression is upregulated, and the
expression levels of the mitophagy-related regulators, PTEN-induced
putative kinase 1 and Parkin, are downregulated in patients with GC
(36). Given that mitophagy can
clear the damaged part of mitochondria and mtDNA, it protects
healthy cells from malignant transformation and tumor cells from
apoptosis (31). It has been
suggested that, in the early stages of GC, mitophagy is associated
with tumor suppression, whereby it can promote tumor growth at the
advanced stages of GC. For example, mitophagy was increased in
advanced-stage GC to sustain the viability and migration of GC
cells (37), since mitophagy in
solids tumor may be activated by two common factors, namely hypoxia
and low nutrient supply (38)
(Fig. 1).
Mitochondrial fusion results in a more
interconnected mitochondrial network and enhances the communication
with the endoplasmic reticulum (39). Fusion allows the diffusion of matrix
content among mitochondria, diluting the accumulated mtDNA
mutations and oxidized proteins (40). Fusion is commonly enhanced by
starvation by triggering the protein kinase A-mediated
phosphorylation of Drp1 (at Ser637) to blunt fission (41). In addition, mitochondrial fusion is
required for mtDNA maintenance (22). Thus, impaired mitochondrial fusion is
often accompanied by bioenergetic defects due to loss of mtDNA
(42). Furthermore, mitochondrial
fusion is also associated with increased OXPHOS and ATP generation
via remodeling of the cristae (43,44), and
downregulation of OPA1, which is responsible for fusion, resulted
in mitochondrial dysfunction and mtDNA stress (45). The number of mitochondria is
regulated by mitochondrial biogenesis to meet the energy demands of
the cells and compensate for their damage (46). A study demonstrated that peroxisome
proliferator-activated receptor gamma coactivator (PGC-1) and the
protein deacetylase sirtuin 1 (SIRT1) can regulate fusion and
OXPHOS (14). Thus, activation of
PGC-1α by SIRT1 induces mitochondrial biogenesis and confers
metabolic advantages (14). Another
study revealed that PGC-1β can induce mitochondrial fusion by
upregulating Mfn2 expression via estrogen-related receptor α
coactivation (47). Mfn2 expression
is downregulated in GC tissues compared with normal gastric mucosal
tissues, and is negatively associated with tumor size, indicating
an antitumor role of Mfn2 (48). In
vitro experiments have demonstrated that overexpression of Mfn2 can
suppress gastric cancer cell proliferation and colony formation
(48). SIRT1 is an enzyme that
mediates NAD+-dependent deacetylation of target
substrates (49). Given that the
cellular redox balance of NAD+ and NADH is highly
associated with catabolic fluxes, SIRT1 can act as a sensor,
directly connecting metabolic perturbations with transcriptional
output (49). SIRT1 expression is
significantly downregulated in GC tissues, which is associated with
poor prognosis (50). It has also
been reported that SIRT1 exerts inhibitory effects on
chemoresistance and cancer stem cell properties via Forkhead box O3
and AMP-activated protein kinase (AMPK) (51). AMPK, another key energy metabolic
sensor, plays a key role in maintaining cellular energy homeostasis
and is activated upon alterations in the cellular AMP/ATP ratio
(52). Previous studies have
demonstrated that, upon energy deficiency, AMPK activation may
result in increased PGC-1α expression and phosphorylation to
modulate the expression of several key players in mitochondrial
biogenesis and OXPHOS of fatty acids (53,54)
(Fig. 1).
Energy metabolism is essential for maintaining
cellular homeostasis and biological functions, and includes ATP
production in the cytosol (glycolysis) and mitochondria (OXPHOS)
(55), which can be reprogrammed
during carcinogenesis (56). Cancer
cells undergo metabolic reprogramming, including enhanced
glycolysis, mutations in genes encoding tricarboxylic acid (TCA)
cycle enzymes, upregulation of de novo lipid synthesis and
glutaminolysis (57). Glycolysis is
characterized by an increased rate of glucose uptake and its
glycolytic conversion to lactate, even under oxygen-rich conditions
(55). There are several pathways
and transcriptional regulators involved in the regulation of
metabolic reprogramming, such as PI3K/AKT pathway and HIF-1
(58,59). The PI3K/AKT pathway can regulate
several aspects of this metabolic program (58). A previous study demonstrated that AKT
activation was sufficient to induce glycolysis by promoting glucose
transporter 1 and phosphorylating pyruvate dehydrogenase kinase to
inhibit pyruvate dehydrogenase and favor lactate dehydrogenase
(LDH) activity (60). It has been
reported that HIF-1 is overexpressed in human cancers as a result
of intratumoral hypoxia, as well as genetic alterations, such as
gain-of-function mutations in oncogenes and loss-of-function
mutations in tumor suppressor genes (61). HIF-1 may also be triggered by the
accumulation of TCA substrates (62), while its degradation is regulated by
O2-dependent prolyl hydroxylation (PHs) (61). HIF-1α maintains its stability by
avoiding the hydroxylation of PHs in cancer cells, since PHs can be
inhibited by the increased levels of cytosolic pyruvate, lactate,
succinate, fumarate and ROS (59).
Most genes encoding glycolytic enzymes and transporters are the
targets of HIF-1α, and its overexpression in cancer cells is
associated with increased levels of glycolytic proteins (63). A study revealed that HIF-1α levels
were high in certain tumors, even under oxygen-rich conditions,
indicating that hormones or growth factors can cause the
stabilization of HIF-1α expression, which may serve important roles
in carcinogenesis (64). A previous
study suggested that HIF-1α can act as a negative regulator of
mitochondrial biogenesis and oxidative phosphorylation to inhibit
the conversion of pyruvate to acetyl-CoA and mitochondrial
respiration and to promote LDH expression (65). HIF-1α activation can also inhibit MYC
transcription to further downregulate PGC-1α and PGC-1β expression,
which in turn regulates mitochondrial biogenesis and OXPHOS
(54). In GC, inhibiting HIF-1α
signaling attenuates the migratory and invasive abilities of GC
cells, and epithelial-to-mesenchymal transition (66), whereas activation of HIF-1α signaling
promotes cell metastasis and glucose metabolism (67).
The tumor microenvironment favors the growth and
expansion of cancer and inflammatory cells, which in turn directly
or indirectly promotes gastric tumorigenesis by secreting soluble
factors or modulating immune responses (68). It has been reported that NF-κB is
activated in chronic inflammation, thus promoting the further
activation of tumor-promoting genes, such as IL-6 and
cyclooxygenase (COX)-2 (69). NF-κB
and HIF-1 can link inflammatory signaling to hypoxia and coordinate
the activation of both COX-2 and IL-6, and the Janus kinase/STAT3
pathway (70). It has been reported
that STAT3 cooperates with NF-κB and HIF-1 in the regulation of
both genes (71). NF-κB can be
strongly induced by hypoxia and chronic inflammation, and is
involved in the reprogramming of tumor glycolysis by interacting
with HIF-1α (70). Given that
inflammation can induce cells lacking oxygen and upregulate HIF-1α,
glycolysis gradually becomes the main energy source instead of
OXPHOS (55) (Fig. 1)
Mitochondrial morphological changes are a type of
primary signal to shape metabolic reprogramming during cellular
quiescence or activation (14,72).
Recent studies have demonstrated that increased mitochondrial
fission promotes a pro-tumorigenic phenotype (12,73,74).
Several studies have been performed in different cell types that
alter their mitochondrial morphology to meet their energy demands,
functions and behaviors. Conversely, certain cells, such as T cells
and stem cells, have higher energy demands to perform their
metabolic and cell-specific functions (75,76).
When T cells recognize major histocompatibility complexes presented
by antigen-presenting cells in response to infection or tumors,
they proliferate and differentiate into different T-cell subsets
(23). Effector T cells display
looser cristae remodeling via fission with reduced electron
transport chain (ETC) complexes, thus attenuating ETC efficiency
and promoting aerobic glycolysis (23). Conversely, in memory T cells, tight
cristae remodeling via fusion with enhanced ETC complex activity is
observed, thus enhancing ETC efficiency and OXPHOS (23). Endothelial progenitor cells (EPCs)
accelerate glycolysis to produce lactate during angiogenesis by
upregulating the expression levels of HIF1α and vascular
endothelial growth factor (77). In
human EPCs, downregulation of Fis1 expression is associated with
mitochondrial dysfunction and may contribute to the impaired
activity of EPCs during the senescence process (73). However, upregulation of Fis1
expression in senescent EPCs restores the younger phenotype
(73). Another study investigated
the function of mitochondrial fission genes in embryonic stem cells
(ESCs). Transmission electron microscopy revealed a significant
increase in the cytoplasm-to-nucleus ratio and mitochondrial
elongation in dynamin-1-like protein (−/−) ESCs caused by
incomplete fission. In addition, increased OXPHOS and intracellular
ATP concentration and reduced glycolysis was observed, which were
associated with mitochondrial elongation (78). The proliferation and invasion of
tumor cells also require faster and increased energy supply
(79). Thus, Drp1 expression is
upregulated in several types of cancer cells, including liver
(80), breast (81) and lung cancers (82), and may be considered as a biomarker
for predicting poor survival in patients with these types of
cancer. A study on ovarian cancer demonstrated that glycolysis is
promoted by activating PI3K/AKT/HIF-1α signaling, while
mitochondrial fission is enhanced by phosphorylation of Drp1 at
Ser616 (83). As a member of the
AMPK family, salt-inducible kinase 2 was demonstrated to be
involved in both pathways (83). In
addition, Drp1 expression was significantly upregulated in
pancreatic cancer (PC) cells and tissues via downregulation of
microRNA-29a expression (74). High
Drp1 expression was associated with poor survival of patients with
PC, while Drp1 promoted both the proliferation and metastasis of PC
cells, mainly through facilitating aerobic glycolysis (74). Another study revealed that Drp1 may
promote KRAS-driven tumor growth by supporting both glycolysis and
mitochondrial function (84). Taken
together, these findings suggest a mutual association between Drp1
and glycolysis, and the promoting effect of Drp1 and glycolysis on
cancer cell proliferation and invasion.
GC is the fifth most common type of cancer and the
third most common cause of cancer-associated mortality, with
784,000 mortalities reported in 2018 worldwide (85). Early detection and treatment can
improve the outcome of patients with GC. Innovative endoscopic
techniques may be more accurate in achieving cytological or even
biological diagnosis. Mitochondria are strongly associated with
carcinogenesis. The present review summarized the role of
mitochondria dynamics, reprogramming of energy metabolism and their
changes in GC. Based on current literature, it can be concluded
that mitochondria in GC are characterized by fission and enhanced
glycolysis to meet the increased energy requirements of cancer
cells, and decrease necrosis via mitophagy. Upregulated expression
levels of Drp1 and HIF-1α are associated with fission and
glycolysis, respectively. The balance of mitochondrial fission and
fusion and the ratio of glycolysis to OXPHOS are positively
associated with different stages of carcinogenesis. When increased
fission and glycolysis and decreased apoptosis and fusion are
detected in high-risk patients, they may indicate that cells are in
the process of malignant transformation. Thus, treatment is
required to inhibit this process, which may be a promising approach
to the detection of early gastric cancer via organelle- and
molecular-level endoscopy in the future.
Not applicable.
The present study was supported by the 1·3·5 project
for disciplines of excellence Clinical Research Incubation Project,
West China Hospital, Sichuan University, China (grant no.
20HXFH016).
Not applicable.
HY designed the present review and drafted the
initial manuscript. BH contributed to designing and reviewing the
manuscript. HY, YL and BH contributed to revising the manuscript
for important intellectual content. Data authentication is not
applicable. All authors have read and approved the final
manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Pimentel-Nunes P, Libânio D, Marcos-Pinto
R, Areia M, Leja M, Esposito G, Garrido M, Kikuste I, Megraud F,
Matysiak-Budnik T, et al: Management of epithelial precancerous
conditions and lesions in the stomach (MAPS II): European Society
of Gastrointestinal Endoscopy (ESGE), European Helicobacter and
Microbiota Study Group (EHMSG), European Society of Pathology
(ESP), and Sociedade Portuguesa de Endoscopia Digestiva (SPED)
guideline update 2019. Endoscopy. 51:365–388. 2019.PubMed/NCBI
|
|
2
|
Banks M, Graham D, Jansen M, Gotoda T,
Coda S, di Pietro M, Uedo N, Bhandari P, Pritchard DM, Kuipers EJ,
et al: British Society of Gastroenterology guidelines on the
diagnosis and management of patients at risk of gastric
adenocarcinoma. Gut. 68:1545–1575. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Take S, Mizuno M, Ishiki K, Kusumoto C,
Imada T, Hamada F, Yoshida T, Yokota K, Mitsuhashi T and Okada H:
Risk of gastric cancer in the second decade of follow-up after
Helicobacter pylori eradication. J Gastroenterol. 55:281–288. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shichijo S and Hirata Y: Characteristics
and predictors of gastric cancer after Helicobacter pylori
eradication. World J Gastroenterol. 24:2163–2172. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Choi SI, Park B, Joo J, Kim YI, Lee JY,
Kim CG, Choi IJ, Kook MC and Cho SJ: Three-year interval for
endoscopic screening may reduce the mortality in patients with
gastric cancer. Surg Endosc. 33:861–869. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Horbay R and Bilyy R: Mitochondrial
dynamics during cell cycling. Apoptosis. 21:1327–1335. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kim SY: Cancer Energy Metabolism: Shutting
Power off Cancer Factory. Biomol Ther (Seoul). 26:39–44. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhou R, Yazdi AS, Menu P and Tschopp J: A
role for mitochondria in NLRP3 inflammasome activation. Nature.
469:221–225. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jung S, Jeong H and Yu SW: Autophagy as a
decisive process for cell death. Exp Mol Med. 52:921–930. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bock FJ and Tait SWG: Mitochondria as
multifaceted regulators of cell death. Nat Rev Mol Cell Biol.
21:85–100. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sun N, Youle RJ and Finkel T: The
Mitochondrial Basis of Aging. Mol Cell. 61:654–666. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Srinivasan S, Guha M, Kashina A and
Avadhani NG: Mitochondrial dysfunction and mitochondrial
dynamics-The cancer connection. Biochim Biophys Acta Bioenerg.
1858:602–614. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu S, Feng M and Guan W: Mitochondrial
DNA sensing by STING signaling participates in inflammation, cancer
and beyond. Int J Cancer. 139:736–741. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Maycotte P, Marín-Hernández A,
Goyri-Aguirre M, Anaya-Ruiz M, Reyes-Leyva J and Cortés-Hernández
P: Mitochondrial dynamics and cancer. Tumour Biol. May 4–2017.(Epub
ahead of print). doi: 10.1177/1010428317698391. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu X, Weaver D, Shirihai O and Hajnóczky
G: Mitochondrial ‘kiss-and-run’: Interplay between mitochondrial
motility and fusion-fission dynamics. EMBO J. 28:3074–3089. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chan DC: Mitochondrial dynamics and its
involvement in disease. Annu Rev Pathol. 15:235–259. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Schrepfer E and Scorrano L: Mitofusins,
from mitochondria to metabolism. Mol Cell. 61:683–694. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cantó C: Mitochondrial dynamics: Shaping
metabolic adaptation. Int Rev Cell Mol Biol. 340:129–167. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ni HM, Williams JA and Ding WX:
Mitochondrial dynamics and mitochondrial quality control. Redox
Biol. 4:6–13. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wai T and Langer T: Mitochondrial dynamics
and metabolic regulation. Trends Endocrinol Metab. 27:105–117.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bhatia D, Capili A and Choi ME:
Mitochondrial dysfunction in kidney injury, inflammation, and
disease: Potential therapeutic approaches. Kidney Res Clin Pract.
39:244–258. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yan C, Duanmu X, Zeng L, Liu B and Song Z:
Mitochondrial DNA: Distribution, mutations, and elimination. Cells.
8:3792019. View Article : Google Scholar
|
|
23
|
Buck MD, O'Sullivan D, Klein Geltink RI,
Curtis JD, Chang CH, Sanin DE, Qiu J, Kretz O, Braas D, van der
Windt GJ, et al: Mitochondrial dynamics controls T cell fate
through metabolic programming. Cell. 166:63–76. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Friedman JR and Nunnari J: Mitochondrial
form and function. Nature. 505:335–343. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Campello S, Lacalle RA, Bettella M, Mañes
S, Scorrano L and Viola A: Orchestration of lymphocyte chemotaxis
by mitochondrial dynamics. J Exp Med. 203:2879–2886. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wu W, Li W, Chen H, Jiang L, Zhu R and
Feng D: FUNDC1 is a novel mitochondrial-associated-membrane (MAM)
protein required for hypoxia-induced mitochondrial fission and
mitophagy. Autophagy. 12:1675–1676. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Twig G, Elorza A, Molina AJ, Mohamed H,
Wikstrom JD, Walzer G, Stiles L, Haigh SE, Katz S, Las G, et al:
Fission and selective fusion govern mitochondrial segregation and
elimination by autophagy. EMBO J. 27:433–446. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kumar V and Maity S: ER Stress-sensor
proteins and ER-mitochondrial crosstalk-signaling beyond (ER)
stress response. Biomolecules. 11:1732021. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Szabadkai G, Simoni AM, Chami M,
Wieckowski MR, Youle RJ and Rizzuto R: Drp-1-dependent division of
the mitochondrial network blocks intraorganellar Ca2+
waves and protects against Ca2+-mediated apoptosis. Mol
Cell. 16:59–68. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang J and Ney PA: Reticulocyte
mitophagy: Monitoring mitochondrial clearance in a mammalian model.
Autophagy. 6:405–408. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Palikaras K, Lionaki E and Tavernarakis N:
Mechanisms of mitophagy in cellular homeostasis, physiology and
pathology. Nat Cell Biol. 20:1013–1022. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xu Y, Shen J and Ran Z: Emerging views of
mitophagy in immunity and autoimmune diseases. Autophagy. 16:3–17.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Xu HM and Hu F: The role of autophagy and
mitophagy in cancers. Arch Physiol Biochem. Oct 9–2019.(Epub ahead
of print). doi: 10.1080/13813455.2019.1675714. View Article : Google Scholar
|
|
34
|
Shida M, Kitajima Y, Nakamura J,
Yanagihara K, Baba K, Wakiyama K and Noshiro H: Impaired mitophagy
activates mtROS/HIF-1α interplay and increases cancer
aggressiveness in gastric cancer cells under hypoxia. Int J Oncol.
48:1379–1390. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kim TW, Lee SJ, Park YJ, Park SY, Oh BM,
Park YS, Kim BY, Lee YH, Cho HJ, Yoon SR, et al: Opa-interacting
protein 5 modulates docetaxel-induced cell death via regulation of
mitophagy in gastric cancer. Tumour Biol. Oct 15–2017.(Epub ahead
of print). View Article : Google Scholar
|
|
36
|
Marzetti E, Lorenzi M, Landi F, Picca A,
Rosa F, Tanganelli F, Galli M, Doglietto GB, Pacelli F, Cesari M,
et al: Altered mitochondrial quality control signaling in muscle of
old gastric cancer patients with cachexia. Exp Gerontol. 87:92–99.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yan H, Qiu C, Sun W, Gu M, Xiao F, Zou J
and Zhang L: Yap regulates gastric cancer survival and migration
via SIRT1/Mfn2/mitophagy. Oncol Rep. 39:1671–1681. 2018.PubMed/NCBI
|
|
38
|
Ferro F, Servais S, Besson P, Roger S,
Dumas JF and Brisson L: Autophagy and mitophagy in cancer metabolic
remodelling. Semin Cell Dev Biol. 98:129–138. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
de Brito OM and Scorrano L: An intimate
liaison: Spatial organization of the endoplasmic
reticulum-mitochondria relationship. EMBO J. 29:2715–2723. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Santel A, Frank S, Gaume B, Herrler M,
Youle RJ and Fuller MT: Mitofusin-1 protein is a generally
expressed mediator of mitochondrial fusion in mammalian cells. J
Cell Sci. 116:2763–2774. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yu R, Liu T, Ning C, Tan F, Jin SB,
Lendahl U, Zhao J and Nistér M: The phosphorylation status of
Ser-637 in dynamin-related protein 1 (Drp1) does not determine Drp1
recruitment to mitochondria. J Biol Chem. 294:17262–17277. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Amati-Bonneau P, Valentino ML, Reynier P,
Gallardo ME, Bornstein B, Boissière A, Campos Y, Rivera H, de la
Aleja JG, Carroccia R, et al: OPA1 mutations induce mitochondrial
DNA instability and optic atrophy ‘plus’ phenotypes. Brain.
131:338–351. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Elezaby A, Sverdlov AL, Tu VH, Soni K,
Luptak I, Qin F, Liesa M, Shirihai OS, Rimer J, Schaffer JE, et al:
Mitochondrial remodeling in mice with cardiomyocyte-specific lipid
overload. J Mol Cell Cardiol. 79:275–283. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yao CH, Wang R, Wang Y, Kung CP, Weber JD
and Patti GJ: Mitochondrial fusion supports increased oxidative
phosphorylation during cell proliferation. Elife. 8:e413512019.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Rodríguez-Nuevo A, Díaz-Ramos A, Noguera
E, Díaz-Sáez F, Duran X, Muñoz JP, Romero M, Plana N, Sebastián D,
Tezze C, et al: Mitochondrial DNA and TLR9 drive muscle
inflammation upon Opa1 deficiency. EMBO J. 37:e965532018.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Yang H and Zhang Z: Sepsis-induced
myocardial dysfunction: the role of mitochondrial dysfunction.
Inflamm Res. Mar 8–2021.(Epub ahead of print). doi:
10.1007/s00011-021-01447-0. View Article : Google Scholar
|
|
47
|
Liesa M, Borda-d'Agua B, Medina-Gómez G,
Lelliott CJ, Paz JC, Rojo M, Palacín M, Vidal-Puig A and Zorzano A:
Mitochondrial fusion is increased by the nuclear coactivator
PGC-1beta. PLoS One. 3:e36132008. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zhang GE, Jin HL, Lin XK, Chen C, Liu XS,
Zhang Q and Yu JR: Anti-tumor effects of Mfn2 in gastric cancer.
Int J Mol Sci. 14:13005–13021. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Tang BL: Sirt1 and the mitochondria. Mol
Cells. 39:87–95. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Li H, He C, Wang X, Wang H, Nan G and Fang
L: MicroRNA-183 affects the development of gastric cancer by
regulating autophagy via MALAT1-miR-183-SIRT1 axis and
PI3K/AKT/mTOR signals. Artif Cells Nanomed Biotechnol.
47:3163–3171. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
An Y, Wang B, Wang X, Dong G, Jia J and
Yang Q: SIRT1 inhibits chemoresistance and cancer stemness of
gastric cancer by initiating an AMPK/FOXO3 positive feedback loop.
Cell Death Dis. 11:1152020. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Gowans GJ and Hardie DG: AMPK: A cellular
energy sensor primarily regulated by AMP. Biochem Soc Trans.
42:71–75. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Shi HJ, Xu C, Liu MY, Wang BK, Liu WB,
Chen DH, Zhang L, Xu CY and Li XF: Resveratrol improves the energy
sensing and glycolipid metabolism of blunt snout bream megalobrama
amblycephala fed high-carbohydrate diets by activating the
AMPK-SIRT1-PGC-1α network. Front Physiol. 9:12582018. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Scarpulla RC: Metabolic control of
mitochondrial biogenesis through the PGC-1 family regulatory
network. Biochim Biophys Acta. 1813:1269–1278. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Yang H, Du L and Zhang Z: Potential
biomarkers in septic shock besides lactate. Exp Biol Med (Maywood).
245:1066–1072. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Bose S and Le A: Glucose metabolism in
cancer. Adv Exp Med Biol. 1063:3–12. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Pavlova NN and Thompson CB: The emerging
hallmarks of cancer metabolism. Cell Metab. 23:27–47. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Alzahrani AS: PI3K/Akt/mTOR inhibitors in
cancer: At the bench and bedside. Semin Cancer Biol. 59:125–132.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Rodríguez-Enríquez S, Marín-Hernández Á,
Gallardo-Pérez JC, et al: Transcriptional regulation of energy
metabolism in cancer cells. Cells. 8:12252019. View Article : Google Scholar
|
|
60
|
Gonnella R, Santarelli R, Farina A,
Granato M, D'Orazi G, Faggioni A and Cirone M: Kaposi sarcoma
associated herpesvirus (KSHV) induces AKT hyperphosphorylation,
bortezomib-resistance and GLUT-1 plasma membrane exposure in THP-1
monocytic cell line. J Exp Clin Cancer Res. 32:792013. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Semenza GL: Targeting HIF-1 for cancer
therapy. Nat Rev Cancer. 3:721–732. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Gottlieb E and Tomlinson IP: Mitochondrial
tumour suppressors: A genetic and biochemical update. Nat Rev
Cancer. 5:857–866. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Koch A, Ebert EV, Seitz T, Dietrich P,
Berneburg M, Bosserhoff A and Hellerbrand C: Characterization of
glycolysis-related gene expression in malignant melanoma. Pathol
Res Pract. 216:1527522020. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Hägg M and Wennström S: Activation of
hypoxia-induced transcription in normoxia. Exp Cell Res.
306:180–191. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Gogvadze V, Orrenius S and Zhivotovsky B:
Mitochondria in cancer cells: What is so special about them? Trends
Cell Biol. 18:165–173. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Zhou Y, Xu Q, Shang J, Lu L and Chen G:
Crocin inhibits the migration, invasion, and epithelial-mesenchymal
transition of gastric cancer cells via miR-320/KLF5/HIF-1α
signaling. J Cell Physiol. 234:17876–17885. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Gan L, Meng J, Xu M, Liu M, Qi Y, Tan C,
Wang Y, Zhang P, Weng W, Sheng W, et al: Extracellular matrix
protein 1 promotes cell metastasis and glucose metabolism by
inducing integrin β4/FAK/SOX2/HIF-1α signaling pathway in gastric
cancer. Oncogene. 37:744–755. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Oya Y, Hayakawa Y and Koike K: Tumor
microenvironment in gastric cancers. Cancer Sci. 111:2696–2707.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Bruning U, Fitzpatrick SF, Frank T,
Birtwistle M, Taylor CT and Cheong A: NFκB and HIF display
synergistic behaviour during hypoxic inflammation. Cell Mol Life
Sci. 69:1319–1329. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
D'Ignazio L, Bandarra D and Rocha S: NF-κB
and HIF crosstalk in immune responses. FEBS J. 283:413–424. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Lavecchia A, Di Giovanni C and Cerchia C:
Novel inhibitors of signal transducer and activator of
transcription 3 signaling pathway: An update on the recent patent
literature. Expert Opin Ther Pat. 24:383–400. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Mishra P and Chan DC: Metabolic regulation
of mitochondrial dynamics. J Cell Biol. 212:379–387. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Wang HH, Wu YJ, Tseng YM, Su CH, Hsieh CL
and Yeh HI: Mitochondrial fission protein 1 up-regulation
ameliorates senescence-related endothelial dysfunction of human
endothelial progenitor cells. Angiogenesis. 22:569–582. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Liang J, Yang Y, Bai L, Li F and Li E:
DRP1 upregulation promotes pancreatic cancer growth and metastasis
through increased aerobic glycolysis. J Gastroenterol Hepatol.
35:885–895. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Almeida L, Lochner M, Berod L and
Sparwasser T: Metabolic pathways in T cell activation and lineage
differentiation. Semin Immunol. 28:514–524. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Van Wyngene L, Vandewalle J and Libert C:
Reprogramming of basic metabolic pathways in microbial sepsis:
therapeutic targets at last? EMBO Mol Med. 10:e87122018. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Ren R, Guo J, Shi J, Tian Y, Li M and Kang
H: PKM2 regulates angiogenesis of VR-EPCs through modulating
glycolysis, mitochondrial fission, and fusion. J Cell Physiol.
235:6204–6217. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Seo BJ, Choi J, La H, Habib O, Choi Y,
Hong K and Do JT: Role of mitochondrial fission-related genes in
mitochondrial morphology and energy metabolism in mouse embryonic
stem cells. Redox Biol. 36:1015992020. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Hsu PP and Sabatini DM: Cancer cell
metabolism: Warburg and beyond. Cell. 134:703–707. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Lin XH, Qiu BQ, Ma M, Zhang R, Hsu SJ, Liu
HH, Chen J, Gao DM, Cui JF, Ren ZG, et al: Suppressing
DRP1-mediated mitochondrial fission and mitophagy increases
mitochondrial apoptosis of hepatocellular carcinoma cells in the
setting of hypoxia. Oncogenesis. 9:672020. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Liu B, Fan Y, Song Z, Han B, Meng Y, Cao P
and Tan K: Identification of DRP1 as a prognostic factor correlated
with immune infiltration in breast cancer. Int Immunopharmacol.
89:1070782020. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Rehman J, Zhang HJ, Toth PT, Zhang Y,
Marsboom G, Hong Z, Salgia R, Husain AN, Wietholt C and Archer SL:
Inhibition of mitochondrial fission prevents cell cycle progression
in lung cancer. FASEB J. 26:2175–2186. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Gao T, Zhang X, Zhao J, Zhou F, Wang Y,
Zhao Z, Xing J, Chen B, Li J and Liu S: SIK2 promotes reprogramming
of glucose metabolism through PI3K/AKT/HIF-1α pathway and
Drp1-mediated mitochondrial fission in ovarian cancer. Cancer Lett.
469:89–101. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Nagdas S, Kashatus JA, Nascimento A,
Hussain SS, Trainor RE, Pollock SR, Adair SJ, Michaels AD, Sesaki
H, Stelow EB, et al: Drp1 promotes KRas-driven metabolic changes to
drive pancreatic tumor growth. Cell Rep. 28:1845–1859.e5. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|