Introduction
Cancer is a leading cause of mortality and
morbidity, with 9.6 million cancer-related deaths in 2018 worldwide
(1), and approximately 18.1 million
newly diagnosed cancer cases (2).
Patients with cancer currently have multiple treatment options
available, including surgery, cytotoxic-chemotherapy, radiotherapy
and immunotherapy (3,4), as well as cryoablation, which is
considered a treatment option for certain types of cancer (5). It is most commonly used to treat liver,
kidney, lung, prostate and breast cancer (6). This minimally invasive percutaneous
procedure is emerging as an alternative to surgery in patients with
early-stage breast cancer (7).
Although cryoablation has yet to be established as a
standard-of-care procedure for breast cancer management, it should
be considered an appropriate therapy during periods where there are
changes in standard procedures due to shifts in healthcare policies
and practices (8,9). It was argued that this non-operative,
resource-saving strategy was pragmatic and appropriate for managing
localized breast cancer during the height of the 2020 COVID-19
pandemic (9).
Cryoablation is a percutaneous ablation technique
that targets neoplastic tissue destruction through freeze/thaw
cycles at low temperatures (5,10). The
process involves inserting a cryoprobe into a tumor mass guided by
imaging devices (6,11,12).
Once the lesion is targeted, the cryoprobe is cooled by passing
liquified gas through the probe, which expands into a gaseous state
at the tip to create low temperatures <-40°C (5,13). In
clinical cryoablation, temperatures below −40°C are maintained to
eradicate all cancer cells (12).
Multiple freeze-thaw cycles are performed to obtain an effective
ablation (13). This approach for
nodule destruction does not allow time for defensive mutations to
occur in cancer cells (14). The
delivery of cryotherapy, combined with the anatomy of the breast
presents technical challenges for its clinical application
(10). Achieving critically low
temperatures throughout the entire tumor mass is difficult
(12). The thermal gradient that
spreads out from the inserted freezing probe may not fully and
completely penetrate the entire mass of the tumor with the desired
freezing temperature (12). In
addition, proximity to blood vessels can interfere with the
freezing process (15). Hence, the
destruction of tumor cells may not be achieved at the freeze
margin. Incomplete ablation of cancerous tissue results in
therapeutic failure (12). The
sensitization of cells to freezing has been suggested as a way to
overcome the problem of under-freezing at the freeze margin
(16,17). Previous studies have revealed the
synergistic effect of administering cryosensitizing agents, such as
5-fluorouracil, vitamin D3 and cisplatin prior to cryotherapy to
improve ablation efficiency (18–20). The
aim of cryosensitization as an adjunct to cryoablation, is to
increase the ablated area achieved by each freeze-thaw cycle
(21). Cellular injury resulting
from both freezing and thawing is associated not merely with simple
freeze rupture but also with molecular-based cell death processes
(apoptosis, autophagy and necrosis) as well as immune responses to
cell damage (17). Therefore,
ablation could be augmented with the use adjuvants, such as
pro-apoptotic, pro-inflammatory and antiproliferative chemicals
(17). The use of cryosensitizing
agents and the ensuing cellular damage results in biochemical
events associated with cell death (13,17).
There is a clinical need to enhance the efficacy of
cryotherapy using adjunctive cryosensitizing agents and the
candidate agents are diverse (12).
Cellular damage resulting from freezing involves various processes
that could be considered targets for cryosensitizing agents
(17). These processes include both
structural stress events, such as rupture of membranes and
cytoskeletal disassembly and chemical stress events, such as
metabolic uncoupling, ATP depletion, ionic imbalances, cellular
acidosis and free radical generation (17). When freezing is initiated, ice
formation creates a hyperosmolar extracellular environment, which
draws water out of the cell and in turn causes cellular dehydration
(7,11,17).
This exposes cells to potentially lethal osmotic pressures
(14,15). Central to this cellular passage of
water are the aquaporin (AQP) integral proteins (22). These transmembrane channels allow
water to flow through cell membranes in response to osmotic
gradients in cells (23). The
presence of AQPs increases plasma membrane permeability to water by
5–50 times compared with that of the plasma membrane alone
(24). Freezing of cancer cells
in vivo has been demonstrated to induce an increase in the
expression of AQP3 and this has been suggested to be an underlying
mechanism for overcoming osmotic stress created by the formation of
ice crystals outside the cell and the subsequent movement of water
from the interior to the exterior of the cell (22). In relation to cold temperature
stress, AQP proteins may be considered cryoprotective (25–28).
Hence, AQP proteins are potential pharmacological targets for
enhancing the efficiency of cryotherapy (22).
AQP proteins are localized to the cytosol of cancer
cells in vivo (29–31). However, these proteins must be
precisely positioned in the plasma membrane in order to function as
transporters (22). In prostate
cancer cells, AQP3 has been found to translocate from the cytosol
to the plasma membrane in response to cryoinjury (22). This cellular localization process
involves a dynamic sequential cascade of events from transcription
to translation and post-translational modifications followed by
recruitment as vesicular cargo transported to appropriate plasma
membrane domains and finally ending in precise docking and fusion
with the cell membranes (29–31).
These complex events are induced as part of the adaptation
mechanisms to cryoinjury (32).
In vitro interference with any of these cellular events may
alter AQP function and, in turn, alter cellular adaptation to
freezing temperatures (22).
Blockade of AQP activity may therefore be a cryosensitizing
process, i.e., inhibition of AQPs may enhance the damage caused by
freezing (17). An increase in AQP3
activity is cryoprotective, while inhibition of AQP3 has been
demonstrated to increase cryodamage (22,26,33).
However, to the best of our knowledge the role of AQP activity in
human breast cancer cells has not been reported in relation to
cryoinjury.
Breast cancer cells express AQP1, AQP3 and AQP5, and
it has been reported that this expression is associated with
severity of histological tumors and patient prognosis (34). It has been demonstrated that AQP3 has
a key role in the migration of breast cancer cells (35), and it has been further suggested that
the level of increased expression of AQP3 and AQP5 may serve as
biomarkers of cancer severity (36).
AQP1 expression is significantly associated with poor clinical
prognosis amongst patients with early breast cancer (37,38).
Disease severity is related to various subtypes of breast cancer,
which are classified based upon defined features (39–41).
Triple-negative breast cancer (TNBC) is characterized by the
absence of estrogen receptors (ER) and progesterone receptors (PR)
and a lack of excess human epidermal growth factor receptor-2
(HER2). Clinically TNBC is associated with poorer prognosis
compared with non-TNBC types of breast cancer (42–45).
TNBC is also associated with the upregulation of AQP3 and AQP5
(36). MDA-MB-231 is a TNBC cell
line that is used as an in vitro model of TNBC (46,47). In
contrast, MCF-7 cells express ER, PR and low levels of HER2 and are
used as an in vitro model of hormone-responsive breast
cancer types (48). The utilization
of these 2 cell lines in experiments allows for the comparison of
both breast cancer types (49–52).
It was hypothesized that the inhibition of AQPs may
function as a possible adjuvant process to cryotherapy that may
enhance cryoablation. The current study aimed to investigate
changes in AQP gene expression and cellular localization of AQPs in
response to cryoinjury. This was performed using 2 breast cancer
cell lines (MDA-MB-231 and MCF-7). In addition, the current study
investigated the synergistic antitumor effect of cryoinjury in
conjunction with aquaporin blockade on breast cancer cells. The
findings of the present study reported a synergy that may bring
about a cryosensitization which may be used as an adjunct to
cryotherapy. This has particular therapeutic importance in the
border area targeted by cryoablation where freezing temperatures
are not cold enough to induce cellular damage.
Materials and methods
Cell culture
Human breast cancer cell lines MDA-MB-231 and MCF-7
were obtained from the American Type Culture Collection. Cells were
cultured in advanced Dulbecco's modified Eagle's medium (Advanced
DMEM; cat. no. 12491015; Thermo Fisher Scientific, Inc.)
supplemented with 2.5% fetal bovine serum (FBS; cat. no. F2442;
Sigma-Aldrich; Merck KGaA), 1% 10,000 U/ml penicillin G sodium
salt/10 mg/ml streptomycin sulphate (cat. no. 15070063; Thermo
Fisher Scientific, Inc.) and 1% 200 mM L-glutamine (cat. no. G7513;
Sigma-Aldrich; Merck KGaA) and maintained at 37°C in a humidified
atmosphere containing 5% CO2.
Cryoinjury
MDA-MB-231 and MCF-7 cells were placed at −13, 0 or
37°C for 10 min. The lowest temperature that could be used to
accomplish freezing while maintaining cell viability was −13°C. In
a previous study, a temperature of −10°C resulted in a high cell
survival rate, whereas −15°C led to >80% cell death (53). In addition, 0°C degree was used as
the control temperature for non-freezing conditions. Freezing for
10 min was the time necessary for the media to be frozen while
preserving the integrity of the cells (53). A temperature of 0°C was achieved by
placing the cells on a MyBlock™ Mini dry bath with cooling
(Benchmark Scientific, Inc.) and −13°C temperature was achieved by
placing the cells in a Sanyo temperature calibrated freezer (Sanyo
Electrical Co. Ltd.), while the control temperature was achieved by
maintaining the cells in a humidified 5% CO2 atmosphere
at 37°C. After cold exposure of −13°C and 0°C temperatures, cells
were placed back in a humidified 5% CO2 atmosphere at
37°C for either 2, 6 or 24 h.
Cell treatment and collection for RNA
and protein analysis
Expression levels of AQP1, AQP3 and AQP5 were
assessed by seeding 1×106 cells in 60-mm tissue culture
dishes. After 24 h, cells were exposed to either-13, 0 or 37°C for
10 min and placed back at 37°C in a humidified atmosphere
containing 5% CO2. Cells were collected after 2, 6 and
24 h incubation. Cells were scraped and pelleted by centrifugation
at 238 × g for 5 min at 4°C and then washed twice with
phosphate-buffered saline (PBS; Sigma-Aldrich; Merck KGaA). The
pellet was subsequently used for RNA or protein extraction.
RNA extraction and cDNA synthesis
Total RNA was isolated using the RNeasy Mini kit
(Qiagen GmbH) according to the manufacturer's instructions. RNA
concentration and purity were measured spectrophotometrically at
260 and 280 nm using a NanoDrop™ 2000 (Thermo Fisher Scientific,
Inc.). cDNA was synthesized using the High-Capacity cDNA Reverse
Transcription kit (Applied Biosystems; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocols. Each reaction
consisted of 1 µg total RNA, 2 µl RT buffer, 2 µl random primer,
0.8 µl dNTP, 1 µl MultiScribe™ reverse transcriptase and 20 µl QSP
of nuclease-free water. Reverse transcription conditions were as
follows: 25°C for 10 min, 37°C for 120 min and then 85°C for 5
min.
Reverse transcription-quantitative
(RT-q) PCR
RT-qPCR was performed to determine the mRNA
expression levels of AQP1, AQP3 and AQP5, which were normalized to
GAPDH expression. Primers were synthesized by Macrogen, Inc., as
shown in Table I. A reaction volume
of 20 µl, which included 1 µl cDNA, 25 nmol forward and reverse
primers, 7 µl nuclease-free water and 10 µl SYBR™ Green PCR Master
mix (Applied Biosystems; Thermo Fisher Scientific, Inc.) was used.
Each reaction was run in duplicate on the Applied Biosystem 7500
system (Applied Biosystems; Thermo Fisher Scientific, Inc.) in
MicroAmp™ Optical 96-well reaction plates (Applied Biosystems;
Thermo Fisher Scientific, Inc.). Samples were initially denatured
for 10 min at 95°C, each cycle was 15 sec at 95°C, followed by
annealing and elongation for 1 min at 60°C for 50 cycles. Relative
gene expression was represented by fold-change relative to GAPDH.
This fold-change was calculated based on the threshold cycle (Ct)
using the 2−ΔΔCq method (54). Fold-change was defined as the
normalized gene expression (2−ΔCq) in the test sample/by
the normalized gene expression (2−ΔCq) in the control
sample (vehicle).
 | Table I.Forward and reverse primer sequences
used for RT-qPCR. |
Table I.
Forward and reverse primer sequences
used for RT-qPCR.
| Gene | Forward Primer
(5′-3′) | Reverse Primer
(5′-3′) |
|---|
| AQP1 |
TATGCGTGCTGGCTACTACCGA |
GGTTAATCCCACAGCCAGTGTAG |
| AQP3 |
CCGTGACCTTTGCCATGTGCTT |
TTGTCGGCGAAGTGCCAGATTG |
| AQP5 |
TACGGTGTGGCACCGCTCAATG |
AGTCAGTGGAGGCGAAGATGCA |
| GAPDH |
GTCTCCTCTGACTTCAACAGCG |
ACCACCCTGTTGCTGTAGCCAA |
Western blotting
Cell pellets were lysed in a protein extraction
buffer composed of 150 mM NaCl, 50 mM Tris HCl, 1 mM
ethylenediaminetetraacetic acid, 10% glycerol, 1% Triton-X-100,
0.5% sodium deoxycholate and 0.1% sodium dodecyl sulphate (SDS)
supplemented with 1X protease inhibitor cocktail (Sigma-Aldrich;
Merck KGaA), 2 mM phenylmethanesulfonylfluoride and 0.01 M sodium
fluoride. Cells were sonicated for 10 sec every 30 sec 3 times for
2 cycles. Following sonication, cell lysates were centrifuged at
13,362 × g for 10 min at 4°C and then the supernatant was
collected. Total proteins were quantified using a bicinchoninic
acid protein assay (Sigma-Aldrich; Merck KGaA) following the
manufacturer's instructions. For western blotting, 300 µg
MDA-MB-231 and 400 µg MCF-7 proteins were resolved on 12% SDS-PAGE
gels and subsequently transferred to polyvinylidene difluoride
membranes. Non-specific binding proteins were blocked in 1% bovine
serum albumin (BSA) (Cell Signaling Technology, Inc.) for 1 h at
room temperature. After blocking, membranes were incubated with
polyclonal rabbit antibodies against AQP1 (cat. no. bs-1506R), AQP3
(cat. no. bs-1253R) and AQP5 (cat. no. bs-1554R) (all 1:1,000; all
BIOSS) in 1% BSA, and monoclonal mouse antibody against β-actin
(1:1,000; cat. no. bsm-51011M; BIOSS) with shaking overnight at
4°C. Subsequently, membranes were washed with 0.1% Tween-20 in
Tris-buffered saline (TBTS) prior to and after the membranes were
incubated in 5% milk with IgG horseradish peroxidase-conjugated
secondary antibodies (1:1,000; cat. nos. 7074 and 7076; Cell
Signaling Technology, Inc.) for 1 h with shaking at room
temperature. Protein bands were detected on the C-DiGit®
Blot Scanner (LI-COR Biosciences) using SignalFire™ ECL reagent
(Cell Signaling Technology, Inc.). Band intensities were quantified
using ImageJ analysis software version 1.46r (National Institutes
of Health) and normalized to loading control (β-actin) band
intensity in each lane.
Immunocytochemistry
MDA-MB-231 and MCF-7 cells (3×105) were
grown on coverslips in 6-well plates and incubated for 48 h in a
humidified atmosphere containing 5% CO2 at 37°C. After
cryoinjury, cells were fixed with 4% paraformaldehyde overnight at
4°C. Then, cells were washed with TBST 3 times, followed by
permeabilization with 0.5% Triton-X-100 in TBST for 30 min at room
temperature. Cells were then blocked with 1.5% goat serum (cat. no.
ab7481; Abcam) in 1% BSA for 1 h at room temperature. Subsequently,
cells were incubated overnight at 4°C with rabbit monoclonal
antibodies against AQP1 (1:200; cat. no. ab168387; Abcam) and AQP5
(1:200; cat. no. cab92320; Abcam), and a rabbit polyclonal against
AQP3 (cat. no. ab125219; 1:200; Abcam). Then, cells were washed
with TBST 3 times prior to and after incubation in the dark with a
goat polyclonal Alexa Fluor® 488 secondary antibody
(1:200; cat. no. ab150077; Abcam) in 1% BSA for 1 h at room
temperature. Slides were then mounted using Fluoroshield™ with DAPI
(Sigma-Aldrich; Merck KGaA) and kept in the dark at 4°C before
visualization at ×40 magnification using a ZEISS LSM 710 confocal
microscope (Zeiss AG).
Transfection of cells with small
interfering (si)RNA
siRNAs against AQP1 (cat. no. sc-29711), AQP3 (cat.
no. sc-29713) and AQP5 (cat. no. sc-29717; all from Santa Cruz
Biotechnology, Inc.) consisting of 3 target-specific 19–25
nucleotides were used to knock down gene expression for 6 h in a
humidified 5% CO2 atmosphere at 37°C. Control siRNA
(cat. no. sc-37007; Santa Cruz Biotechnology, Inc.) consisting of a
scrambled sequence was used as the negative control. MDA-MB-231 and
MCF-7 cells were transfected with Lipofectamine® 3000
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocols. The concentrations of siRNA, including
the control siRNA, used for each transfection were: RT-qPCR, 10
µg/ml; immunocytochemistry, 1.5 µg/ml; and cell viability assay,
0.12 µg/ml (Table II). After 6 h,
the media was aspirated and fresh Advanced DMEM was added. After 48
h, cells were cryoinjured for 10 min at −13°C and processed the
following day. The transfection efficiency was assessed by RT-qPCR
as described above.
| Table II.Volumes, cell densities and
conditions used for RT-qPCR, immunocytochemistry and the cell
viability assays. |
Table II.
Volumes, cell densities and
conditions used for RT-qPCR, immunocytochemistry and the cell
viability assays.
| Method | Plate | Cell
density/well | Solution A: siRNA
(µl) | Solution B:
Lipofectamine (µl) | Reduced serum media
(µl) |
|---|
| RT-qPCR | 6-well |
2.0×105 | 8.0+125.0 µl | 3.7+125.0 | 750 |
|
|
|
| Opti-MEM | Opti-MEM |
|
|
Immunocytochemistry | 24-well |
2.5×105 | 1.2+25.0 µl | 0.7+25.0 µl | 250 |
|
|
|
| Opti-MEM | Opti-MEM |
|
| Cell viability
assay | 96-well |
1×104 | 0.1+5.0 µl | 0.2+5.0 µl | 80 |
|
|
|
| Opti-MEM | Opti-MEM |
|
Cell viability assay
Cell viability was assessed using a Sulforhodamine B
(SRB) assay (Sigma-Aldrich; Merck KGaA) as described by Skehan
et al (55). Cells were
seeded (1×104) in 96-well plates in triplicate for 24 h
followed by transfection for 48 h. At 24 h post cryoinjury, cells
were fixed with 10% trichloroacetic acid (TCA) for 30 min at 4°C.
TCA was then aspirated and dried in a 37°C oven before the cells
were stained with 0.4% SRB dissolved in 1% acetic acid for 10 min
at room temperature. To remove the unbound dye, cells were washed
with 1% acetic acid and then dried in a 37°C oven. The bound dye
was then solubilized with 10 mM Tris base solution (pH 10.5) and
shaken before measuring the absorbance. Absorbance was measured at
a wavelength of 540 nm with a reference wavelength of 650 nm using
a microplate reader. Cell viability was calculated according to the
following formula: Cell viability=100-[(absorbance of treated
cells/absorbance of untreated cells) ×100].
Statistical analysis
Data are presented as the mean ± SD of at least 3
repeats. Means were compared using one-way ANOVA with the post hoc
Tukey's test used for pairwise comparisons. All statistical tests
were carried out using IBM SPSS Statistics v.25 (IBM Corp). Graphs
were drawn using Microsoft Excel v.2002 (Microsoft Corporation).
P<0.05 was considered to indicate a statistically significant
difference.
Results
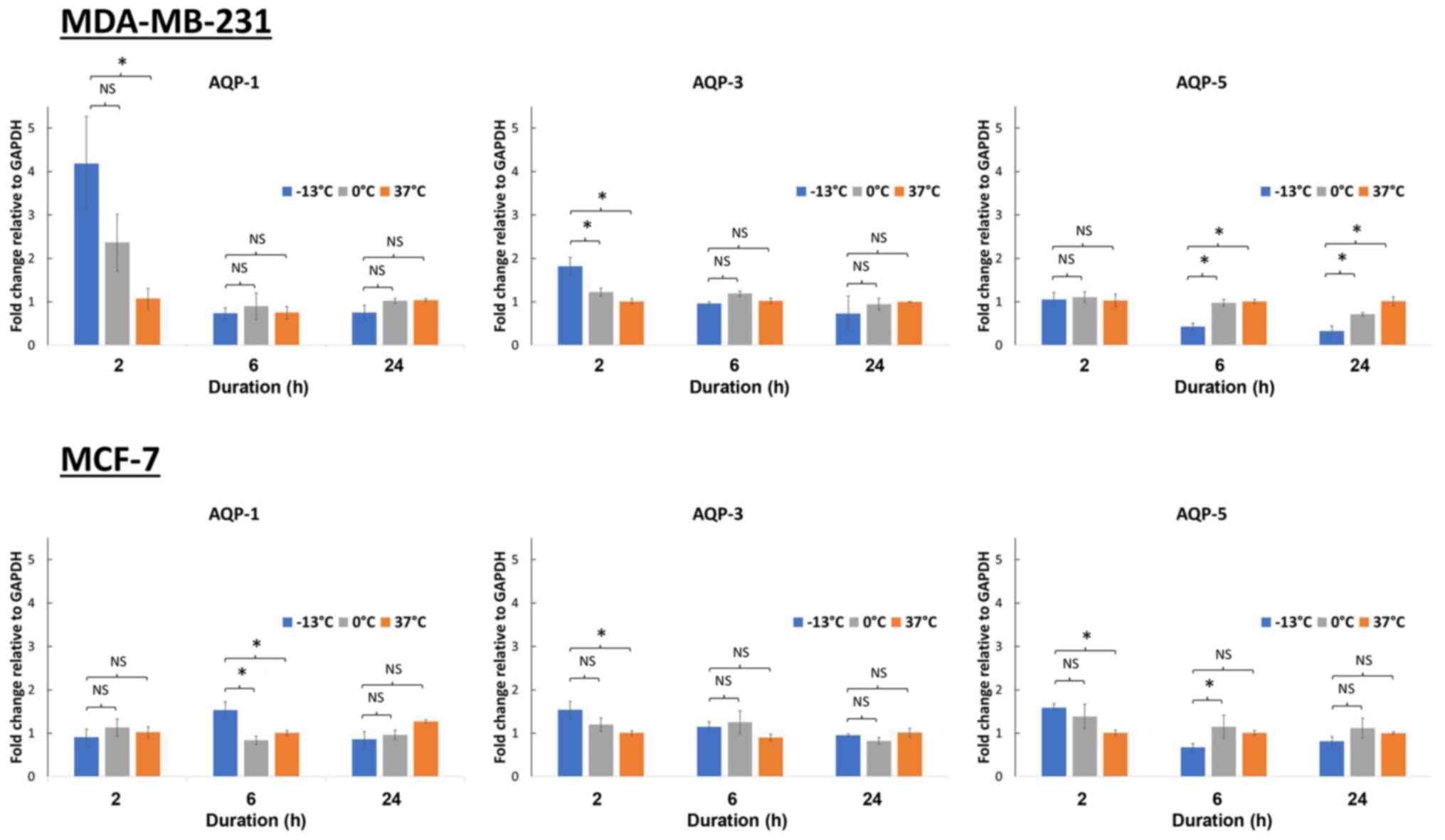
Changes in mRNA expression levels of
AQP1, AQP3 and AQP5 in breast cancer cells upon cryoinjury after 2,
6 and 24 h
Breast cancer cells (MCF-7 and MDA-MB-231) were
incubated at −13, 0 and 37°C for 10 min. A temperature of 0°C was
chosen as the control for non-freezing condition, while −13°C was
the lowest temperature capable of achieving freezing whilst
maintaining cell viability. Following this, AQP1, AQP3 and AQP5
mRNA expression levels were analyzed at 2, 6 and 24 h of
incubation. Several different time points were used as the rate of
gene expression varies depending on the gene. With the exception of
AQP1 expression in MDA-MD-231 cells (P<0.05), no significant
fold-change in AQP expression was observed in the MCF-7 and
MDA-MB-231 upon treatment at different temperatures or incubation
times (Fig. 1). A 4-fold increase in
the AQP1 gene expression was observed in MDA-MB-231 cells, 2 h
following freezing at −13°C (P<0.05), while a 2-fold increase
was seen at 0°C when compared with control cells kept at 37°C
(Fig. 1).
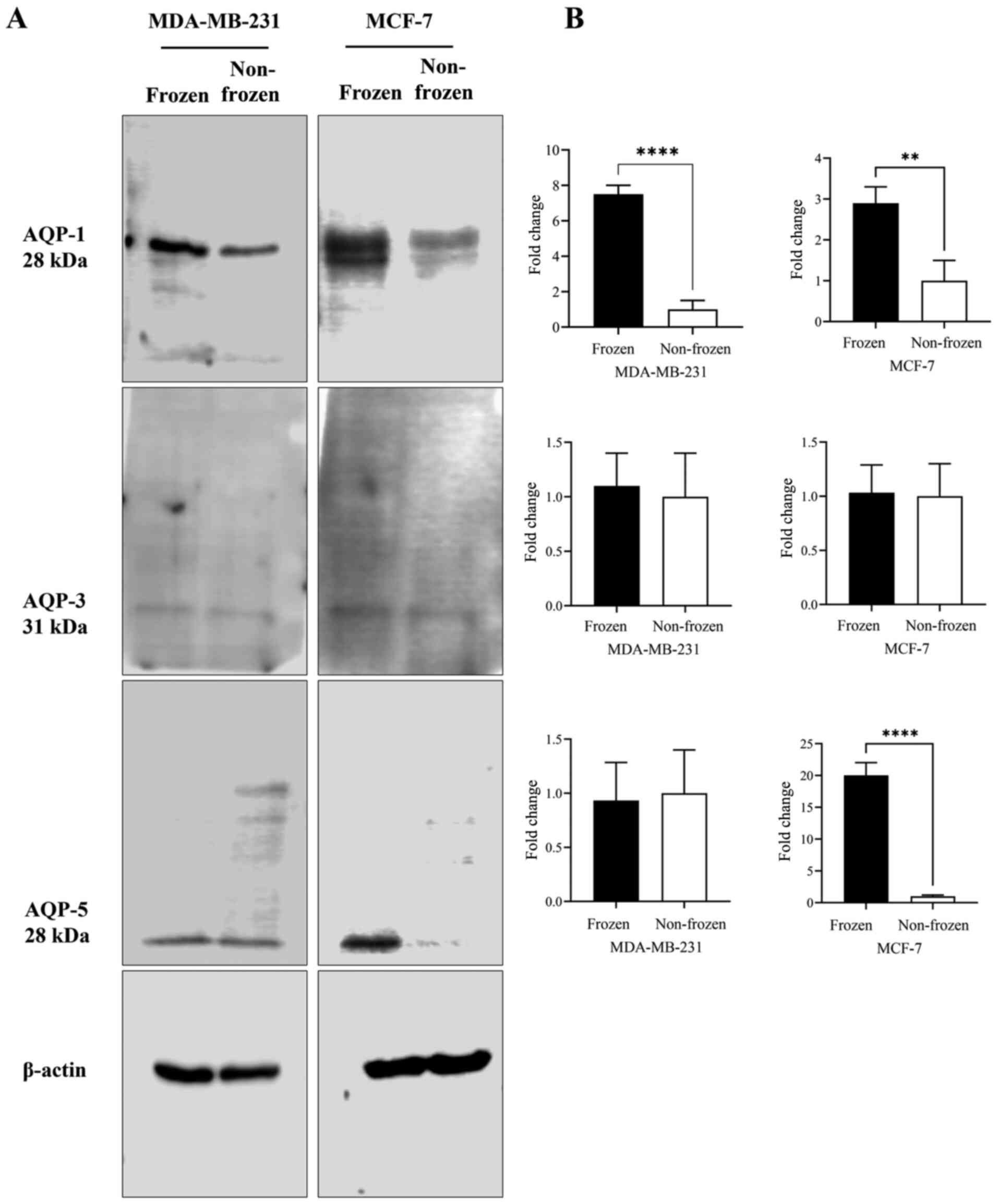
Changes in AQP1, AQP3 and AQP5 protein
expression levels in breast cancer cells upon cryoinjury as
determined via western blotting
AQP1, AQP3 and AQP5 protein expression levels in
MCF-7 and MDA-MB-231 cells were quantified following exposure to
frozen (−13°C) and non-frozen (37°C) temperatures (Fig. 2). No clear difference in AQP-3
protein content was observed for either cell line at the two
temperatures (Fig. 2A and B).
Following exposure to freezing temperature, AQP-1 protein content
was increased in both cell lines by 7- and 3-fold in MDA-MB-231
(P<0.001) and MCF-7 (P<0.05) cells, respectively, when
compared with non-frozen cells maintained at 37°C (Fig. 2B). AQP5 protein content was increased
by 20-fold (P<0.001) following exposure to freezing temperature
only in MCF-7 cells (Fig. 2B).
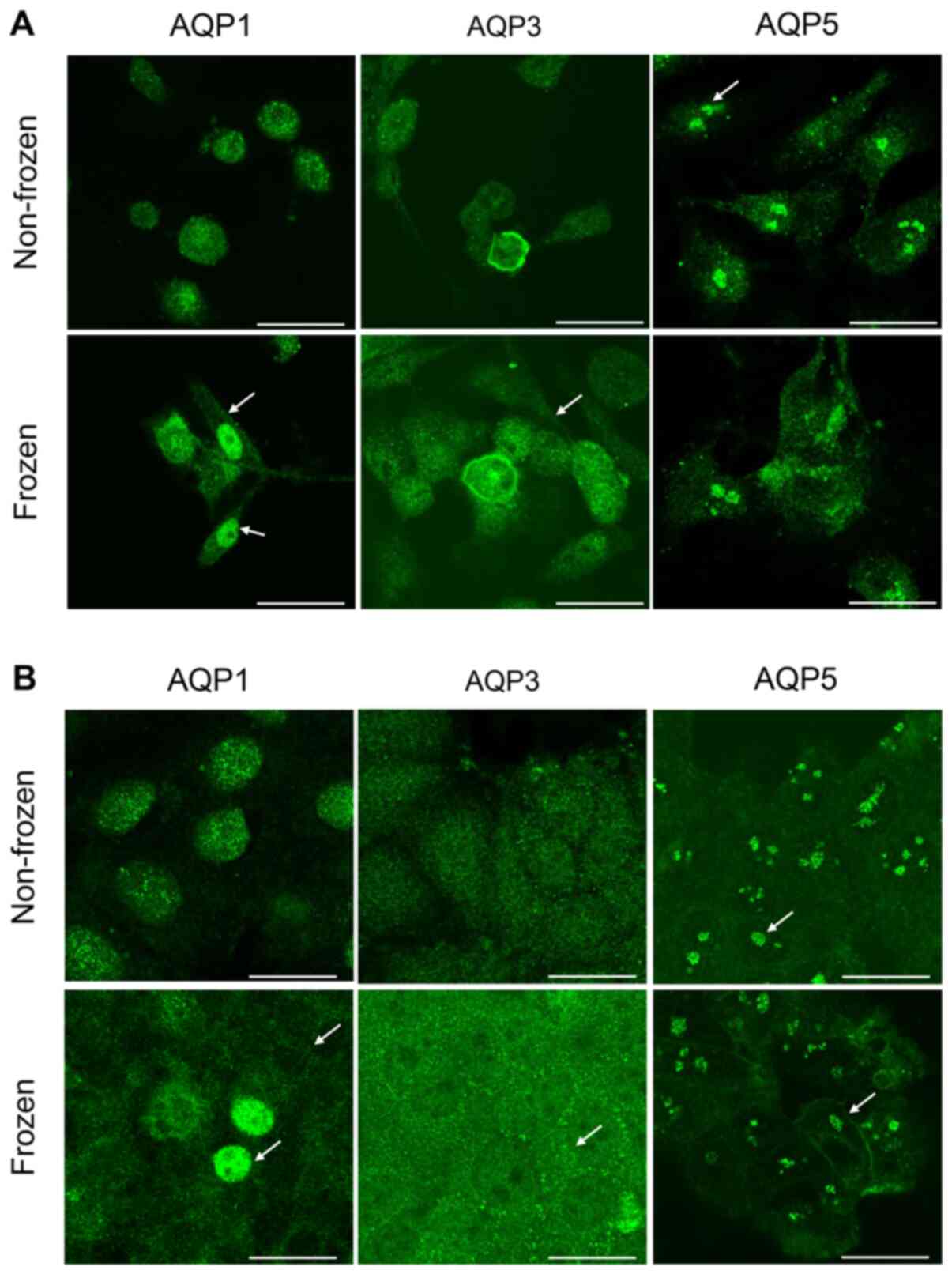
Localization of AQP1 AQP3 and AQP5 in
breast cancer cells
The intracellular localization of AQP proteins in
both cell lines was determined by immunocytochemistry. The exposure
to freezing temperature altered the localization of AQPs in
MDA-MB-231 cells (Fig. 3A), but not
in MCF-7 cells (Fig. 3B), when
compared with non-frozen cells maintained at 37°C. With regards to
MDA-MB-231 cells, a strong AQP1 staining was observed within the
cytosol in non-frozen cells and within the plasma and nuclear
membranes in frozen cells (Fig. 3A).
However the staining was more intense in the nuclear membrane when
compared with the plasma membrane in MDA-MB-231 cells exposed to
freezing. Similar staining for AQP3 expression was observed in the
plasma membrane following freezing of MDA-MB-231 cells (Fig. 3A). Whereas AQP5 protein expression
was clustered and localized in the nucleus in both treatment
conditions (Fig. 3A). No observable
difference in intensity was seen between the plasma and nuclear
membranes for AQP3 and AQP5 staining in MDA-MB-231 cells when
compared to non-frozen cells (Fig.
3A). In MCF-7 cells, staining for AQP1 demonstrated that the
freezing temperature affected the protein localization (Fig. 3B). AQP1 had a diffused cytoplasm
staining and accumulated in the plasma membrane when compared with
AQP1 localization in non-frozen cells (Fig. 3B). Few cells had an intense nuclear
staining as presented by the arrows (Fig. 3B). AQP3 proteins formed small
clusters localized at the plasma membrane in MCF-7 cells exposed to
the freezing temperature when compared to non-frozen cells
maintained at 37°C. (Fig. 3B). The
staining of AQP5 in MCF-7 cells revealed a similar clustering
pattern observed previously in MDA-MB-231 cells (Fig. 3A) and was not affected by the
freezing temperature (Fig. 3B).
However, the plasma membrane localization was more intense
following exposure to freezing temperature as presented by the
arrows (Fig. 3A and B).
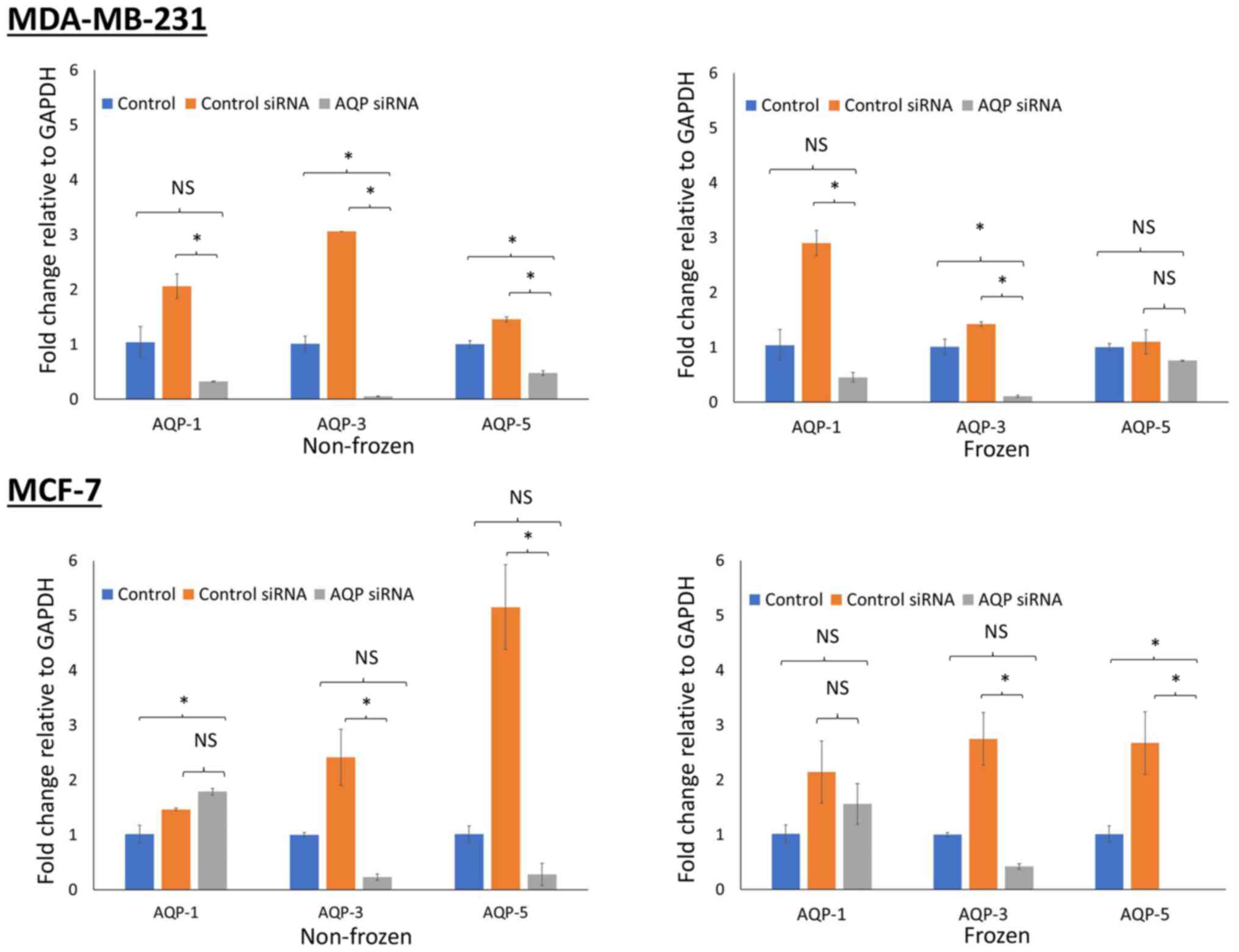
Assessment of gene knockdown following
siRNA transfection
AQP gene silencing was assessed via RT-qPCR. A
successful knockdown was achieved after 48 h of transfection, with
an efficiency of 90%. The expression of AQP genes was normalized to
GAPDH and compared with control cells maintained at 37°C.
MDA-MB-231 cells demonstrated a reduction in gene expression for
all 3 types of aquaporin siRNA transfected in both non-frozen cells
(P<0.05) maintained at 37°C and frozen cells (P<0.05 for AQP1
and AQP3) exposed to −13°C (Fig. 4).
Similar reductions in gene expression were observed in MCF-7 cells
(P<0.05 for AQP3 and AQP5; both frozen and non-frozen
exposures). However, this was not the case for AQP1 whose
expression was increased following transfection of AQP1 siRNA
(Fig. 4).
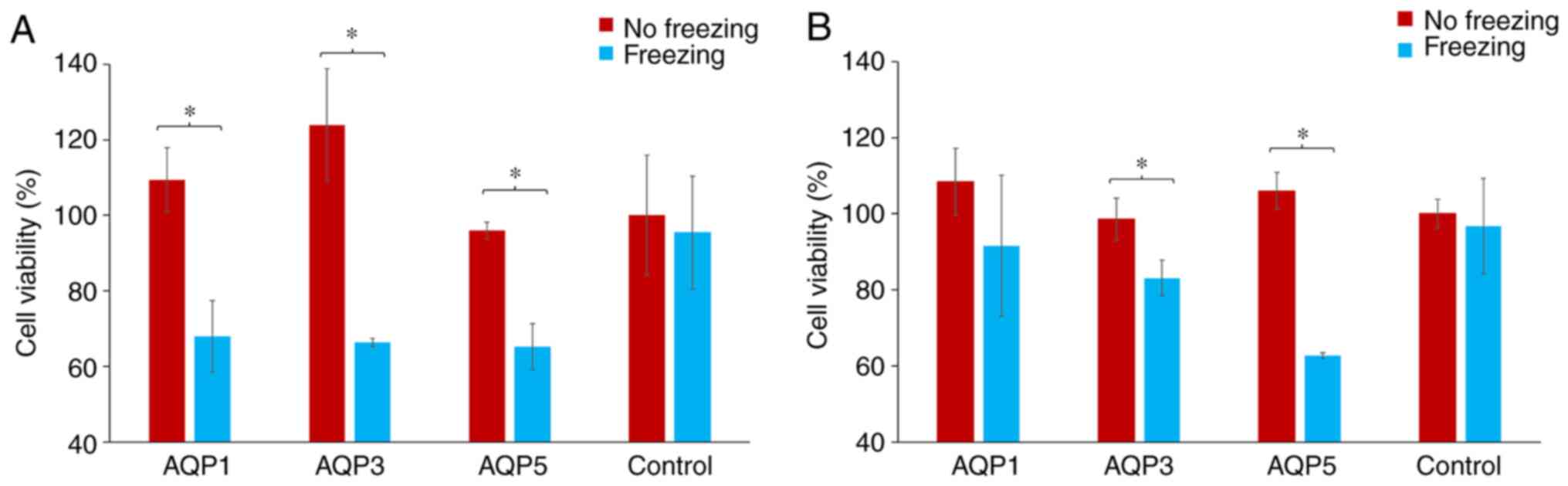
Assessment of cell viability following
siRNA transfection and exposure to freezing in breast cancer
cells
Following transfection with AQP-specific or control
siRNA, cells were either exposed to freezing temperature or not.
Cell viability was then assessed using the SRB assay and
comparisons were made between non-frozen and frozen cells. The
percentage change in cell viability for each AQP-specific siRNA
group was calculated by normalizing the cell viability in the
transfected cells to the cell viability in the cells transfected
with scramble control siRNA. These were then represented as
percentage differences. The exposure of both MCF-7 and MDA-MB-231
cells to freezing temperature did not affect their viability when
transfected with the scrambled siRNA.
With regards to MDA-MD-231 cells, there was a
significant decrease in cell viability for frozen cells transfected
with AQP1, AQP3 and AQP5 siRNA compared with the non-frozen cells
(P<0.05; Fig. 5A). This reduction
in cell viability was 32.6, 46.4 and 32.0% for AQP1, AQP3 and AQP5
siRNA transfected cells, respectively (Fig. 5A). There was a similarly significant
reduction in the cell viability of MCF-7 cells that were frozen
compared with non-frozen cells, for cells transfected with siRNA
targeting AQP3 and AQP5 only (P<0.05; Fig. 5B). There was no statistical
difference between MCF-7 cells transfected with siRNA against AQP1,
which was the one transfection that failed to silence the test-gene
under investigation (Fig. 5B). The
reductions observed in cell viability were 15.5, 15.7 and 40.8% for
AQP1, AQP3 and AQP5 siRNA transfected cells, respectively (Fig. 5B).
Discussion
In the present study, cryoinjury did not have a
major influence on gene expression of AQPs in breast cancer cells.
Based upon RT-qPCR results, the freezing of cells induced an
increase in the gene expression of AQP1 only in the MDA-MD-231 cell
line. In contrast in the present study, freezing did not appear to
have a major effect on the expression of AQP3 and AQP5 in
MDA-MB-231 cells or on AQP-1, 3 and 5 in MCF-7 cells. In addition
to mRNA-based gene expression, AQP protein levels were also
assessed as part of the present study. Bands of immunoreactivity
were present on western blots, which indicated that freezing cells
had no major influence on the expression of AQP3 protein. However,
the AQP1 protein expression was sensitive to the variation of
temperature in both cell lines as observed by the increase band
intensity following the exposure to freezing temperature when
compared with control cells maintained at 37°C. The expression of
AQP5 was sensitive to the exposure of freezing temperature only in
MCF-7, but not in MDA-MB-231 cells. This finding of the present
study demonstrated the heterogeneity of breast cancers. The use of
an additional quantitative assay to investigate the expression of
this protein may further elucidate the effect of freezing on
protein expression in these cell lines. In the present study, the
measure of the presence or absence of AQP protein does not give an
indication of the cellular location of the detected AQP protein
immunoreactivity. Hence, these results should be evaluated in
tandem with the immunocytochemistry images. Immunofluorescence
staining in the present study demonstrated that freezing cells had
the effect of translocating aquaporins from the cytosol to the
plasma membrane and incorporating these protein channels in the
cell membranes. This redistribution observed in the present study,
may reflect the involvement of aquaporin in the movement of
intercellular water associated with freezing. Results from the cell
lines in the present study suggested that freezing did not only
influence the amount of AQP protein expression in the cell, but
also influenced the cellular localization of these proteins.
Cryoablation is considered an efficacious breast
cancer treatment and this efficacy is due to the destructive
effects of cold temperatures on the cellular ultrastructure of
tumor tissue (5). The destruction of
neoplastic tissue is achieved though freeze/thaw cycles using a
cryoprobe (12). The goal of the
present study was to assess cryo-damage in association with
aquaporin downregulation using breast cancer cells in vitro.
Clinically, the first freeze cycle in cryoablation has a duration
of 5–13 min (56,57). Hence, a freezing time of 10 min in
the in vitro setting in the present study was considered
appropriate. The time taken for the temperature to decrease from
37°C to below 0°C was 5 min and the actual freezing time of the
cells at −13°C was 5 min in the present study. In the present
study, 10 min was the time needed for the media to freeze whilst
still preserving the integrity of the cells. The present study
replicated elements of the in vivo clinical application of
cryoinjury. However, this study could not mimic or simulate the
in vivo environment. The freezing protocols employed
clinically use temperatures below −40°C and consist of freeze/thaw
cycles as freezing alone once does not uniformly damage the
targeted tumor (12). Multiple
freezing cycles extend the zone of destruction ensuring maximum
lethality (17). However, on the
margins of the targeted area, the cells are exposed to subzero
temperatures where survival of the cancer cells is possible.
Cells that express AQPs acquire a tolerance to
freezing by evading cell membrane damage (33). It was previously reported that AQP3
gene silencing increased cryosensitization in prostate cancer cells
and that freezing caused the relocation of AQP3 from the cytosol to
the membrane (22). To the best of
our knowledge, the present study is the first to report similar
cellular responses to freezing in breast cancer cell lines. Fujita
et al (58) previously found
that enhanced expression of AQP3 in cancer cells was an attempt to
overcome osmotic stress. An upregulation on exposure to mild
hypothermia was reported in earlier studies (58,59). It
has also been demonstrated that cryoinjury leads to increased
expression of AQP3 in prostate cancer cells (22). However, an increase in AQP3
expression was not observed in the 2 breast cancer cell lines used
in the present study, as determined using RT-qPCR and
immunocytochemistry. A cellular redistribution of AQPs was observed
following freezing at −13°C with MDA-MB-231 cells, but not with
MCF-7 cells. This difference in localization between the 2 cell
lines may be related to differences in growth patterns between the
two. Temperatures lower than −13°C may have brought about a similar
effect with MCF-7 as was observed with MDA-MB-231 cells. However,
the low temperatures of −30°C or −40°C that are typically used
clinically would likely destroy cells and alter the fluidity of the
proteins within them (17).
Temperatures of ~-40 C cause both intra- and extra-cellular water
to solidify (17). Hence, a
temperature of −13°C was chosen in the present study to mimic the
behavior of cancer cells located near the zone targeted by the
ultra-cold temperatures administered during clinical cryotherapy
procedures. Clinically, temperatures located on the margins of the
cryo-ablated area, such as −13°C, will recover (12,22).
This damage to cells may be influenced by the expression of
aquaporins (12,22).
AQP expression is associated with numerous
pathologies, which includes tumor metastasis (34,60,61).
However, there are currently no definitive, small molecule AQP
inhibitors available for therapeutic use (62). Heavy metal compounds, such as
mercury, are effective AQP inhibitors (22), but in the present study they were
considered inappropriate for use in a biological experiment due to
their non-specificity and toxicity. Hence, the present study
employed a gene-silencing technique to investigate the role of AQPs
in cryosensitization. In the present study, the transfection of
breast cancer cells with siRNA had an influence on cell viability
following cryodamage. The viability of frozen MDA-MD-231 cells was
decreased in the present study following transfection with AQP1,
AQP3 and AQP5 siRNA when compared with MDA-MB-231 cells transfected
with siRNA-NC. A similar reduction in cell viability was observed
in the present study with MCF-7 cells transfected with AQP3 siRNA,
which was more pronounced with AQP5 siRNA. In the present study,
the transfection of AQP1 siRNA in MCF-7 cells led to an increase in
AQP1 mRNA expression probably due to the siRNA failing to target
AQP1 mRNA efficiently in MCF-7 cells. The aforementioned findings
of the present study suggested that the silencing of AQP genes
exacerbated cell damage associated with freezing. Notably the
expression levels of some aquaporins were significantly higher in
the control siRNA groups as compared to those in the untransfected
control groups. The process of transfection may have had an impact
on the aquaporin expression which in turn resulted in the increase
expression of aquaporin in the present study as transfection
effects the permeability of cells (63).
It has been demonstrated that AQP1 is upregulated in
breast cancer cells, which is associated with poor patient
prognosis resulting from the induction of angiogenesis which leads
to metastasis (64). It is
speculated that AQP1 upregulation is stimulated by estrogen acting
via the ER (34). MDA-MB-231 cells
are lacking in these receptors (65). However, freezing these ER-deficient
cells in the present study increased AQP1 expression. This implied
that the underlying mechanism of induction was independent of ER.
In addition, an increase in the expression of AQP1 was not observed
in the MCF-7 cells in the present study, which do possess an ER
(65).
Previous studies have demonstrated AQP1 to be
upregulated in breast cancer cells and this has been associated
with enhanced cell proliferation and invasion, which may make AQP1
a potential prognostic marker for breast cancer (38,66). The
increase of AQP1 in MDA-MB-231 following freezing that was observed
in the present study may promote the cell's tolerance to
cryo-damage as the knockdown of AQP1 expression decreased
MDA-MB-231 cell viability. It is therefore reasonable to assume
that increase expression of AQP1 allows the cells to recover from
cryoinjury. The techniques employed in the current study measured
quantitative fold-changes in RNA and the presence or absence of
aquaporin proteins. However, no measurement of aquaporin channel
function activity was assessed. A functional assay of channel
activity would add value to this investigation as cryoinjury and
sensitization to injury are active processes (17). These processes depend upon the
activity of the channels studied rather than the mere absences,
presences or location of the protein (34,61,64). It
has been demonstrated that AQP1 activity is regulated by cyclic
nucleotides, such as cAMP and protein kinase pathways, such as
focal adhesion kinase (64).
Post-translational modifications, such as phosphorylation regulate
AQP1 activity (67). The methods
adopted in the current study did not consider such necessary
modifications to aquaporin proteins.
Clinically, multiple freeze-thaw cycles are crucial
to the therapeutic application cry-injury (17). However, the laboratory experimental
setting employed in the present study did not allow for multiple
freezing. This is a limitation of the laboratory approach to
investigating cryoinjury using cell culture. The limitations of the
present study also included the use of a cell monolayer instead of
tissue. In addition, the in vitro system did not incorporate
estrogen exposure as part of the experimental design. Future
studies should examine the relationship between estrogen and AQP1
in this ER-expressing cell line in the context of cryodamage.
In summary, in the present study, freezing breast
cancer cells induced the redistribution of AQP proteins from the
cytosol to the cell membrane. Inhibition of AQP function
exacerbated cell damage associated with freezing. This indicated
that reduced aquaporin function may be used as an adjunct to
cryotherapy. The combination of cryotherapy and AQP inhibition may
result in less aggressive freezing protocols whilst achieving more
complete ablation of tumors and, ultimately, lower treatment
failure for patients with breast cancer.
Acknowledgements
The authors would like to thank Mrs. Asma Mostafa
(Sharjah Institute of Research and Technology, University of
Sharjah) and Dr Nabil El Zein (Faculty of Science, Lebanese
University) for their technical help, advice and assistance
regarding the breast cancer cell lines.
Funding
This study was supported by a RCSI (Bahrain)
internal research grant (no. BR00070).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HA performed the cell and molecular laboratory
experiments, data analysis and wrote the manuscript. FM was
involved data conception and analysis. SeT conceived the
experimental design, conducted data analysis and revised the
manuscript for important intellectual content. KG performed the
data analysis of the laboratory work. SaT performed and supervised
laboratory work. SF was involved in data collection and analysis
and drafted the initial manuscript. HA, SeT, FM, and SF confirmed
the authenticity of all the raw data. All the authors have read and
approved the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Global Burden of Disease Cancer
Collaboration, ; Fitzmaurice C, Allen C, Barber RM, Barregard L,
Bhutta ZA, Brenner H, Dicker DJ, Chimed-Orchir O, Dandona R, et al:
Global, regional, and national cancer incidence, mortality, years
of life lost, years lived with disability, and disability-adjusted
life-years for 32 cancer groups, 1990 to 2015: A systematic
analysis for the global burden of disease study. JAMA Oncol.
3:524–548. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
PDQ Adult Treatment Editorial Board, .
Breast cancer treatment (Adult) (PDQ®): Patient version.
PDQ Cancer Information Summaries. National Cancer Institute (USA);
Bethesda, MD: 2002
|
|
4
|
Sharma GN, Dave R, Sanadya J, Sharma P and
Sharma KK: Various types and management of breast cancer: An
overview. J Adv Pharm Technol Res. 1:109–126. 2010.PubMed/NCBI
|
|
5
|
Pusceddu C, Melis L, Ballicu N, Meloni P,
Sanna V, Porcu A and Fancellu A: Cryoablation of primary breast
cancer in patients with metastatic disease: Considerations arising
from a single-centre data analysis. Biomed Res Int.
2017:38390122017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Erinjeri JP and Clark TW: Cryoablation:
Mechanism of action and devices. J Vasc Interv Radiol. 21 (Suppl
8):S187–S191. 2010. View Article : Google Scholar
|
|
7
|
Pusceddu C, Paliogiannis P, Nigri G and
Fancellu A: Cryoablation in the management of breast cancer:
Evidence to date. Breast Cancer (Dove Med Press). 11:283–292.
2019.PubMed/NCBI
|
|
8
|
Beji H, Pilleul F, Picard R, Tredan O,
Bouhamama A, Peix M, Mavrovi E and Mastier C: Percutaneous
cryoablation of breast tumours in patients with stable metastatic
breast cancer: Safety, feasibility and efficacy. Br J Radiol.
91:201705002018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Holmes DR: Breast cancer care during a
pandemic: An opportune time for cryoablation? Breast Cancer Res
Treat. 182:515–521. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mahnken AH, König AM and Figiel JH:
Current technique and application of percutaneous cryotherapy.
Rofo. 190:836–846. 2018.(In English, German). View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Niu L, Mu F, Zhang C, Li Y, Liu W, Jiang
F, Li L, Liu C, Zeng J, Yao F, et al: Cryotherapy protocols for
metastatic breast cancer after failure of radical surgery.
Cryobiology. 67:17–22. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Baust JG, Gage AA, Bjerklund Johansen TE
and Baust JM: Mechanisms of cryoablation: Clinical consequences on
malignant tumors. Cryobiology. 68:1–11. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Aarts BM, Klompenhouwer EG, Rice SL, Imani
F, Baetens T, Bex A, Horenblas S, Kok M, Haanen JBAG, Beets-Tan RGH
and Gómez FM: Cryoablation and immunotherapy: An overview of
evidence on its synergy. Insights Imaging. 10:532019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gage AA and Baust J: Mechanisms of tissue
injury in cryosurgery. Cryobiology. 37:171–186. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hoffmann NE and Bischof JC: The
cryobiology of cryosurgical injury. Urology. 60 (Suppl 1):S40–S49.
2002. View Article : Google Scholar
|
|
16
|
Santucci KL, Snyder KK, Baust JM, Van
Buskirk RG, Mouraviev V, Polascik TJ, Gage AA and Baust JG: Use of
1,25α dihydroxyvitamin D3 as a cryosensitizing agent in a murine
prostate cancer model. Prostate Cancer Prostatic Dis. 14:97–104.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Baust JG, Snyder KK, Santucci KL,
Robilotto AT, Van Buskirk RG and Baust JM: Cryoablation: Physical
and molecular basis with putative immunological consequences. Int J
Hyperthermia. 36 (Suppl 1):S10–S16. 2019. View Article : Google Scholar
|
|
18
|
Clarke DM, Baust JM, Van Buskirk RG and
Baust JG: Chemo-cryo combination therapy: An adjunctive model for
the treatment of prostate cancer. Cryobiology. 42:274–285. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Clarke DM, Baust JM, Van Buskirk RG and
Baust JG: Addition of anticancer agents enhances freezing-induced
prostate cancer cell death: Implications of mitochondrial
involvement. Cryobiology. 49:45–61. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Le Pivert P, Haddad RS, Aller A, Titus K,
Doulat J, Renard M and Morrison DR: Ultrasound guided combined
cryoablation and microencapsulated 5-Fluorouracil inhibits growth
of human prostate tumors in xenogenic mouse model assessed by
luminescence imaging. Technol Cancer Res Treat. 3:135–142. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Baust JM, Klossner DP, Robilotto A,
Vanbuskirk RG, Gage AA, Mouraviev V, Polascik TJ and Baust JG:
Vitamin D(3) cryosensitization increases prostate cancer
susceptibility to cryoablation via mitochondrial-mediated apoptosis
and necrosis. BJU Int. 109:949–958. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ismail M, Bokaee S, Morgan R, Davies J,
Harrington KJ and Pandha H: Inhibition of the aquaporin 3 water
channel increases the sensitivity of prostate cancer cells to
cryotherapy. Br J Cancer. 100:1889–1895. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Khajah MA and Luqmani YA: Role of
aquaporins in breast cancer progression and metastasis. Tumor Met.
59–83. 2016.
|
|
24
|
Wang L, Zhang Y, Wu X and Yu G:
Aquaporins: New targets for cancer therapy. Technol Cancer Res
Treat. 15:821–828. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hagedorn M, Lance SL, Fonseca DM,
Kleinhans FW, Artimov D, Fleischer R, Hoque AT, Hamilton MB and
Pukazhenthi BS: Altering fish embryos with aquaporin-3: An
essential step toward successful cryopreservation. Biol Reprod.
67:961–966. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Edashige K, Yamaji Y, Kleinhans FW and
Kasai M: Artificial expression of aquaporin-3 improves the survival
of mouse oocytes after cryopreservation. Biol Reprod. 68:87–94.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tanghe A, Van Dijck P, Colavizza D and
Thevelein JM: Aquaporin-mediated improvement of freeze tolerance of
Saccharomyces cerevisiae is restricted to rapid freezing
conditions. Appl Environ Microbiol. 70:3377–3382. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tanghe A, Van Dijck P and Thevelein JM:
Why do microorganisms have aquaporins? Trends Microbiol. 14:78–85.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tietz PS, McNiven MA, Splinter PL, Huang
BQ and Larusso NF: Cytoskeletal and motor proteins facilitate
trafficking of AQP1-containing vesicles in cholangiocytes. Biol
Cell. 98:43–52. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Riethmüller C, Oberleithner H, Wilhelmi M,
Franz J, Schlatter E, Klokkers J and Edemir B: Translocation of
aquaporin-containing vesicles to the plasma membrane is facilitated
by actomyosin relaxation. Biophys J. 94:671–678. 2008. View Article : Google Scholar
|
|
31
|
Conner AC, Bill RM and Conner MT: An
emerging consensus on aquaporin translocation as a regulatory
mechanism. Mol Membr Biol. 30:1–12. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cohen E: Roles of aquaporins in
osmoregulation, desiccation and cold hardiness in insects. Entomol
Ornithol Herpetol. S1:1–17. 2012.
|
|
33
|
Kato Y, Miyauchi T, Abe Y, Kojić D, Tanaka
M, Chikazawa N, Nakatake Y, Ko SB, Kobayashi D, Hazama A, et al:
Unprecedented cell-selection using ultra-quick freezing combined
with aquaporin expression. PLoS One. 9:e876442014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang J, Feng L, Zhu Z, Zheng M, Wang D,
Chen Z and Sun H: Aquaporins as diagnostic and therapeutic targets
in cancer: How far we are? J Transl Med. 13:962015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Satooka H and Hara-Chikuma M: Aquaporin-3
controls breast cancer cell migration by regulating hydrogen
peroxide transport and its downstream cell signaling. Mol Cell
Biol. 36:1206–1218. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhu Z, Jiao L, Li T, Wang H, Wei W and
Qian H: Expression of AQP3 and AQP5 as a prognostic marker in
triple-negative breast cancer. Oncol Lett. 16:2661–2667.
2018.PubMed/NCBI
|
|
37
|
Otterbach F, Callies R, Adamzik M, Kimmig
R, Siffert W, Schmid KW and Bankfalvi A: Aquaporin 1 (AQP1)
expression is a novel characteristic feature of a particularly
aggressive subgroup of basal-like breast carcinomas. Breast Cancer
Res Treat. 120:67–76. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Qin F, Zhang H, Shao Y, Liu X, Yang L,
Huang Y, Fu L, Gu F and Ma Y: Expression of aquaporin1, a water
channel protein, in cytoplasm is negatively correlated with
prognosis of breast cancer patients. Oncotarget. 7:8143–8154. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Nielsen TO, Hsu FD, Jensen K, Cheang M,
Karaca G, Hu Z, Hernandez-Boussard T, Livasy C, Cowan D, Dressler
L, et al: Immunohistochemical and clinical characterization of the
basal-like subtype of invasive breast carcinoma. Clin Cancer Res.
10:5367–5374. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hugh J, Hanson J, Cheang MC, Nielsen TO,
Perou CM, Dumontet C, Reed J, Krajewska M, Treilleux I, Rupin M, et
al: Breast cancer subtypes and response to docetaxel in
node-positive breast cancer: Use of an immunohistochemical
definition in the BCIRG 001 trial. J Clin Oncol. 27:1168–1176.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Blows FM, Driver KE, Schmidt MK, Broeks A,
van Leeuwen FE, Wesseling J, Cheang MC, Gelmon K, Nielsen TO,
Blomqvist C, et al: Subtyping of breast cancer by
immunohistochemistry to investigate a relationship between subtype
and short and long term survival: A collaborative analysis of data
for 10,159 cases from 12 studies. PLoS Med. 7:e10002792010.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Carey LA, Dees EC, Sawyer L, Gatti L,
Moore DT, Collichio F, Ollila DW, Sartor CI, Graham ML and Perou
CM: The triple negative paradox: Primary tumor chemosensitivity of
breast cancer subtypes. Clin Cancer Res. 13:2329–2334. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Dent R, Trudeau M, Pritchard KI, Hanna WM,
Kahn HK, Sawka CA, Lickley LA, Rawlinson E, Sun P and Narod SA:
Triple-negative breast cancer: Clinical features and patterns of
recurrence. Clin Cancer Res. 13:4429–4434. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Liedtke C, Mazouni C, Hess KR, André F,
Tordai A, Mejia JA, Symmans WF, Gonzalez-Angulo AM, Hennessy B,
Green M, et al: Response to neoadjuvant therapy and long-term
survival in patients with triple-negative breast cancer. J Clin
Oncol. 26:1275–1281. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Li X, Yang J, Peng L, Sahin AA, Huo L,
Ward KC, O'Regan R, Torres MA and Meisel JL: Triple-negative breast
cancer has worse overall survival and cause-specific survival than
non-triple-negative breast cancer. Breast Cancer Res Treat.
161:279–287. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Cailleau R, Olive M and Cruciger QV:
Long-term human breast carcinoma cell lines of metastatic origin:
Preliminary characterization. In vitro. 14:911–915. 1978.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Chavez KJ, Garimella SV and Lipkowitz S:
Triple negative breast cancer cell lines: One tool in the search
for better treatment of triple negative breast cancer. Breast Dis.
32:35–48. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Sweeney EE, McDaniel RE, Maximov PY, Fan P
and Jordan VC: Models and mechanisms of acquired antihormone
resistance in breast cancer: Significant clinical progress despite
limitations. Horm Mol Biol Clin Investig. 9:143–163.
2012.PubMed/NCBI
|
|
49
|
Jung HJ, Park JY, Jeon HS and Kwon TH:
Aquaporin-5: A marker protein for proliferation and migration of
human breast cancer cells. PLoS One. 6:e284922011. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Ahmad AE, Khajah MA, Khushaish S and
Luqmani YA: Aquaporin expression in breast cancer and their
involvement in bleb formation, cell motility and invasion in
endocrine resistant variant cells. Int J Oncol. 56:1014–1024.
2020.PubMed/NCBI
|
|
51
|
Wei M, Yu H, Cai C, Gao R, Liu X and Zhu
H: MiR-3194-3p inhibits breast cancer progression by targeting
Aquaporin1. Front Oncol. 10:15132020. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Zhu F, Chen L, Zhou Y, An J, Wu Z, Hu J,
Ma Z and Cao F: Cellular-mesenchymal to epithelial transition
factor upregulates aquaporin 3 expression in human breast cancer
cells. Res Sq. 2020.
|
|
53
|
Snyder KK, Van Buskirk RG, Baust JG and
Baust JM: Breast cancer cryoablation: Assessment of the impact of
fundamental procedural variables in an in vitro human breast cancer
model. Breast Cancer (Auckl). 14:11782234209723632020.PubMed/NCBI
|
|
54
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Skehan P, Storeng R, Scudiero D, Monks A,
McMahon J, Vistica D, Warren JT, Bokesch H, Kenney S and Boyd MR:
New colorimetric cytotoxicity assay for anticancer-drug screening.
J Natl Cancer Inst. 82:1107–1112. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Littrup PJ, Jallad B, Chandiwala-Mody P,
D'Agostini M, Adam BA and Bouwman D: Cryotherapy for breast cancer:
A feasibility study without excision. J Vasc Interv Radiol.
20:1329–1341. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Cazzato RL, de Lara CT, Buy X, Ferron S,
Hurtevent G, Fournier M, Debled M and Palussière J: Single-centre
experience with percutaneous cryoablation of breast cancer in 23
consecutive non-surgical patients. Cardiovasc Intervent Radiol.
38:1237–1243. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Fujita Y, Yamamoto N, Sobue K, Inagaki M,
Ito H, Arima H, Morishima T, Takeuchi A, Tsuda T, Katsuya H and
Asai K: Effect of mild hypothermia on the expression of aquaporin
family in cultured rat astrocytes under hypoxic condition. Neurosci
Res. 47:437–444. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Salman MM, Kitchen P, Woodroofe MN, Brown
JE, Bill RM, Conner AC and Conner MT: Hypothermia increases
aquaporin 4 (AQP4) plasma membrane abundance in human primary
cortical astrocytes via a calcium/transient receptor potential
vanilloid 4 (TRPV4)- and calmodulin-mediated mechanism. Eur J
Neurosci. 46:2542–2547. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Papadopoulos MC and Saadoun S: Key roles
of aquaporins in tumor biology. Biochim Biophys Acta.
1848:2576–2583. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Dajani S, Saripalli A and Sharma-Walia N:
Water transport proteins-aquaporins (AQPs) in cancer biology.
Oncotarget. 9:36392–36405. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Abir-Awan M, Kitchen P, Salman MM, Conner
MT, Conner AC and Bill RM: Inhibitors of mammalian aquaporin water
channels. Int J Mol Sci. 20:15892019. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Prevette LE, Mullen DG and Holl MM:
Polycation-induced cell membrane permeability does not enhance
cellular uptake or expression efficiency of delivered DNA. Mol
Pharm. 7:870–883. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Tomita Y, Dorward H, Yool AJ, Smith E,
Townsend AR, Price TJ and Hardingham JE: Role of Aquaporin 1
signalling in cancer development and progression. Int J Mol Sci.
18:2992017. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Taurin S, Allen KM, Scandlyn MJ and
Rosengren RJ: Raloxifene reduces triple-negative breast cancer
tumor growth and decreases EGFR expression. Int J Oncol.
43:785–792. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Chong W, Zhang H, Guo Z, Yang L, Shao Y,
Liu X, Zhao Y, Wang Z, Zhang M, Guo C, et al: Aquaporin 1 promotes
sensitivity of anthracycline chemotherapy in breast cancer by
inhibiting β-catenin degradation to enhance TopoIIα activity. Cell
Death Differ. 28:382–400. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Han Z and Patil RV: Protein Kinase
A-dependent phosphorylation of aquaporin-1. Biochem Biophys Res
Commun. 273:328–332. 2000. View Article : Google Scholar : PubMed/NCBI
|