Introduction
Synovial sarcoma (SS) is a malignant mesenchymal
tumor that is characterized by partial epithelial differentiation,
and it constitutes 8–10% of soft tissue sarcoma (1,2). Despite
its low incidence, SS is not easy to diagnose, progresses rapidly,
and has a low 5-year survival rate that ranges between 21–40%
(1,2). While sensitive to a variety of
chemotherapeutic drugs, distant metastasis is another malignant
manifestation of SS. Patients with SS often exhibit distant
metastasis at early stages, and the majority of metastatic sites
are concentrated in the lymph nodes, lungs and the liver (3,4). In
fact, the main cause of death in patients with SS is tumor
recurrence and metastasis, wherein ~40% of patients with SS develop
lung, liver and regional lymph node metastasis within 2 years
(5,6). Therefore, elucidating the mechanisms
underlying SS recurrence and metastasis would greatly improve the
survival rate of patients.
Long non-coding RNAs (lncRNAs) are a type of RNA,
that are >200 bp in length, which generally have no open reading
frame and do not encode a protein. In addition, they are usually
transcribed by RNA polymerase II and are composed of multiple
spliced exons (7,8). Recently, various lncRNAs have been
associated with the development of different types of cancer
(7,9), and multiple studies have shown that
lncRNA HOTAIR can promote the progression of neuroglioma (10,11). In
this regard, the lncRNA HOTAIR has been shown to act as an
oncogene, which is aberrantly expressed in multiple malignant tumor
tissues, including colorectal cancer (12), pancreatic cancer (13), hepatocellular carcinoma (14), rhinitis (15) and cervical cancer (16). In addition, HOTAIR has been
demonstrated to promote cancer metastasis (17). It has been shown that HOTAIR can
regulate the biological characteristics of gastric cancer cells and
glioma cells by binding to and inhibiting microRNA (miR)-126
expression (11,18). On the other hand, stromal
cell-derived factor-1 (SDF-1), a target of miR-126 (19), has been shown to be highly expressed
in SS tissues and to be associated with poor prognosis in patients
with SS (20).
However, the expression of HOTAIR in SS tissues and
its association with miR-126 remains unclear. In this regard, the
hypothesis used in the present study was that the association
between HOTAIR expression and the prognosis of patients with SS is
associated with the regulation of SDF-1 via miR-126 inhibition.
Materials and methods
Synovial sarcoma tissues and cell
lines
As a prospective study, a total of 54 synovial
sarcoma tissues from the limbs were collected from patients at the
Fourth Hospital of Hebei Medical University (Shijiazhuang, China)
between January 2011 and December 2017. The inclusion criteria were
as follows: i) Synovial sarcoma identified by pathological
diagnosis; ii) complete clinical and histological features,
including sex, age, tumor size, histological grade, distant
metastasis and the 8th Edition of American Joint Committee on
Cancer (AJCC) (21) staging; iii) no
radiotherapy, chemotherapy, immunotherapy or molecular targeting
treatment prior to surgery; iv) regular follow-up or review
following surgery and a clear cause or time of death; v) signed
informed consent. The exclusion criteria were: i) Combined with
other malignant tumors; ii) postoperative death due to non-tumor
related causes; iii) combined with other severe diseases, such as
chronic infection, organ disorders and cardiovascular and
cerebrovascular diseases; iv) poor physical fitness, unable to
tolerate surgery or other related examinations, poor postoperative
mental state, and the prognosis affected by the basic conditions;
v) postoperative follow-up is lost or the cause of death is
unclear. In addition, 10 normal synovium tissues from the limbs
were collected at the same hospital, over the same time period,
from patients who underwent limb amputation with no history of
joint disease. The tissues were snap-frozen in liquid nitrogen
until required. Informed consent was obtained from all patients
participating in the study or from their family members. The study
was approved and monitored by the Ethics Committee of The Fourth
Hospital of Hebei Medical University.
Wild-type (WT) SW982 (HTB-93; American Type Culture
Collection) cell lines were cultured at 37°C and 5% CO2
in DMEM (Thermo Fisher Scientific, Inc.) supplemented with 10% FBS
(Thermo Fisher Scientific, Inc.). SDF-1 knockout (KO) SW982 cell
lines were obtained from Synbio Technologies LLC., and were
cultured under the same conditions as the WT-SW982. WT cells were
used as the untreated control group.
Cell proliferation assay
A MTT Cell Proliferation and Cytotoxicity Assay kit
(Beyotime Institute of Biotechnology) was used to assess cell
proliferation according to the manufacturer's instructions. A total
of 2×103 SW982 cells/well were initially seeded in a
96-well plate for the MTT assay. Following 24-h culture, the medium
was removed, the cells were washed twice with PBS, and MTT reagent
was added to incubate for ≥2 h at 37°C. The MTT solvent included in
the assay kit was used to dissolve the purple formazan, and the
absorbance was measured at 570 nm.
Cell transfection
Small interfering (si)-negative control (NC),
si-HOTAIR-1, si-HOTAIR-2, miR-126-NC, miR-126-mimic and
miR-126-inhibitor were designed and synthesized by Shenggong
Bioengineering Co., Ltd. For siRNA or miRNA mimic/inhibitor
transfection, 50 nmol/l siRNA, miRNA mimic or inhibitor was
transfected into 2.5×106 cells using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) for 6 h at 37°C according to the manufacturer's
protocol. At 48 h post-transfection, the follow-up experiments were
conducted. The sequences were as follows: si-NC:
5′-GAAACAUUAUGAAAAUCAAUU-3′; si-HOTAIR-1:
5′-AGACUAAGACGGAUAACGCGU-3′; si-HOTAIR-2:
5′-ACAAUAUCUACUUUGGAUCAC-3′; mimic-NC: 5′-AUCUCAACUUCGCGGCA-3′;
miR-126-mimic: 5′-UCGUACCGUGAGUAAU-3′; inhibitor-NC:
5′-AUUGCUGGCUAAUAACG-3′; miR-126-inhibitor:
5′-AGCAUGGCACUCAUUA-3′.
Dual-luciferase reporter assay
First, WT-SDF-1 and mutant (MUT)-SDF-1 were
integrated into pmirGLO plasmids (Promega Corporation) and
co-transfected into cells (1×106 cells/well) with
miR-126-NC, miR-126-mimic and miR-126-inhibitor (50 nmol/l) using
Lipofectamine® 2000 for 6 h at 37°C. At 48 h
post-transfection, the Dual-Lucy Assay kit (Beijing Solarbio
Science & Technology Co., Ltd.) was used to detect the activity
of luciferase according to the manufacturer's instructions.
Briefly, cells were collected at 48 h post-transfection and lysed
for 5 min on ice before centrifugation (12,000 × g for 1 min at
room temperature) to collect the cell supernatant. Subsequently, 5X
firefly or Renilla luciferase reaction solution were added
to the cell lysate, and the enzyme activity was detected using a
multifunctional microplate reader (Thermo Fisher Scientific, Inc.),
and the enzyme activity was standardized to that of Renilla
luciferase.
Reverse transcription-quantitative PCR
(RT-qPCR)
The mRNA expression levels of HOTAIR and miR-126
were detected using the RT-qPCR Fluorescence
quantitative-polymerase chain reaction kit (Hangzhou Bori
Technology Co., Ltd.), as previously described (22). Total RNA was extracted using RNAiso
Plus (Takara Bio, Inc.) and reverse transcribed into cDNA using the
PrimeScript RT reagent kit with gDNA Eraser (Takara Bio, Inc.) at
37°C for 15 min and 85°C for 5 sec. U6 was used as a reference gene
for miRNA (miR-126 and miR-429) measurements, whereas GAPDH was
used as a reference gene for lncRNA HOTAIR. The following PCR
primers were used: HOTAIR forward, 5′-GGCAAATGTCAGAGGGTT-3′ and
reverse, 5′-GTGTAACAGGCAGGTGGA-3′; GAPDH forward,
5′-GGCTGTATTCCCCTCCATCG-3′ and reverse,
5′-CCAGTTGGTAACAATGCCATGT-3′; U6 forward, 5′-CTCGCTTCGGCAGCACA-3′;
and reverse, 5′-AACGCTTCACGAATTTGCGT-3′; and miR-126 forward,
5′-ACACTCCAGCTGGGTCGTACCGTG-3′ and reverse, 5′-TGGTGTCGTGGAGTCG-3′.
The 2−ΔΔCq method was used to calculate quantitative
gene expression (23).
Western blot analysis
Protein expression in SW982 cells was measured using
western blot analysis as previously described (22), and GAPDH was used as the a loading
control. Proteins were separated using 10% SDS-PAGE at 90 V for 1.5
h, and transferred on to PVDF membranes (EMD Millipore) at 300 mA
for 1 h. The membranes were then blocked in 5% skimmed milk at room
temperature for 1 h, then incubated with the primary antibody for 2
h at room temperature, followed by the secondary antibody for 1 h
at room temperature. The antibodies used were as follows: Anti-CDK1
[EPR165] (1:1,000; cat. no. ab133327; Abcam), anti-Cdk2 [E304]
(1:1,000; cat. no. ab32147; Abcam), anti-cyclin D1 (1:2,000; cat.
no. ab226977, Abcam), anti-p21 [HUGO291] (1:1,500; cat. no.
ab107099; Abcam), anti-p53 (DO-1) (1:500; cat. no. sc-126; Santa
Cruz Biotechnology, Inc.), anti-MMP9 (1:1,000; cat. no. ab38898;
Abcam), anti-vimentin [VI-10] (1:2,000; cat. no. ab20346; Abcam),
anti-E-cadherin [4A2] (1:500; cat. no. ab231303; Abcam),
anti-N-cadherin [EPR1791-4] (1:1,000; cat. no. ab76011; Abcam),
SDF1/CXCL12 (D32F9) Rabbit mAb (1:1,500; cat. no. 3350; Cell
Signaling Technology, Inc.), HRP Conjugate Goat Anti-Rabbit IgG
(H+L) (1:3,000; cat. no. HS101-01) and HRP Conjugate Goat
Anti-Mouse IgG (H+L) (1:3,000; cat. no. HS201-01) (both TransGen
Biotech Co., Ltd.). The proteins were visualized with ECL solution
(Beijing Xinjingke Biotechnologies Co., Ltd), followed by
densitometry analysis using ImageJ v1.8.0 (National Institutes of
Health). GAPDH was loaded as control.
Cell invasion assay
A 24 mm Transwell® with 3.0 µm Pore
Polycarbonate Membrane Insert (Corning Inc.) was used to assess the
invasive capacity of SW982 cells, as previously described (24). In brief, 25 µl Matrigel (diluted with
3X serum-free medium; Sigma-Aldrich; Merck KGaA) was added to the
upper chamber of the Transwell plate (Corning, Inc.) for 30 min at
37°C. A total of 0.5×106 cells/ml were added into the
Transwell upper chamber in serum-free DMEM, and media containing
20% FBS (Gibco; Thermo Fisher Scientific, Inc.) was added into the
lower chamber. The plate was incubated at 37°C for 24 h.
Subsequently, the Transwell insert was removed and washed twice
with PBS, and 100% methanol was used for fixation for 30 min at
room temperature, followed by drying. The membrane was stained with
crystal violet for 20 min at room temperature, and the relative
migration was determined by measuring the absorbance at 595 nm.
Wound healing assay
A total of 5×105 cells/well were seeded
with 2 ml medium in a 6-well plate. A scratch perpendicular to the
back of the horizontal line was created using a vertically
positioned (not tilted) pipette tip against a ruler, and images
were captured determine the distance the wound at the start of the
assay using a light microscope (Olympus Corporation) (×40
magnification). Serum-free DMEM medium was added after washing
three times with PBS to remove the scratched cells. Images were
captured after incubation for 24 h at 37°C in a humidified
incubator with 5% CO2. ImageJ v1.8.0 was used for
quantification.
Cell cycle and apoptosis assay
The cell cycle phases were detected using the Cell
Cycle and Apoptosis Analysis kit (cat. no. 40301ES50; Shanghai
Yeasen Biotechnology Co., Ltd.) using the MACSQuant®
Analyzer 10 Flow cytometer (Miltenyi Biotec GmbH), according to the
manufacturer's instructions. Data were analyzed by FlowJo X 10.0.7
software (Beckman Coulter, Inc.).
Immunofluorescence assay
A total of 3×104 cells/well were seeded
onto a Lab-Tek Chambered cover glass (cat. no. LOT1228622; Thermo
Fisher Scientific, Inc.). After incubation for 24 h, the medium was
removed, and the cells were washed twice with PBS. The cells were
incubated with 0.3% Triton X-100 in PBS-Tween-20 (PBS-T) for 30 min
at room temperature, washed twice with PBS-T and incubated with the
SDF-1 antibody (1:50; cat. no. ab155090; Abcam) for 30 min at room
temperature. After washing three times with PBS-T, the cells were
incubated with the Goat Anti-Rabbit IgG H&L (Alexa
Fluor® 488) secondary antibody (1:500; cat. no.
ab150077; Abcam) for 1 h at room temperature. The nuclei were
finally stained with 5 µg/ml DAPI for 5 min at room temperature. A
Leica TCS SP5 microscope with the LAS AF Lite v4.0 image browser
software (DMI3000; Leica Microsystems GmbH) was used for
fluorescence detection and analysis.
Statistical analysis
The data was analyzed using SPSS v20.0 software
(IBM, Corp.). Pairwise comparisons between groups were performed
using either the unpaired Student's t-test, the χ2 test
or Fisher's exact test, while multi-group statistical analysis was
performed using one-way ANOVA followed by the Tukey's post hoc
test. Pearson's coefficient was used for correlation analysis.
P<0.05 was considered to indicate a statistically significant
difference.
Results
HOTAIR is highly expressed in SS
tissues
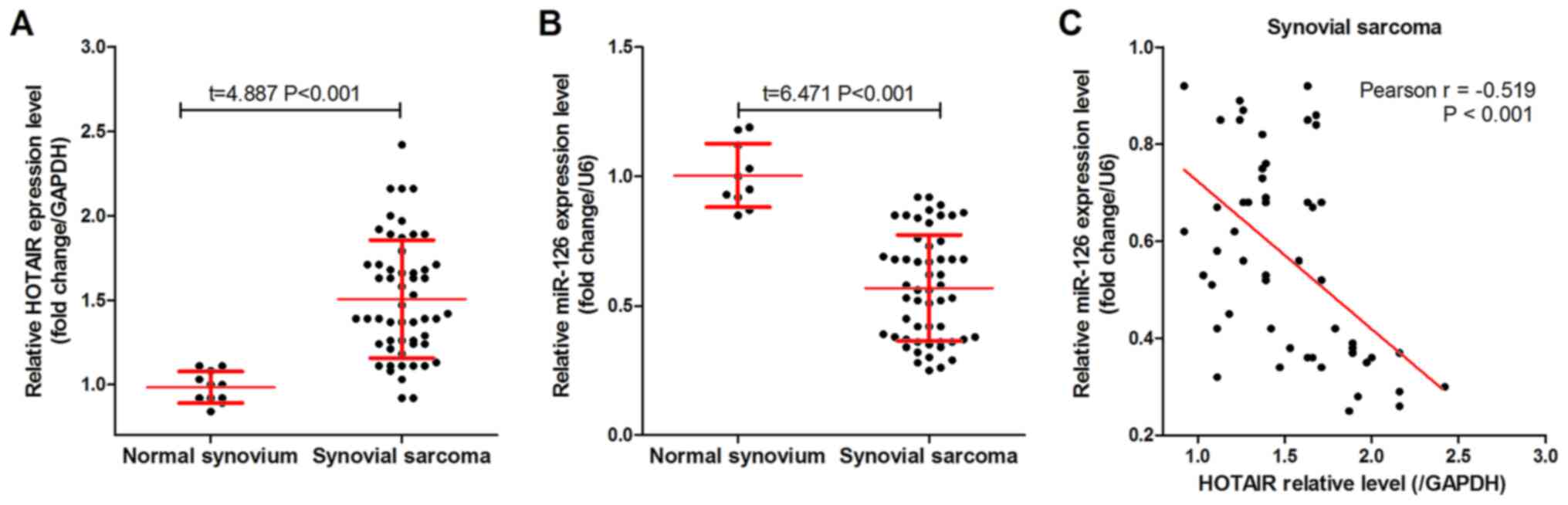
As shown in Fig. 1A,
HOTAIR was highly expressed in the 54 SS tissues compared with that
in the 10 normal synovium tissues. By contrast, miR-126 was
expressed at low levels in the SS tissues (Fig. 1B) compared with that in the normal
synovium tissues, and was negatively correlated with HOTAIR
expression level in the SS tissues (Fig.
1C). In addition, patients with SS were divided into groups
based on the median HOTAIR or miR-126 expression. Analysis using
either χ2 or Fisher's exact tests found that both HOTAIR
and miR-126 expression levels were significantly associated with
histological grade, distant metastasis and AJCC staging in the
patients with SS (Table I).
 | Table 1.Clinicopathological variables and
HOTAIR/miR-126 expression level. |
Table 1.
Clinicopathological variables and
HOTAIR/miR-126 expression level.
|
|
| HOTAIR | miR-126 |
|---|
|
|
|
|
|
|---|
| Variable | Number | Low, n | High, n | P-value | Low, n | High, n | P-value |
|---|
| Sex |
|
|
| 0.846 |
|
| 0.409 |
|
Female | 23 | 12 | 11 |
| 13 | 10 |
|
|
Male | 31 | 17 | 14 |
| 14 | 17 |
|
| Age, years |
|
|
| 0.816 |
|
| 0.413 |
|
≥30 | 25 | 13 | 12 |
| 14 | 11 |
|
|
<30 | 29 | 16 | 13 |
| 13 | 16 |
|
| Tumor size, cm |
|
|
| 0.918 |
|
| 0.580 |
|
<5 | 32 | 17 | 15 |
| 15 | 17 |
|
| ≥5 | 22 | 12 | 10 |
| 12 | 10 |
|
| Histological
grade |
|
|
| 0.033 |
|
| 0.021 |
| I | 12 | 3 | 9 |
| 10 | 2 |
|
| II | 20 | 8 | 12 |
| 9 | 11 |
|
|
III | 22 | 15 | 7 |
| 8 | 14 |
|
| Distant
metastasis |
|
|
| 0.001 |
|
| 0.001 |
| No | 34 | 24 | 10 |
| 11 | 23 |
|
|
Yes | 20 | 5 | 15 |
| 16 | 4 |
|
| AJCC staging |
|
|
| 0.015 |
|
| 0.003 |
|
I/II | 25 | 9 | 16 |
| 7 | 18 |
|
|
III/IV | 29 | 20 | 9 |
| 20 | 9 |
|
HOTAIR knockdown inhibits cellular
proliferation and blocks the cells in the G1 phase
A MTT assay was used to measure the proliferation of
the SW982 cells in vitro following knockdown of HOTAIR,
using si-HOTAIR-1 and si-HOTAIR-2 (Fig.
2A). The results showed that HOTAIR knockdown significantly
reduced the cell proliferation of the SW982 cells (Fig. 2B). In addition, HOTAIR knockdown
significantly increased the ratio of the SW982 cells in the
G1/G0 phase of the cell cycle (Fig. 2C). In addition, HOTAIR knockdown
decreased the protein expression level of CDK1, CDK2 and cyclin D1,
while it increased the expression level of p21 and p53 (Fig. 2D).
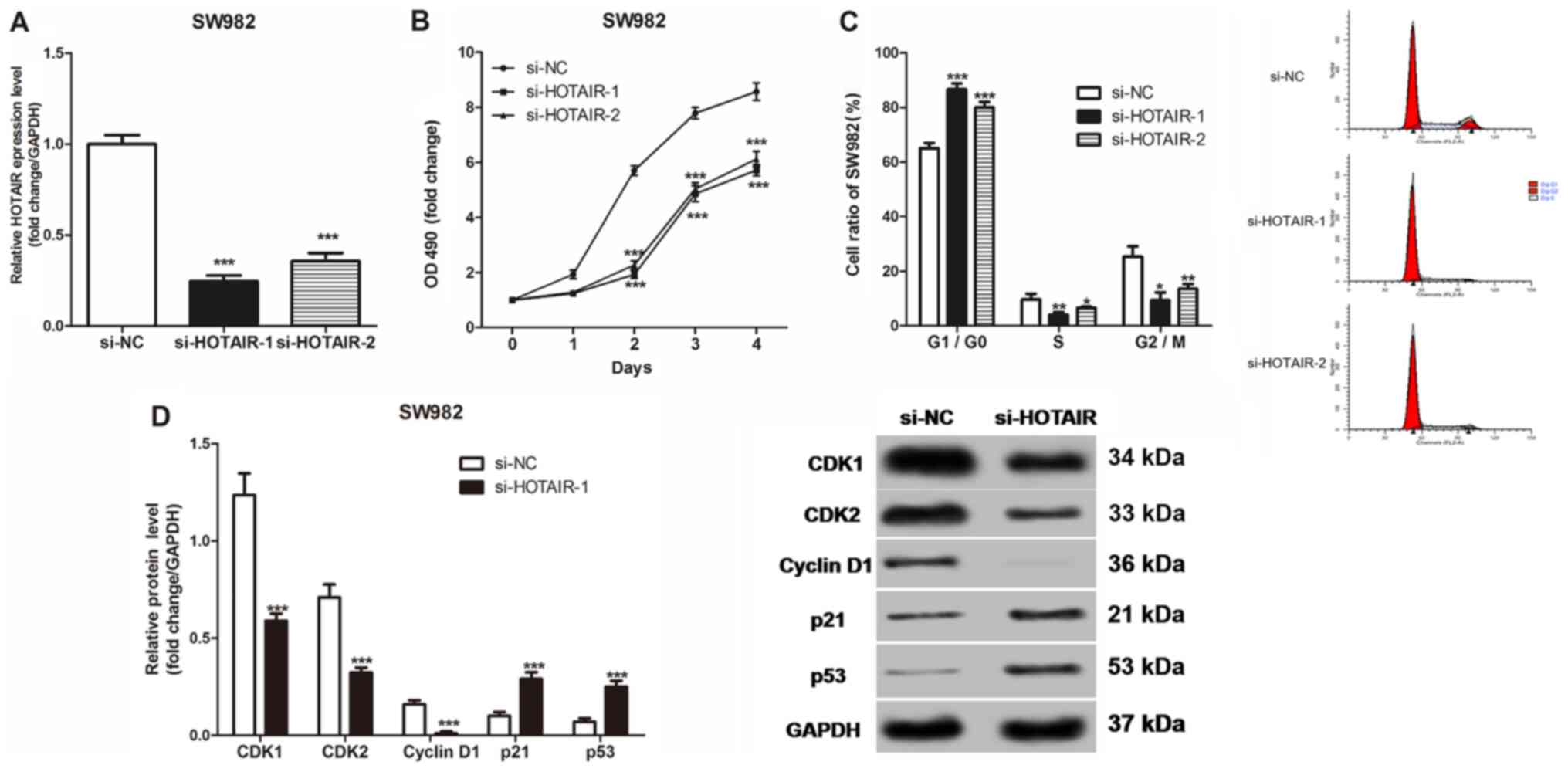 | Figure 2.Effects of HOTAIR expression on the
proliferation, cell cycle and cell cycle-related protein expression
of the SS cells in vitro. (A) HOTAIR expression level in the
SW982 cells transfected with si-NC or 2 siRNAs. (B) The
proliferation of the SW982 cells was measured using MTT assays. (C)
The cell cycle of SW982 was detected using flow cytometry. (D)
Western blot analysis was used to measure CDK1, CDK2, cyclin D1,
p21, and p53 protein expression levels in the SW982 cells.
*P<0.05, **P<0.01, ***P<0.001 vs. si-NC. si, short
inhibiting; NC, negative control; OD, optical density. |
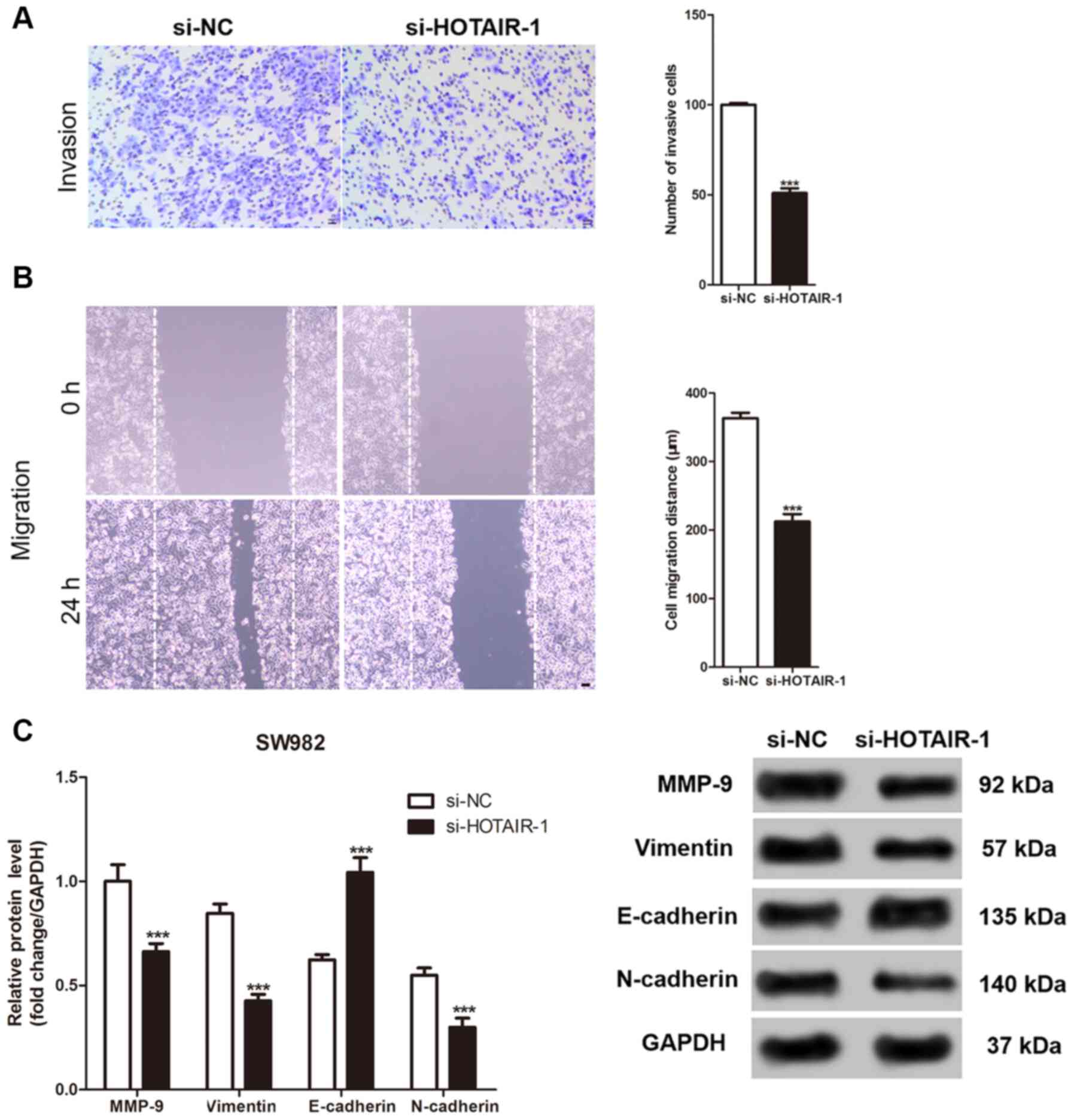
HOTAIR knockdown inhibits invasion and
migration of the SS cells
The Transwell and wound healing assays were used to
evaluate the invasive and migration abilities of the SS cells
following knockdown of HOTAIR, respectively. As shown in Fig. 3A and B, the number of invasive SS
cells was significantly reduced after HOTAIR knockdown. In
addition, the migration distance by the SS cells was also
significantly reduced. Similarly, the expression level of the
proteins involved in cellular invasion and migration were measured.
While HOTAIR knockdown significantly decreased the protein
expression level of MMP-9, vimentin and N-cadherin, it increased
the expression level of E-cadherin in the SW982 cells compared with
that in the control cells (Fig.
3C).
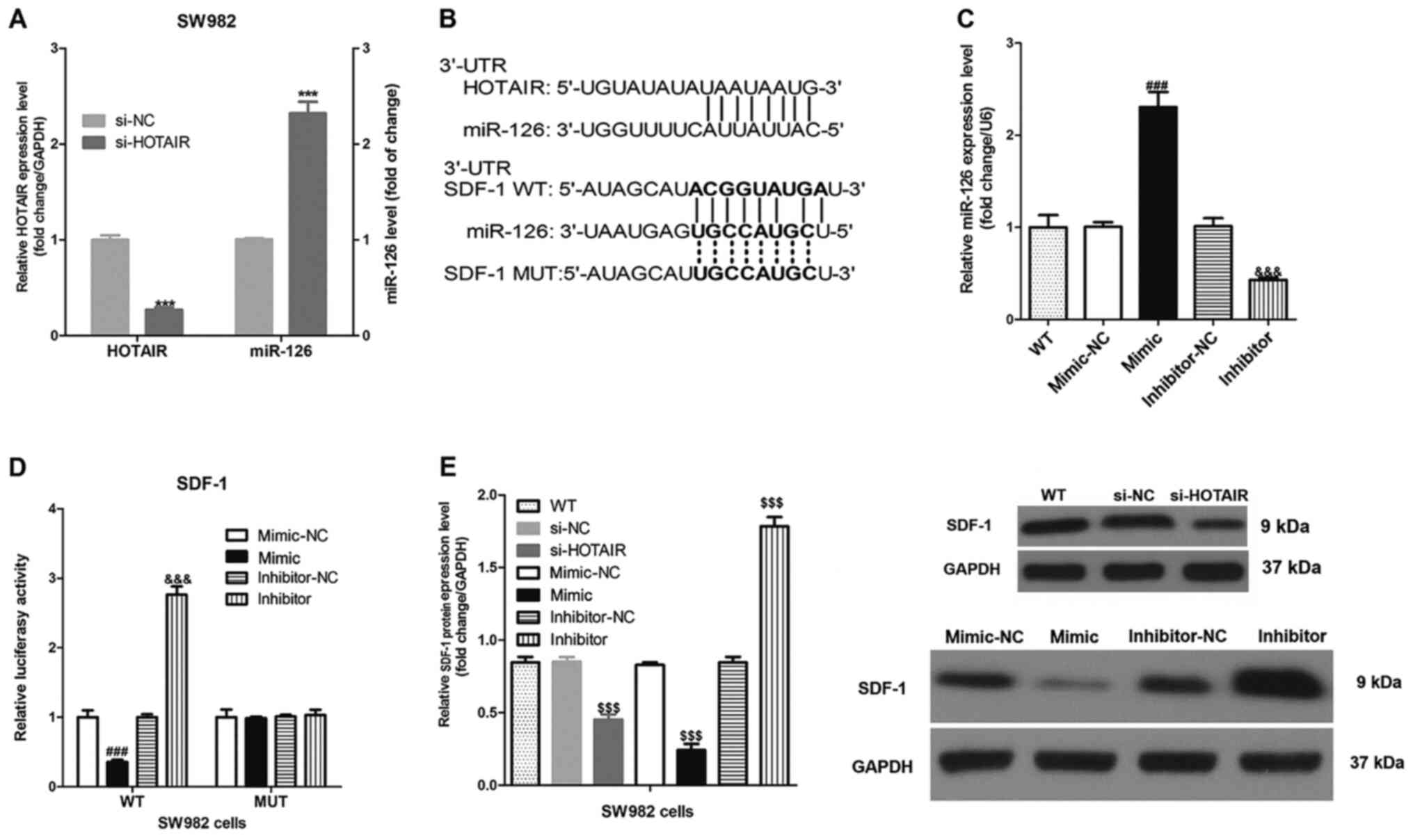
HOTAIR promotes SDF-1 expression level
by inhibiting miR-126
HOTAIR knockdown significantly increased miR-126
expression level in the SW982 cells (Fig. 4A). Notably, sequence analysis showed
that HOTAIR and SDF-1 contained complementary sequences to miR-126
(Fig. 4B). In accordance with the
sequence analysis, a luciferase gene reporter assay indicated that
that the miR-126-mimics significantly reduced, and the
miR-126-inhibitor significantly increased the luciferase activity
in SW982 cells transfected with WT-SDF-1, whereas the
miR-126-mimics and miR-126-inhibitor did not affect the luciferase
activity in SW982 cells transfected with MUT-SDF-1 (Fig. 4C and D). Both HOTAIR knockdown and
miR-126-mimics significantly decreased SDF-1 protein expression
level, whereas treatment with miR-126 inhibitor significantly
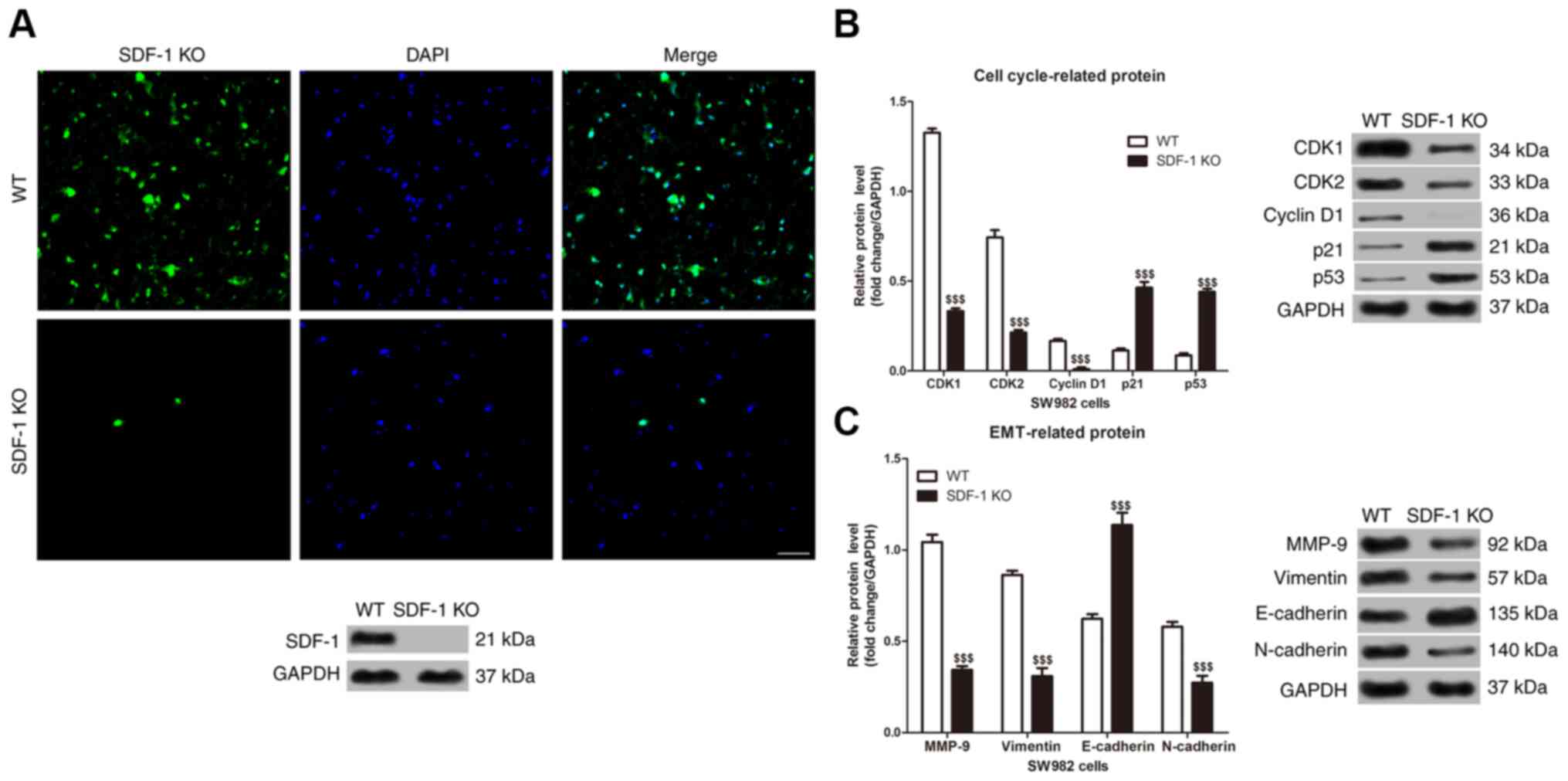
increased the protein expression level in the SW982 cells (Fig. 4E). Furthermore, immunofluorescence
and western blot analysis showed that the expression of SDF-1
protein in SDF-1 KO SW982 cells was significantly lower than that
of wild-type SW982 cells (Fig. 5A).
In addition, SDF-1 knockdown suppressed the protein expression
level of CDK1, CDK2, cyclin D1, MMP-9, vimentin and N-cadherin,
while it induced the expression level of p21, p53 and E-cadherin in
the SW982 cells (Fig. 5B and C).
Discussion
It has been shown in our previous study that SDF-1
was highly expressed in SS tissues, which was also associated with
poor prognosis in patients with SS (20). Notably, SDF-1 was a target gene of
miR-126. Indeed, van Solingen et al (25) showed that miR-126 increased the
number of endothelial cells by inhibiting SDF-1 expression in the
endothelium. Similarly, Zhang et al (19) demonstrated that miR-126 could inhibit
the recruitment of mesenchymal stem cells and inflammatory
monocytes into the tumor tissues by inhibiting SDF-1 expression
level, thereby inhibiting the metastasis of breast cancer cells.
Notably, HOTAIR was shown to promote the metastasis of cancer cells
by inhibiting miR-126 expression level in different malignant tumor
cells, such as breast, gastric and liver cancer cells (17). In the present study, it was shown
that HOTAIR mRNA was highly expressed and was negatively correlated
with miR-126 expression level in SS tissues, but was also
significantly associated with histological grade, distant
metastasis and AJCC staging in patients with SS. These results
suggested that HOTAIR was associated with the development of
SS.
The knockdown of HOTAIR in the SS cells inhibited
proliferation, and the invasion and migration abilities of the SS
cells in vitro, which is consistent with previous studies
reporting HOTAIR as an oncogene in various malignancies. For
example, Gupta et al (17)
showed that HOTAIR could promote cancer metastasis through
epigenetic reprograming of the cellular chromatin. As a non-coding
RNA, HOTAIR may only act indirectly to regulate cellular functions
through miRNA and mRNA regulation. In this regard, the results from
the present study showed that HOTAIR knockdown could increase
miR-126 expression level, which is consistent with previous studies
reporting miR-126 to be a target of HOTAIR (11,18).
miRNAs are non-coding, single-stranded RNAs, that
are 20–25 nucleotides in length, which can regulate cellular
proliferation, differentiation, apoptosis, metabolism, invasion and
migration through the post-transcriptional regulation of target
genes (26,27). Multiple miRNAs have been recognized
as biomarkers for the diagnosis of multiple diseases and have been
demonstrated to serve important roles in the development of cancer,
acute myocardial infarction and rheumatoid arthritis (28). Previous studies have shown (29,30) that
the development of breast cancer was associated with the abnormal
expression of miR-1271 and miR-192-5p. miR-126, a tumor suppressor,
was shown to be abnormally expressed in numerous tumors, where it
regulated multiple biological processes in tumors, such as
proliferation, apoptosis, invasion, and migration (31). Feng et al (32) demonstrated that miR-126 could inhibit
the proliferation, invasion and migration of gastric cancer cells
by inducing cell-cycle arrest at the G1/G0
phase. Hamada et al (33), on
the other hand, showed that miR-126 could play a tumor suppressor
role in pancreatic tumor cells through the regulation of ADAM9.
Taken together, these findings indicated that HOTAIR could regulate
the proliferation, invasion and migration of SS cells via miR-126
regulation.
There have been two major interaction mechanisms
reported between lncRNA and miRNA. The first mechanism suggested
that lncRNA could act as ‘molecular sponges’ to block the
post-transcriptional inhibitory effects of miRNAs on downstream
target genes. These molecular sponge lncRNAs are called competitive
endogenous RNA (34). The other
mechanism suggested that lncRNAs and miRNAs could increase the
stability of target genes by competing with miRNAs to bind target
genes (34). In the present study,
it was shown that HOTAIR knockdown increased miR-126 expression
level, and that miR-126-mimics inhibited the protein expression
level of SDF-1 in the SS cells. This suggested that the high
expression level of HOTAIR may act as an oncogene in the SS cells
by promoting SDF-1 expression level by regulating miR-126.
Furthermore, it was also shown that SDF-1 knockout and HOTAIR
knockdown resulted in similar molecular signatures in the SW982
cells, whereby they significantly reduced CDK1, CDK2, cyclin D1,
MMP-9, vimentin and N-cadherin protein expression levels, while
inducing p21, p53 and E-cadherin expression level.
Cyclin network systems, including CDKs and CDKIs,
play important roles in the regulation of the G1/S phase
of the cell cycle (35,36). p21 is a negative regulator of the
cell cycle and is a member of the CDKI family. p21 can induce cell
cycle arrest and block cell proliferation by binding to cyclins,
CDKs or cyclin-CDKs complexes (37,38). In
addition, the p21 gene has a specific binding site to the p53
protein and is considered one of the most important target genes of
p53; when the cell is subjected to various stressors (such as
chemical drugs or radiation), the WT p53 protein acts on the p21
gene and rapidly induces its expression (39). The p21 protein, encoded by the p21
gene, is a cell cycle inhibitor that is currently considered to
have the broadest kinase inhibitor activity (39). Indeed, p21 can bind to and inhibit a
wide range of cyclin-CDK complexes, such as cyclinD1-CDK4,
cyclinE-CDK2, and cyclinA-CDK2, but also has a weak inhibitory
activity against cyclinB-related complexes. p21 inhibits the
activity of cyclinD1-CDK4 and cyclinE-CDK2, and hence, can block Rb
protein phosphorylation and the release of the E2F protein, thereby
inducing cell cycle arrest in the G1 phase (40). On the other hand, MMP-9, vimentin,
E-cadherin and N-cadherin are key regulators of
epithelial-mesenchymal transition, and hence, can regulate the
metastatic ability of cancer cells (41,42).
In summary, the results from the present study
showed that HOTAIR could be highly expressed in SS tissues, where
it can promote the proliferation, invasion and migration of the SS
cells by promoting SDF-1 protein expression through the regulation
of miR-126 expression. Despite these important results, the number
of SS tissues, as well as their freezing time and conditions may
affect the expression level of HOTAIR and miR-126. Therefore,
future studies, using larger sample sizes and more cell lines are
required to further support the findings.
Acknowledgements
Not applicable.
Funding
This research was supported by the fund of Youth
Science and Technology Project of Hebei Provincial Department of
Health (grant no. 20170692).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
QF conceived and designed the current study and
contributed to writing the manuscript. QF and DW performed the
experiments. QF and DW confirm the authenticity of all the raw
data. DW, PG, ZZ and JF analyzed and interpreted the data. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
The study was approved and monitored by the Ethics
Committee of The Fourth Hospital of Hebei Medical University
(Hebei, China). Informed consent was provided by all the patients
participating in the study or from their family members.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Palmerini E, Staals EL, Alberghini M,
Zanella L, Ferrari C, Benassi MS, Picci P, Mercuri M, Bacci G and
Ferrari S: Synovial sarcoma: Retrospective analysis of 250 patients
treated at a single institution. Cancer. 115:2988–2998. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bergh P, Meis-Kindblom JM, Gherlinzoni F,
Berlin O, Bacchini P, Bertoni F, Gunterberg B and Kindblom LG:
Synovial sarcoma: Identification of low and high risk groups.
Cancer. 85:2596–2607. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Baccari-Ezzine S, Chelbi E and Bouzaidi K:
Intracardiac metastasis of primary synovial sarcoma of the lung.
Asian Cardiovasc Thorac Ann. 21:623. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Amankwah EK, Conley AP and Reed DR:
Epidemiology and therapies for metastatic sarcoma. Clin Epidemiol.
5:147–162. 2013.PubMed/NCBI
|
|
5
|
Wushou A and Miao XC: Tumor size predicts
prognosis of head and neck synovial cell sarcoma. Oncol Lett.
9:381–386. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bakri A, Shinagare AB, Krajewski KM,
Howard SA, Jagannathan JP, Hornick JL and Ramaiya NH: Synovial
sarcoma: Imaging features of common and uncommon primary sites,
metastatic patterns, and treatment response. AJR Am J Roentgenol.
199:208–215. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bhan A, Soleimani M and Mandal SS: Long
Noncoding RNA and Cancer: A new paradigm. Cancer Res. 77:3965–3981.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hon CC, Ramilowski JA, Harshbarger J,
Bertin N, Rackham OJ, Gough J, Denisenko E, Schmeier S, Poulsen TM,
Severin J, et al: An atlas of human long non-coding RNAs with
accurate 5′ends. Nature. 543:199–204. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chandra Gupta S and Nandan Tripathi Y:
Potential of long non-coding RNAs in cancer patients: From
bio-markers to therapeutic targets. Int J Cancer. 140:1955–1967.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rao AKDM, Rajkumar T and Mani S:
Perspectives of long non-coding RNAs in cancer. Mol Biol Rep.
44:203–218. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu L, Cui S, Wan T, Li X, Tian W, Zhang
R, Luo L and Shi Y: Long non-coding RNA HOTAIR acts as a competing
endogenous RNA to promote glioma progression by sponging
miR-126-5p. J Cell Physiol. 233:6822–6831. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kogo R, Shimamura T, Mimori K, Kawahara K,
Imoto S, Sudo T, Tanaka F, Shibata K, Suzuki A, Komune S, et al:
Long noncoding RNA HOTAIR regulates polycomb-dependent chromatin
modification and is associated with poor prognosis in colorectal
cancers. Cancer Res. 71:6320–6326. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kim K, Jutooru I, Chadalapaka G, Johnson
G, Frank J, Burghardt R, Kim S and Safe S: HOTAIR is a negative
prognostic factor and exhibits pro-oncogenic activity in pancreatic
cancer. Oncogene. 32:1616–1625. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yang Z, Zhou L, Wu LM, Lai MC, Xie HY,
Zhang F and Zheng SS: Overexpression of long Non-coding RNA HOTAIR
predicts tumor recurrence in hepatocellular carcinoma patients
following liver transplantation. Ann Surg Oncol. 18:1243–1250.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nie Y, Liu X, Qu S, Song E, Zou H and Gong
C: Long non-coding RNA HOTAIR is an independent prognostic marker
for nasopharyngeal carcinoma progression and survival. Cancer Sci.
104:458–464. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kim HJ, Lee DW, Yim GW, Nam EJ, Kim S, Kim
SW and Kim YT: Long non-coding RNA HOTAIR is associated with human
cervical cancer progression. Int J Oncol. 46:521–530. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gupta RA, Shah N, Wang KC, Kim J, Horlings
HM, Wong DJ, Tsai MC, Hung T, Argani P, Rinn JL, et al: Long
non-coding RNA HOTAIR reprograms chromatin state to promote cancer
metastasis. Nature. 464:1071–1076. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yan J, Dang Y, Liu S, Zhang Y and Zhang G:
LncRNA HOTAIR promotes cisplatin resistance in gastric cancer by
targeting miR-126 to activate the PI3K/AKT/MRP1 genes. Tumor Biol.
Nov 30–2016.(Epub ahead of print). doi: 10.1007/s13277-016-5448-5.
View Article : Google Scholar
|
|
19
|
Zhang Y, Yang P, Sun T, Li D, Xu X, Rui Y,
Li C, Chong M, Ibrahim T, Mercatali L, et al: miR-126 and miR-126*
repress recruitment of mesenchymal stem cells and inflammatory
monocytes to inhibit breast cancer metastasis. Nat Cell Biol.
15:284–294. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Feng Q, Guo P, Wang J, Zhang X, Yang HC
and Feng JG: High expression of SDF-1 and VEGF is associated with
poor prognosis in patients with synovial sarcomas. Exp Ther Med.
15:2597–2603. 2018.PubMed/NCBI
|
|
21
|
Tanaka K and Ozaki T: New TNM
classification (AJCC eighth edition) of bone and soft tissue
sarcomas: JCOG Bone and Soft Tissue Tumor Study Group. Jpn J Clin
Oncol. 49:103–107. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tao J, Zhang J, Ling Y, McCall CE and Liu
TF: Mitochondrial Sirtuin 4 resolves immune tolerance in monocytes
by rebalancing glycolysis and glucose oxidation homeostasis. Front
Immunol. 9:4192018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xie Z, Chen W, Chen Y, Wang X, Gao W and
Liu Y: miR-768-3p is involved in the proliferation, invasion and
migration of non-small cell lung carcinomas. Int J Oncol.
51:1574–1582. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
van Solingen C, de Boer HC, Bijkerk R,
Monge M, van Oeveren-Rietdijk AM, Seghers L, de Vries MR, van der
Veer EP, Quax PH, Rabelink TJ and van Zonneveld AJ: MicroRNA-126
modulates endothelial SDF-1 expression and mobilization of
Sca-1(+)/Lin(−) progenitor cells in ischaemia. Cardiovasc Res.
92:449–455. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bartel DP: MicroRNA: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lu J, Getz G, Miska EA, Alvarez-Saavedra
E, Lamb J, Peck D, Sweet-Cordero A, Ebert BL, Mak RH, Ferrando AA,
et al: MicroRNA expression profiles classify human cancers. Nature.
435:834–838. 2009. View Article : Google Scholar
|
|
28
|
Wang J, Chen J and Sen S: MicroRNA as
biomarkers and diagnostics. J Cell Physiol. 231:25–30. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang Y, Xu L and Jiang L: miR-1271
promotes non-small-cell lung cancer cell proliferation and invasion
via targeting HOXA5. Biochem Biophys Res Commun. 458:714–719. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ye M and Zhang J and Zhang J, Miao Q, Yao
L and Zhang J: Curcumin promotes apoptosis by activating the
p53-miR-192-5p/215-XIAP pathway in non-small cell lung cancer.
Cancer Lett. 357:196–205. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Meister J and Schmidt MH: miR-126 and
miR-126*: New players in cancer. ScientificWorldJournal.
10:2090–2100. 2014. View Article : Google Scholar
|
|
32
|
Feng R, Chen X, Yu Y, Su L, Yu B, Li J,
Cai Q, Yan M, Liu B and Zhu Z: miR-126 functions as a tumour
suppressor in human gastric cancer. Cancer Lett. 298:50–63. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hamada S, Satoh K, Fujibuchi W, Hirota M,
Kanno A, Unno J, Masamune A, Kikuta K, Kume K and Shimosegawa T:
miR-126 acts as a tumor suppressor in pancreatic cancer cells via
the regulation of ADAM9. Mol Cancer Res. 10:3–10. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Erratum: The lncRNA XIST exhibits
oncogenic properties via regulation of miR-449a and Bcl-2 in human
non-small cell lung cancer. Acta Pharmacol Sin. 38:4432017.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lim S and Kaldis P: Cdks, cyclins and
CKIs: Roles beyond cell cycle regulation. Development.
140:3079–3093. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Schwartz GK and Shah MA: Targeting the
cell cycle: A new approach to cancer therapy. J Clin Oncol.
23:9408–9421. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Waldman T, Kinzler KW and Vogelstein B:
p21 is necessary for the p53-mediated G1 arrest in human cancer
cells. Cancer Res. 55:5187–5190. 1995.PubMed/NCBI
|
|
38
|
Kim EM, Jung CH, Kim J, Hwang SG, Park JK
and Um HD: The p53/p21 complex regulates cancer cell invasion and
apoptosis by targeting Bcl-2 family proteins. Cancer Res.
77:3092–3100. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Karimian A, Ahmadi Y and Yousefi B:
Multiple functions of p21 in cell cycle, apoptosis and
transcriptional regulation after DNA damage. DNA Repair (Amst).
42:63–71. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Dutto I, Tillhon M, Cazzalini O, Stivala
LA and Prosperi E: Biology of the cell cycle inhibitor p21(CDKN1A):
Molecular mechanisms and relevance in chemical toxicology. Arch
Toxicol. 89:155–178. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zavadil J, Haley J, Kalluri R, Muthuswamy
SK and Thompson E: Epithelial-mesenchymal transition. Cancer Res.
68:9574–9577. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lamouille S, Xu J and Derynck R: Molecular
mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell
Biol. 15:178–196. 2014. View Article : Google Scholar : PubMed/NCBI
|