Introduction
Lung cancer is a common malignant tumor with high
morbidity and mortality rates (1,2). In
2015, ~158,040 patients succumbed to lung cancer in the United
States (3). Non-small cell lung
cancer (NSCLC) accounts for 87% of all lung cancers, and 40% of
NSCLC cases are diagnosed with tumor metastasis (3). Although targeted therapies, such as
gefitinib, are available for patients with gene mutations,
including EGFR mutations and anaplastic lymphoma kinase
rearrangements, these mutations are not common in NSCLC (4). Therefore, most patients are treated
with conventional chemotherapy (5).
Systemic chemotherapy can improve survival and alleviate
disease-related symptoms. However, a large proportion of patients
develop drug resistance after several chemotherapy courses,
resulting in tumor relapse, invasion and metastasis (6,7).
Therefore, it would be of great clinical value to identify novel,
potent targets for the treatment of NSCLC, and to explore their
association with invasion and metastasis of NSCLC.
Receptor tyrosine-protein kinase erbB-2 (ERBB2) is a
185 kDa cell membrane receptor encoded by the oncogene erbB-2,
which is a member of the EGFR family. ERBB2 is involved in multiple
cell events, such as proliferation, differentiation and apoptosis
(8,9). ERBB2 is highly expressed in multiple
malignant tumors, such as breast cancer, ovarian cancer and
hepatocellular carcinoma (8,10–13).
Downregulation of ERBB2 suppresses the proliferation and invasion
of breast cancer and gastric cancer cells (12,14,15).
ERBB2 expression is positively associated with NSCLC, and high
expression levels of ERBB2 are associated with resistance to
radiotherapy and chemotherapy (16,17).
Therefore, ERBB2 may be a potential prognostic biomarker and
therapeutic target for NSCLC.
MicroRNAs (miRNAs/miRs) are endogenous short
non-coding sequences with a length of 20–24 bases. miRNAs serve a
role in multiple biological processes, such as cell proliferation,
differentiation and apoptosis, by regulating protein expression at
the posttranscriptional level (18,19).
miRNAs regulate mRNAs by binding to the 3′ untranslated region
(3′UTR) end of mRNA, degrading mRNA or repressing protein
translation. miR-133a is highly conserved and widely expressed in
various organisms, and can be expressed as the subtypes miR-133a-3p
and miR-133a-5p (20). In a variety
of cancer types, including gastric cancer, bladder cancer and oral
squamous cell carcinoma, miR-133a-3p expression is downregulated
(21–23). Yang et al (24) reported that patients with NSCLC with
high miR-133a expression in tumor cells had a favorable prognosis.
To the best of our knowledge, no reports have demonstrated the
association between miR-133a-3p and NSCLC pathogenesis.
The present study aimed to investigate the
association between dysregulated miR-133a-3p expression and NSCLC
pathogenesis, and to explore the role of miR-133a-3p and ERBB2 in
NSCLC carcinogenesis. The expression of miR-133a-3p and ERBB2 in
NSCLC tissues and cell lines was detected using reverse
transcription-quantitative (RT-q)PCR and western blotting,
respectively. The effects of miR-133a-3p on the malignant
phenotypes of NSCLC cells were then evaluated by multiple
experiments, such as MTT and Transwell assays, and the underlying
mechanisms were further investigated. The present data indicate
that the miR-133a-3p/ERBB2 axis could be a potential therapeutic
target of NSCLC.
Materials and methods
Sample collection
Material was collected from 77 patients (45 males
and 32 females, aged 66.58±14.38 years) primarily diagnosed with
NSCLC who underwent primary surgery at Zhejiang Hospital (Hangzhou,
China) between January 2010 and January 2017. None of the
participants received chemotherapy or radiotherapy before surgery
and none of the participants was diagnosed other malignant diseases
before. The present study was conducted in accordance with the
Helsinki Declaration, and all patients or direct relatives
consented to participate in the study. Furthermore, the present
study was approved by the Ethics Committee of Zhejiang Hospital.
After surgery, the tumor and adjacent healthy tissues (≥1-cm from
the edge of tumor) were collected and stored in liquid nitrogen
(−196°C).
Cell culture and transfection
The normal human lung epithelial cell line BEAS-2B
and human NSCLC HCC827 and H1299 cell lines were purchased from the
Institute of Shanghai Biochemistry and Cell Biology. BEAS-2B was
cultured in BEBM medium (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS (HyClone; Cyvita). HCC827 and H1299 cells
were cultured in RPMI-1640 medium (Invitrogen; Thermo Fisher
Scientific, Inc.) supplemented with 10% FBS and 1%
penicillin/streptomycin (100 U/ml penicillin and 0.1 mg/ml
streptomycin; Thermo Fisher Scientific, Inc.). All cells were
cultured with 5% CO2 at 37°C.
For transfection, HCC827 and H1299 were collected
and seeded in 6-well plates at the density of
1×105/well. After incubated for 24 h, cells treated with
100 nM mimic control (5′-CAGCUGGUUGAAGGGGACCAAA-3′; Shanghai
GenePharma Co., Ltd.), 75 nM miR-133a-3p mimic
(5′-UUUGGUCCCCUUCAACCAGCUG-3′; Shanghai GenePharma Co., Ltd.), 600
ng/well pcDNAn empty vector negative control (pcDNA-NC; Shanghai
GenePharma Co., Ltd.) or 600 ng/well pcDNA-ERBB2 (Shanghai
GenePharma Co., Ltd.) using Lipofectamine 3000 (Invitrogen; Thermo
Fisher Scientific, Inc.) according to the manufacturers' protocols.
The 1 µg nucleic acid fragment and 2 µl Lipofectamine 3000 were
mixed with 1 ml RPMI-1640 medium without serum, and then incubated
for 15 min at room temperature. Then, the mixture was mixed with 4
ml serum-free RPMI-1640 medium and added into plates. After
cultured for 24 h, the culture medium was removed and replaced with
RPMI-1640 medium containing 10% FBS. After 24–72 h, cells were used
for subsequent experiments.
MTT assay
Following treatment with control mimics, miR-133a-3p
mimics or pcDNA-ERBB2, H1299 and HCC827 cells were collected and
seeded into 96-well plates at the density of 1×104
cells/well. After culturing for 24, 48 and 72 h, medium was removed
and replaced with medium containing MTT (KGA312; Nanjing KeyGen
Biotech Co., Ltd.) at a final concentration of 0.5 mg/ml and cells
were cultured for another 4 h at 37°C. Finally, the medium was
replaced with 200 µl DMSO. After 10 min, the absorbance at 490 nm
was measured using a spectrometer (Thermo Fisher Scientific,
Inc.).
Western blotting
The protein expression levels of ERBB2 were assessed
by western blotting. Following treated with different nucleotide
fragments as aforementioned, cells were collected and lysed in RIPA
buffer (Nanjing KeyGen Biotech Co., Ltd.). Samples were quantified
using a Bicinchoninic Acid Assay kit (Beyotime Institute of
Biotechnology). Samples (50 µg per lane) were separated using 10%
SDS-PAGE and then transferred onto 0.45-µm PVDF membranes (Merck
KGaA). The membranes were blocked with 5% skim milk powder
solution, followed by incubation overnight at 4°C with antibodies
against ERBB2 (dilution, 1:1,000; 18299-1-AP; ProteinTech Group,
Inc.) and GAPDH (dilution, 1:3,000; 10494-1-AP; ProteinTech Group,
Inc.). Subsequently, the membranes were incubated with
HRP-conjugated Affinipure Rabbit Anti-Goat IgG(H+L) (dilution,
1:5,000; SA00001-4; ProteinTech Group, Inc.) for 2 h at 37°C.
Subsequently, bands were detected by incubation with ECL mixture
solution (Advansta Inc.), and immediately detected using a Bio-Rad
Gel Imaging System (Bio-Rad Laboratories, Inc.). ImageJ software
(version 1.8.0; National Institutes of Health) was used to analyze
the intensity of the bands.
Reverse transcription-quantitative PCR
(RT-qPCR)
After undergoing different treatments, cells were
collected and lysed using TRIzol (Invitrogen; Thermo Fisher
Scientific, Inc.) to extract total RNA. RNA was transcribed into
cDNA using a cDNA reverse transcription kit (Takara Bio, Inc.). The
conditions used for reverse transcription were as follows: 42°C For
15 min, then 5 sec at 85°C and storage at 4°C for further analysis.
Subsequently, cDNA samples were amplified using an RT-qPCR (Applied
Biosystems; Thermo Fisher Scientific, Inc.) system and
SYBR® Green Premix Ex Taq™ (Takara Bio, Inc.) according
to the manufacturer's protocols. Thermocycling conditions for qPCR
were: Pre-denaturation for 30 sec at 95°C; followed by 40 cycles of
5 sec at 95°C and 30 sec at 60°C; and dissociation at 95°C for 15
sec, 60°C for 30 sec and 95°C for 15 sec. GAPDH and U6 were used as
the reference genes to normalize the expression levels of ERBB2 and
miR-133a-3p, respectively. mRNA and miRNA expression levels were
quantified using the 2−∆∆Cq method (25). The primers used were as follows:
ERBB2 forward, 5′-CCAGCCTTCGACAACCTCTATT-3′ and reverse,
5′-TGCCGTAGGTGTCCCTTTG-3′; miR-133a-5p forward,
5′-CTTTAACCATTCTAGCTTTTCCAGGTA−3′ and reverse,
5′-GACTTCGGCTGTGGACAAGATTAG−3′; U6 forward,
5′-CGCTTCGGCAGCACATATACTA-3′ and reverse,
5′-CGCTTCACGAATTTGCGTGTCA-3′; and GAPDH forward,
5′-AGGTCGGTGTGAACGGATTTG−3′ and reverse,
5′-GGGGTCGTTGATGGCAACA-3′.
Wound healing assay
The present study measured the cell migration
ability using a wound healing assay. After seeding into 24-well
plates at the density of 3×105 cells per well, H1299 and
HCC827 cells were transfected with different nucleotide fragments
as aforementioned. After cell reached 80% confluence, scratches
were created across the monolayer using a 10-µl pipette tip,
followed by washing with cold PBS. An image of each scratch was
captured under a light microscope. Subsequently, cells were
incubated in RPMI-1640 medium containing 1% FBS (26). After culturing for 24 h at 37°C,
images of the scratches were captured again. The migration distance
of each group was measured using ImageJ software, and analyzed with
the following formula: (W 0 h-W 24 h)/W 0 h ×100%, where W is the
wound.
Cell invasion assay
A Transwell assay was performed to detect the cell
invasion ability following the overexpression of miR-133a-3p or
ERBB2. The Transwell inserts were purchased from Corning, Inc.
Prior to the experiment, the upper chambers were covered with 100
µl of a 1:1 mixture of Matrigel and RPMI-1640 medium and incubated
at 37°C for 1 h. NSCLC cells were trypsinized, resuspended in
serum-free RPMI-1640 medium, and then seeded into the upper
chambers (2×104 cells/well), while medium containing 20%
FBS was placed in the lower chambers. After culturing at 37°C for
24 h, non-invaded cells in the upper chamber were wiped off using
sterile cotton swabs. The invaded cells were fixed using 4%
paraformaldehyde at room temperature for 10 min and stained with
0.1% crystal violet solution for 20 min at room temperature. The
invaded cells were imaged using an light microscope (magnification,
×400).
Dual-luciferase reporter gene
assay
Following bioinformatics analysis of the association
between miR-133a-3p and the 3′UTR of the ERBB2 transcript
(http://www.targetscan.org/vert_72/),
the dual-luciferase reporter gene assay was employed using
TargetScanHuman 7.2 to verify this prediction. The 3′UTR clones of
the ERBB2 transcript, including mutant (MUT) 3′UTRs and wild type
(WT) 3′UTRs, were designed and synthesized by Sangon Biotech Co.,
Ltd. The nucleotide fragments were ligated into the luciferase
plasmid (Promega Corporation). H1299 cells were seeded into 24-well
plates at the density of 1×105/well, and co-transfected
with 300 ng/well plasmid for pmir/ERBB2-WT or pmir/ERBB2-MUT and 50
ng/well miR-133a-3p mimics or miR-negative control (NC) and 100 ng
Renilla luciferase plasmid (pRL-TK; Promega Corporation) using
Lipofectamine 3000 (Invitrogen; Thermo Fisher Scientific, Inc.),
and H1299 cells transfected with pRL-TK alone were regarded as the
negative control, followed by incubation for 48 h at 37°C. Finally,
cells were lysed and the luciferase activities in different groups
were detected using a Dual-Luciferase Reporter Assay System
(Promega Corporation) following the manufacturer's guidance.
Firefly luciferase activities were normalized to Renilla
luciferase activities.
Immunofluorescence assay
Immunofluorescence assays were carried out to
evaluate E-cadherin and N-cadherin expression. A glass slide was
placed in a 6-well plate, and 2×105 cells/well were
seeded into the plate. After 24 h, the medium was removed and cells
were fixed with 4% paraformaldehyde at 4°C for 30 min. Then, cells
were blocked with 10% goat serum at room temperature (22±5°C) for
30 min. Subsequently, cells were incubated with anti-N-cadherin
(dilution, 1:1,000; 22018-1-AP; ProteinTech Group, Inc.) or
anti-E-cadherin (dilution, 1:1,000; 20874-1-AP; ProteinTech Group,
Inc.) antibodies overnight at 4°C. Afterwards, cells were incubated
with the Fluorescein-conjugated Affinipure Goat Anti-Rabbit IgG
(H+L) secondary antibody (dilution, 1:100; SA00003-2; ProteinTech
Group, Inc.) at room temperature for 1 h, followed by staining
using DAPI at room temperature for 5 min. Finally, images were
captured using a fluorescence microscope (magnification, ×400).
Statistical analysis
The data analysis in the present study was conducted
using SPSS 23.0 (IBM Corp.) and GraphPad 6.0 software (GraphPad
Software, Inc.). Student's t-test was carried out to compare
differences between two groups. Statistical differences among three
groups were analyzed using one-way ANOVA followed by Duncan's post
hoc test (LSR) at the 95% confidence interval (P<0.05 was
considered to indicate statistical significance). Statistical
differences among four groups were analyzed using one-way ANOVA
followed by Tukey's post hoc test. P<0.05 was considered to
indicate a statistically significant difference. Data were
expressed as mean ± standard deviation. Each assay was repeated
three times.
Results
Expression levels of miR-133a-3p and
ERBB2 are dysregulated in NSCLC tissues and cell lines
The present study assessed miR133a-3p expression at
the mRNA level and ERBB2 expression at the mRNA and protein levels
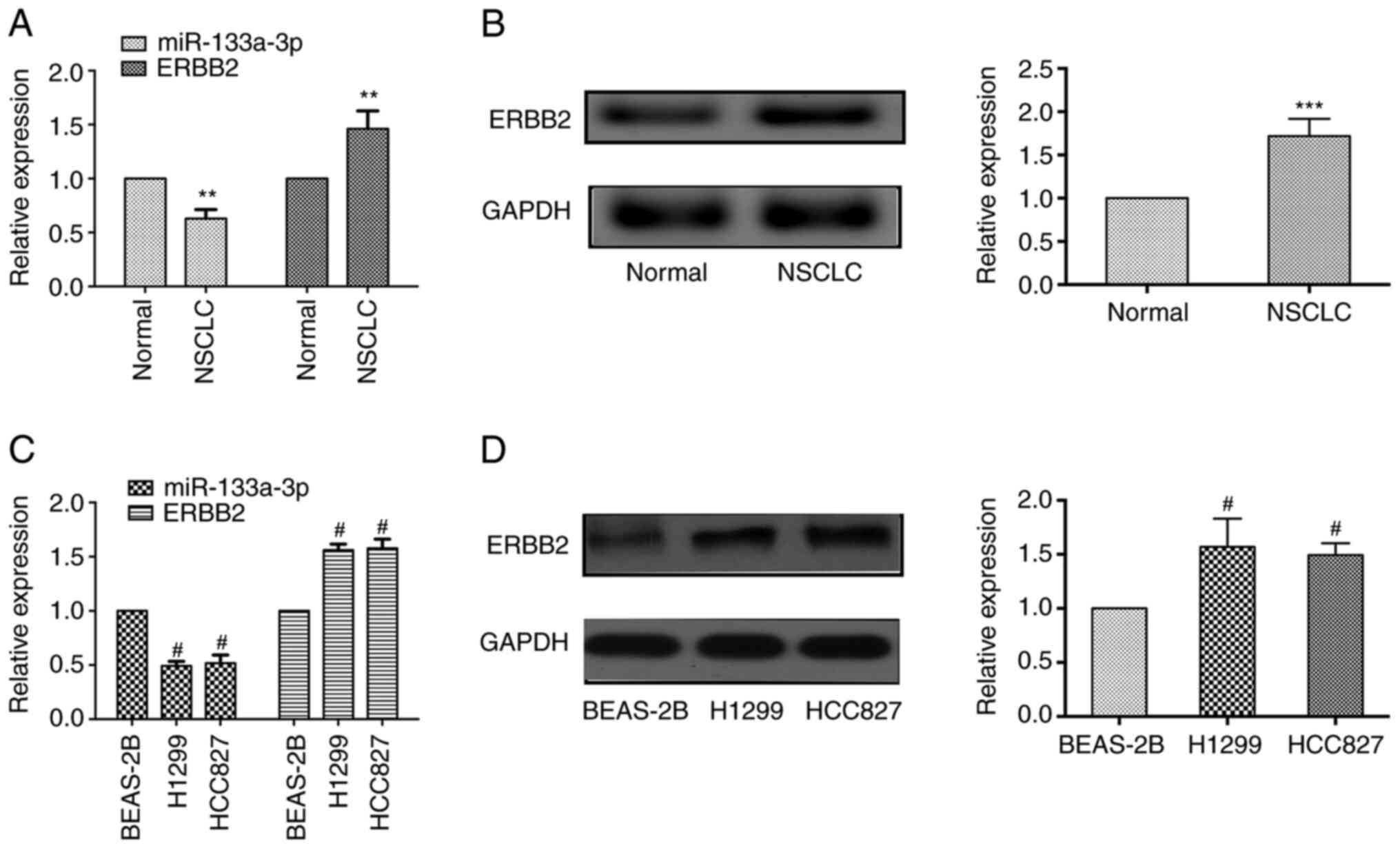
in NSCLC tissues and normal adjacent tissues. As shown in Fig. 1A, RT-qPCR results revealed that
miR-133a-3p expression was markedly downregulated in NSCLC tissues
compared with adjacent healthy tissues, while the mRNA expression
levels of ERBB2 were upregulated in cancer tissues. Western
blotting demonstrated that the protein expression levels of ERBB2
were elevated in NSCLC tissues (Fig.
1B).
In addition, the present study evaluated the
expression levels of these molecules in NSCLC cells. Consistent
with the results identified in tissue samples, lower miR-133a-3p
expression was observed in NSCLC cells (H1299 and HCC827 cell
lines), while ERBB2 expression was upregulated both at the mRNA and
protein levels compared with that in the normal pulmonary
epithelial cells (BEAS-2B; Fig. 1C and
D). These results indicated that miR-133a-3p expression was
decreased, while ERBB2 expression was increased in NSCLC tissues
and cells.
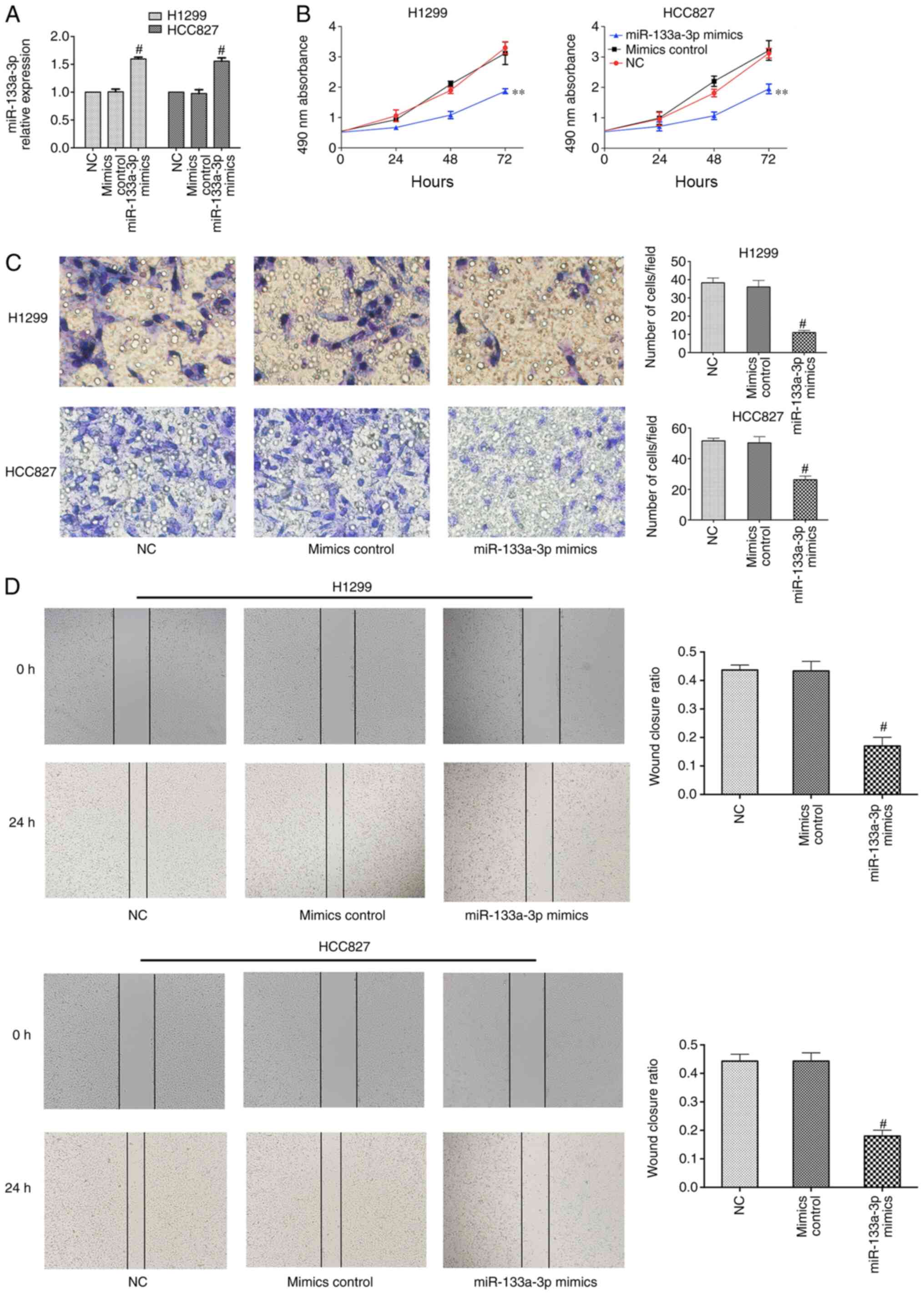
Overexpression of miR-133a-3p inhibits
NSCLC cell proliferation, migration and invasion
To determine the biological role of miR-133a-3p,
H1299 and HCC827 cells were transfected with control or miR-133a-3p
mimic. RT-qPCR was performed to evaluate transfection efficiency,
which demonstrated that miR-133a-3p expression was significantly
increased after transfection with miR-133a-3p mimic (Fig. 2A). An MTT assay was used to assess
the effect of miR-133a-3p on NSCLC cell proliferation. As shown in
Fig. 2B, overexpression of
miR-133a-3p led to a suppression of proliferation in NSCLC cells.
The Transwell assay indicated that overexpression of miR133a-3p
inhibited the invasion abilities of these cells (Fig. 2C). In addition, the wound healing
assay indicated the inhibitory effects of miR-133a-3p on migration
(Fig. 2D). Therefore, the present
results demonstrated that overexpression of miR-133a-3p inhibited
the proliferation, migration and invasion of NSCLC cells.
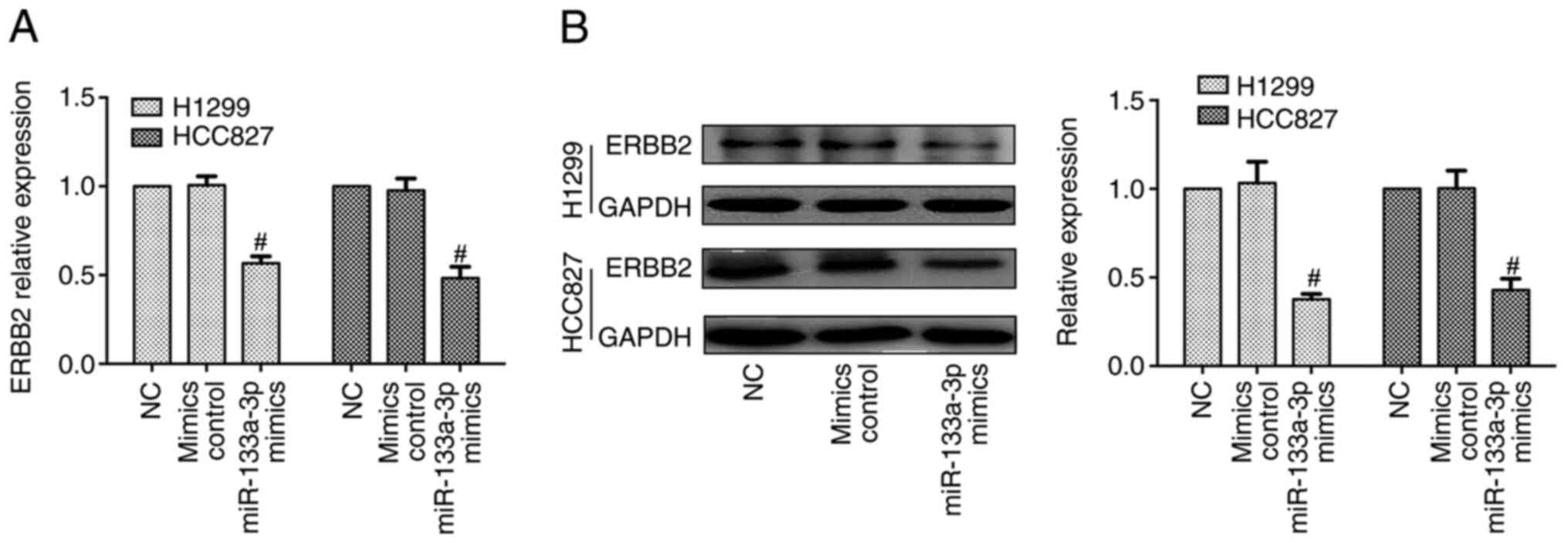
Furthermore, RT-qPCR and western blotting data indicated that the
mRNA and protein expression levels of ERBB2 were decreased in the
miR-133a-3p mimics group, compared with mimics control groups
(Fig. 3).
Overall, upregulation of ERBB2 and decreased
expression levels of miR-133a-3p were observed in NSCLC tissues and
cell lines. Downregulation of ERBB2 expression was observed in
cells transfected with miR-133a-3p mimic, indicating that ERBB2 may
be an indirect or direct target of miR-133a-3p.
miR-133a-3p directly targets the 3′UTR
of the ERBB2 transcript in NSCLC cells
As aforementioned, a negative association between
ERBB2 and miR-133-3p expression was noted. To further analyze this
interaction, bioinformatics analysis was performed using TargetScan
(Fig. 4A). This suggested that the
3′UTR of the ERBB2 transcript is a potential target of miR-133a-3p.
A dual-luciferase reporter assay was performed to verify this
prediction. As shown in Fig. 4B,
miR-133a-3p overexpression markedly suppressed the luciferase
activity in cells transfected with pmir/ERBB2-WT, while the
luciferase activity was not changed in cells transfected with
pmir/ERBB2-MUT. Therefore, these data provide evidence that
miR-133a-3p directly targets the 3′UTR of the ERBB2 transcript in
NSCLC cells.
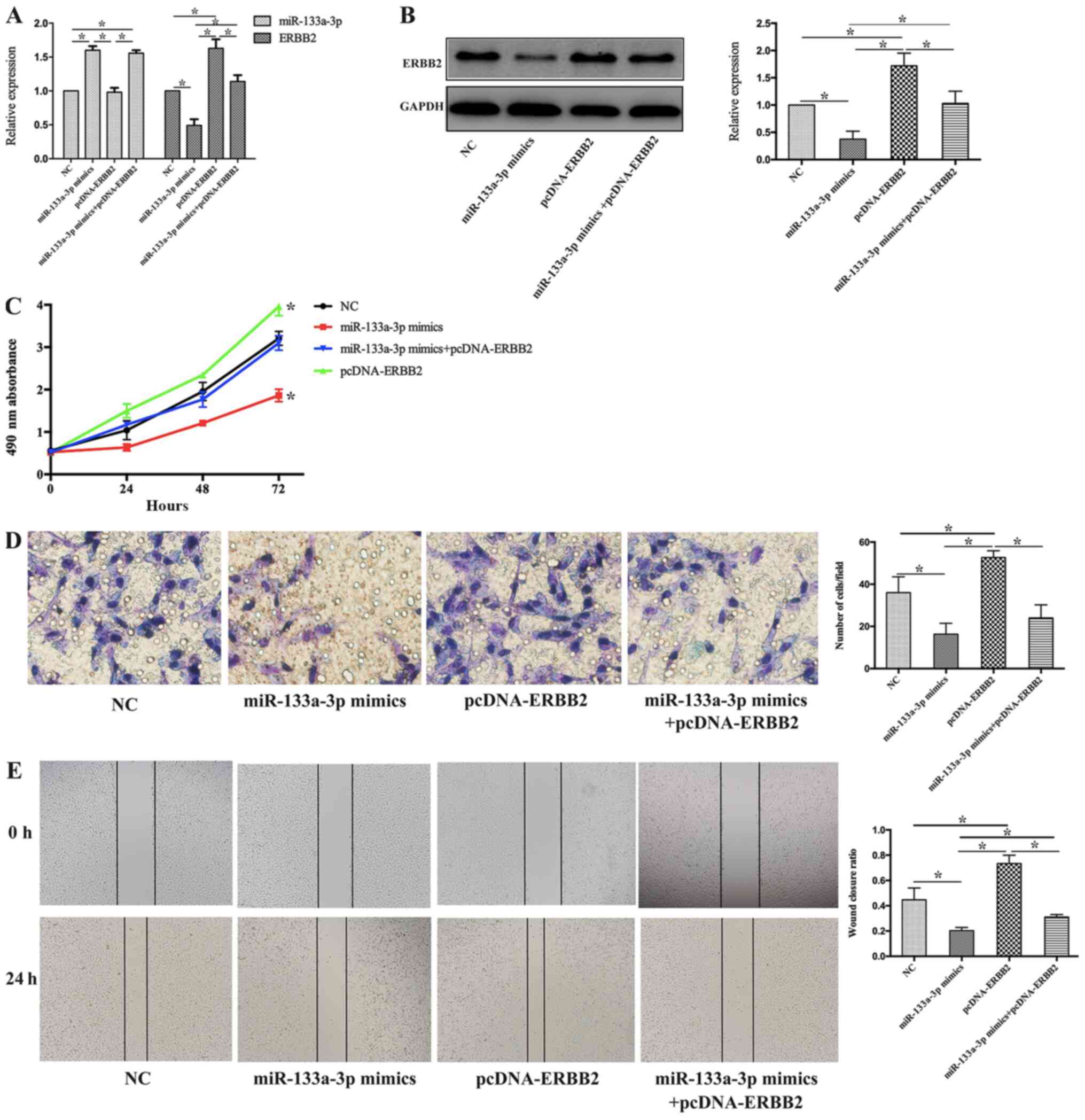
miR-133a-3p inhibits the
proliferation, invasion and migration of NSCLC cells by negatively
regulating ERBB2 expression
To further explore the biological functions of the
miR-133a-3p/ERBB2 axis in NSCLC progression, H1299 cells were
treated with miR-133a-3p mimic, pcDNA-ERBB2 or miR-133a-3p mimic +
pcDNA-ERBB2. pcDNA-NC and pcDNA-ERBB2 were transfected into H1299
cells, and pcDNA-NC did not affect the expression levels of ERBB2
in H1299 cells (Fig. S1).
RT-qPCR and western blotting were performed to
detect miR-133a-3p and ERBB2 expression in each treatment group. As
shown in Fig. 5A, miR-133a-3p
expression was increased in both the miR-133a-3p mimic and
miR-133a-3p mimic + pcDNA-ERBB2 groups but was not significantly
altered in the pcDNA-ERBB2 group compared with the NC group.
Furthermore, ERBB2 expression was increased in the pcDNA-ERBB2
group, while it was decreased in the miR-133a-3p mimics group,
compared with non-treated (NC) group. Compared with the pcDNA-ERBB2
group, ERBB2 expression was suppressed in the miR-133a-3p mimic +
pcDNA-ERBB2 group (Fig. 5B).
MTT, Transwell and wound healing assays were
performed to demonstrate the potential role of the
miR-133a-3p/ERBB2 axis in NSCLC cell phenotypes. As shown in
Fig. 5C, the MTT data demonstrated
that overexpression of miR-133a-3p and ERBB2 in combination
preserved the proliferation activity of H1299 cells, which was
suppressed in miR-133a-3p mimics group. Compared with pcDNA-ERBB2
group, the proliferation of H1299 cells in miR-133a-3p
mimics+pcDNA-ERBB2 group decreased. In addition, the invasive
ability was attenuated in H1299 cells transfected with both
miR-133a-3p mimics and pcDNA-ERBB2 (miR-133a-3p mimics+pcDNA-ERBB2
group), compared with pcDNA-ERBB2 group (Fig. 5D). Finally, the wound healing assay
suggested that the co-transfection of miR-133a-3p mimic and
pcDNA-ERBB2 decreased the wound closure ratio in H1299 cells
compared with that in cells in the pcDNA-ERBB2 group (Fig. 5E). These results suggest that
miR-133a-3p inhibits cell proliferation, invasion and migration of
NSCLC cells by negatively regulating ERBB2 expression.
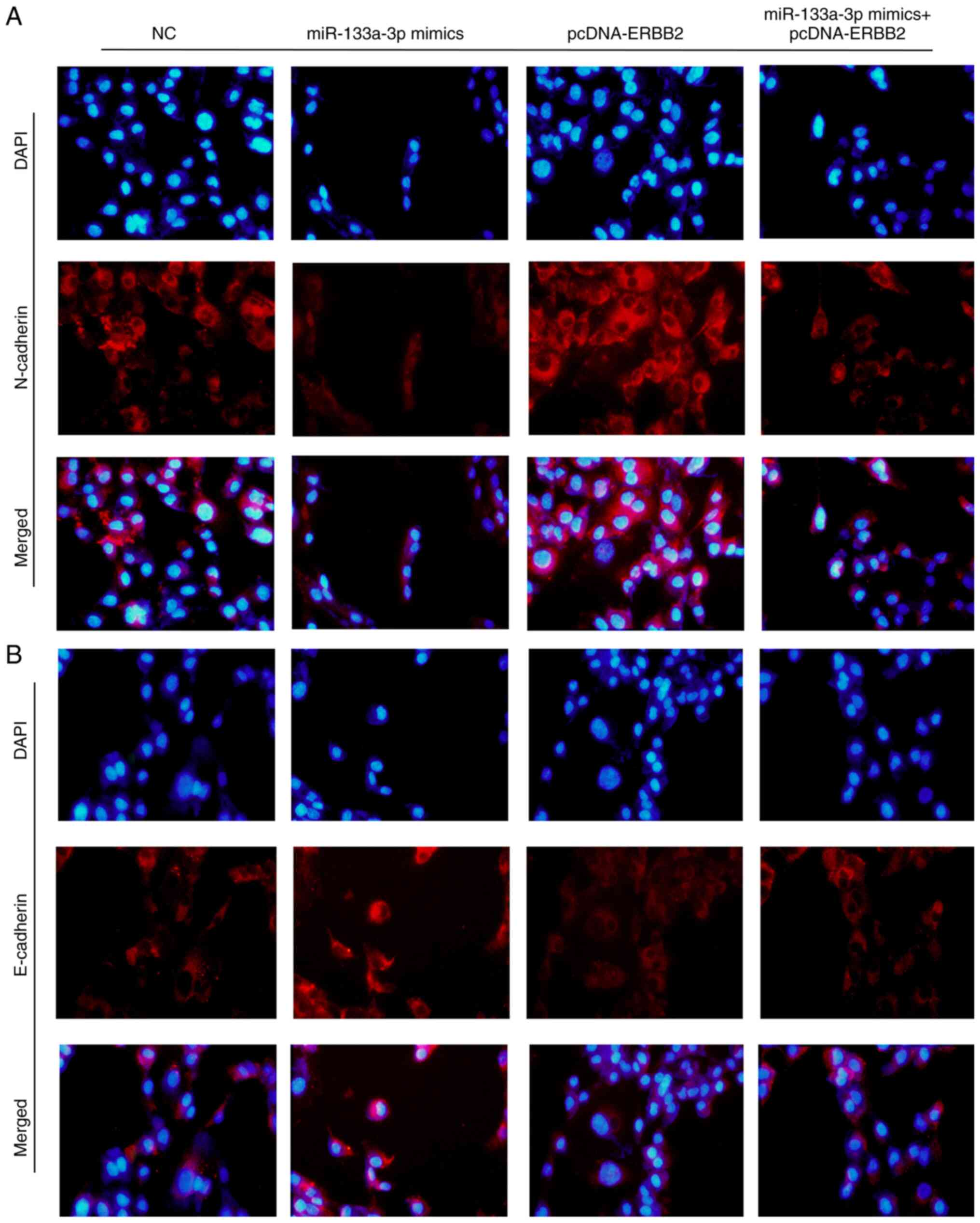
Overexpression of miR-133a-3p
regulates the expression levels of epithelial-mesenchymal
transition (EMT) biomarkers
E-cadherin and N-cadherin are two important
biomarkers of the EMT process. The present study detected
E-cadherin and N-cadherin expression using an immunofluorescence
assay following transfection with miR-133a-3p mimics, pcDNA-ERBB2
or miR-133a-3p mimics + pcDNA-ERBB2. As shown in Fig. 6A, N-cadherin expression was decreased
in the miR-133a-3p mimics group but increased in the pcDNA-ERBB2
group. Consistently, E-cadherin exhibited the opposite trends
(Fig. 6B). These results indicate
that the overexpression of miR-133a-3p suppresses the EMT process
in NSCLC cells.
Discussion
At present, surgery and chemotherapy are the
mainstream treatment modalities for NSCLC (27). However, the outcome of these
treatments is unsatisfactory, since postoperative recurrence and
therapy resistance are common in patients with NSCLC (28). With the development of molecular
biology, targeted therapy has become available for malignant tumors
(29). However, its efficacy remains
limited, since most patients eventually develop drug resistance.
Therefore, there is still an urgent need for the identification of
novel therapeutic targets. In the last decade, miRNAs have become a
hot topic in cancer diagnosis and treatment (18). miRNAs are a type of short, non-coding
RNA molecules that serve critical regulatory roles in multiple
cellular events. Increasing evidence has suggested that miRNA
dysregulation is closely associated with the pathogenesis and
progression of a variety of diseases, in particular cancer types
(30), for example miR-133a-3p
expression is downregulated in gastric cancer (21). With the increased understanding of
miRNAs, the feasibility of using miRNAs as therapeutic targets is
supported (31,32).
Previous studies have suggested a tumor-suppressive
role of miR-133a-3p in multiple malignant tumors, including
gastric, breast and bladder cancer (21,22,33). Low
expression levels of miR-133a-3p are associated with poor prognosis
and faster progression (22). Shi
et al (33) reported that
downregulation of miR-133a-3p promotes the proliferation and
invasion of breast cancer cells. Li et al (34) suggested that miR-133a-3p promotes
autophagy in gastric cancer cells by negatively regulating forehead
box P3. The present study identified decreased miR-133a-3p
expression in NSCLC cancer tissues and cell lines. A series of
cellular and molecular experiments were performed to identify its
biological function in NSCLC pathogenesis. The present results
indicated that overexpression of miR-133a-3p effectively suppressed
the cell proliferation and invasion of NSCLC cells.
To further assess the molecular mechanisms
underlying the tumor-suppressive effect of miR-133a-3p,
bioinformatics analysis was performed using TargetScan 7.2. Among
the potential target genes, the present study focused on ERBB2, a
well-known oncogene in NSCLC (35).
ERBB2 is a member of the EGFR family, which is involved in a series
of biological events in malignant diseases (36). It has been suggested that ERBB2 could
trigger several cellular processes, including cell proliferation,
survival and differentiation (8,37).
Spencer et al (38) provided
evidence that the ERBB2 receptor assists the binding of p130Cas to
CRK, and thus, promotes the invasion and migration of breast cancer
cells. Furthermore, ERBB2 expression is upregulated in various
malignancies, and ERBB2-targeted therapy has become an important
component of therapeutic strategies in a number of cancer types
(12). In the present study, the
luciferase assay supported the notion that miR-133a-3p could target
the 3′UTR of ERBB2. Furthermore, a pcDNA-ERBB2 vector was employed
to increase ERBB2 expression, and this reversed the effects of the
miR-133a-3p mimic on cell proliferation, invasion and migration.
Therefore, the tumor-suppressive effect of miR-133a-3p may be
mediated, at least in part, by inhibition of ERBB2 expression.
EMT is an essential event in tumor carcinogenesis
and progression. As a member of the EGFR family, ERBB2 has been
demonstrated to promote EMT in previous study (39). The present study detected the typical
EMT biomarkers E-cadherin and N-cadherin and demonstrated that
overexpression of miR-133a-3p markedly suppressed the EMT process
in NSCLC cells, while overexpression of ERBB2 ameliorated this
effect. Therefore, the present findings suggest that miR-133a-3p
may suppress EMT by targeting ERBB2.
There were some limitations to the present study.
Commonly in wound healing assays non-FBS medium should be used;
however, 1% FBS was used in the present study to prevent cell
starving. Also, the present study only provided in vitro
data, therefore further in vivo experiments, such as
tumorigenesis assay, should be performed in future. Besides,
further analysis of whether miR-133a-3p can regulate other
signaling pathways impacting the phenotypes of NSCLC are
needed.
In conclusion, the present results indicate that
miR-133a-3p acts as a tumor-suppressive gene via inhibition of the
proliferation, invasion and EMT of NSCLC cells. In particular, the
inhibitory effects of miR-133a-3p on ERBB2 indicated that
miR-133a-3p may be a potent antitumor target for NSCLC
treatment.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
The majority of experiments and manuscript were
finished by YX. YX performed statistical analysis and interpreted
the data. LZ contribute to bioinformatics analysis and dual
luciferase reporter gene experiment. LX and XZ designed the
research, collected the tissues and edited the manuscript. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was conducted in accordance with
the Helsinki Declaration guidelines, and written consent to
participation in the present study were obtained from all patients
or their direct relatives. Furthermore, the present study was
approved by The Ethics Committee of Zhejiang Hospital (Hangzhou,
China; approval no. 20100103).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Duffy SW and Field JK: Mortality reduction
with low-dose CT screening for lung cancer. N Engl J Med.
382:572–573. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Qi M, Dai D, Liu J, Li Z, Liang P, Wang Y,
Cheng L, Zhan Y, An Z, Song Y, et al: AIM2 promotes the development
of non-small cell lung cancer by modulating mitochondrial dynamics.
Oncogene. 39:2707–2723. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhang H, Qian G, Zong D, Fan S, Owonikoko
TK, Ramalingam SS and Sun SY: Overcoming acquired resistance of
epidermal growth factor receptor-mutant non-small cell lung cancer
cells to osimertinib by combining osimertinib with the histone
deacetylase inhibitor panobinostat (LBH589). Cancer. 126:2024–2033.
2020. View Article : Google Scholar
|
|
5
|
Morgensztern D, Ng SH, Gao F and Govindan
R: Trends in stage distribution for patients with non-small cell
lung cancer: A national cancer database survey. J Thorac Oncol.
5:29–33. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Du L and Morgensztern D: Chemotherapy for
advanced-stage non-small cell lung cancer. Cancer J. 21:366–370.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhang Q, Zhou L, Guan Y, Cheng Y and Han
X: BENC-511, a novel PI3K inhibitor, suppresses metastasis of
non-small cell lung cancer cells by modulating β-catenin/ZEB1
regulatory loop. Chem Biol Interact. 294:18–27. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yang ZY, Yang L, Xu CW, Wang XJ and Lei L:
An insertion mutation of ERBB2 enhances breast cancer cell growth
and confers resistance to lapatinib through AKT signaling pathway.
Biol Open. 9:bio0476622020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Honkoop H, de Bakker DE, Aharonov A, Kruse
F, Shakked A, Nguyen PD, de Heus C, Garric L, Muraro MJ, Shoffner
A, et al: Single-cell analysis uncovers that metabolic
reprogramming by ErbB2 signaling is essential for cardiomyocyte
proliferation in the regenerating heart. Elife. 8:e501632019.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Albrecht T, Rausch M, Rössler S, Albrecht
M, Braun JD, Geissler V, Mehrabi A, Vogel MN, Pathil-Warth A,
Mechtersheimer G, et al: HER2 gene (ERBB2) amplification is a rare
event in non-liver-fluke associated cholangiocarcinogenesis. BMC
Cancer. 19:11912019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Menderes G, Bonazzoli E, Bellone S, Black
J, Altwerger G, Masserdotti A, Pettinella F, Zammataro L, Buza N,
Hui P, et al: SYD985, a novel duocarmycin-based HER2-targeting
antibody-drug conjugate, shows promising antitumor activity in
epithelial ovarian carcinoma with HER2/Neu expression. Gynecol
Oncol. 146:179–186. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ahmed S, Sami A and Xiang J: HER2-directed
therapy: Current treatment options for HER2-positive breast cancer.
Breast Cancer. 22:101–116. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li L, Jia L and Ding Y: Upregulation of
miR-375 inhibits human liver cancer cell growth by modulating cell
proliferation and apoptosis via targeting ErbB2. Oncol Lett.
16:3319–3326. 2018.PubMed/NCBI
|
|
14
|
Rimawi MF, Schiff R and Osborne CK:
Targeting HER2 for the treatment of breast cancer. Annu Rev Med.
66:111–128. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xiang Z, Huang X, Wang J, Zhang J, Ji J,
Yan R, Zhu Z, Cai W and Yu Y: Cross-database analysis reveals
sensitive biomarkers for combined therapy for ERBB2+
gastric cancer. Front Pharmacol. 9:8612018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wu J, Li S, Ma R, Sharma A, Bai S, Dun B,
Cao H, Jing C, She J and Feng J: Tumor profiling of co-regulated
receptor tyrosine kinase and chemoresistant genes reveal different
targeting options for lung and gastroesophageal cancers. Am J
Transl Res. 8:5729–5740. 2016.PubMed/NCBI
|
|
17
|
Liang CH, Shiu LY, Chang LC, Sheu HM and
Kuo KW: Solamargine upregulation of Fas, downregulation of HER2,
and enhancement of cytotoxicity using epirubicin in NSCLC cells.
Mol Nutr Food Res. 51:999–1005. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lu TX and Rothenberg ME: MicroRNA. J
Allergy Clin Immunol. 141:1202–1207. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Olejniczak M, Kotowska-Zimmer A and
Krzyzosiak W: Stress-induced changes in miRNA biogenesis and
functioning. Cell Mol Life Sci. 75:177–191. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ohanian M, Humphreys DT, Anderson E,
Preiss T and Fatkin D: A heterozygous variant in the human cardiac
miR-133 gene, MIR133A2, alters miRNA duplex processing and strand
abundance. BMC Genet. 14:182013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang X, Li Z, Xuan Z, Xu P, Wang W, Chen
Z, Wang S, Sun G, Xu J and Xu Z: Novel role of miR-133a-3p in
repressing gastric cancer growth and metastasis via blocking
autophagy-mediated glutaminolysis. J Exp Clin Cancer Res.
37:3202018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gao L, Li SH, Tian YX, Zhu QQ, Chen G,
Pang YY and Hu XH: Role of downregulated miR-133a-3p expression in
bladder cancer: A bioinformatics study. Onco Targets Ther.
10:3667–3683. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
He B, Lin X, Tian F, Yu W and Qiao B:
miR-133a-3p inhibits oral squamous cell carcinoma (OSCC)
proliferation and invasion by suppressing COL1A1. J Cell Biochem.
119:338–346. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yang ZQ, Wu CA and Cheng YX: Prognostic
value of microRNA-133a expression and its clinicopathologic
significance in non-small cell lung cancer: A comprehensive study
based on meta-analysis and the TCGA database. Oncol Res Treat.
41:762–768. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tian L, Chen M, He Q, Yan Q and Zhai C:
MicroRNA-199a-5p suppresses cell proliferation, migration and
invasion by targeting ITGA3 in colorectal cancer. Mol Med Rep.
22:2307–2317. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gamerith G, Kocher F, Rudzki J and Pircher
A: ASCO 2018 NSCLC highlights-combination therapy is key. Memo.
11:266–271. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tian Y, Sun C, Zhang L and Pan Y: Clinical
significance of miRNA-106a in non-small cell lung cancer patients
who received cisplatin combined with gemcitabine chemotherapy.
Cancer Biol Med. 15:157–164. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hu Y, Liu C and Muyldermans S:
Nanobody-based delivery systems for diagnosis and targeted tumor
therapy. Front Immunol. 8:14422017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
McGuire A, Brown JA and Kerin MJ:
Metastatic breast cancer: The potential of miRNA for diagnosis and
treatment monitoring. Cancer Metastasis Rev. 34:145–155. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shin VY and Chu KM: miRNA as potential
biomarkers and therapeutic targets for gastric cancer. World J
Gastroenterol. 20:10432–10439. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Takahashi RU, Prieto-Vila M, Kohama I and
Ochiya T: Development of miRNA-based therapeutic approaches for
cancer patients. Cancer Sci. 110:1140–1147. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Shi W, Tang T, Li X, Deng S, Li R, Wang Y,
Wang Y, Xia T, Zhang Y, Zen K, et al: Methylation-mediated
silencing of miR-133a-3p promotes breast cancer cell migration and
stemness via miR-133a-3p/MAML1/DNMT3A positive feedback loop. J Exp
Clin Cancer Res. 38:4292019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li JP, Zhang HM, Liu MJ, Xiang Y, Li H,
Huang F, Li HH, Dai ZT, Gu CJ, Liao XH and Zhang TC:
miR-133a-3p/FOXP3 axis regulates cell proliferation and autophagy
in gastric cancer. J Cell Biochem. 121:3392–3405. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Byeon S, Lee B, Park WY, Choi YL, Jung HA,
Sun JM, Ahn JS, Ahn MJ, Park K and Lee SH: Benefit of targeted DNA
sequencing in advanced non-small-cell lung cancer patients without
EGFR and ALK alterations on conventional tests. Clin Lung Cancer.
21:e182–e190. 2020. View Article : Google Scholar
|
|
36
|
Liberelle M, Jonckheere N, Melnyk P, Van
Seuningen I and Lebègue N: EGF-containing membrane-bound mucins: A
hidden ErbB2 targeting pathway? J Med Chem. 63:5074–5088. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Voelker R: Another targeted therapy for
ERBB2-positive breast cancer. JAMA. 323:4082020. View Article : Google Scholar
|
|
38
|
Spencer KS, Graus-Porta D, Leng J, Hynes
NE and Klemke RL: ErbB2 is necessary for induction of carcinoma
cell invasion by ErbB family receptor tyrosine kinases. J Cell
Biol. 148:385–397. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jung DH, Bae YJ, Kim JH, Shin YK and Jeung
HC: HER2 regulates cancer stem cell activities via the Wnt
signaling pathway in gastric cancer cells. Oncology. 97:311–318.
2019. View Article : Google Scholar : PubMed/NCBI
|